
Abstract
This study recorded and analyzed cumulative vertebrate roadkill data on 107 km of roads in rural South Korea and reports the first intensive roadkill study results from Asia. Over 30 months, roadkill strikes and adjacent landscape factors were recorded daily or every other day on three types of roads: highway, local, and riverside. For analysis, roads were segmented into 250-m units, and roadkill hotspot segments were calculated with the Getis–Ord Gi* statistic. Eighteen road characteristics and landscape factors were correlated to roadkill hotspots using multiple logistic regression analysis. Mammal mortality increased from spring to fall, whereas bird mortality peaked in summer. Reptile and amphibian mortality peaked during fall. Seasonal focal species’ mortality tracked the taxonomic groups they represent, except oriental scops owl (Otus scops), for which yearly mortality peaked 2 months earlier than for most birds. Mammal and bird roadkill hotspots were concentrated on the highway, which passes across mountains, whereas amphibian and reptile hotspots were on the riverside road because of movements related to breeding, juvenile dispersal, and hibernation. However, many species used the spatially complex agricultural fields along the local road, especially during harvest season. The significant site and landscape factors that influenced overall roadkill hotspots were a high landscape percentage of water and rice paddies, low traffic volumes, high percentage of natural vegetation, an absence of road banking, high roadside grass presence, and an absence of drainage. South Korea has an active wildlife-crossing management program, and these findings can inform avoidance, minimization, and mitigation strategies and practices.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Despite economic, quality of life (QOL), and other benefits of roads to humans, there is widespread recognition of the adverse impacts of roads on natural environments (National Research Council 2005; Forman et al. 2003; Trombulak and Frissell 2000). These impacts include direct and cumulative mortality to species hit by vehicles along roads (Forman et al. 2003; Riley et al. 2006; Grilo et al. 2009), reduced dispersal capacity (Forman and Alexander 1998) and associated impediments to gene flow (Epps et al. 2005; Riley et al. 2006; Balkenhol and Waits 2009), increased spread of invasive species (Gelbard and Belnap 2003), disturbance effects such as traffic noise that can affect breeding birds (Reijnen and Foppen 1995; Reijnen et al. 1995), impedance to recolonization of depopulated habitats, and indirect effects including but not limited to the road-effect zone (Bissonette and Rosa 2009), landscape fragmentation (Jaeger et al. 2005; Girvetz et al. 2008) and greenhouse gas emissions (Fuglestvedt et al. 2008).
Roadkill studies have focused on a wide range of vertebrate species (Finder et al. 1999; Nielsen et al. 2003; Glista et al. 2007; Gunson et al. 2009; Grilo et al. 2009; Gomes et al. 2009; Langen et al. 2009). However, few studies have examined multiple taxonomic vertebrate groups across time periods long enough to determine predictive landscape factors by taxonomic group or season (e.g., Farmer and Brooks 2012).
The tremendous expansion of roads associated with human population growth increases the need to understand their impact on ecosystems and wildlife. Recognition of this need has led to the development of the subdiscipline of road ecology, recognized by textbooks (Forman 1995; Turner et al. 2001; Havlick 2002; Forman et al. 2003), conferences (Forman et al. 1997; West 2006) and >20 years of publications (Harris and Scheck 1991; Crist and Wilmer 2002; Johnson et al. 2002; Smith et al. 2005). Areas of research interest include developing landscape-scale metrics of fragmentation due to roads (Jaeger et al. 2005; Li et al. 2010), landscape-connectivity studies (Alexander and Waters 2000; Donaldson 2007; Crooks et al. 2008; Beier et al. 2008), and predictive assessments of the impacts of new roads (Thorne et al. 2009). Much of this research is meant to support actions to reduce the negative effects of roads through infrastructure alterations such as wildlife-crossing structures, fencing, decommissioning of roads, and optimizing the alignment of new roads (Fahrig and Rytwinski 2009; Gunson et al. 2011). Efforts that monitor roadkill of multiple taxa can provide information to transportation planners on where to place modifications to roads in order to minimize roadkill (e.g., Clevenger et al. 2003).
This study analyzed roadkill-monitoring data from three types of roads in southern South Korea and is the first to report results of multiple roadkill taxonomic groups and species from eastern Asia. The study investigated three questions: What are the rates of roadkill for different taxonomic groups and focal species on the Korean peninsula, and does this mortality vary by month? What are the spatial and seasonal patterns of roadkill hotspots for the four different taxonomic groups (mammals, birds, amphibians, and reptiles) and their representative focal species? What landscape and site-level factors contribute to roadkill hotspots?
Materials and methods
Study area
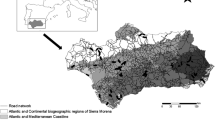
To address these questions, we monitored roadkill strikes for 30 months from 1 July 2004 to 10 January 2007 on a 107-km stretch of road in a rural region of southern South Korea comprising three types of construction. The road studied forms a semicircle around the northern and western sides of Jirisan National Park (JNP) and is a barrier between the park and mountainous areas further west. The study road passes through natural and seminatural landscapes (Fig. 1).

Study area
There are three types of roads in the study area: (1) A two-lane, undivided express highway (highway 12, herein called the highway), 44 km in length with an 80-km/h speed limit and in mountainous terrain. This road is located north of JNP and passes through forest and agriculture lands. It traverses the Baekdu Daegan, the central mountain range of South Korea, which is the main ecocorridor for wildlife movement in Korea. (2) National road 19 is the second type (herein called the local road). It is a four-lane divided national road 30 km long with an 80-km/h speed limit. It passes through hilly terrain and agricultural lands including rice paddies and vegetable and orchard crops interspersed with hedgerows and small water courses in a wide valley west of JNP. Adjacent hill slopes have limited tree cover and some shrublands. Where the road passes through these hills, it was created using a cut-and-fill technique. (3) The third is another section of national road 19 comprising a two-lane, undivided road 33 km long, with a 60-km/h speed limit. It runs parallel to the Sumjin River (herein called the riverside road). The riverside road passes through stands of mature upland trees and riparian forest, through some settlements, and follows the natural topography about 20 m above the river.
Data collection
Roadkill surveys were conducted daily from 1 July 2004 to 30 June 2005 and every other day from 1 July 2005 to 10 January 2007, for a total of 559 road surveys (Choi 2007). After 1 year of daily surveys, it was determined through statistical analysis that all roadkill strikes could be uniquely identified using every-other-day surveys (Bager and Rosa 2011). Corpses were removed after counting, and the strike location was spray painted to avoid duplicate counts. At each observed roadkill site, we recorded location, species, and date, as well as 18 different landscape and site variables such as fencing, road dividers, drainage culverts, and roadside vegetation (Table 1). Data were collected on four taxonomic groups: mammals, birds, amphibians, and reptiles. Each taxonomic group was further represented by an endangered species list, which was independently analyzed: leopard cat (Prionailurus bengalensis), oriental scops owl (Otus scops), Asiatic toad (Bufo bufo gargarizans), and red-sided water snake (Rhabdophis tigrinus).
Summary of roadkill statistics
We summed annual, monthly, and seasonal mortality by taxonomic group and focal species. We examined rates of roadkill for the three types of roads and assessed temporal variations for taxonomic groups and focal species.
Roadkill hotspot analysis
We expected roadkill hotspots to be nonrandom and to have a high spatial association with neighborhood type. It was therefore necessary to use local measures to depict the variability of spatial autocorrelation (Anselin 1995; Ord and Getis 1995). Using the Getis–Ord Gi* statistic (Ord and Getis 1995; Baruch-Mordo et al. 2008), we identified clusters of roadkill of higher magnitude than expected by random chance. To do this, we used road segments as units of analysis for all tests and described roadkill data, roadside variables, and landscape variables within each segment. We divided the road into 429 segments of 250 m in length and measured Gi for each road segment. The delineation of 250 m resulted in an average number of roadkill strikes of 12 per segment overall and of four per segment for focal species. Road segments shorter or longer than 250 m were also examined, but they either reduced or increased the number of roadkill occurrences in each segment to the point where we could not identify hotspot patterns. We used the fixed distance option (500 m; two road segments) for calculating hotspots and set a Z value of 2 from Gi scores to identify hotspot segments (Anselin 1995). Spatial and seasonal patterns of roadkill hotspots were examined for each taxonomic group and focal species. We used ArcGIS 9.3 for hotspot analysis (ESRI Inc. 2009).
Driving factors for roadkill
We examined 18 predictor variables in a multiple logistic regression framework to identify roadside and landscape factors consistently present at roadkill hotspots (Table 1). Thirteen variables were collected at each roadkill reported. Five landscape variables, consisting of general landcover categories, were sampled using a 500-m buffer from the road center line, and the percentage of each landcover type was summarized for every 250 m of road segment. The 500-m buffer distance was selected after a sensitivity analysis on a range of buffer sizes because it best captured the distribution of landcover types in our study area. The pattern of roadkill hotspots from the previous analysis (above) identified each road segment as having (1) or not (0) a hotspot. This pattern was used as the response variable. Logistic regression was selected because we used continuous and categorical data for explanatory variables (Press and Wilson 1978). The 18 predictor variables were quantified for each road segment. We analyzed roadkill hotspots for the four taxonomic groups and focal species.
All roadkill hotspot segments were used, but only twice as many randomly selected nonroadkill hotspot segments were included (rather than all nonhotspot locations) (Kvamme 1985). We used two analyses to evaluate each model. First, we used AIC to assess goodness of fit. Second, to assess model accuracy, we used three procedures: area under the relative operating characteristic curve (AUC, Hanley and McNeil 1982), Cohen’s K (Monserud and Leemans 1992), and true skill statistics (TSS, Allouche et al. 2006). Used in combination, these three procedures can provide more ways to assess the accuracy of each model. We therefore provide all three results. We assessed the relative importance of each predictor variable within each model by comparing Pearson correlation between standard predictions and predictions with the randomly permutated investigation variable. We also validated each model using 100 bootstrapping runs of a cross validation with a 70 % training set and a 30 % test set. We used BIOMOD in R 2.13.2 for all analyses (Thuiller et al. 2009).
Results
Summary statistics by taxonomic group, season, and road type
Road surveys totaled 59,813 km (559 surveys), and recorded 5,715 fatal road strikes. We used 5,044 records, and excluded 145 recorded strikes of invertebrates including crustaceans and millipedes and 526 locations with GPS error above 10 m from a roadside. Roadkill yearly totals were 2,532 (July 1, 2004–June 30, 2005), 1,655 (July 1, 2005–June 30, 2006), and 857 (July 1, 2006–January 10, 2007) (Table 2). A total of 5,044 vertebrates were killed during the study, comprising 1,582 mammals (1.71/day), 1,205 birds (1.3/day), 1,394 amphibians (1.5/day), and 863 reptiles (0.93/day) (Table 2). Roadkill species richness totaled 101 vertebrates, comprising 22 mammals, 59 birds, 6 amphibians, and 14 reptile species (Supplementary Appendix 1). Among these are 15 which have both national monument status (species designated by the Cultural Heritage Administration of South Korea for their significance), and endangered species status in Korea.
Unspecified rodents were the most frequently struck mammals, followed by the focal species raccoon dog (Nyctereutes procyonoides) (86.7/year) and water deer (Hydropotes inermis) (62.5/year). Mortality for the focal species leopard cat was 37.6/year (see all impacts in Supplementary Appendix 1). Mammal mortality increased in the spring and fall and peaked in October (Fig. 2). Maximum bird strikes were on ring-necked pheasant (Phasianus colchicus) (138.1/year), with 37.2 strikes/year on the focal species, oriental scops owl. Bird mortality peaked in summer. Reptile mortality peaked in the fall. Among reptiles, snakes were most affected; the focal species, red-sided water snake, was hit 116.3 times/year. The Asiatic toad had the dubious distinction of most frequently killed species, with 523.9 strikes/year. Amphibian mortality peaked during fall, and there were 3 months with peak strikes: March, June, and October (Fig. 2). All taxonomic groups had minimal mortality during the winter due to hibernation and inactivity.

Cumulative monthly roadkill for four taxonomic groups and for focal species; results of 2.5 years of monitoring (1 July 2004–10 January 2007). a Mammal, bird, amphibian, reptile. b–a Leopard cat (Prionailurus bengalensis), oriental scops owl (Otus scops), b–b Asiatic toad (Bufo bufo gargarizans) and red-sided water snake (Rhabdophis tigrinus)
Roadkill hotspot analysis
The distribution of yearly roadkill strikes across the study area for all species and taxonomic groups was nonrandom. For all species combined for the entire year, six roadkill hotspots were identified with a mean length of 1.2 km and a Gi Z statistic value > 2 (Fig. 3a). However, mammals and birds each had eight yearly hotspots, with an average 0.8-km kill zone for mammals and 1.2-km zone for birds. There were similar numbers of hotspots for amphibians (6) and reptiles (7), but these were longer kill zones, averaging 1.7 km for each taxonomic group (Fig. 3b).

Roadkill hotspots of four taxonomic groups and four seasons. a Total, b–a mammal, b–b bird, b–c amphibian, b–d reptile, c–a spring, c–b summer, c–c fall, c–d winter. Grey segment means less aggregated.; black segments show where roadkill hotspots are more spatially clustered (Z ≥ 2); grey segments are less clustered (1 ≤ Z ≤ 2)
The number and location of all-taxa hotspots varied by season, with fall having the most (11, averaging 727 m in hotspot length), followed by summer (7 hotspots, averaging 500 m). Spring and winter had six and five hotspots, respectively, but they were longer (1,000 and 900 m, respectively) than summer hotspots (Fig. 3c). There were nine yearly roadkill hotspots for leopard cat, with a mean road length of 611 m. This number doubled in the summer to 16 (Fig. 4a), and the hotspot length for this species was longest in spring, at 844 m (n = 8), and shortest in winter, at 406 m (n = 8). Mean roadkill hotspot length (875 m, n = 7) for oriental scops owl (Fig. 4b) was shorter than for all birds, and the number of hotspots was constant (10) in spring, summer, and fall; there was no mortality in winter. Hotspots lengths for this species varied slightly by season through spring, summer, and fall, at 450, 600, 800 m, respectively. For the Asiatic toad (Fig. 4c), the mean roadkill hotspot length (786 m, n = 7) was shorter than for all amphibians combined; the number of hotspots in spring and fall was similar (6 and 7, respectively) but was halved in summer, likely due to seasonal movement because Asiatic toads move from mountain to river habitats in spring for breeding and in the opposite direction in fall for hibernation. The length of hotspots for this species was longest in fall (1,036 m). Mean hotspot length (615 m, n = 13) for the red-sided water snake (Fig. 4d) was shorter than for all reptiles combined and was longest in spring (958 m); from spring to fall, hotspot length was 11 and 13 m, respectively, with none in winter due to hibernation.

Seasonal roadkill hotspots of four focal species. a Leopard cat (Prionailurus bengalensis), b oriental scops owl (Otus scops), c Asiatic toad (Bufo bufo gargarizans), and d red-sided water snake (Rhabdophis tigrinus). Columns: a total, b spring, c summer, d fall, e winter. Black segments roadkill hotspots are more spatially clustered (Z ≥ 2); grey segments are less clustered (1 ≤ Z ≤ 2)
The spatial pattern of hotspots varied by season and by taxonomic group and focal species (Figs. 3, 4), with hotspots most prevalent along the riverside road due to the many roadkill occurrences of amphibians and reptiles. However, spatial distributions of mammal and bird fatalities were concentrated on the mountainous highway. Roadkill occurrences on the local road were few compared with highway and riverside roads. Across the year, roadkill hotspots for the leopard cat occurred primarily on the local road and were widespread in summer but more spatially clustered in winter; occurrences were concentrated on the local road in fall. Roadkill hotspots of oriental scops owl were concentrated on the highway; those of the Asiatic toad were concentrated on the riverside road with more hotspots found in spring and fall. Hotspots of the red-sided water snake mostly concentrated on the highway and riverside road across the entire year but were widespread in spring.
Driving factors for roadkill
Statistically significant site and landscape factors that influence overall roadkill hotspots were a high landscape percentage of water, low traffic volumes, a high percentage of natural vegetation, a high percentage of rice paddies, an absence of road banking, high roadside grass presence, and an absence of drainage. For mammals, rank-ordered significant factors that increased the risk of roadkill were being far from water, high roadside tree presence, low traffic volumes, less mowing, and low road slopes. A high percentage of rice paddies, median barrier presence, and presence of road banking were the most important roadkill factors for leopard cat. Locations with fewer traffic lanes, high vehicle speed, median barrier presence, high numbers of roadside trees or shrubs in forests and mixed forest–open areas correlated with a high risk of roadkill for birds. Roadkill occurrences of oriental scops owl were correlated with locations where roads crossed lands with less vegetation or rice paddies that had fewer traffic lanes and were close to villages. Amphibian roadkill occurrences were associated with roads near streams, vegetation along the roadside, locations without road banking, near to rice paddies, and areas with low traffic volume. Roadkill of Asiatic toad occurred at similar locations. Reptile roadkill occurrences were correlated with roads having low vehicle speeds and roadside shrubs that were nearby to forest and agriculture land. Roadkill occurrences of the red-sided water snake were correlated with roads with low traffic volume, far from villages, and without road banking (Table 3).
The final models for taxonomic groups and focal species were built using a multiple logistic regression model with AIC, and model performance was calculated with AUC, K statistics, and TSS (Table 3): overall values for total roadkill hotspots were 0.868, 0.715, and 0.733, respectively, and were highest for amphibians (0.872, 0.730, 0.787, respectively), followed by reptiles (0.833, 0.651, 0.662, respectively), mammals (0.798, 0.545, 0.598, respectively), and birds (0.784, 0.551, 0.579, respectively). The Asiatic toad (0.849, 0.846, 0.742, respectively) had the highest value among focal species. The leopard cat (0.784, 0.596, 0.590, respectively) had similar values to those of the Asiatic toad, followed by the red-sided water snake (0.746, 0.481, 0.499, respectively) and oriental scops owl (0.726, 0.491, 0.492, respectively).
Discussion
Spatial patterns of roadkill varied according to taxonomic group, seasonal pattern, road type, and landscape and site factors. We found the majority of mammals and birds were killed on the highway, which passes across a forested mountain range where these groups are residents and have to move for breeding and migration. On the other hand, amphibian and reptile mortality was higher along the riverside road because of breeding and juvenile dispersal near water (Langen et al. 2009). Whereas some of these patterns are similar to what might be expected in other countries, the magnitude of mortality is higher and the spatial patterns are driven by landscape heterogeneity in South Korea. Measuring the patterns and intensity of mortality hotspots permitted the modeling of their driving factors. The resulting information can be used for making more general models of expected mortality hotspots for various species or taxonomic groups and as a general set of guidelines that could be used in developing mitigation practices to avoid roadkill.
Road characteristics and roadkill
Road density for highway, national, and local roads in South Korea is a little higher than in the USA (1.05 vs. 0.68 km/km2) and much lower than in Japan (3.18 km/km2). However, road length per vehicle (5.88 km/1,000 vehicle) is much lower than in the USA (26.45 km/1,000 vehicle) and Japan (15.85 km/1,000 vehicle) (Statistics Korea 2011, IRF 2010). This means that the probability of roadkill is higher in South Korea compared to other countries because of the higher traffic volume. This high traffic volume is a possible contributor to the high rates of roadkill we recorded (Table 2). Amphibians and reptiles drove the high rates along the river. Roadkill rate on the local road was smallest because four-lane roads in rural areas with high traffic volumes may present greater hurdles and thus fewer animals would attempt to cross them; two-lane roads in forests and along the river have a higher incidence of collisions with animals (Hughes et al. 1996). These rates are much higher than those reported in other, roughly comparable, studies we examined, which range from 1.25 to 5.29/km/year for all vertebrates (Clevenger et al. 2003; Taylor and Goldingay 2004), and 0.47/km/year for mammalian mesocarnivores in a Mediterranean system (Grilo et al. 2009) compared with 1.47/km/year for the mesocarnivores in this study. Possible reasons for the high numbers in this study (in addition to traffic volume) include proximity to a national park; animal abundance levels are higher in South Korea than in locations used for other studies; and other human activities in our study region have relatively low impact on the species studied, leading to a greater abundance of animals.
Characterization of roadkill hotspots
Seasonal patterns of vertebrate roadkill were evident for all four taxonomic groups. These patterns are consistent with species’ life history, particularly breeding and juvenile dispersal (Davies et al. 1987; Joyce and Mahoney 2001; Clevenger et al. 2003; Conard and Gipson 2006). For example, the leopard cat is more active in summer, and its roadkill hotspots were widely spread at that time; however, occurrences became concentrated and reached their highest levels on the local road in the fall. This road traverses agricultural lands, which comprise a dense network of small rice paddies and vegetable plots, footpaths, and extensive hedgerows or natural vegetation in patches among a large number of small human-constructed waterways. Increased roadkill for this species is possibly due to their prey species being found on these farmlands during harvest time (Rajaratnam et al. 2007).
Bird mortality peaked in July. The overall pattern is likely driven by juvenile dispersal of ring-necked pheasants (Giudice and Ratti 2001) and seasonal migration of other species. Roadkill of oriental scops owl, the bird focal species, peaked in June and continued in summer and fall due to summer migration (Marshall 1978); most occurrences were on the highway, which is near the forest habitat it requires and that essentially forms a barrier from east to west, across the north/south migration routes for this species (Figs. 4b, 5).

Congruence map summing roadkill hotspots of the four taxonomic groups and four wildlife crossings, which were constructed by local governments as a result of this research
Reptile mortality peaked in the fall, potentially due to seasonal migrations and juvenile dispersal (Brito 2003). Roadkill hotspots of the focal species red-sided water snake were concentrated on the riverside road across the entire year but also occurred on the highway in the fall. Hotspots were widespread in spring because these snakes are active after hibernation, when they migrate downhill on mountain slopes; possibly, fall roadkill on the highway represented a return to nesting sites for hibernation (Fig. 4d).
The tripeaked mortalities for amphibians in March, June, and October (Fig. 2) could be due to seasonal migrations of the Asiatic toad for reproduction, juvenile dispersion, and hibernation (Schmetterling and Young 2008); roadkill was nearly exclusive to the riverside road because Asiatic toads have to move to water for breeding.
AUC values of three taxonomic groups were >0.784. This showed that roadkill hotspots of these groups were well detected, and the predictor variables for roadside and landscape factors derived from models described their roadkill characteristics reasonably well. AUC values of three focal species were >0.726 but lower than their respective taxonomic groups, suggesting slightly lower confidence in the results. Roadkill hotspots of the oriental scops owl were found on roads along mountains near villages, but these were not strongly associated with specific predictor variables. Further studies on the behavioral characteristics of target species and carrying capacity of the land for different species are needed to enhance the low AUC of some target species.
Implications for roadkill mitigation
The importance of reducing the impact of roads on wildlife is well recognized in South Korea. As part of the National Ecological Network Plan (Ministry of Environment 2007), >317 wildlife crossing structures have been built or retrofitted since 1998, including 147 overpasses and 170 underpasses. These crossings were built by three agencies: 153 crossings on the national roads by the Ministry of Construction and Transportation, 119 on local roads by local governments, and 45 on the highways by the Korea Expressway Corporation (Ministry of Environment 2011).
This study shows that ongoing roadkill mitigation efforts can anticipate seasonal patterns of animal movement as they progress through their yearly cycle. These movements are dictated by the Korean Peninsula’s strong seasonal weather patterns. For example, considerations about where to place additional wildlife crossing structures or to modify existing infrastructure can use the hotspot analysis to identify locations that optimize safety for multiple taxa or that target specific migration locations (Fig. 5). In addition, predictor variables that proved important in the model should be used to determine crossing locations outside the study area by identifying the types of locations that are most suitable. As long as target species and road types are similar and predictor variables are available or can be measured, a model could be constructed for most of South Korea.
The study also suggests ways in which South Koreans can improve local-level roadkill mitigation practices. Roadkill hotspot locations and identification of disconnected wildlife habitats can be evaluated in geographic information systems (GIS) by branch offices of the Road Maintenance Bureau to create metrics of road crossing “demand” and evaluate ongoing mitigation efforts in terms of location and function. In the study area, presentation of these results to transportation officials led to the construction of four wildlife crossing structures on the study road (Fig. 6), one of which is being monitored with heat sensor cameras.

Four wildlife crossings constructed as a result of this study: a Overpass 1 crosses from hill to hill where a mammal roadkill hotspot occurred on local road 19. b Underpass 1, located across the edge of agricultural land and mountains, is under construction. c Underpass 2 is a crossing on the riverside road that links the upland, a habitat to the river, where mammal, amphibian, and reptile roadkill hotspots occurred. d Underpass 3 has three subunderpasses that go under a road; a exclusion fence have also been installed along the road to guide wildlife to the underpass
Eight natural monument species and seven endangered species (including five natural monument species) were killed during this study. Mortality totals for two endangered species, the leopard cat and oriental scops owl, were >100, emphasizing a need to consider mitigation options for these protected species, even if background rates of demographic condition are not known. Further studies on these species are needed to determine whether wildlife crossings, exclusion fences, vegetation management, or other mitigation measures are effective.
Conclusion
This study provides roadkill information from an intensive 2.5-year survey near a South Korean national park in northeastern Asia, a region for which road impacts have not yet been reported. The study represents a significant improvement over previous, presence–absence analysis methods through a roadkill hotspot analysis that used the Getis–Ord Gi* statistic. However, there were some limitations, including the number of replicates for a comparative study of road types, the use of the same landscape factors for different taxonomic groups, and that few studies were available addressing the phenology of wildlife in South Korea. Future work is necessary to focus on: (1) extending the study to include replicates of the three road types, and possibly other classes of roads; (2) reclassifying landscape factors according to taxonomic groups and habitat types; and (3), improving the roadkill hotspot analysis through the calibration of parameters according to taxonomic groups and species characteristics. Ultimately, we hope these findings can inform countries that have similar landscapes to develop avoidance, minimization, and mitigation practices to lower the impact of roads on wildlife.
References
Alexander SM, Waters NM (2000) The effects of highway transportation corridors on wildlife: a case study of Banff National Park. Transp Res Part C 8:307–320
Allouche O, Tsoar A, Kadmon R (2006) Assessing the accuracy of species distribution models: prevalence, kappa and the true skill statistic (TSS). J Appl Ecol 43:1223–1232
Anselin L (1995) Local indicators of spatial association—LISA. Geogr Anal 27:93–115
Bager A, Rosa CA (2011) Influence of sampling effort on the estimated richness of road-killed vertebrate wildlife. Environ Manag 47:851–858
Balkenhol N, Waits LP (2009) Molecular road ecology: exploring the potential of genetics for investigating transportation impacts on wildlife. Mol Ecol 18:4151–4164
Baruch-Mordo S, Breck SW, Wilson KR, Theobald DM (2008) Spatiotemporal distribution of black bear human conflicts in Colorado, USA. J Wildl Manag 72:1853–1862
Beier P, Majka DR, Spencer WD (2008) Forks in the road: choices in procedures for designing wildland linkages. Conserv Biol 22:836–851
Bissonette JA, Rosa SA (2009) Road zone effects in small-mammal communities. Ecol Soc 14 (online). http://www.ecologyandsociety.org/vol14/iss1/art27/
Brito CJ (2003) Seasonal variation in movements, home range, and habitat use by male Vipera latastei in northern Portugal. J Herpetol 37:155–160
Choi T (2007) Road-kill mitigation strategies for mammals in Korea: data based on survey of road-kill non-wildlife passage use, and home range. Dissertation, Seoul National University, Seoul, South Korea
Clevenger PA, Chruszcz B, Gunson KE (2003) Spatial patterns and factors influencing small vertebrate fauna road-kill aggregations. Biol Conserv 109:15–16
Conard JM, Gipson PS (2006) Spatial and seasonal variation in wildlife–vehicle collisions. Prairie Nat 38:251–260
Crist MR, Wilmer B (2002) Roadless areas: the missing link in conservation. An analysis of biodiversity and landscape connectivity in the northern Rockies. The Wilderness Society, Washington, DC
Crooks K, Haas C, Baruch-Mordo S, Middledorf K, Magle S, Shenk T, Wilson K, Theobald D (2008) Roads and connectivity in Colorado: animal–vehicle collisions, wildlife mitigation structures, and lynx–roadway interactions. Colorado Department of Transportation Research Branch, Denver
Davies JM, Roper TJ, Shepherson DJ (1987) Seasonal distribution of road kills in the European badger (Meles meles). J Zool 211:525–529
Donaldson B (2007) Use of highway underpasses by large mammals and other wildlife in Virginia: factors influencing their effectiveness. Transp Res Rec 2007:157–164
Epps CW, Palsboll PJ, Wehausen JD, Roderick GK, Rmaey RR II, McCullough DR (2005) Highways block gene flow and cause a rapid decline in genetic diversity of desert bighorn sheep. Ecol Lett 8:1029–1038
ESRI (2009) ArcGIS desktop: release 9.3. Environmental Systems Research Institute, Redlands
Johnson ML et al (2002) North coast river loading study: road crossing on small streams. Status of salmonids in the watershed, vol I. CTSW-RT-02-040. Sacramento, CA, Caltrans, California Department of Transportation Sacramento, California
Fahrig L, Rytwinski T (2009) Effects of roads on animal abundance: an empirical review and synthesis. Ecol Soc 14 (online). http://www.ecologyandsociety.org/vol14/iss1/art21/
Farmer RG, Brooks RJ (2012) Integrated risk factors for vertebrate roadkill in southern Ontario. J Wildl Manag 76:1215–1224
Finder RA, Roseberry JL, Woolf A (1999) Site and landscape conditions at white-tailed deer/vehicle collision locations in Illinois. Landsc Urban Plan 44:77–85
Forman RTT (1995) Land mosaics: the ecology of landscapes and regions. Cambridge University Press, Cambridge
Forman RTT, Alexander LE (1998) Roads and their major ecological effects. Annu Rev Ecol Syst 29:207–231
Forman RTT, Friedman DS, Fitzhenry D, Martin JD, Chen AS, Alexander LE (1997) Ecological effects of roads: toward three summary indices and an overview for North America. In: Canters K, Piepers A, Hendriks-Heersma D (eds) Proceedings habitat fragmentation and infrastructure. Ministry of Transport, Public Works and Water Management, Delft, pp 40–54
Forman RTT et al (2003) Road ecology: science and solutions. Island Press, Washington
Fuglestvedt J, Berntsen T, Myhre G, Rypdal K, Skeie RB (2008) Climate forcing from the transport sectors. Proc Natl Acad Sci 105:454–458
Gelbard JL, Belnap J (2003) Roads as conduits for exotic plant invasions in a semiarid landscape. Conserv Biol 17:420–432
Getis A, Ord JK (1996) Local spatial statistics: an overview. In: Longley P, Batty M (eds) Spatial analysis: modelling in a GIS environment. Wiley, New York
Girvetz EH, Thorne JH, Berry AM, Jaeger JAG (2008) Integration of landscape fragmentation analysis into regional planning: a state-wide multiscale case study for California. Landsc Urban Plan 86:205–218
Giudice JH, Ratti JT (2001) Ring-necked pheasant (Phasianus colchicus). In: Poole A, Gill F (eds) The birds of North America, number 572. Academy of Natural Sciences, Philadelphia, pp 1–32
Glista DJ, DeVault TL, DeWoody JA (2007) Vertebrate road mortality predominantly impacts amphibians. Herpetol Conserv Biol 3:77–87
Gomes L, Grilo C, Silva C, Mira A (2009) Identification methods and deterministic factors of owl roadkill hotspot locations in Mediterranean landscapes. Ecol Res 24:355–370
Grilo C, Bissonette JA, Margarida S (2009) Spatial–temporal patterns in Mediterranean carnivore road casualties: consequences for mitigation. Biol Conserv 142:301–313
Gunson KE, Clevenger AP, Ford AT, Bissonette JA, Hardy A (2009) A comparison of data sets varying in spatial accuracy used to predict the locations of wildlife–vehicle collisions. Environ Manag 44:268–277
Gunson KE, Mountrakis G, Quackenbush L (2011) Spatial wildlife–vehicle collision models: a review of current work and its application to transportation mitigation projects. J Environ Manag 92:1074–1082
Hanley JA, McNeil BJ (1982) The meaning and use of the area under a receiver operating characteristic (ROC) curve. Radiology 143:29–36
Harris LD, Scheck J (1991) From implications to applications: the dispersal corridor principle applied to the conservation of biological diversity. In: Saunders DA, Hobbs RJ (eds) Nature conservation 2: the role of corridors. Surrey beatty, Chipping Norton, pp 189–220
Havlick DG (2002) No place distant: roads and motorized recreation on America’s public lands. Island Press, Washington
Hughes WE, Saremi AR, Paniati JF (1996) Vehicle–animal crashes: an increasing safety problem. Inst Transp Eng J 66:24–28
IRF (International Road Federation) (2010) World road statistics 2010. IRF, Alexandria
Jaeger J, Bowman J, Brennan J, Fahrig L, Bert D, Bouchard J, Charbonneau N, Frank K, Gruber B, Toschanowitz K (2005) Predicting when animal populations are at risk from roads: an interactive model of road avoidance behavior. Ecol Model 185:329–348
Joyce TL, Mahoney SP (2001) Spatial and temporal distributions of moose–vehicle collisions in Newfoundland. Wildl Soc Bull 29:281–291
Kvamme KL (1985) Determining empirical relationships between the natural environment and pre-historic site locations: a hunter–gatherer example. In: Carr C (ed) For concordance in archeological analysis. Kansas City, pp 208–238
Langen AT, Ogden KM, Schwarting LL (2009) Predicting hot spots of herpetofauna road mortality along highway networks. J Wildl Manag 73:104–114
Li R, Shilling F, Thorne J, Li F, Schott H, Boynton R, Berry AM (2010) Fragmentation of China’s landscape by roads and urban areas. Landsc Ecol 25:839–853
Marshall TJ (1978) Systematics of smaller Asian night birds based on voice. Ornithol Monogr 25:1–58
Ministry of Environment (2007) The plan for Korean Peninsula Ecological Network. Ministry of Environment, Gwacheon
Ministry of Environment (2011) The status of national Eco-Corridor establishment. Ministry of Environment, Gwacheon
Monserud RA, Leemans R (1992) Comparing global vegetation maps with the Kappa statistic. Ecol Model 62:275–293
National Research Council (2005) Assessing and managing the ecological impacts of paved roads. National Academies Press, Washington, DC
Nielsen CK, Anderson RG, Grund MD (2003) Landscape influences on deer vehicle accident areas in an urban environment. J Wildl Manag 67:46–51
Ord JK, Getis A (1995) Local spatial autocorrelation statistics: distribution issues and an application. Geogr Anal 27:286–306
Press SJ, Wilson S (1978) Choosing between logistic regression and discriminant analysis. J Am Stat Assoc 73:699–705
Rajaratnam R, Sunquist M, Rajaratnam L, Ambu L (2007) Diet and habitat selection of the leopard cat (Prionailurus bengalensis borneoensis) in an agricultural landscape in Sabah, Malaysian Borneo. J Trop Ecol 23:209–217
Reijnen R, Foppen R (1995) The effects of car traffic on breeding bird populations in woodland. IV. Influence of population size on the reduction of density close to a highway. J Appl Ecol 32:481–491
Reijnen R, Foppen R, Braak CT, Thissen J (1995) The effects of car traffic on breeding bird populations in woodland. III. Reduction of density in relation to the proximity of main roads. J Appl Ecol 32:187–202
Riley SPD, Pollinger JP, Sauvajot RM, York EC, Bromley C, Fuller TK, Wayne RK (2006) A southern California freeway is a physical and social barrier to gene flow in carnivores. Mol Ecol 15:1733–1741
Ripley BD (1981) Spatial statistics. Wiley, New York
Schmetterling AD, Young MK (2008) Summer movements of boreal toads (Bufo boreas boreas) in two western Montana basins. J Herpetol 42:111–123
Smith LL, Smith KG, Barichivich WJ, Dodd CK Jr, Sorensen K (2005) Roads and Florida’s herpetofauna: a review and mitigation case study. In: Meshaka WE, Babbitt KJ Jr (eds) Status and conservation of Florida amphibians and reptiles. Krieger Publications, Malabar
Spooner PG, Lunt ID, Okabe A, Shiode S (2004) Spatial analysis of roadside Acacia populations on a road network using the network K-function. Landsc Ecol 19:490–499
Statistics Korea (2011) Road statistics of OECD countries. Statistics Korea, Seoul, South Korea (online). http://www.index.go.kr/egams/stts/jsp/potal/stts/PO_STTS_IdxMain.jsp?idx_cd=1207
Taylor BD, Goldingay RL (2004) Wildlife roadkills on three major roads in north-eastern New South Wales. Wildl Res 31:83–91
Thorne JH, Girvetz EH, McCoy M (2009) Evaluating aggregate terrestrial impacts of road construction projects for advanced regional mitigation. Environ Manag 43:936–948
Thuiller W, Lafourcade B, Engler R, Araujo MA (2009) BIOMOD—a platform for ensemble forecasting of species distributions. Ecography 32:369–373
Trombulak SC, Frissell CA (2000) Review of ecological effects of roads on terrestrial and aquatic communities. Conserv Biol 14:18–30
Turner MG, Gardner RH, O’Neill RV (2001) Landscape ecology in theory and practice: pattern and process. Springer, New York
West E (2006) Bioacoustic profiles: evaluating potential masking of wildlife vocal communication by highway noise. In: Irwin CL, Garrett P, McDermott KP (eds) Proceedings of the 2005 international conference on ecology and transportation. Center for Transportation and the Environment, North Carolina State University, Raleigh, p 29
Acknowledgments
Funding for this study was provided by the Korean Technology Development Agency of the Ministry of the Environment and the Eco-STAR Project. The work was accomplished due to the field efforts of technicians, and we greatly appreciate their dedication. We acknowledge, in particular, the efforts of Cheonkwon Choi and Donggi Choi in coordinating the surveys used as the basis of this work.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article
Cite this article
Seo, C., Thorne, J.H., Choi, T. et al. Disentangling roadkill: the influence of landscape and season on cumulative vertebrate mortality in South Korea. Landscape Ecol Eng 11, 87–99 (2015). https://doi.org/10.1007/s11355-013-0239-2
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11355-013-0239-2