
Abstract
Aiming to evaluate responses in terms of growth rates, physiological parameters, and degree of sensitivity to SO2 and SPMFe in Eugenia uniflora L. (Myrtaceae, a C3 species) and Clusia hilariana Schlecht (Clusiaceae, a CAM species); saplings were exposed to emissions from a pelletizing factory for 7 months. The species were distributed along a transect (200, 500, 800, 1400, and 1700 m away from the emission source), and analyses were performed after 71, 118, and 211 days of exposure to the pollutants. E. uniflora received higher superficial deposition of particulate iron. The highest total iron foliar contents were observed 200 m away from the emission source in both plant species, while the highest total sulfur foliar contents were observed 200 m away in C. hilariana and 800 m away in E. uniflora. E. uniflora presented decreased values of height growth rate, number of necrotic leaves, chlorophyll analysis (SPAD index) and transpiration, in relation to the distances from the emission source. C. hilariana showed decreased values of height growth rate, number of leaves, number of necrotic leaves, total ionic permeability, stomatal conductance, transpiration, net CO2 assimilation, and total dry matter, in relation to distances from the emission source. In relation to the days of exposure, both species presented increased number of necrotic leaves and foliar phytotoxicity index, and decreased values in the chlorophyll analysis. The two native plant species, both of which occur in the Brazilian Restinga, showed damage when exposed to emissions from an iron ore pelletizing factory. C. hilariana was considered the most sensitive species due to the decreased values in a higher number of variables after exposition.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Brazil is the second largest producer of iron (Fe) ore worldwide (Instituto Brasileiro de Mineração–IBRAM 2011), and an increasing activity has been observed in mining and processing industries of this metal (Kuki et al. 2009).
Located at the Brazilian coast, the Restinga can be defined as coastal areas constituted by Quaternary sandy deposits of marine origin and dunes built over these deposits by wind action (Araujo and Maciel 1998). This biome presents a great variety of communities with typical fauna and flora (Freire 1990; Rocha et al. 2003; Assis et al. 2004), which are subjected to abiotic adversities such as high temperatures, dry periods, constant wind, high salinity, and nutrient scarcity. This makes the structure and function of the Restinga different from those of any other environment (Scarano 2002).
However, this coastal biome is being impacted by the emission of pollutants from mining industries, which causes alterations in the structure, dynamics, and diversity of the exposed vegetation (Silva et al. 2006; Kuki et al. 2008, 2009; Pereira et al. 2009).
On the coast of Espírito Santo state, in the district of Ubu, there is an iron pelletizing factory which processes ore for exportation. This factory has records of suspended particulate matter emission levels that are considered to be below the air quality secondary standard allowed in Brazil (Lopes et al. 2002; CONAMA 1990). Nevertheless, some plant species can be sensitive to levels of air pollution that are regarded as low and may present symptoms, which would qualify them as bioindicators of air quality (De Temmerman et al. 2004).
Pelletizing factories are responsible for increasing atmospheric levels of several pollutants, especially sulfur dioxide (SO2) and iron suspended particulate matter (SPMFe) (Lopes et al. 2002; Grantz et al. 2003).
Sulfur dioxide is one of the six major air pollutants and can be absorbed by roots or through stomata, the latter by means of photosynthesis and respiration (Manninen and Huttunen 2000; Zhang et al. 2013). Its absorption by plants causes different responses depending on the species and on environmental conditions. Studies have reported damage to the photosynthetic apparatus (Swanepoel et al. 2007), stomatal density (Haworth et al. 2012), and carbon fixation efficiency (Chung et al. 2011).
Suspended particulate matter is composed of a mixture of several pollutants and contains particles of different sizes, origins, and chemical compositions. The dispersion dynamics of trace elements such as Fe may vary with different factors such as air temperature, chimney height, wind frequency and intensity, precipitation, and the presence of suspended water (marine aerosol) in the atmosphere (Grantz et al. 2003). In the ecosystem, particulate deposition can alter nutrient cycling, thus inhibiting their absorption; reducing growth, flowering, and fruiting; and altering species diversity due to local elimination of the most sensitive species, among other effects (Silva et al. 2006; Kuki et al. 2008).
Along with such problem, in coastal environments, the marine aerosol can adhere to suspended particles. Depending on its size, particulate deposition will either precipitate in the vicinity of the factory or be dispersed for miles inland. The high sodium and chloride concentration in the marine aerosol is extremely toxic to plants, and when adhered to particulate iron, they may produce leaf necrosis. Thus, this may also contribute to the elimination of local species that are intolerant to salt spray (Grantz et al. 2003).
The species Eugenia uniflora L. (Myrtaceae) and Clusia hilariana Schlecht (Clusiaceae) are sensitive to some air pollutants (Silva et al. 2005; Alves et al. 2008) and are indicated for active biomonitoring studies of air quality (Alves et al. 2008; Pereira et al. 2009; Neves et al. 2009).
E. uniflora and C. hilariana have distinct photosynthetic metabolisms, namely C3 and crassulacean acid metabolism (CAM), respectively. Unlike plants with C3 metabolism, CAM plants close their stomata during the day, and only at night, when their stomata are open, does atmospheric CO2 fixation occur, by the activity of the phosphoenolpyruvate carboxylase enzyme (Cushman and Bohnert 1997). Furthermore, the two species also present different anatomical features and metabolic pathways, which can determine their differential susceptibility to pollutants (Silva et al. 2005). Thus, we believed that E. uniflora and C. hilariana would present different responses to a SO2- and SPMFe-contaminated environment.
As the impacts of atmospheric pollution are poorly studied in tropical ecosystems, the aim of this study was to evaluate the responses in terms of growth and physiological parameters, and the degree of sensitivity of E. uniflora and C. hilariana to the atmospheric pollutants SO2 and SPMFe.
Material and methods
Studied species
The chosen species for the study were E. uniflora L. (Myrtaceae) and C. hilariana Schlecht. (Clusiaceae), due to their representativeness and frequency in the Restinga of Paulo César Vinha State Park (PCVSP); to the ease of their cultivation; and to their phenological, morphological, and physiological traits.
E. uniflora L. (Myrtaceae) is a shrub ca. 3 m high with C3 metabolism and occurs both in the backshore and in the Myrtaceae forest formations, on the Setiba Restinga. It possesses opposite phyllotaxis and simple, glossy, glabrous leaves. Its fruit is widely commercially exploited due to its sweet and tangy taste (Neves and Donato 1989).
C. hilariana Schlecht (Clusiaceae) is a tree with CAM metabolism (Franco 1989) that is widely distributed along the Setiba Restinga, especially in the open Clusia scrub, where its individuals predominate, forming bushes. It possesses opposite phyllotaxis and its leaves are thick and waxy. It is commercially exploited as an ornamental species and for the extraction of wood and resin, the latter for medicinal purposes (Schneider 1985).
Characterization of the studied area
The studied area belongs to the industrial complex of a pelletizing factory located in the municipality of Anchieta, on the coast of Espírito Santo state, at coordinates 20° 46′ 21.0″ S and 40° 34′ 52.3″ W (Fig. 1).

Map indicating the region of the pelletization factory and the stations of active biomonitoring
Climatic conditions were monitored at the meteorological station installed at the same line of the exposure transect, where wind speed and direction, rainfall, and ambient temperature were registered. Data were recorded on a daily basis, measurements being taken from 12:00 AM to 11:59 PM.
Concentrations of emitted SO2 and SPM, which were obtained at the station located 1700 m away from the emission source on the same transect of the studied area, were determined with a Tri-Gas equipment (Tri-Gas 1/110 V—Energética Indústria e Comércio LTDA, Brazil) and a Hi-Vol air sampler (AGVPTS1—Energética Indústria e Comércio LTDA, Brazil), respectively. Sulfur dioxide concentrations were determined by the hydrogen peroxide method.
Cultivation conditions and plant exposure
Saplings of both species ca. 4 months old provided by the PCVSP nursery garden were standardized according to height, number of leaves, and phytosanitary status. After being transferred to 2-kg plastic pots filled with a mixture of sand, soil, and humus (3:1:1), plants were put in an automatic irrigation system, in which the pots were connected through a wick to water reservoirs that guaranteed the maintenance of soil humidity at an adequate level.
Plants were subjected to the action of the gaseous and particulate emissions from the pelletizing factory for 7 months, under field conditions, from August 14, 2001 to March 12, 2002. They were placed in standardized stations according to the international standards for air quality control using bioindicators (Arndt and Schlutler 1985). The stations were distributed along a transect, which was determined according to the predominant wind direction (NE) in the region and placed 200 m (20° 46′ 25.0″ S and 40° 34′ 58.9″ W), 500 m (20° 46′ 28.0″ S and 40° 35′ 10.1″ W), 800 m (20° 46′ 35.8″ S and 40° 35′ 14.9″ W), 1400 m (20° 46′ 51.2″ S and 40° 35′ 29.8″ W), and 1700 m (20° 46′ 42.2″ S and 40° 35′ 47.3″ W) away from the emission source. This distribution had been previously established by a statistical study coupled to a sulfur dioxide dispersion modeling (Queiroz 1998), which was performed with an Industrial Source Complex Short Term simulator (ISCST3, Environmental Protect Agency—EPA, USA). The location of each station was determined with global positioning system (GPS).
Throughout the experiment, four evaluations were performed, regarding height, number of leaves, total chlorophyll content (SPAD index), number of necrotic leaves, and necrosis-based foliar phytotoxicity index. Measurements were taken at the beginning of the experiment and after 71, 118, and 211 days of exposure. Simultaneously with the last evaluation, measurements of gas exchange, membrane permeability, iron deposition on leaves, iron and sulfate quantification in plant matter, and total dry matter were performed.
Deposition of particulate matter on leaf surface
In order to determine particulate deposition on leaves, three leaves from all E. uniflora plants and two leaves from all C. hilariana plants were randomly chosen and individually washed with 20 ml of distilled water in pre-weighed glass flasks. The collected percolate was set aside at room temperature to allow deposition of the solid particulate matter on the bottom of the flasks. After 48 h, the flasks were carefully placed into a non-ventilated oven at 40 °C until complete water evaporation. After cooling, flasks were reweighed. Each washed leaf had its area measured by a plan meter (Delta MK2—Delta Devices Ltd, England). The amount of particulate matter deposited on the leaf was expressed as milligrams per square millimeter of leaf area (Prusty et al. 2005).
Iron and sulfur quantification in leaf dry matter
To determine iron and sulfur amounts in plant matter, 15 leaves of E. uniflora and 5 leaves of C. hilariana were rinsed with running water, then with deionized water, and dried with filter paper. After oven-drying at 75 °C, samples were ground through a 1-mm-mesh sieve. Iron content was determined by atomic absorption spectrophotometry and sulfate content by inductively coupled plasma (ICP). Measurements were taken in composite samples.
Growth analysis
In the two studied species among all distances from the emission source, the following parameters were analyzed: growth rate, number of leaves, and dry matter weight. Growth rate was obtained by the difference between the height values at the end of the exposition and the ones at the beginning. Total dry matter weight was obtained by drying roots and shoots in a drying oven with forced ventilation, until constant weight.
Foliar phytotoxicity
All leaves presenting necroses were quantified. Foliar phytotoxicity was determined by attributing grades to the percentage of necrotic leaf area, according to the following scale: 0—no necrosis, 1—with necrosis in up to 25 % of the leaf area, 2—with necrosis in 26 to 50 % of the leaf area, 3—with necrosis in 51 to 75 % of the leaf area, and 4—with necrosis in 76 to 100 % of the leaf area, as proposed by Bustamante et al. (1993). A general sampling was performed with the most representative adult leaves per pot.
Chlorophyll content (SPAD index)
The relative leaf greenness was measured by the nondestructive chlorophyll meter SPAD (model 501, Minolta, Japan). Each repetition was represented by the mean value of four measurements taken on adult leaves, located on the third (C. hilariana) and fourth (E. uniflora) nodes. All measured leaves were adult and non-senescent. All noninvasive measurements were performed on a leaf region whose distance from the tip was equivalent to one third of the entire leaf length.
Evaluation of gas exchange
Stomatal conductance (g s), transpiration (E), and net CO2 assimilation (A) were evaluated in adult leaves from the third node, with an infrared gas analyzer (IRGA; model LCA-2, Analytical Development Co. Ltda, Hoddesdon, England) with PLC-B chambers (model ADC, BioScientific Limited, Herts, England) between 7:00 and 10:00 AM in E. uniflora and between 9:00 and 11:00 PM in C. hilariana, due to the crassulacean acid metabolism of the latter (Liebig et al. 2001).
Membrane permeability
Total ionic permeability was determined in 15 leaf discs of E. uniflora and in 10 discs of C. hilariana (5 mm diameter, each), due to the higher leaf blade thickness of the latter. The leaf discs, which had been previously weighed and rinsed, were placed in a recipient with deionized water for 4 h, and the initial electrical conductivity of the solution (i) was then measured. After this, the solutions were transferred to an oven at 90 °C for 2 h, in order to provoke cell membrane rupture. After cooling, the final electrical conductivity of the solution (f) was measured. Membrane permeability was calculated by the formula i/(i + f) × 100, as proposed by Tarhanen et al. (1999).
Statistical analysis
The experimental design was completely randomized, and each species was evaluated separately since they have completely different physiologies. For each species, two factors were set. The first one was the station, with five levels (200, 500, 800, 1400, and 1700 m away from the emission source). The second one was the time of exposure (0, 71, 118, and 211 days). Data were transformed whenever they did not present homogeneity of variance nor normal distribution and were then submitted to analysis of variance. Whenever a factor or the interaction between factors was significant by F test at 5 % probability, means were submitted to Tukey’s test at 5 % probability for comparison.
Results
Characterization of the studied area
The climate of the studied region is type Aw; tropical hot and wet with two dry months (Köppen 1918). During the experimental period, northeast wind predominated in all months, with an average maximum speed of 1.9 m · s−1. Air temperature along the experimental period ranged from 20.71 to 25.62 °C, with the average value of 23.4 °C. Precipitation ranged from 53.9 to 201.4 mm per month, the heaviest rainfall being registered in November. The average for the experimental period was 88.7 mm.
Regarding the amount of total suspended particulates, only in February did the values exceed the secondary standard (66.8 μg · m−3, for a standard of 60 μg · m−3) (Fig. 2a). As for sulfur dioxide, the emission values remained below the secondary standard along the entire exposure period (Fig. 2b).

Emissions from the mining industry. a Total suspended particles and b sulfur dioxide. Secondary standard according to CONAMA resolution, March 1990
Deposition of particulate matter on leaf surface
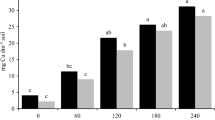
No difference regarding deposited particulate matter per leaf area in relation to distances from the emission source was detected in either species (Fig. 3a).

Deposition of particulate matter per leaf area (a), total iron content (b), and total sulfur content (c) in leaves of Eugenia uniflora and Clusia hilariana at different distances from the emission source (200, 500, 800, 1400, and 1700 m). Upper case letters compare Eugenia uniflora and lower case letters compare Clusia hilariana. Means followed by the same letter do not differ by Tukey’s test at 5 % probability. Error bars indicate the standard error
Iron and sulfur quantification in leaf dry matter
Iron and sulfur quantification in plant matter was performed using composite samples, which were obtained by gathering samples from repetitions of a same station. Iron amounts in leaf dry matter ranged between 895 and 273 mg · kg−1 and between 596 and 93 mg · kg−1 in E. uniflora and C. hilariana, respectively (Fig. 3b). Sulfur foliar contents in E. uniflora ranged between 764 and 151 mg · kg−1, while in C. hilariana, it ranged between 2762 and 146 mg · Kg−1 (Fig. 3c).
Growth analysis
Emissions from the factory interfered with growth of neither species along the analyzed distances (data not shown). E. uniflora plants showed the highest height growth rates at stations 200, 500, and 1700 m away from the emission source (Table 3), while C. hilariana plants showed the highest growth rates at 500, 800, and 1700 m away from the emission source (Table 4).
In E. uniflora, the number of leaves per pot increased along with the collection dates (Table 1). However, no significant difference was observed in the number of leaves in relation to distances from the emission source (Table 3). C. hilariana showed no difference along the days of exposure (Table 2), but there was a significant decrease in relation to distances from the emission source (Table 4).
In E. uniflora, there was no difference in total dry matter in relation to distances from the emission source (Table 3). In C. hilariana, the highest dry matter production occurred in the nearest station to the factory (Table 4).
Foliar phytotoxicity
Leaf necrosis was the main visible symptom observed in the plants. The highest number of necrotic leaves was detected in the last data collection in E. uniflora and from the penultimate collection in C. hilariana (Tables 1 and 2). Reduction in relation to distances from the emission source was observed at 500 m in E. uniflora and at 200 m in C. hilariana (Tables 3 and 4). In E. uniflora, there was interaction between evaluation dates and the analyzed stations (Fig. 4). Plants located 1400 m away from the emission source showed a higher number of necrotic leaves from October. In December, plants located 800, 1400, and 1700 m away showed the highest number of necrotic leaves. In the last evaluation date, all stations showed the highest number of necrotic leaves. Plants located at the nearest station to the emission source became necrotic only by the time of the last evaluation date (Fig. 4).

Interactions between collection dates and distances from the emission source in plants of Eugenia uniflora exposed to the depositions from the factory for 7 months, from August 2001 to March 2002, regarding number of necrotic leaves. Asterisks indicate significance in the interactions. Error bars indicate the standard error
From the second collection date, the percentage of necrotic leaf area (necrosis-based foliar phytotoxicity index) in both species was higher than in the first collection and did not change on the subsequent dates (Tables 1 and 2). From the second collection, there was a high percentage of necrotic leaf area in E. uniflora (Tables 1 and 2). The distance to the emission source did not influence the percentage of necrotic leaves on the studied species (Tables 3 and 4). In E. uniflora, the interaction between collection dates and distances was significant, the highest percentages of necrotic leaves being registered at 800 and 1700 m away, in February (Fig. 4).
Chlorophyll content (SPAD index)
Chlorophyll concentration decreased gradually along with the evaluation dates in both species (Tables 1 and 2). Only in E. uniflora where there was a significant difference on total chlorophyll content among the evaluated distances, the highest average being registered 800 m away from the emission source (Table 3).
Evaluation of gas exchange
Stomatal conductance in E. uniflora did not change along with distances from the emission source, while in C. hilariana, there was a decrease from 800 m (Tables 3 and 4).
On both studied species, a decreasing tendency on transpiration rates was observed in relation to distances from the emission source (Tables 3 and 4).
Net CO2 assimilation in relation to distances from the emission source did not vary in E. uniflora, while in C. hilariana, the lowest values were observed on the stations located 800 and 1400 m away (Tables 3 and 4).
When comparing gas exchange between the plants with the highest and lowest foliar iron amounts, a significant reduction was observed on photosynthesis values in E. uniflora. In C. hilariana, both stomatal conductance and transpiration increased with increasing iron contents (Table 5).
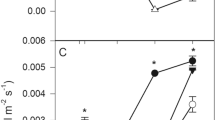
Nonlinear correlation coefficients between foliar sulfur contents and parameters g s and A were estimated in C. hilariana as 98 and 88 %, respectively (Fig. 5).

Gas exchange according to sulfur foliar content, in Clusia hilariana exposed to particulate deposition of metallic iron for 7 months. a Stomatal conductance (g s) and b net CO2 assimilation (A)
Membrane permeability
Significant difference on total ionic permeability of cell membranes in relation to distances from the emission source was observed only in C. hilariana, the highest values being detected on plants located 500 m away from the emission source, followed by plants located 200 m away (Tables 3 and 4).
Discussion
Data on SPMFe and SO2 emissions along the 7 months throughout which the present experiment was conducted proved that the pelletizing factory was responsible for the increased levels of these pollutants in the atmosphere, which, in February, exceeded the secondary standard for deposition of suspended particles. Regarding sulfur dioxide, the emission values were acceptable according to CONAMA resolution 03/1990 (National Council for the Environment, the Brazilian agency that legislates over the National Environmental Policy) (CONAMA 1990). The emissions from the factory could be confirmed by the foliar iron deposition and by the quantification of iron and sulfur contents on the E. uniflora and C. hilariana plants exposed at different distances from the emission source.
C. hilariana leaves are extremely waxy (Schneider 1985; Silva et al. 2005). Epicuticular wax features might have contributed to the lower iron retention on the leaf of this species, in relation to E. uniflora. The plaque-shaped epicuticular wax visualized by Silva et al. (2005) might have favored the non-adherence of iron particles to the leaves. Some specific features of the epicuticular wax, such as its chemical composition and the structure of its layers, might be related to its potential for capturing atmospheric particulates (Kaupp et al. 2000; Jouraeva et al. 2002; Dzierżanowski et al 2011). Along with the epicuticular wax, other factors, such as precipitation and wind, may have been capable of reducing foliar iron deposition, as verified with other pollutants (MacLean et al. 1989).
The total iron content found on E. uniflora and C. hilariana leaves after 7 months of exposure to particulate deposition exceeded the needs of crop plants cited by Larcher (1995) and Dobermann and Fairhurst (2000). In C. hilariana, on plants located 1400 m away from the emission source, iron content was within the limits of normality for crop plants, although the plants presented considerable necrosis. Iron contents on plants located 200 m away from the emission source reached 895 and 596 mg · kg−1 in E. uniflora and C. hilariana, respectively, exceeding the toxicity limit stipulated for crop plants, of 500 mg · kg−1 (Marschner 1995; Dobermann and Fairhurst 2000). These data suggest that for these native species, the critical level is lower than the one stipulated for crop plants.
E. uniflora seems to demonstrate a capacity for acclimation in the presence of the factory emissions. This could be verified by the production of a higher number of leaves with increasing days of exposure. C. hilariana, on the other hand, showed higher number of new leaves on the nearest stations to the emission source. Iron and sulfur, both essential for plants, may have acted as foliar fertilizers on the studied species, thus stimulating the production of leaf biomass, as reported to Pinus ponderosa by Momen et al. (2002).
Although no difference among the sampled stations could be observed, the height of the factory chimney as well as climatic conditions seem to have produced a protective effect against the deposition of particulate matter on the C. hilariana plants located nearest to the chimney, since plants of the station located 200 m away from the emission source presented the highest total dry matter and the lowest number of necrotic leaves. In E. uniflora, however, the lowest number of necrotic leaves was found on plants located 500 m away from the emission source. These results are in agreement with those presented by Lopes et al. (2002), according to which saplings of native plants from the Restinga exposed to similar depositions to the ones of the present work showed the highest number of necroses on plants located 800 and 1400 m away from the emission source. According to Grantz et al. (2003), depending on climatic conditions and on topographic features, nanoparticles can remain in the air for days or even months and may be transported from 1000 to 10,000 km away from the emission source.
According to Spiro and Stigliani (2002), the longer the time of exposure to atmospheric pollutants, the higher is the percentage of damage presented by the plants. This was also proved in the present study, with both E. uniflora and C. hilariana, since the longer was the period of exposure to the factory emissions, the higher were the number of necrotic leaves and the foliar phytotoxicity index found, and the lower were the total chlorophyll contents.
The foliar iron and sulfur amounts found in C. hilariana plants located 200 m away from the emission source were responsible for damage to cell membranes, as shown by the high values of total ionic permeability observed in plant samples from that station. The significant increase in electrolyte leakage suggests that the stress caused by the excess iron, and sulfur may be related to a possible oxidative stress in this species, through formation of reactive oxygen species (ROS), which results in several physiological disturbances such as peroxidation of the phospholipid membrane and damage to plant tissues (Becana et al. 1998; Hänsch and Mendel 2005; Sinha and Saxena 2006; Karuppanapandian et al. 2011).
The decreased SPAD index in relation to days of exposure in both studied species indicates that two processes may have been affected: chlorophyll synthesis and/or its degradation. The synthesis may have probably been reduced due to the iron deposited on the leaf, which causes shading of the organ, limits incident radiation, and increases foliar temperature (Pereira et al. 2009). Chlorophyll degradation, on the other hand, could be the consequence of oxidative stress produced by the excess free iron ions, after iron absorption by the leaf (Jeong and Guerinot 2009; Jucoski et al. 2013).
The decrease in g s, E, and A occurred at the largest distances from the emission source, especially in C. hilariana, which suggests a protective effect regarding deposition of particulate matter from the chimney on the nearest plants to the emission source. Chimney height, wind speed, and the presence of marine aerosol can favor the deposition of particulate iron at larger distances from the emission source (Grantz et al. 2003).
The obstruction of stomatal pores (Rocha et al. 2014) and the decreased incident luminous radiation by SPMFe may have been responsible for the decreased g s and E in C. hilariana. The reduction on A, on the other hand, was the consequence of such stomata obstruction and of the decreased g s and E, which may have possibly also decreased the carboxylation capacity on this species. Similar results were observed by Pereira et al. (2009), according to which the increased leaf temperature, which is caused mainly by the decreased incident luminous radiation, and the decreased conductance may be the explanation for alterations in gas exchange parameters in C. hilariana plants exposed to SPMFe.
Plants that accumulated the highest iron amounts in their leaves seem to have presented higher damage to their stomata. In E. uniflora, this fact could be verified due to decreased photosynthesis (A), although no variation in g s and E was observed. In C. hilariana, the increase in g s and E in plants with higher iron amounts did not reflect in an increased A. In that case, it seems that an alteration on the capacity of stomatal movements has occurred. In rice plants subjected to excess iron, the responsible factor for decreased photosynthetic parameters was stomatal limitation (Pereira et al. 2013).
C. hilariana accumulated ca. 10 g · kg−1 of sulfur in plant matter, a much higher amount than the one found by Klumpp et al. (1998), who exposed plants of Psidium guajava and Psidium cattleianum on several sites of the industrial complex of Cubatão city (São Paulo state, Brazil).
The correlation coefficients between foliar sulfur and g s and between foliar sulfur and A in C. hilariana suggest an effect of SO2 on stomatal conductance and net CO2 assimilation in this species. According to Rennenberg et al. (1996a, b), since plants that present crassulacean acid metabolism keep stomata open at night, they get exposed to a higher amount of gaseous air pollutants. During the day, the high temperatures allow a greater dissipation of gaseous emissions, conversely to what happens at night, when the lower temperatures hamper the diffusion of gases to the environment, rendering leaves to be exposed for a longer period of time to the gaseous pollutants. Thus, CAM plants show higher sensitivity to SO2 and this may be related to the diurnal differences in cellular capacity for SO2 detoxification (Swanepoel et al. 2007). These factors may have been the reason for the greater damage caused by SO2 exposure to C. hilariana than the one caused to E. uniflora.
Conclusions
The two species showed different strategies when exposed to the pollutants. E. uniflora presented decreased values of height growth rate, number of necrotic leaves, chlorophyll analysis (SPAD index), and transpiration, in relation to the distances from the emission source. C. hilariana presented decreased values of height growth rate, number of leaves, number of necrotic leaves, total ionic permeability, stomatal conductance, transpiration, net CO2 assimilation, and total dry matter, in relation to the distances from the emission source. In relation to days of exposure, both species presented increased values of number of necrotic leaves and foliar phytotoxicity index, and decreased values in the chlorophyll analysis. E. uniflora seems to demonstrate a capacity for acclimation in the presence of the stressing agent, producing a higher number of new leaves with increasing days of exposure to the pollutants.
The native plant species that occur in the Brazilian Restinga showed damage when exposed to emissions from an iron ore pelletizing factory. C. hilariana presented higher sensitivity to SO2 and greater alterations in the analyzed parameters in comparison to E. uniflora, thus being considered the most sensitive species.
References
Alves ES, Tresmondi F, Longui EL (2008) Leaf anatomy of Eugenia uniflora L. (Myrtaceae) in urban and rural environments, São Paulo State, Brazil. Acta Bot Bras 22:241–248 (in Portuguese)
Araujo DSD, Maciel NC (1998) Restingas of Rio de Janeiro state: biodiversity and preservation. Bol FBCN 25:27–51 (in Portuguese)
Arndt U, Schlutler C (1985) Effect-based determination of the immission load in specific forest decline areas in Baden-Württemberg by the use of bio-indicators (active monitoring with indicator plants). Statuskoll. z. Arbeit des PEF, Karlsruhe 5.bis 7.3, pp. 17-41 (in German)
Assis AM, Pereira OJ, Thomaz LD (2004) Phytosociology of a restinga forest in the “Paulo César Vinha” State Park, Setiba, Guarapari (Espírito Santo). Rev Bras Bot 27:349–361 (in Portuguese)
Becana M, Moran JF, Iturbe-Ormaetxe I (1998) Iron-dependent oxygen free radical generation in plants subjected to environmental stress: toxicity and antioxidant protection. Plant Soil 201:137–147
Bustamante M, Oliva MA, Sant’Anna R, Lopes NF (1993) Fluorine sensitivity on soybean plants. Rev Bras Fisiol Veg 5:151–157 (in Portuguese)
Chung CY, Chung PL, Liao SW (2011) Carbon fixation efficiency of plants influenced by sulfur dioxide. Environ Monit Assess 173:701–707
CONAMA (1990) Resolução no 003, de 28 de junho de 1990. Brasil. Ministério do Meio Ambiente. Conselho Nacional de Meio Ambiente. Diário Oficial da União, Brasília, 22 ago. 1990, Seção 1, p. 15937–15939. Available at: http://www.mma.gov.br/port/conama/index.cfm. Accessed 1 July 2014
Cushman JC, Bohnert HJ (1997) Molecular genetics of crassulacean metabolism. Plant Physiol 113:667–676
De Temmerman L, Bell JNB, Garrec JP, Klumpp A, Krause GHM, Tonneijck AEG (2004) Biomonitoring of air pollutants with plants—considerations for the future. In: Klumpp A, Ansel W, Klumpp G (eds) Urban air pollution, bioindication and environmental awareness. Cuvillier Verlag, Göttingen
Dobermann A, Fairhurst T (2000) Rice: nutrient disorders & nutrient management. Potash & Phosphate Institute, Potash & Phosphate Institute of Canada, and International Rice Research Institute, Singapore and Los Baños, pp. 1-4
Dzierżanowski K, Popek R, Gawrońska H, Sæbø A, Gawroński SW (2011) Deposition of particulate matter of different size fractions on leaf surfaces and in waxes of urban forest species. Int J Phytoremediation 13:1037–1046
Franco AC (1989) The genus Clusia as example of studies on plant responses to stress in tropical environments. In: Scarano FR, Franco AC (eds) Ecophysiological strategies of xerophytic and amphibious plants in the neotropics, vol IV. Series Oecologia Brasiliensis., Rio de Janeiro, Brazil, pp 65–85
Freire MSB (1990) Floristic survey on Natal Sandy Dunes State Park. Acta Bot Bras 4:41–59 (in Portuguese)
Grantz DA, Garner JHB, Johnson DW (2003) Ecological effects of particulate matter. Environ Int 29:213–239
Hänsch R, Mendel RR (2005) Sulfite oxidation in plant peroxisomes. Photosynth Res 86:337–343
Haworth M, Elliott-Kingston C, Gallagher A, Fitzgerald A, McElwain JC (2012) Sulphur dioxide fumigation effects on stomatal density and index of non-resistant plants: implications for the stomatal palaeo-[CO2] proxy method. Rev Palaeobot Palynol 182:44–54
Instituto Brasileiro de Mineração–IBRAM (2011) Information and analysis of the Brazilian mineral economy. IBRAM, Brasília (in Portuguese)
Jeong J, Guerinot ML (2009) Homing in on iron homeostasis in plants. Trends Plant Sci 14:280–285
Jouraeva VA, Johnson DL, Hassett JP, Nowak DJ (2002) Differences in accumulation of PAHs and metals on the leaves of Tilia × euchlora and Pyrus calleryana. Environ Pollut 120:331–338
Jucoski GO, Cambraia J, Ribeiro C, Oliveira JA, de Paula SO, Oliva MA (2013) Impact of iron toxicity on oxidative metabolism in young Eugenia uniflora L. plants. Acta Physiol Plant 35:1645–1657
Karuppanapandian T, Moon JC, Kim C, Manoharan K, Kim W (2011) Reactive oxygen species in plants: their generation, signal transduction, and scavenging mechanisms. Aust J Crop Sci 5:709–725
Kaupp H, Blumenstock M, McLachlan MS (2000) Retention and mobility of atmospheric particle-associated organic pollutant PCDD/Fs and PAHs on maize leaves. New Phytol 148:473–480
Klumpp A, Domingos M, Moraes RM, Lima JS (1998) Effects of complex air pollution on tree species of the Atlantic Rain Forest near Cubatão, Brazil. Chemosphere 36:989–994
Köppen W (1918) Klassifikation der Klimate nach Temperatur, Niederschlag und Jahreslauf. Petermanns Mitt 64:193–203
Kuki KN, Oliva MA, Pereira EG (2008) Iron ore industry emissions as a potential ecological risk factor for tropical coastal vegetation. Environ Manag 42:111–121
Kuki KN, Oliva MA, Costa AC (2009) The simulated effects of iron dust and acidity during the early stages of establishment of two coastal plant species. Water Air Soil Pollut 196:287–295
Larcher W (1995) Physiological plant ecology, 3rd edn. Springer, Germany, p 506
Liebig M, Scarano FR, Mattos EA, Zaluar HLT, Lüttge U (2001) Ecophysiological and floristic implications of sex expression in the dioecious neotropical CAM tree Clusia hilariana Schltdl. Trees 15:278–288
Lopes SA, Oliva MA, Martinez CA (2002) Impact of sulfur dioxide emissions and iron particulate matter deposition on Restinga plant species (Anchieta city, Espírito Santo state, Brazil): ecophysiological evaluation. In: Espíndola ELG, Paschoal CMR, Rocha O, Bohrer MBC, Oliveira Neto AL (eds) Ecotoxicology—perspectives for the XXI century. RiMa Artes e Textos, São Carlos, pp 53–71 (in Portuguese)
Maclean DC, Schneider RE, Hansen KS, Troiano JJ (1989) Effects of simulated acid rain on uptake, accumulation, and retention of fluoride in forage crops. Water Air Soil Pollut 43:191–198
Manninen S, Huttunen S (2000) Response of needle sulphur and nitrogen concentrations of Scots pine versus Norway spruce to SO2 and NO2. Environ Pollut 107:421–436
Marschner H (1995) Mineral nutrition of higher plants, 2nd edn. Academic Press, California, California
Momen B, Anderson PD, Houpi JLJ, Helms JA (2002) Growth of ponderosa pine seedlings as affected by air pollution. Atmos Environ 36:1875–1882
Neves LJ, Donato AM (1989) Contributions to the study on Eugenia uniflora L. (Myrtaceae). Bradea 5:275–284 (in Portuguese)
Neves NR, Oliva MA, da Cruz CD, Costa AC, Ribas RF, Pereira EG (2009) Photosynthesis and oxidative stress in the restinga plant species Eugenia uniflora L. exposed to simulated acid rain and iron ore dust deposition: potential use in environmental risk assessment. Sci Total Environ 407:3740–3745
Pereira EG, Oliva MA, Kuki KN, Cambraia J (2009) Photosynthetic changes and oxidative stress caused by iron ore dust deposition in the tropical CAM tree Clusia hilariana. Trees 23:277–285
Pereira EG, Oliva MA, Rosado-Souza L, Mendes GC, Colares DS, Stopato CH, Almeida AM (2013) Iron excess affects rice photosynthesis through stomatal and non-stomatal limitations. Plant Sci 201–202:81–92
Prusty BAK, Mishra PC, Azzez PA (2005) Dust accumulation and leaf pigment content in vegetation near the national high way at Sambalpur, Orissa, India. Ecotoxicol Environ Saf 60:228–235
Queiroz RS (1998) Evaluation of the Samarco Mining Company environmental monitoring network of sulfur dioxide and particulate matter. Report, 22p (in Portuguese)
Rennenberg H, Herschbach C (1996) Responses of plants to atmospheric sulphur. In: Yunus M, Iqbal M (eds) Plant response to air pollution. John Wiley, Chincester, pp 285–293
Rennenberg H, Herschbach C, Polle A (1996) Consequences of air pollution on shoot-root interactions. J Plant Physiol 148:269–301
Rocha CFD, Bergallo HG, Alves MAS, Van Sluys M (2003) Biodiversity in the largest forest remnants in the state of Rio de Janeiro and in the Restingas of the Atlantic Rain Forest. Editora RiMa, São Carlos (in Portuguese)
Rocha DI, Silva LC, Pereira EG, Sant’Anna-Santos BF, Gontijo ER, Oliva MA (2014) Early detection of injuries in leaves of Clusia hilariana Schletlendal (Clusiaceae) caused by particulate deposition of iron. Revista Árvore 38:423–432
Scarano FR (2002) Structure, function and floristic relationships of plant communities in stressful habitats marginal to the Brazilian Atlantic Rainforest. Ann Bot London 90:517–524
Schneider SZ (1985) ‘Leaf anatomy of Clusia hilariana Schlechtendal and Clusia spiritu-sanctensis G. Mariz et Weinberg (Guttiferae) occurring in the state of Espírito Santo, Brazil’, M.S. Thesis, Department of Biological Sciences, Universidade Federal do Rio de Janeiro, 162 pp (in Portuguese)
Silva LC, Oliva MA, Azevedo AA, Araújo JM, Aguiar RM (2005) Micromorphological and anatomical alterations caused by simulated acid rain in restinga plants: Eugenia uniflora and Clusia hilariana. Water Air Soil Pollut 168:129–143
Silva LC, Oliva MA, Azevedo AA, Araújo JM (2006) Responses of restinga plant species to pollution from an iron pelletization factory. Water Air Soil Pollut 175:241–256
Sinha S, Saxena R (2006) Effect of iron on lipid peroxidation, and enzymatic and non-enzymatic antioxidants and bacoside—a content in medicinal plant Bacopa monnieri L. Chemosphere 62:1340–1350
Spiro TG, Stigliani MW (2002) Chemistry of the environment. Prentice Hall of India, New Delhi
Swanepoel JW, Kruger GHJ, Van Heerden PDR (2007) Effects of sulphur dioxide on photosynthesis in the succulent Augea capensis Thunb. J Arid Environ 70:208–221
Tarhanen S, Metsärinne S, Holopainen T, Oksanen J (1999) Membrane permeability response of lichen Bryoria fuscescens to wet deposited heavy metals and acid rain. Environ Pollut 104:121–129
Zhang X, Zhou P, Zhang W, Zhang W, Wang Y (2013) Selection of landscape tree species of tolerant to sulfur dioxide pollution in subtropical China. Open Journal of Forestry 3:104–108
Acknowledgments
The authors thank for the research productivity scholarships granted to L.C. Silva (309170/2012-5) and A.A. Azevedo (312190/2013-1) and the agreement firmed between the Federal University of Viçosa (UFV) and SAMARCO Mining Company.
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible editor: Philippe Garrigues
Rights and permissions
About this article

Cite this article
da Silva, L.C., de Araújo, T.O., Martinez, C.A. et al. Differential responses of C3 and CAM native Brazilian plant species to a SO2- and SPMFe-contaminated Restinga. Environ Sci Pollut Res 22, 14007–14017 (2015). https://doi.org/10.1007/s11356-015-4391-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11356-015-4391-0