
Abstract
Road-killed mammals, birds, and reptiles were collected weekly from highways in southern Brazil in 2002 and 2005. The objective was to assess variation in estimates of road-kill impacts on species richness produced by different sampling efforts, and to provide information to aid in the experimental design of future sampling. Richness observed in weekly samples was compared with sampling for different periods. In each period, the list of road-killed species was evaluated based on estimates the community structure derived from weekly samplings, and by the presence of the ten species most subject to road mortality, and also of threatened species. Weekly samples were sufficient only for reptiles and mammals, considered separately. Richness estimated from the biweekly samples was equal to that found in the weekly samples, and gave satisfactory results for sampling the most abundant and threatened species. The ten most affected species showed constant road-mortality rates, independent of sampling interval, and also maintained their dominance structure. Birds required greater sampling effort. When the composition of road-killed species varies seasonally, it is necessary to take biweekly samples for a minimum of one year. Weekly or more-frequent sampling for periods longer than two years is necessary to provide a reliable estimate of total species richness.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Highways are important for socioeconomic development (Perz and others 2007). For the maintenance of biodiversity, however, highways have negative effects, involving habitat fragmentation (Forman and Deblinger 2000; Soares-Filho and others 2004), hydrological changes (Jones and others 2000), sound pollution (Reijnen and others 1995), introduction of exotic species (Forman and others 2003), and road accidents (Bager 2003). Road mortality has been shown to be one of the main causes of biodiversity loss of several vertebrate species (Coffin 2007; Trombulak and Frissell 2000), exceeding other impacts such as hunting (Forman and Alexander 1998). Other species are affected indirectly, through changes in rates of movement (McGregor and others 2008), which may segregate populations and alter their structure.
Studies that monitor highway mortality of the fauna use different sampling designs. Kleist and others (2007) used weekly intervals, Gomes and others (2009) and Grilo and others (2009) used biweekly samplings, Coelho and others (2008) used monthly intervals, and Smith-Patten and Patten (2008) used varied and irregular intervals. Other investigators did not report the intervals used (Brockie and others 2009). In Brazil, the majority of studies on highway mortality of the fauna have been descriptive, resulting merely in lists of species, without any standardization of the methods used or a clear definition of sampling effort (Bager and others 2007).
Identification and quantification of the fauna killed on roads is required by environmental agencies in different countries, in order to evaluate the impact of highways on biodiversity. The reliability of estimates of the impact of highway mortality on the wild fauna is often compromised by limitations of time and financial resources. Consequently, critical areas (roadkill hotspots) cannot be identified; threatened species, which are normally rare, do not appear on the list of affected species; and costly apparatuses intended to mitigate roadkill impact are proposed and installed with little effect on conservation.
The aims of the present study were: (1) to test the effect of different sampling efforts on the estimation of richness of road-killed species; (2) to evaluate the extent of sampling necessary for reptiles, birds, and mammals; and (3) to analyze the variation in abundance and composition of the species that are most often killed on roads, using different sampling intervals.
Methods
Study Area
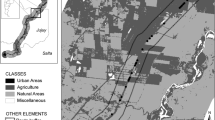
Sampling was carried out along 117 km of federal highways (BR 392 and 471) in extreme southern Brazil. The study area extended from the municipality of Pelotas (52°20′37′′ W; 31°47′34′′ S) to a federally protected area, the Taim Ecological Station (ESEC Taim) (52°35′42′′ W; 32°40′36′′ S) (Fig. 1). The region lies on the coastal plain of the state of Rio Grande do Sul. The low-relief terrain contains restingas (coastal dune forests) and extensive temporarily or permanently flooded areas, together with fresh- and brackish-water ponds (Carvalho and Rizzo 1994). The climate is subtropical (Cfa; Köeppen 1948) and rainfall is evenly distributed throughout the year. Between January 1991 and July 2002 the mean monthly rainfall was 125 mm, and the mean monthly temperature was 18°C in the region.

Study area between the municipality of Pelotas and the Taim Ecological Station in southern Brazil
Sampling Design
We monitored the roads weekly from January through December in the years 2002 and 2005, always in the direction from Pelotas to the ESEC Taim, at a speed of 50 km/h. The highway is one lane in each direction, which allows road-killed animals (reptiles, birds, and medium and large-sized mammals) to be seen over its entire width. The sample of mammals included species as large as Lutreolina crassicaudata (~300 g). For birds and reptiles, all the specimens observed were included in the analyses. Each animal found was identified, and geographical position (determined with a GPS) was noted.
Data Analysis
For the analysis of the effect of sampling effort on the richness of road-killed species, we considered the richness observed in the weekly monitoring as the maximum impacted richness. The sampling effort consisted of 35 samples in 2002 and 42 in 2005. We then carried out resamplings that used biweekly, monthly, bimonthly, and seasonal samples. Each resampling resulted from the mean of ten randomizations. The analysis was done for the years 2002 and 2005 individually, and for both years together. The analyses of individual years were done in order to identify temporal changes in the roadkill rates. For annual samplings we used 24 monitoring events for the biweekly samplings, 12 for monthly, 6 for bimonthly, and 4 for seasonal; for the combined years, the number of monitoring events was doubled. The roadkill rates refer to the number of road-killed animals per sampling day (number of animals killed on the 117 km).
Using each day of monitoring as the sampling unit (Sample-Based), the species accumulation curve was analyzed using the species richness estimator Chao 1, in the program EstimateS 7.5 (Colwell 2005), calculating the estimated richness and the standard deviation for each of the intervals (Chazdon and others 1998; Colwell and Coddington 1994). We considered that the sampling effort of the resamplings was reached when the estimated richness plus one standard deviation was equal to or greater than the richness observed by weekly sampling. The comparison of observed richness between the years 2002 and 2005 was carried out with rarefaction analyses, using the program Ecosim 7 (Gotelli and Entsminger 2007), with 10,000 iterations and a significance level of 0.05.
For each species, the abundance of road kills in 2002 and 2005 was summed, identifying the 10 most affected species in the weekly samples (Reptiles: Helicops infrataeniatus—water snake; Trachemys dorbigni—Orbigny’s slider turtle; Phrynops hilarii—Hilaire’s side-necked turtle; Philodryas patagoniensis—Patagonia Green Racer; Mammals: Myocastor coypus—Nutria; Didelphis albiventris—White-eared Opossum; Hydrochaeris hydrochaeris—Capybara; Conepatus chinga—Molina’s hog-nosed skunk; Cerdocyon thous—Crab-eating fox; Bird: Chrysomus ruficapillus—Chestnut-capped blackbird). These ten species were analyzed in the different resampling periods, with respect to (1) the minimum number of samplings that was necessary so that they would be represented in the affected community, (2) the percentage of species present, and (3) the variation in roadkill rates. Using ten resamplings, we calculated the mean sample number that was necessary for all ten species to be included in the list of affected species (1). The percentage of species present (2) was used to establish which resampling period was capable of identifying all ten species, and which resulted in the absence of one or more species. When all the species were present in the ten resamplings, we considered this as 100% success; the absence of one species from one of the resamplings represented 90% success; and so on. We evaluated the existence of variation in the road-mortality rate of each of the ten species between the different resampling periods (3) by means of Friedman’s ANOVA, using a significance level of 0.05. Complementarily, we assessed the association of roadkill rate of each of the ten species between the different sampling intervals with Kendall’s coefficient of concordance. Both analyses were executed with Biostat 5.0 (Ayres and others 2007).
We evaluated the presence of the two threatened species that were affected by road mortality (Leopardus geoffroyi—Geoffroy’s cat, and Lontra longicaudis—Neotropical River Otter) in the different resampling intervals, using the same method of percentage of species present (2). Finally, we tested the effect of short sampling periods (n = 10 weeks) on the determination of roadkill rates. We calculated the mean rate in the first 10 weeks, then we eliminated the first week and calculated the mean rate between the second and 11th week, and so on until the 35th or 42nd week, for 2002 and 2005 respectively.
Results
Species richness was higher for 2002 than for 2005 (Table 1). The rarefaction curve identified a difference in richness between the 2 years, and was lower than expected in 2005 (S est = 76 ± 4). This difference was related to birds, which in 2002 showed a richness of 53 species but only 21 in 2005, whereas for reptiles and mammals, richness was similar between the two years (Table 1). Observed richness was 56.5% of that estimated by Chao 1, independently of the year. The lowest sampling success was obtained for birds, but for mammals, sampling sufficiency was 100% in both years (Table 1).
The richness obtained with weekly samples was reached in the estimates calculated for biweekly and monthly resamplings in 2002, and for biweekly resamplings in 2005. Similar results were found when the classes were analyzed separately (Table 1). The combination of the 2 years confirmed the necessity for biweekly or monthly samples, according to the class of animals; however, the richness of reptiles was not sufficiently estimated in any of the resampling periods.
The ten species with the highest roadkill rates in the weekly samples, were also present in all the biweekly and monthly resamplings, but were absent from some samplings in the other intervals. The distribution of roadkill rates among the most-affected species remained constant, and the pattern identified in the weekly samples was maintained in all the resampling periods (Fig. 2). This was more evident in 2002, where the snake H. infrataeniatus dominated. The only change in the pattern was the absence of C. thous from the seasonal resamplings. In 2005 the pattern was repeated, but with changes in the order of the most-affected species. H. infrataeniatus remained the most abundant species, except in the bimonthly resamplings, where it was replaced by D. albiventris. Friedman’s ANOVA demonstrated that there were no differences in the roadkill rates of each species in the different resampling intervals (2002: Fr² (N = 9, df = 4) = 5.207; P < 0.267; 2005: Fr² (N = 10, df = 4) = 2.640; P < 0.620). Kendall’s coefficient of concordance was low, 0.145 and 0.066 for 2002 and 2005, respectively.

Frequency distribution of the roadkill rates of the ten most-affected species in the years 2002 (a) and 2005 (b). The bars indicate the different resampling intervals. H.i., Helicops infrataeniatus; M.c., Myocastor coypus; C.r., Chrysomus ruficapillus; H.h., Hydrochaeris hydrochaeris; D.a., Didelphis albiventris; C.c., Conepatus chinga; C.t., Cerdocyon thous; T.d., Trachemys dorbigni; P.p., Philodryas patagoniensis; P.h., Phrynops hilarii
Rates of detection of the ten most-affected species varied significantly between the two years (Table 2). In 2002, all the species were identified after three weekly samplings, whereas in 2005, 14 samplings were necessary. This difference was related to C. ruficapillus, which was included after eight samplings of the next-to-last species, H. hydrochaeris. C. ruficapillus was the third most-impacted in 2002, but only the tenth in 2005 (Fig. 2). There was also a difference between years in the percentage of resampling. In 2002, all ten species were present in 100% of the biweekly and monthly resamplings, and in 2005 only in the biweekly samplings. Bimonthly and seasonal samples were not efficient in either year, with two to eight species absent.
The two threatened species (L. geoffroyi and L. longicaudis) showed different results for the resamplings and in the different years. In 2002, in the weekly samplings, both species were sampled in the first monitoring event. In 2005, they were only identified after longer sampling periods (Table 2). For the resamplings, only the biweekly resampling showed high efficiency in the identification of these species.
The means for the ten periods of consecutive weekly samplings demonstrated that short-term sampling can result in erroneous analyses of the impact of road mortality on seasonal species (Fig. 3). Pronounced seasonality was apparent in the roadkill rates of H. infrataeniatus (in summer and the beginning of autumn) and M. coypus (in the winter months), and a slight seasonality for D. albiventris. The roadkill rates of the two species that were most influenced by seasonality were asynchronous in the 2 years.

Sazonal variation of roadkill rates of Helicops infrataeniatus, Myocastor coypus, and Didelphis albiventris in 2002 (a) and 2005 (b). Each point in X-axis is a mean roadkill rates for the ten weekly sampling periods
Discussion
The weekly samplings did not attain sampling sufficiency in either of the years when all the classes were considered together, demonstrating that even this sampling effort is insufficient to identify all of the species affected by road mortality. The Chao 1 estimator is conservative (Gotelli 2009), and therefore it emphasized the inefficiency of weekly sampling in evaluations of the entire community. Weekly sampling was adequate for reptiles and middle and large-sized mammals, although it was not effective in sampling all of the bird species. These results reflect the high richness of bird species in the region, which supports between 176 and 220 species depending on the area considered (Dias and Maurício 1998; Mähler and others 1996). The observed richness of road-killed avifauna represented less than one-fourth of the species that occur near the highways, but the estimated richness indicated that more than half of the species reported from the region may be affected.
The total species richness varied between the two sampling years, and a high proportion of species occurred in only one of the years. If the samples had been taken only in 2005, the estimated richness would be 92 species, which would be an underestimate because the richness observed in the 2 years was equal to this value and did not reach sampling sufficiency. This outcome serves as an alert that the total number of species affected by road mortality can only be determined with difficulty, and that the curve of sampling sufficiency will only stabilize after years of weekly or more-frequent sampling. Esbérard and Bergallo (2008) stated that an increase in sampling effort reduces the number of singleton and doubleton species; this was confirmed by the significant reduction in the estimated richness of birds when we analyzed the 2 years together. Sampling carried out during only one year may increase the proportion of species of low occurrence, because this will sample seasonal or migratory species that do not repeat in subsequent samplings (Gotelli 2009). The effect of clumped distributions on our analyses must also be considered, because in this case the rarefaction analyses inevitably overestimate the expected richness (Gotelli and Colwell 2001).
Surveys of specific classes require different efforts. For birds, the biweekly samples resulted in samples that were similar to those done weekly, but in all cases did not adequately characterize the estimated bird community. The effects of highways on the avifauna must be evaluated with caution and can require intensive monitoring for at least a year, if the objective is to identify the largest number of species. However, considering that the percentage of abundant species is low, the same effort as proposed for the reptiles results in statistically meaningful data for the road-killed avifauna. For medium- to large-sized mammals, which normally have a lower richness and are less influenced by seasonal climate variations (at least in subtropical and tropical regions) (Caro and others 2000; Coelho and others 2008; Taylor and Goldingay 2004), year-round monitoring is not required, and therefore the sampling effort can be concentrated. The richness of mammals was little influenced by sampling effort. For the reptiles, monthly monitoring also resulted in representative sampling of the community affected by road mortality. The region is dominated by wetlands, and although no population studies exist for the majority of the reptile species, it is known that highways constructed through or near wetlands are significant sources of mortality for turtle populations (Aresco 2003; Ashley and Robinson 1996). Factors such as recruitment and migration in breeding seasons increase road mortality of turtles and snakes (Bager 2003; Bonnet and others 1999). For the ten species that were most affected by road mortality, two, a reptile and a mammal, are strongly influenced by seasonality (H. infrataeniatus and M. coypus). Helicops infrataeniatus shows high rates of road mortality in its breeding season and when the young are born (Aguiar and Di-Bernardo 2005). Myocastor coypus shows the highest road-mortality rates during rainy periods, when its burrows are flooded and the nutrias must seek out dry areas on the highway embankments. Probably different species, whether or not they are significantly affected by road mortality, have different degrees of seasonal variation in the rates of roadkill, as well as asynchronies that make it necessary to carry out continuous sampling for at least one annual cycle.
Except in specific cases, measures to mitigate the impact of roadkills are directed toward the species whose abundances are most affected (Bager 2003), generally medium and large-sized animals that cause personal and material harm to humans (Romin and Bissonette 1996), threatened species (Philcox and others 1999), or a combination of these (Huijser and others 2009). Species that are rarely killed on roads, or those that are rare but important for the maintenance of ecosystem functions, will not appear in the samples in sufficient numbers to allow identification of those localities where the incidence of roadkills is highest (hotspots), and therefore the proper design of mitigation measures. For the species that are killed in greater numbers, biweekly monitoring for one year gave satisfactory results, similar to the weekly samples. Thus, in the case of monitoring for the purpose of placing structures to mitigate highway mortality, we suggest carrying out at least biweekly sampling. Therefore, sampling design should be analyzed with care, because of the possible impact on threatened species with low population densities, where the loss of even a few animals can severely impact the long-term maintenance of the population, principally for species with a long life cycle and low fecundity (Grelle and others 2006). In this study, L. longicaudis and L. geoffroyi were present in the biweekly and monthly resamplings, and were incorporated into the list of affected species after different sampling periods. Conservation of community structure, and consequently its influence on the maintenance of ecological processes, has raised questions about the importance of the identification and inclusion of rare species in studies of community structure. The inclusion of these species requires a more-detailed treatment and consequently a greater sampling effort in time and frequency. We must distinguish between what is a rare species among those killed on the road, and a rare species in the community surrounding the highway. No existing studies have demonstrated a relationship between these two “rarities”.
Independently of the sampling effort used, none sampling schemes allows evaluation of the impact of road mortality on the populations of the species that inhabit the areas surrounding the highway. Species that are abundant but that are highly sensitive to effects such as noise, fragmentation, and alteration of the vegetation structure, are only occasionally impacted (Fahrig and Rytwinski 2009; Jaeger and others 2005). In this context, the lack of data on the abundance of the species in the surrounding area makes it difficult to evaluate the impact of a highway on the populations and communities affected. Future studies must give priority to analyses of the selectivity of road mortality, in order to increase the effectiveness of their conservation proposals when the objective is to maintain ecological processes.
Conclusions
Defining an appropriate sampling effort is fundamental in order adequately to represent the richness of road-killed species. The sampling design must consider the specific objective of the proposed study (e.g., total richness, most-affected species, specific classes). In the case of monitoring studies focusing on the total vertebrate richness, including reptiles, birds, and mammals, the minimum effort must involve weekly or more-frequent samples, for periods longer than 1 year. In regions with pronounced environmental seasonality, the priority is biweekly samplings for a period of at least 1 year. When the objective is the identification of the most-affected species, biweekly monitoring for 1 year is sufficient.
References
Aguiar LFS, Di-Bernardo M (2005) Reproduction of the water snake Helicops infrataeniatus (Colubridae) in southern Brazil. Amphibia-Reptilia 26:527–533
Aresco MJ (2003) Highway mortality of turtles and other herpetofauna at Lake Jackson, Florida, USA and the efficacy of a temporary fence/culvert system to reduce roadkills. In: Irwin CL, Garrett P, McDermott KP (eds) Proceedings of International Conference on Ecology and Transportation, Center for Transportation and the Environment, North Carolina State University, Raleigh, North Carolina, pp 433–449
Ashley PE, Robinson JT (1996) Road mortality of amphibians, reptiles and other wildlife on the Long Point Causeway, Lake Erie. Canadian Field-Naturalist 110:403–412
Ayres M, Ayres M Jr, Ayres DL, Santos AS (2007) Bioestat Version 5.0: Aplicações estatísticas nas áreas das ciências biológicas e médicas. Sociedade Civil Mamirauá, Brasil
Bager A (2003) Repensando as medidas mitigadoras impostas aos empreendimentos rodoviários associados a Unidades de Conservação – Um estudo de caso, In: Bager A (ed) Áreas Protegidas: Conservação no Âmbito do Cone Sul, Bager A, Pelotas, Brasil, pp 159–172
Bager A, Piedras SRN, Pereira TSM, Hobus Q (2007) Fauna selvagem e atropelamento.- diagnóstico do conhecimento científico Brasileiro. In: Bager A (ed) Áreas Protegidas: Repensando as Escalas de Atuação. Armazém Digital, Porto Alegre, pp 49–62
Bonnet X, Naulleau G, Shine R (1999) The dangers of leaving home: dispersal and mortality in snakes. Biological Conservation 89:39–50
Brockie RE, Richard MFSS, Wayne LL (2009) Long-term wildlife road-kill counts in New Zealand. New Zealand Journal of Zoology 36:123–134
Caro TM, Shargel JA, Stoner CJ (2000) Frequency of medium-sized mammal road kills in an agricultural landscape in California. American Midland Naturalist 144:362–369
Carvalho VC, Rizzo HG (1994) A zona costeira Brasileira: subsídios para uma avaliação ambiental. Ministério do Meio Ambiente e da Amazônia Legal, Brasil, 211 p
Chazdon RL, Colwell RK, Denslow JS, Guariguata MR (1998) Statistical methods for estimating species richness of woody regeneration in primary and secondary rain forests of NE Costa Rica. In: Dallmeier F, Comiskey JA (eds) Forest biodiversity research, monitoring and modeling: conceptual background and old world case studies. Parthenon Publishing, New York, pp 285–309
Coelho IP, Kindel A, Coelho AVP (2008) Roadkills of vertebrate species on two highways through the Atlantic Forest Biosphere Reserve, southern Brazil. European Journal of Wildlife Research 54:689–699
Coffin AW (2007) From roadkill to road ecology: a review of the ecological effects of roads. Journal of Transport Geography 15:396–406
Colwell RK (2005) EstimateS Version 7.5: Statistical estimation of species richness and shared species from samples. http://purl.oclc.org/estimatesq. Accessed online 15 Nov 2009
Colwell RK, Coddington JA (1994) Estimating terrestrial biodiversity through extrapolation. Philosophical Transactions of the Royal Society B 345:101–118
Dias RA, Maurício GN (1998) Lista preliminar da avifauna da extremidade sudoeste do saco da Mangueira e arredores, Rio Grande, Rio Grande do Sul. Atualidades Ornitológicas 86:10–11
Esbérard CEL, Bergallo HG (2008) Influência do esforço amostral na riqueza de espécies de morcegos no sudeste do Brasil. Revista Brasileira de Zoologia 25:67–73
Fahrig L, Rytwinski T (2009) Effects of roads on animal abundance: an empirical review and synthesis. Ecology and Society 14(1):21 (online article)
Forman RTT, Alexander LE (1998) Roads and their major ecological effects. Annual Review of Ecology, Evolution and Systematics 29:207–231
Forman RTT, Deblinger RD (2000) The ecological road-effect zone of a Massachusetts (USA) suburban highway. Conservation Biology 14:36–46
Forman RTT, Sperling D, Bissonette JA, Clevenger AP, Cutshall CD, Dale VH, Fahrig L, France R, Goldman CR, Heanue K, Jones JA, Swanson FJ, Turrentine T, Winter TC (2003) Road ecology: science and solutions. Island Press, Washington
Gomes L, Grilo C, Silva C, Mira A (2009) Identification methods and deterministic factors of owl roadkill hotspot locations in Mediterranean landscapes. Ecological Research 24:355–370
Gotelli NJ (2009) Ecologia. Planta Press, Brasil
Gotelli NJ, Colwell RK (2001) Quantifying biodiversity: procedures and pitfalls in the measurement and comparison of species richness. Ecology Letters 4:379–391
Gotelli NJ, Entsminger GL (2007) EcoSim, Version 7.0: Null models software for ecology. http://homepages.together.net/~gentsmin/ecosim.htm
Grelle CEV, Paglia AP, Silva HS (2006) Análise dos fatores de ameaça de extinção: estudo de caso com os mamíferos brasileiros. In: Rocha CFD, Bergallo HG, Alves MAS, Sluys MV (eds) Biologia da Conservação. Rima Press, Brasil, pp 361–374
Grilo C, Bissonette JA, Santos-Reis M (2009) Spatial-temporal patterns in Mediterranean carnivore road casualties: consequences for mitigation. Biological Conservation 142:301–313
Huijser MP, Duffield JW, Clevenger AP, Ament RJ, McGowen PT (2009) Cost-benefit analyses of mitigation measures aimed at reducing collisions with large ungulates in the United States and Canada: a decision support tool. Ecology and Society 14(2):15 (online article)
Jaeger JAG, Bowman J, Brennan J, Fahrig L, Bert D, Bouchard J, Charbonneau N, Frank K, Gruber B, von Toschanowitz KT (2005) Predicting when animal populations are at risk from roads: an interactive model of road avoidance behavior. Ecological Modelling 185:329–348
Jones JA, Swanson FJ, Wemple BC, Snyder KU (2000) Effects of roads on hydrology, geomorphology and disturbance patches in stream networks. Conservation Biology 14:76–85
Kleist AM, Lancia RA, Doerr PD (2007) Using video surveillance to estimate wildlife use of a highway underpass. Journal Wildlife Management 71:2792–2800
Köeppen W (1948) Climatologia. Fondo de Cultura Económica, México, p 479
Mähler JK Jr, Kindel A, Kindel EAI (1996) Lista comentada das espécies de aves da Estação Ecológica do Taim, Rio Grande do Sul, Brasil. Acta Biolologica Leopoldensia 18:69–103
McGregor RL, Bender DJ, Fahrig L (2008) Do small mammals avoid roads because of the traffic? Journal of Applied Ecology 45:117–123
Perz SG, Caldas MM, Arima E, Walker RJ (2007) Unofficial road building in the Amazon: socioeconomic and biophysical explanations. Development and Change 38:529–551
Philcox CK, Grogan AL, Macdonald DW (1999) Patterns of otter Lutra lutra road mortality in Britain. Journal of Applied Ecology 36:748–762
Reijnen R, Foppen R, Braak CT, Thissen J (1995) The effects of car traffic on breeding bird populations in woodland. Reduction of density in relation to the proximity of main roads. Journal of Applied Ecology 32:187–202
Romin LA, Bissonette JA (1996) Temporal and spatial distribution of highway mortality of mule deer on newly constructed roads at Jordanelle Reservoir, Utah. Great Basin Naturalist 56:1–11
Smith-Patten BD, Patten MA (2008) Diversity, seasonality, and context of mammalian roadkills in the Southern Great Plains. Environmental Management 41:844–852
Soares-Filho B, Alencar A, Nepstad D, Cerqueira G, Diaz MCV, Rivero S, Solórzano L, Voll E (2004) Simulating the response of land-cover changes to road paving and governance along a major Amazon highway: the Santarém-Cuiabá corridor. Global Change Biology 10:745–764
Taylor BD, Goldingay RL (2004) Wildlife road-kills on three major roads in north-eastern New South Wales. Wildlife Research 31:83–91
Trombulak SC, Frissell CA (2000) Review of ecological effects of roads on terrestrial and aquatic communities. Conservation Biology 14:18–30
Acknowledgments
We thank the Fundação O Boticário de Proteção à Natureza, FAPERGS, and FAPEMIG for financial support, which allowed us to carry out the project that resulted in this manuscript. We are grateful for suggestions made by Dr. S. R. Freitas and C. Bueno on the initial versions of the study.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Bager, A., da Rosa, C.A. Influence of Sampling Effort on the Estimated Richness of Road-Killed Vertebrate Wildlife. Environmental Management 47, 851–858 (2011). https://doi.org/10.1007/s00267-011-9656-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00267-011-9656-x