
Abstract
In the coastal zone of the Espírito Santo state, Brazil, fragments of restinga, which form a natural ecosystem, share their space with an increasing number of iron ore industries. The iron ore dust and SO2 originating from the industry processing activities can interfere with the vegetation of the adjacent ecosystems at various levels. This study was undertaken in order to evaluate the effects of industry emissions on representative members of the restinga flora, by measuring physiological and phenological parameters. Foliar samples of Ipomoea pes caprae, Canavalia rosea, Sophora tomentosa, and Schinus terebinthifolius were collected at three increasing distances from an ore industry (1.0, 5.0, and 15.0 km), and were assessed for their dust deposition, chlorophyll, and Fe content. Phenological monitoring was focused on the formation of shoots, flowers, and fruits and was also performed throughout the course of a year. The results showed that the edaphic characteristics and the mineral constitutions of the plants were affected by industry emissions. In addition, the chlorophyll content of the four species increased with proximity to the industry. Phenological data revealed that the reproductive effort, as measured by fruit production, was affected by emissions and S. tomentosa was the most affected species. The use of an integrative approach that combines biochemical and ecological data indicates that the restinga flora is under stress due to industry emissions, which on a long-term basis may put the ecosystem at risk.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
It is widely accepted that atmospheric pollution can trigger environment-associated problems. A stationary source of pollutants, such as ore industry, can generate and emit considerable amounts of particulate matter (PM) and sulfur dioxide (SO2), thus increasing the deposition of wet and dry materials on ecosystems in the vicinity (fLopes et al. 2000; Grantz et al. 2003).
Coarse and fine PM can have a variety of effects on living organisms, including chemical and physical injuries. In particular, plants are prone to suffer from PM on a larger scale, due mainly to their inability to escape from agents causing stress (Yunus and Iqbal 1996). Depending on its size, the PM deposited on the leaf surface can affect the plant’s metabolism by blocking light, obstructing stomatal apertures, increasing leaf temperature, and altering pigment and mineral contents of the leaf (Paling et al. 2001; Naidoo and Chirkoot 2004). Vegetation damage imposed by PM can also be enhanced if trace metals are present in the PM or are delivered through simultaneous acid deposition. Once deposited in the soil, the metals may accumulate to phytotoxic levels, and in association with low pH, they can modify substrate composition and nutrient availability to the plants (Farmer 1993; Uhling and Juntilla 2001). The combination of these effects can contribute to the development of oxidative stress and alter plant growth and reproduction, altering the dynamics and composition of ecosystems (Wong et al. 1978; Gallego et al. 1996; Bartakova et al. 2001; Alvarez et al. 2003; Kozlov and Zvereva 2004). Nonetheless, much of the impact caused by atmospheric pollution are time- and space-related; therefore, the effects may vary with the sensitivity of the organism but also with season or period of exposure to pollutants.
Restinga is a tropical coastal ecosystem of quaternary origin found along the Brazilian littoral. The vegetation that comprises this ecosystem has a crucial role on the stability of the sand dunes and the security of its biodiversity (Pereira 1990). The coastline of the Espírito Santo state, Brazil, hosts fragments of restinga, and a growing number of iron ore industries. The industrial activities involve processing, beneficiation, stocking, and ship-loading of ore. As a result, SO2 and iron ore dust, a type of dry PM, are constantly added into the atmosphere, and may occur through spontaneous or unpredictable spills.
The onset of ecological risk in a natural ecosystem due to industry-based pollutants can be detected in its vegetation. Although coastal vegetation is adapted to marine aerosol, a natural form of airborne particles (McCune 1991; Barrick 2003), the addition of PM as iron dust can impose a new form of stress to the vegetation. The impact of this pollutant can affect multiple levels of biological organization. While isolated effects of pollutants on plants may indicate the integrity of a particular species, an integrated view of the effects on different species can help evaluate the integrity of the vegetation as a whole (Dale and Beyeler 2001).
The restinga ecosystem is protected by law, and many efforts have been implemented to ensure its preservation, despite intense anthropogenic disturbance. However, few studies concerning the impact caused by the ore industry activities are available. Through ecological and physiological analyses, the aim of the present study was to investigate the effects of industry emissions on representative members of the restinga flora.
Materials and Methods
Study Site
Fragments of restinga occur in Anchieta, in the state of Espírito Santo. According to Köppen’s classification, the climate in this region is Aw, which is tropical, warm, and humid with a nonpronounced dry season during the winter months. The annual precipitation is about 900 mm, the average air humidity is 70 %, and the average temperature is 23.4° C.
An iron ore industry complex, located near a highway and 0.5 km from the seashore (20° 46′ 21.0″ S and 40° 34′ 52.3″ W), is surrounded by native flora. The industry plant includes a stockyard, two furnaces and a port terminal for shipment of the iron pellets and sinter, the final products. The main ore exploited is itabirito, which is basically composed of hematite (Fe2O3) and quartz (SiO3). The beneficiation and handling of the ore are the processes that generate iron ore particles, mainly coarse particulates (Table 1). In addition, SO2 is emitted by the furnaces during the formation of pellets, which increases the total particulate matter in suspension (TPM) or dust. Personal communications, which have also been confirmed by visual observations, suggest that the airborne iron particles may precipitate as far as 10.0 km from the source. To collect data regarding pollutant emissions and climate, emissions and microclimate record stations were positioned within the industry plant. Due to vandalism, no stations were assembled outside of the industry complex.
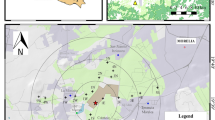
To study the effects of these pollutants on vegetation, three sites were selected along the coast, based on their proximity to the source and the occurrence of the plant species of interest. The distances of the first, second, and third site were 1.0, 5.0, and 15.0 km, respectively, from the industry complex (Fig. 1).

Map of Anchieta municipality, Espírito Santo state-Brazil, showing the location of the 3 assessed sites (1.0, 5.0, and 15.0 km). The (⋆) represents the location of the iron ore industry plant
Plant Material and Experiments
Four species were chosen to have their phenological and biochemical traits assessed: Canavalia rosea DC, (Leguminosae-Papilionoideae), Ipomoea pes caprae L. (Convolvulaceae), Schinus terebinthifolius Rardii (Anarcadiaceae), and Sophora tomentosa L. (Leguminosae-Papilionoideae). The selection was based on ecological importance and abundance in the areas of study. Canavalia rosea and I. pes caprae are both crawling, fast growing herbaceous species. Their pattern of growth helps protect sand dunes from shifting (Gross 1993; Patiño et al. 2002). Schinus terebinthifolius and S. tomentosa are arbustive species. Schinus terebinthifolius is a species with great ecological plasticity, commonly found in several tropical ecosystems, while the distribution of S. tomentosa is restricted to tropical coastal ecosystems (Lenzi and Orth 2004; Nogueira and Arruda 2006).
At each sample site, 10 individuals per species were selected and marked. Plants were selected for health and size homogeneity. Since S. terebinthifolius is a dioecious species, only female plants were marked. To mark C. rosea and I. pes caprae, 10-m2 plastic frames were used to mark out individual parts of the dense ground cover produced by these crawling species.
The deposition of TPM on the leaf surface and the iron (Fe) and pigment content in the leaves were analyzed every trimester in the year 2005. The TPM was determined for plants of all three sites, while the Fe and pigment analysis was only performed on plants at the first and third sites.
The TPM deposition was estimated for five leaves, from the third or fourth nodes, each randomly collected from 5 out of the 10 marked plants. The leaves were individually washed using a soft paintbrush with 20 ml of distilled water in pre-weighed glass flasks. The collected percolate was set aside at room temperature to allow the deposition of solid particulate matter at the bottom of the flasks. After 48 hours, the flasks were carefully transferred into a nonventilated oven at 40° C, until the water completely evaporated. After cooling, the flasks were re-weighed. Each washed leaf had its area measured by a plan meter (Delta MK2 – Delta Devices Ltd, England). The amount of TPM deposit on the leaf was expressed as mg per mm2 of leaf area (Prusty et al. 2005).
To estimate the content of foliar pigments, three foliar discs (59 mm2) were extracted from the leaves used to determine TPM deposition. The discs were placed in darkened glass vessels containing 7 ml of dimethyl sulfoxide (DMSO) at room temperature. The pigments were extracted into the DMSO without grinding. The incubation period varied from 48 hours for I. pes caprae, S. terebinthifolius and S. tomentosa, to 72 hours for the thicker foliar discs of C. rosea. A 3.0-ml aliquot of the pigment extract was transferred to a cuvette and the absorbances were read at 665.1 nm and 649.1 nm for chlorophyll a (Chl a) and chlorophyll b (Chl b), respectively. All readings were carried out in semi-dark conditions using a spectrophotometer (U-2000 UV/Vis – Hitachi Ltd, Japan). The chlorophyll content was calculated following the equations proposed by Wellburn (1994), Chl a = 12.47A 665.1−3.45A 649.1 and Chl b = 21.99A 649.1−5.32A 665.1. The chlorophyll concentrations were expressed as pigment content per area (μg mm−2).
For determination of Fe content, 10 to 15 leaves were randomly collected from three sample plants. These leaves were thoroughly washed in distilled water and dried at 70° C for 72 hours in a ventilated oven. The dry leaves were ground in an electrical stainless steel mill and 0.5 g samples (n = 3) were digested in concentrated HNO3:HClO4 acid solution (3:1 v/v) at 200° C. Digests were analyzed for Fe by atomic absorption spectrophotometry (GBC Avanta – GBC Scientific Equipment Ltd, Australia) (Kampfenkel et al. 1995).
Phenological phases of the four species were monitored monthly over the year of 2005. This monitoring survey was carried out on the marked plants from the first (1.0 km) and third (15.0 km) sites. The intensity of shoot, flower, and fruit production was estimated for each marked plant, on a scale of 0–4, using the semi-quantitative score method proposed by Fournier (1974). The results were then transformed into percentages.
Samples of the soil from the 1.0 and 15.0 km sites were analyzed for their physical and chemical composition by a commercial soil laboratory. Meteorological and emission data were also collected from microclimate stations located in the industry plant. The amount of SO2 in the atmosphere was determined by a Tri-gas monitor (TriGas 1/110 V - Energética Indústria e Comércio LTDA, Brazil) and the amount of TPM in suspension in the air was determined by a Hi-Vol air sampler (AGVPTS1 – Energética Indústria e Comércio LTDA, Brazil).
Statistical Analysis
The data were submitted to analysis of variance (ANOVA) using a statistical program package (SAEG/UFV), and the effects of significant interactions were examined in detail. The means of each parameter were further compared by Tukey’s test at p ≤ 0.05.
Results
Meteorology, Emissions, and Soil Properties
The rainfall precipitation in the area occurred mainly in the first semester of the year when the research was conducted, with its peak at the beginning of the winter season (Fig. 2a). The maximum and the minimum average temperatures in the region did not exceed 25° C and 15° C, respectively. The wind direction was predominantly northeastern, with an average high speed of 1.9 m s−1.

Meteorological (a) and emission (b, c) data from 2005 collected from a microclimate station located in the vicinity of the iron ore plant
Throughout the year, the local emission reports showed that the SO2 level in the air was below the primary standard (80 μg m−3) established by the Brazilian National Council for the Environment (Resoluções CONAMA 1992). Higher values of TPM in suspension were detected in the second semester, probably due to low levels or an absence of precipitation. In three specific months, January, August, and October, the TPM in suspension (Fig. 2b, c) exceeded the threshold of the primary standard (80 μg m−3) as established by CONAMA. The nitrogen oxide levels (NOx) were not available at the time of the study.
The soil from the sites located at 1.0 and 15.0 km from the industry showed different characteristics (Table 2). The 1.0-km site showed lower mineral content and high acidity when compared to the site located at 15.0 km from the source of the emissions. The iron content of the soil samples collected from the 1.0-km site was also higher than the 15.0-km site.
Plant Experiments
Total Particulate Matter Deposition
The deposition of dust or TPM on the leaves of I. pes caprae, C. rosea, S. tomentosa, and S. terebinthifolius, was significantly affected by the interaction between time and location of analyses (p ≤ 0.05, f test). The amount of suspended particulate matter tended to be greater during the second semester rather than in the first semester of the year. This pattern is expected, since in the second semester the rainfall was scarce, which would lead to greater precipitation of solid particulate matter (Fig. 2c).
Indeed, all four species examined, and notably S. tomentosa, showed high TPM deposition in the period of low rainfall. The third quarterly analyses (in September) registered the highest amount of dust intercepted by most species. In addition, the pattern of TPM deposition varied between the sites (Fig. 3). The plants located at the 1.0-km site showed a greater deposit of TPM on their leaves, while the plants growing at the 15.0 km site had less particles deposited on their leaves. These results confirm that the influence of the airborne particles is minimized with increased distance from the pollutant source.

Mean spatial and temporal variation of the Total Particulate Matter (TPM) deposited on the leaf surface (n = 5) of Ipomoea pes caprae, Canavalia rosea, Sophora. Tomentosa, and Schinus. terebinthifolius. Bars marked with different capital and lower case letters differ from each other on a temporal and spatial basis, respectively. Probability level p ≤ 0.05 (Tukey’s test)
Total Iron Content
The total Fe content in the leaves of I. pes caprae, C. rosea, S. tomentosa, and S. terebinthifolius was significantly affected by the interaction between location and time (p ≤ 0.05, f test) This indicated that exposure to the source of emissions might influence the chemical constitution of the plants, either by modifying the soil properties, or by deposition of iron particulates on the leaf surface according to the temporal deposition pattern.
The accumulation of total Fe by all four studied species, especially in S. tomentosa, was greater on plants growing adjacent to the industry (1.0-km site) rather than on those growing at the farther site (15.0 km). A variation in the total Fe content was also observed during the year, but was less pronounced in S. tomentosa. For the other three species, higher Fe contents were generally found in the first semester which corresponded to the rainy period (Fig. 4).

Mean spatial and temporal changes of the total iron content in the leaves of Ipomoea pes caprae, Canavalia rosea, Sophora. Tomentosa, and Schinus. terebinthifolius (n = 3). Bars marked with different capital and lower case letters differ from each other on a temporal and spatial basis, respectively. Probability level p ≤ 0.05 (Tukey’s test)
Total Chlorophyll Content
The effect of time and location on chlorophyll content of the leaves of the four species studied was variable. The total chlorophyll content of I. pes caprae and S. tomentosa was significantly affected by the interaction between time and location (p ≤ 0.05, f test). The total chlorophyll content of C. rosea was only affected by the location where the plants were growing (p ≤ 0.05, f test). The total chlorophyll content of S. terebinthifolius was influenced both by the location and the time of analysis (p ≤ 0.05, f test).
The total chlorophyll content of I. pes-caprae, C. rosea, and S. terebinthifolius was generally higher on the 1.0-km site (Fig. 5), where the amount of Fe in the soil (Table 2) and the load of particulate matter on the leaves (Fig. 3) were higher than those observed at the 15.0-km site. S. tomentosa did not show a significant difference in chlorophyll content, except in March when the content was the lowest (Fig. 5b).

Mean spatial and temporal variation of total chlorophyll content (chla + chlb) in the leaves of Ipomoea pes caprae, Canavalia rosea, Sophora. Tomentosa, and Schinus. terebinthifolius (n = 5). (a, b) Bars marked with different capital and lower case letters differ from each other on a temporal and spatial basis, respectively. (c, d) Bars marked with different lower case letter differ from each other. Probability level p ≤ 0.05 (Tukey’s test)
The influence of time on the chlorophyll content was variable among the studied species along the course of the year. This variation may be linked to the pattern of production of new leaves.
Phenology Observation
The phenological monitoring of I. pes caprae, C. rosea, S. tomentosa, and S. terebinthifolius revealed that all phenophases were affected by the proximity to the emission source. Fluctuations in the intensity and pattern of the phenophases between plants at different distances from the emission source were observed (Fig. 6a–d).

(a, b) Phenology of Ipomoea pes caprae and Canavalia. rosea (n = 10) growing at 1.0 km and 15.0 km away from the iron ore industry plant at Anchieta restinga / ES. (c, d) Phenology of Sophora tomentosa and Schinus terebinthifolius (n = 10) growing at 1.0 km and 15.0 km away from the iron ore industry plant at Anchieta restinga / ES
The production of new leaves by all four species growing at the 1.0-km site tended to be lower than those plants growing at the 15.0-km site. Most of the shoot production in I. pes caprae and C. rosea occurred in the first semester (Fig. 6a, b), during the rainy period (days 5 up to 185). The maximum shoot production observed in I. pes caprae was around day 65 in plants at the 15.0-km site, while the plants growing at the 1.0-km site showed delayed shoot production, and this delay was nearly 120 days (Fig. 6a). In C. rosea, no obvious signs of a shooting delay were observed between the plants growing at the two sites; however, the intensity of the shoot production was lower for the plants growing near the industry (Fig. 6b). S. tomentosa and S. terebinthifolius had two peaks of shoot production in the year, one in the early rainy period (first semester) and the other in the early dry period (second semester) (Fig. 6c, d). The intensity of this phenophase in S. tomentosa was affected, although not severely. New leaves on S. tomentosa were primarily found in plants at the 1.0-km site. This was followed 30 days later by the plants at the 15.0-km site. In the second semester of the year, the opposite pattern was observed, with the plants at the 15.0-km site releasing new leaves prior to those plants growing at the 1.0-km site (Fig. 6c). Shoot production in S. terebinthifolius was simultaneously observed in plants at both sites in the early rainy period (first semester), while in the early dry period (second semester), the plants at the 15.0 km site were the first to release the new leaves (Fig. 6d).
The flowering pattern of I. pes caprae and C. rosea was similar for the plants growing at both sites. I. pes caprae had two peaks of flowering, the first and greatest occurred in the early rainy period (first semester). This was followed 180 days later (second semester) by a weaker flowering peak (Fig. 6a). Flowering in C. rosea occurred all year round at both sites, although the highest percentages were observed on two occasions in the first semester of the year (Fig. 6b). S. tometosa also showed a similar pattern of flowering, but flowering was greater in plants at the 1.0-km site (Fig. 6c). Plants of S. terebinthifolius growing at both sites flowered simultaneously at two distinctive periods. The first peak occurred around day 65 (first semester), while a second and weaker peak occurred around day 185 (second semester) (Fig. 6d).
The fruit production in all the species, except for I. pes caprae, was affected by the proximity of the plants to the source of pollutants (Fig. 6a–d). Fruit production in I. pes caprae plants growing at both sites was highest in the first semester, with its maximum in the early rainy period (day 35) (Fig. 6a). In contrast, C. rosea plants growing at both sites spread their fruit production throughout the year, peaking in the middle of the first semester (day 95) (Fig. 6b). Although S. tomentosa plants at both sites showed the highest levels of fruit production throughout the first semester, this event was less intense in plants growing at the 1.0-km site (25%), and started about 60 days later when compared to the plants growing at the 15.0-km site (Fig. 6c). The fruit production in S. terebinthifolius was also affected by proximity to the source of pollutants. In the middle of the first semester, the plants growing at the 1.0-km site reached their maximum of fruit production of 50%, this was in contrast with the 95% of the plants growing at the 15.0-km site that reached their maximal fruit production. A second peak of fruit production was observed at the end of the second semester, but it was less intense (Fig. 6d).
Discussion
Both wet and dry acid precipitation can alter soil pH and its mineral content. The pronounced dust load at the 1.0 km site confirms that this area is affected on a more regular basis than the 15.0-km site (Fig. 3). Modifications on soil properties are one of the consequences of intense exposure to atmospheric pollution upon ecosystems. Such effects have an important role on the chemical composition of plant tissues (Monni et al. 2001; Pugh et al. 2002). The nature of the iron ore particulate pollutants in this study, in addition to the acidification potential of the SO2 emitted by the industry, may be responsible for the differences in the soil composition of the studied sites (Table 2).
Regardless of the pollution, we found no indication that the amounts of iron found in the soils of 1.0- and 15.0-km sites exceed typical values (Azevedo and Chasin 2003). However, the lower pH of the soil at the 1.0-km site may enhance the solubility of essential micronutrients, including Fe (Engleman and McDiffett 1996; Connolly and Guerinot 2002; Momen et al. 2002). Increased nutrient availability may interfere in plant physiology and nutrient content, and if the nutrients do not reach toxic levels, then plant growth and development may be improved. The physical and chemical characteristics of sandy soils, such as those found at the studied sites, are often limiting factors for plant establishment and survival. Therefore, few species are able to overcome the harshness of the environment. Any permanent modifications on the soil properties may interfere with the plant composition of the ecosystem.
A higher deposition of TPM was observed in all four studied species in the second semester, which is the less pronounced rainy period (Fig. 3). Although, in September, the rainfall was unusually pronounced, therefore the higher deposition of TPM on all four species may be explained by the timing of sampling, which occurred before the precipitation event.
The amount of TPM deposited on leaves of S. tomentosa was generally higher than in other species. The presence of trichomes (data not measured) on both leaf surfaces of this species might have helped trap particulate matter, in contrast to glabrous species or those with fewer trichomes on their leaf surfaces (I. pes caprae, C. rosea, and S. terebinthifolius). During rainy periods, most of the particulate matter, either in suspension or deposited on surfaces, is usually washed out. The coarse fraction (2.5–10 μm∅) of the TPM is most affected by this meteorological component, while the fine fraction (< 2.5 μm∅) often stays on surfaces as a persistent layer. Depending on the leaf surface characteristics, more or less particles may adhere (Grantz et al. 2003). The size of the particulate matter also dictates much of their depositional fate, therefore the coarse fraction often precipitates at areas near the source. This is opposite to the observations for fine particles, which tend to be carried by the wind for greater distances (Farmer 1993).
Physical alterations of plants have been attributed to dust coarse particles (Hirano et al. 1995). This fraction of PM can abrade or smother the leaves, causing weakness, a rise in temperature, or impair light from reaching the photosynthetic tissues (Naidoo and Chirkoot 2004). The fine fraction of PM can affect plants by affecting stomatal apertures and if the PM penetrates within the mesophyll, it may modify the chemical balance of the mesophyll (Silva et al. 2006). Depending on the sensitivity of the individual species, the outcome may be either beneficial or detrimental.
In all four studied species, the Fe content was higher in the site near the source of pollutants (Fig. 4). In S. tomentosa, the higher content of total Fe occurred during the first semester (rainy period) and may be due to greater water availability, elevated temperature and irradiation. Together, these factors can increase plant growth rates and uptake of mineral nutrients by the roots. In addition, since TPM deposition in this species is higher, due to the presence of trichomes, previous accumulation of TPM could have led to an increased total Fe content in the leaves. Foliar absorption is also a possible route of nutrient intake, and this process may be aided by the combination of constant acid and particulate matter deposition onto leaves (Grantz et al. 2003). Similar conditions are found at the location of assessment in this study, thus increasing the possibility of higher Fe accumulation by the leaves of exposed species.
Fe is an essential micronutrient involved in chlorophyll synthesis and many metabolic reactions in plant tissues. Even though Fe content in the leaves was significantly affected by spatial and temporal components, the values found in I. pes caprae, C. rosea, and S. terebinthifolus never reached values considered to be phytotoxic (>500 ppm) (Levy et al. 1999). It is possible that these species are able to resist iron toxicity by avoiding over-accumulation of the mineral. However in S. tomentosa plants growing at the 1.0-km site, Fe content clearly exceeded phytotoxic levels, indicating that this particular species might not be able to regulate its nutrient uptake. Excess Fe in foliar tissue may elicit oxidative stress or enhance photo damage leading to a reduction in photosynthetic capacity (Sinha et al. 1997; Suh et al. 2002).
The plant response to toxicity caused by trace elements depends on the individual species sensitivity and environmental conditions (Narayan et al. 1994). Synergic pollution stress, such as acid deposition and overload of particulate matter, can collaborate to diminish the development and reproduction of susceptible species, while resistant species will thrive in a disturbed environment (Salemaa et al. 2001; Alvarez et al. 2003).
Near the source of emissions, the TPM deposition and chlorophyll content in all four studied species were higher (Figs. 3 and 5), despite the fact that the presence of heavy dust layers on leaves can block light and thus affect chlorophyll synthesis (Prusty et al. 2005). Although shading can also increase chlorophyll in an area, it is unlikely that the smothering caused by TPM deposition is the only factor promoting the increase in chlorophyll in the present study. That is because for shading to effectively promote a rise in chlorophyll content, it should be a constant factor. However, the deposition of particulate matter upon field conditions, such as the ones in the coastal regions is often disturbed by the wind.
Chlorophyll synthesis and content can also be affected by the age of the leaf and nutrient availability. The chlorophyll content in leaves is often negatively correlated to atmospheric pollution (Monni et al. 2001; Prusty et al. 2005), but increases can also be observed depending on the type of the pollutants or mineral excesses to which the plants are exposed (Päivöke and Simola 2001; Silva et al. 2006).
The higher Fe content in the leaves of all four species at the 1.0-km site clearly shows that Fe accumulates in larger quantities in the plants of this site. Since Fe is necessary for chlorophyll synthesis (Briat et al. 1995), these plants might be favored by the circumstances present at the 1.0-km site. The increased Fe present in S. tomentosa at the 1.0-km site might be partially stored as chelates in the apoplastic space (Connolly and Guerinot 2002; Bartakova et al. 2001). This may prevent the onset of phytotoxic stress and avoid cellular damage. However, the toxic levels of Fe in S. tomentosa leaves could generate oxygen reactive intermediates at some point, which may then affect plant productivity (Becana et al. 1998). Even though the Fe content in the leaves of I. pes caprae, C. rosea, and S. terebinthifolius was higher at the 1.0-km site, toxic levels were never observed. Therefore, this component is probably not associated with trace metal toxicity. Since the increase in chlorophyll content of all studied species was favored by proximity to the source of pollutant, this component was probably not a limiting factor for the observed decreases in the phenophases of the four species assessed.
The observed temporal influences on chlorophyll content (Fig. 5) in all four studied species are probably related to the phenological component of the species. This is driven by seasonal variation and self-regulation of the plants (Van Schaick et al. 1993). During hot rainy periods, tropical plants often invest in the outburst production of new leaves, while senescence is synchronized with periods of low rainfall (Van Schaick et al. 1993). In the cases of I. pes caprae, C. rosea, and S. terebinthifolius, leaf production occurred throughout the year, although in an uneven pattern (Fig. 6a, b, d). This strategy ensures that new and mature leaves are always present, and sampling of leaves can be performed with low variation along the whole year. S. tomentosa invested in the production of new leaves in two different periods of the year. The lower content in chlorophyll in the plants at the 15.0-km site in March 2005 is probably due to the age of the leaves that are not yet mature at the sampling time (Fig. 5b). The sampling time almost overlapped the summer outburst of leaves in these species (Fig. 6c).
The setting of new leaves, flowers, and fruits relies on a series of factors, including the ones mentioned above that were analyzed in this study. Organic and mineral nutrition and water availability are major requirements for the investment in new organs and reproductive structures. Under exposure to severe pollutants, plant growth and productivity can be reduced (Momen et al. 2002; Brun et al. 2003; Chauhan et al. 2004). The greater amount of particulate matter deposited on the plants of all studied species growing at the 1.0-km site might decrease their overall production. Iron ore dust may contribute by overheating or smothering the leaves (Naidoo and Chikoot 2004; Silva et al. 2006).
Fruit production at the 1.0-km site was the phenological aspect most affected in two out of the four species assessed, S. tomentosa, and S. terebinthifolius. Since the chlorophyll content and the iron levels in the soil were not limiting factors, the decrease in flowering and fruiting are probably linked to other ecological factors. Nonetheless, in S. tomentosa, toxic iron accumulation may have interfered in reproductive success, since iron excess may trigger oxidative stress (Becana et al. 1998). The formation of seeds and fruits generally depend on pollination and fertilization processes. Unfit environmental conditions due to pollution can affect the behavior of pollinators, as well as the fertilization process, thus contributing to a decline in fruit production (Paoletti and Bellani 1990; Kevan 1999; Iannotti et al. 2000; Gottardini et al. 2004). As a result of poor fruit production, natural germplasm resources can be depleted, causing changes in the vegetational structure and dynamics of the ecosystem (Brun et al. 2003; Wen et al. 2006).
Understanding how and which restinga plant species respond to ore industry emissions may provide new approaches to dealing with issues concerning ecosystem conservation and restoration.
Conclusion
Even though the effects differed among the species, the overall outcome was that the majority of plants growing in the vicinity of the source of emissions were affected. Although phenological monitoring was only performed over one year, it provided evidence that the assessed species may be under stress. Since fruit production was diminished in the plants near the pollution source, the heavy load of TPM observed at the 1.0-km site might contribute to subtle changes in the reproductive effort of all four species studied. In the long term, it is possible that iron ore industry emissions will alter the structure and diversity of the exposed vegetation. To avoid bias, this conclusion was not based solely on one variable.
Different ecophysiological parameters and a unified view of the data were used to evaluate the impact caused by the ore industry on restinga vegetation. Adverse effects were pronounced in S. tomentosa, which accumulated Fe to toxic levels and exhibited a decreased reproductive effort. This species may also be used as an indicator organism, and may help to assess the ecological risks due to particulate matter emitted by iron ore industry activities.
References
Alvarez E, Fernández Marcos ML, Vaamonde C, Fernández-Sanjurjo MJ (2003) Heavy metals in the dump of an abandoned mine in Galicia (NW Spain) and in the spontaneously occurring vegetation. Science of Total Environment 313:185–197
Azevedo FA, Chasin AAM (eds) (2003) Metais: gerenciamento da toxicidade. Atheneu, São Paulo, pp 554
Barrick KA (2003) Comparison of the nutrient ecology of coastal Banksia grandis elfin wood (windswept shrub-like from) and low trees, Cape Leeuwin-Natural site National Park, Western Australia. Austral Ecology 28:252–262
Bartakova I, Kummerova M, Mandl M, Pospisil M (2001) Phytotoxicity of iron in relation to its solubility conditions and the effect of ionic strength. Plant and Soil 235:45–51
Becana M, Moran JF, Inturbe-Ormaetxe I (1998) Iron-dependent oxygen free radical generation in plants subjected to environmental stress: toxicity and antioxidant protection. Plant and Soil 201:137–147
Briat JF, Fobis-Loisy I, Grignon N (1995) Cellular and molecular aspects of iron metabolism in plants. Cell Biology 84:69–81
Brun LA, Le Corff J, Maillet J (2003) Effects of elevated soil copper on phenology, growth and reproduction of five ruderal plant species. Environmental Pollution 122:361–368
Chauhan SV, Chaurasia B, Rana A (2004) Impact of air pollution on floral morphology of Cassia siamea Lamk. Journal of Environmental Biology 25:291–297
Connolly EL, Guerinot ML (2002) Iron stress in plants. Genome Biology 3:10241–10245
Dale VH, Beyeler SC (2001) Challenges in the development and use of ecological indicators. Ecological Indicators 1:3–10
Engleman CJ Jr, McDiffet WF (1996) Accumulation of aluminum and iron by bryophytes in streams affected by acid – mine drainage. Environmental Pollution 94:67–74
Farmer A (1993) The effects of dust on vegetation: a review. Environmental Pollution 79:63–75
Fournier LAO (1974) Um método cuantitativo para la medición de característica fenológica em árboles. Turrialba 24:422–423
Gallego SM, Benavides PM, Tomaro ML (1996) Effect of heavy metal ion excess on sunflower leaves: evidence for involvement of oxidative stress. Plant Science 121:151–159
Gottardini E, Cristofolini F, Paoletti E, Lazzeri P, Pepponi G (2004) Pollen viability for air pollution bio-monitoring. Journal of Atmospheric Chemistry 49:149–159
Grantz DA, Garner JHB, Johnson DW (2003) Ecological effects of particulate matter. Environment International 29:213–239
Gross CL (1993) The reproductive ecology of Canavalia rosea (Fabaceae) on Anak Krakatau, Indonesia. Australian Journal of Botany 41:591–599
Hirano T, Kiyota M, Aiga I (1995) Physical effects of dust on leaf physiology of cucumber and kidney bean plants. Environmental Pollution 89:255–261
Iannotti O, Mincigrucci G, Bricchi E, Frenguelli G (2000) Pollen viability as a bio-monitor of air quality. Aerobiology 16:361–365
Kampfenkel K, Van Montagu M, Inze D (1995) Effects of iron excess on Nicotiana plumbaginifolia plants. Plant Physiology 107:725–735
Kevan PG (1999) Pollinators as bioindicators of the state of the environment: species, activity and diversity. Agriculture Ecosystems and Environment 74:373–393
Kozlov MV, Zverva EL (2004) Reproduction of mountain birch along a strong pollution gradient near Monchegorsk, Northwestern Russia. Environmental Pollution 132:443–451
Lenzi M, Orth AI (2004) Caracterização functional do sistema reprodutivo de aroeira vermelha (Schinus terebinthifolius Raddi), em Florianópolis-SC, Brasil. Revista Brasileira de Fruticultura 26:198–201
Levy DB, Redente EF, Uphoff GD (1999) Evaluating the phytotoxicity of Pb-Zn tailing to big bluestem (Andropogon gerardii Vitman) and switchgrass (Panicum virgatum L.). Soil Science 164:363–375
Lopes SA, Oliva MA, Martinez CA (2000) Impacto das imissões de dióxido de enxofre e deposição de material particulado de ferro em espécies vegetais de restinga: avaliação ecofisiológica. In: Espíndola E, Paschoal C, Rocha O, Bohrer M, Oliveira Neto A (eds) Ecotoxicologia. RiMa, São Paulo, pp 53–71
McCune DC (1991) Effects of airbone saline particle on vegetation in relation to variables of exposure and other factors. Environmental Pollution 74:176–203
Momen B, Anderson PD, Houpi JLJ, Helms JA (2002) Growth of ponderosa pine seedlings as affected by air pollution. Atmospheric Environment 36:1875–1882
Monni S, Uhling C, Hansen E, Magel E (2001) Ecophysiological responses of nigrum to heavy metal pollution. Environmental Pollution 112:121–129
Naidoo G, Chirkoot D (2004) The effects of coal dust on photosynthetic performance of the mangrove, Avicennia marina in Richards Bay, South Africa. Environmental Pollution 127:359–366
Narayan D, Agrawal M, Pandey J, Singh J (1994) Changes in vegetation characteristics downwind of an aluminum factory in India. Annals of Botany 73:557–565
Nogueira EML, Arruda VLV (2006) Fenologia reprodutiva, polinização e sistema reprodutivo de Sophora tomentosa L. (Leguminosae – Papilionoideae) em restinga da praia da Joaquina, Florianópolis, sul do Brasil. Biotemas 19:29–36
Päivöke AEA, Simola LK (2001) Arsenate toxicity to Pisum sativum: mineral nutrients, chlorophyll content, and phytase activity. Ecotoxicology and Environmental Safety 49:111–121
Paling EI, Humphries G, McCardle I, Thomson G (2001) The effects of iron ore dust on mangroves in Western Australia: lack evidence for stomatal damage. Wetland Ecology Management 9:363–370
Paoletti E, Bellani LM (1990) The in vitro response of pollen germination and tube length to different types of acidity. Environmental Pollution 67:279–286
Patiño S, Jeffree C, Grace J (2002) The ecological role of orientation in tropical convolvulaceous flowers. Oecologia 130:373–379
Pereira OJ (1990) Caracterizações fitofisionômica da restinga de Setiba, Guarapari - ES. In: Simpósio sobre ecossistemas da costa sul e sudeste brasileira, III. Anais, São Paulo, ACIESP, pp 207–219
Prusty BAK, Mishra PC, Azzez PA (2005) Dust accumulation and leaf pigment content in vegetation near the national high way at Sambalpur, Orissa, India. Ecotoxicology and Environmental Safety 60:228–235
Pugh RE, Dick DG, Fredeen AL (2002) Heavy metal (Pb, Zn, Cd, Fe, and Cu) contents of plant foliage near the Anvil Range lead/zinc mine, Faro, Yukon Territory. Ecotoxicology and Environmental Safety 52:273–279
Resoluções CONAMA (1992) Resoluções CONAMA n° 003/90, 4th edn. Brasília
Salemaa M, Vanha-Majamaa I, Derome J (2001) Understory vegetation along a heavy metal pollution gradient in SW Finland. Environmental Pollution 112:339–350
Silva LCD, Oliva MA, Azevedo AA, Araújo JM (2006) Responses of restinga plant species to pollution from an iron pelletization factory. Water, Air and Soil Pollution 17:241–256
Sinha S, Gupta M, Chandra P (1997) Stress induced by iron in Hydrella verticillata, (I. F.) Royele: response of antioxidants. Ecotoxicology and Environmental Safety 38:286–291
Suh HJ, Kim CS, Lee JY, Jung J (2002) Photodynamic effect of iron excess on photosystem II function in pea plants. Photochemistry and Photobiology 75:513–518
Uhling C, Juntilla O (2001) Airborne heavy metal pollution and its effects on foliar elemental composition of Empetrum hermaphroditum and Vaccinium myrtillus in Sor-Varanger, northern Norway. Environmental Pollution 114:461–469
Van Schaick CP, Terbogh JW, Wright SJ (1993) The phenology of tropical forests: adaptive significance and consequences for primary consumers. Annual Review of Ecology and Systematics 24:353–377
Wellburn AR (1994) The spectral determination of chlorophyll a and b, as well as total carotenoids, using various solvent with spectrophotometers of different resolutions. Journal of Plant Physiology 144:307–313
Wen D, Kuang Y, Liu S, Zhang D, Lu Y, Li J (2006) Evidences and implications of vegetation damage from ceramic industrial emission on a rural site in the Pearl River delta of China. Journal of Forestry Research 17:7–12
Wong MH, Chan KC, Choy CK (1978) The effect of the iron ore tailing on the costal environment of Tolo harbor, Hong Kong. Environmental Research 15:342–356
Yunus M, Iqbal M (eds) (1996) Plant response to air pollution. John Wiley & Sons, New York, pp 447
Acknowledgments
This research was made possible by Universidade Federal de Viçosa (UFV), Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), Fundação de Amparo à Pesquisa do Estado de Minas Gerais (FAPEMIG), and SAMARCO Mineração S.A.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Kuki, K.N., Oliva, M.A. & Pereira, E.G. Iron Ore Industry Emissions as a Potential Ecological Risk Factor for Tropical Coastal Vegetation. Environmental Management 42, 111–121 (2008). https://doi.org/10.1007/s00267-008-9093-7
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00267-008-9093-7