
Abstract
An increasing number of wildlife-vehicle collisions occur each year worldwide, which involves extensive economic costs and constitutes one of the main anthropogenic causes of animal mortality. Because of this, there is an urgent need to identify the factors leading to collision hotspots and thus implementing effective mitigation measures. By using a stratified random sampling survey, we investigated the fine-scale determinants of roadkill probability in small and medium-sized birds and mammals across a country-size region of Southern Spain, Andalusia (87,000 km2), located within a global biodiversity hotspot. During two consecutive seasons, we regularly surveyed 45 road transects of 10 km each and characterized the site-specific attributes of both roadkill and random points, including traffic density, road design (embankments, medians, fences, roadside vegetation and distance to curves), and adjacent landscape matrix. Based on this information, we investigated variation in collision risk according to landscape and road features, and the life history of the affected taxa. Mortality rates of mammals and birds increased with traffic density, and were also significantly affected by the distance to the nearest curve, slope of embankments, height of roadside vegetation, and land use adjacent to roads. Road mortality of both birds and mammals was related to the presence and typology of fences and center medians, so more densely vegetated medians and smaller mesh sizes reduced roadkill probability. Overall, our results indicate that roadkill risk may vary at exceedingly small spatial scales. The information provided by this extensive survey may be used to identify taxa-specific factors associated to roadkill risk and priority points for action. Our findings will therefore be relevant for the design of safer roads for both drivers and wildlife through the application of effective mitigation measures.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Roads have multiple ecological impacts, as they can act as barriers by limiting connectivity among populations, contaminate adjacent ecosystems, alter animal behavior, and facilitate dispersal of exotic species, among others (reviewed in Sandberg et al. 1998; Trombulak and Frissell 2000; Forman et al. 2003; Coffin 2007; van der Ree et al. 2015b). However, traffic-related mortality due to wildlife-vehicles collisions (WVCs, hereafter) appears to be the most important ecological impact of roads (Trombulak and Frissell 2000; Coffin 2007).
WVCs are an important traffic safety issue that involves significant monetary costs, primarily due to human injury and material damage (Bissonette et al. 2008; Huijser et al. 2009), but also high environmental costs (Forman et al. 2003; Erritzøe et al. 2003; Bissonette et al. 2008). Traffic related mortality is considered one of the most important sources of non-natural mortality in wildlife populations (Forman et al. 2003; Erritzøe et al. 2003; Colino-Rabanal and Lizana 2012). For instance, considering only birds, 27 and 80 millions of fatalities are estimated to occur annually in Europe (Erritzøe et al. 2003) and the United States (Erickson et al. 2005), respectively, although actual numbers may be higher. Traffic related mortality may dramatically affect population dynamics (e.g. through differential incidence into a gender or age class; Madsen et al. 2002; Colino-Rabanal and Lizana 2012) and constitutes a major threat for endangered species (Mumme et al. 2000; Gibbs and Shriver 2002). Importantly, because of the expansion of the road network and the increase in traffic volume, the ecological impact of roads on wildlife is expected to increase over the next decades in both developed and developing countries (Fulton and Eads 2004; Meyer et al. 2012; van der Ree et al. 2015c). Thus, quantifying the impact of roads on wildlife and developing effective mitigation measurements is urgently needed to balance future development requirements and biodiversity conservation (van der Ree et al. 2015c).
Road characteristics have long been recognized as a crucial determinant of roadkills. Factors such as traffic density and velocity, road sinuosity, and the presence of road crosses and elevation changes are frequently associated with collision risk (e.g. Trombulak and Frissell 2000; Forman et al. 2003; Clevenger et al. 2003; Malo et al. 2004; Seiler 2005; Gomes et al. 2009; Langen et al. 2012; Zuberogoitia et al. 2014; D’Amico et al. 2015). Further, roadside strips of vegetation and land-use adjacent to roads may influence roadkill risk by determining the presence and movements of animals (e.g. Forman et al. 2003; Clevenger et al. 2003; Malo et al. 2004; Grilo et al. 2009; Gunson et al. 2011).
Mitigation measures commonly used to reduce animal roadkill include, among others, wildlife crossing structures (e.g. underpasses and overpasses), warning signs, animal detection systems and a variety of fences (reviewed in Glista et al. 2009; van der Grift et al. 2013). However, none of these measures has been fully effective in preventing WVCs, since their effectiveness strongly depends on the interplay between particular life history traits (e.g. foraging strategy, dispersal or migratory movements) of the species affected by roadkills and environmental factors influencing collision risk. Given that the implantation of mitigation measures along the entire road network is economically and logistically unfeasible, and that WVCs are typically clustered (Gunson et al. 2011), identifying the factors that increase the risk of collision is essential to implement effective mitigation measures (Gunson et al. 2011). Furthermore, recent calls have highlighted the need to conduct additional research that broadens the taxonomic, spatial, and temporal scale of roadkill data sets to optimize the implementation of the mitigation measures (van der Ree et al. 2015c).
In this study, we investigated the fine-scale determinants of roadkill probability in small and medium-sized birds and mammals at a large spatio-temporal scale. Over 22 months, we regularly surveyed 45 road sections of 10 km each distributed across Andalusia (South Spain), an extensive Mediterranean region (87,268 km2) located in a biodiversity hotspot (Myers et al. 2000). During the surveys, we characterized site-specific attributes at both WVC and control points (i.e. randomly-generated points without casualties recorded), using a landscape-level and road level approach (Table 1). This information was used to investigate the WVC risk in relation to the species’ biology, adjacent land use, and road design based on the predictions presented in Table 1.
Material and methods
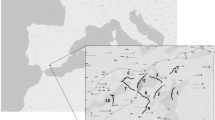
The study was conducted during two consecutive periods (July 2009–June 2010—except September and October—and April 2011–March 2012) in the Autonomous Community of Andalusia, Spain, a region that stretches from the southeast to the southwest of the Iberian Peninsula (Fig. 1). The ecosystems in Andalusia, characterized by an extraordinary diversity of species and landscapes, are considered to be highly sensitive to global-change drivers and are thus predicted to experience dramatic biodiversity changes in the next decades (Myers et al. 2000).

Situation of the road sections and main ecological units (ecoregions) surveyed during the study period. Surveyed roads (roadkill and control points) are highlighted in black
Briefly, the climate in Andalusia is Mediterranean, but due to a marked interannual variation in rainfall (it may varies from 170 mm/year to more than 1800 mm/year) and a wide elevation range (from sea level to approximately 3500 m.a.s.l.), there is a high diversity in vegetation and landscape conditions (including semiarid zones, forest, mountains or marshland). To capture such environmental diversity, the study region was divided into five ecoregions, defined as areas characterized by similar landscape characteristics and environmental conditions (GIASA et al. 2006). For a detailed description of characteristics and environmental conditions on these ecoregions see Canal et al. (2018) and GIASA et al. (2006).
Besides environmental conditions, the selection of the sampling roads aimed at representing the road network in Andalusia. Thus, according to the physic characteristics of the roads (number of lines, speed limit or traffic density), we grouped the surveyed roads into three categories: (1) Highways characterized by a dual carriageway and 120 km/h speed limit; (2) National roads, including all roads belonging to the State Network, Regional and Interregional network except highways and (3) Local roads, all roads belonging to Complementary Regional Network and the Provincial Councils. Both national and local roads are characterized by a single carriageway and a 90 km/h speed limit.
Nine road sections (three replicates per road category) were randomly selected from the road network crossing each of the five ecoregions (3 replicates × 3 road categories × 5 ecoregions = 45 road sections). For each road, a random number was generated to set the starting point (kilometric point) of the sampling stretch. Overall, we monthly surveyed 450 km along 45 road segments crossing all ecoregions included in Andalusia (Fig. 1 and Table 1 in Supplementary material).
Monthly surveys were carried out by two experienced observers by driving a vehicle at low speed (~ 25–30 km/h) along the shoulder of the road with the emergency lights flashing. The sampling order of the surveyed sections was set at random from month to month and survey session. Roadkilled animals encountered on the paved road or the road verge were identified at the species level (whenever possible) and its location was recorded using a GPS. All carcasses were removed from roads to avoid duplicating records during posterior surveys. At each point, we recorded site-specific attributes of roads and their immediate vicinity, including structures potentially influencing animal accessibility to roads (see Table 1 for a description of the measured variables and its predicted effect on WVC). Traffic density of the surveyed roads, defined as the average number of vehicles per day, was obtained from official data at http://www.fomento.es/MFOM/LANG_CASTELLANO/DIRECCIONES_GENERALES/CARRETERAS/TRAFICO_VELOCIDADES/MAPAS/ and http://www.juntadeandalucia.es/fomentoyvivienda/portal-web/web/areas/carreteras/aforos. Twenty control points were randomly selected as outlined above (without previous knowledge of roadkill points) within each of the sampling sections (20 × 45 road sections = 900 control points) and characterized following the same procedure as for the collision points.
Statistical analyses
We used Generalized Linear Mixed Models (GLMM) to model the probability of WVC in relation to landscape and road features. Separate GLMMs were fitted for birds and mammals (see below) according to the noticeable differences between their life-history strategies (e.g. spatial ecology or locomotor capacity). Even though the classification of roadkill species at the class level might appear simplistic due to major species-specific differences in life history traits, such a classification may broaden the applicability of the mitigation measures derived from our survey. Further support for the use of a coarse-grained approach comes from the common implementation of similar mitigation measurements for different animal groups. For instance, similar measures are applied for ground birds and large terrestrial mammals, whereas the same applies for bats and flying birds (Abbott et al. 2015; Kociolek et al. 2015). Finally, an analysis of the probability of WVC at a lower taxonomic level (higher functional similarity) including passerines, carnivores and lagomorphs, the groups most affected by roadkills, showed similar results to those found at the class level (see Table 3 in Supplementary material for details).
GLMMs analyzing the probability of WVC separately in birds and mammals included the presence (1) or absence (0, control points) of collision as the dependent variable (binomial distribution and logit link function) and seven explanatory variables as descriptors of each point: road type, distance to the nearest curve and its quadratic term, maximum height of roadside vegetation and its quadratic term, adjacent land use, and type of embankment. Road ID, nested within the ecoregion, was fit as a random factor.
Based on exploratory analyses, the multiple levels of road embankments was reduced to two classes: roads with embankments in any of the road sides instigating birds to fly high above the road (e.g., steep, buried sections) and those allowing animals to fly close to the road surface (e.g. roads sites at ground level). Similarly, for mammals, we reduced the type of road embankments to four classes: roads at ground level, raised, buried and roads with opposing types of embankment at each side (e.g. buried on the right side and raised on the left side).
Note that traffic density was not included in the two models above as the values for roadkill and control points within a given road section would be the same. Thus, to test if the accumulated number of roadkills in a road section was related to traffic density, we used a Pearson correlation.
Special considerations for fences and center medians
Because all highways in Spain are fenced, the effect of fences on vertebrate roadkills could only be investigated using data from secondary and local roads. The latter roads, however, lack median centers and thus, the influence of this structure on WVC was explored using exclusively data from highways.
Road points were categorized into four classes according to the presence and type of structures preventing the access of wildlife to roads (unfenced points, presence of barbed fences, wire mesh fences and walls; Table 1). The presence and type of barrier (fence/wall) was recorded at both roadsides and the difficulty of accessing roads was then determined according to the roadside having the less restrictive type of barrier. Exploratory analyses revealed no differences between mesh fences and walls in roadkill likelihood, and thus the difficulty of wildlife to access roads was assessed using a three-level variable: easy (unfenced point), medium (barber fenced) and high (mesh fences and walls) difficulty of access.
Median strips were initially categorized as Jersey barriers and structures with absent, medium or much vegetation (Table 1). However, frequency diagrams of the types of median strips revealed that the number of points with Jersey barriers as medians was very small. These points were therefore excluded from the models to avoid a disproportionate influence of rare categories on model outputs and, consequently, only medians varying in the amount of vegetation cover were analyzed.
The influence of barriers (fence/walls; except for highways) and medians (only in highways) on roadkill likelihood was analyzed separately for mammals and birds using GLMMs with the same structure as described above.
Model selection
In total, we fitted three GLMMs per vertebrate class analyzing the probability of roadkill in relation to (i) the landscape and road attributes, (ii) the type of barriers and (iii) the amount of vegetation cover in the median strips.
Selection of the final models-i.e. containing only statistically significant terms- was carried out by sequentially dropping non-significant terms from fully saturated models (containing all main effects and interactions) in a hierarchical way, starting with the least significant order terms. To confirm whether the inclusion of a predictor was significantly informative, we compared the models including and excluding the focal term using Chi square likelihood ratio tests (through maximum likelihood estimations).
We systematically performed model diagnostics statistics to avoid misleading conclusions based on statistical artifacts. Accordingly, we visually checked assumptions about the distribution of residuals through diagnostics plots, and examined collinearity and the existence of influential data points. To meet statistical assumptions, the distances to the nearest curve and traffic intensity were log10-transformed. After these transformations, diagnostics analyses did not show obvious deviation from GLMM assumptions.
Our dataset was unbalanced since twenty control points were systematically recorded per road section, whereas the number of recorded roadkill varied among roads (Fig. 2). Even though the accuracy of binomial models is robust to unbalanced sampling (Crone and Finlay 2012), we repeated the analyses above after creating a balanced dataset (roadkill and control points were randomly selected) to check for consistency between the results based on balanced and unbalanced samples. Because the results obtained using the balanced and raw datasets were similar, we present along the paper the models using the whole dataset to make full use of the available data as suggested by Crone and Finlay (2012). During our surveys, we found a small fraction of domestic mammals (mostly dogs; see Canal et al. 2018) killed by vehicles. Results from the analyses excluding and including domestic animals were qualitatively similar. For this reason, the results of the analyses including the whole dataset are presented here.

Number of recorded bird (left) and mammal (rigth) roadkills in relation to traffic density (average number of vehicles per day) of the roads
Statistical analyses were carried out in R 3.5.1 (R Core Team 2018). For running the GLMMs, we used the packages lme4 (Bates et al. 2014), lmerTest (Kuznetsova et al. 2017) and Rcpp (Eddelbuettel and François 2011). For a part of the model diagnostics, we used the package DHARMa (Hartig 2018) and the VIF function from the package car (Fox and Weisberg 2011).
Results
A total of 835 mammals and 555 birds belonging to 19 and 70 species, respectively, were recorded as killed by vehicles during the two study seasons (Tables 1 and 2 in Supplementary material; Canal et al. 2018). 2.8% of roadkills could not be identified at the species level due to severe damage and/or poor conservation status.
Road mortality analyses revealed common factors associated with the occurrence of roadkills in birds and mammals (Tables 2 and 3). In both groups, roadkills were related to traffic density (mammals: R = 0.57, P < 0.001; birds: R = 0.39, P = 0.01; Fig. 2), the distance to the nearest curve, and the height of the roadside vegetation. For the latter two factors, roadkill probability showed an inverted-U shape, increasing until a maximum distance and height, and decreasing afterwards (Tables 2, 3 and Fig. 3). Also, in both groups, roadkill risk was affected by the adjacent land use type and the slope of road embankments. Roads crossing forests showed the highest probability of roadkill in birds (Table 2 and Fig. 4), whereas, in mammals, forests and farmlands were the habitats with highest mortality rates (Table 3 and Fig. 4). The presence and type of road embankments also affected roadkill risk. In mammals, the probability of roadkill was lowest in elevated road sites, whereas roads with embankments boosting “high-altitude” flights (e.g. buried roads sections) reduced roadkill probability in birds (Tables 2, 3).

Probability of vehicle collision in birds (left) and mammals (right) according to height of roadside vegetation (upper figures) and distance to the nearest curve (lower figures). Grey dots are predicted values, the solid line denotes the fitted response of GLMMS and dashed lines show the 95% confidence intervals. Distances to the nearest curve were log transformed to meet normality assumptions

Probability of vehicle collision in birds (upper) and mammals (bottom) according to type of habitat surrounding the road. Boxplots show the extreme of the lower whisker, the lower hinge, the median, the upper hinge, and the extreme of the upper whisker. Dots are data points that lie beyond the extremes of the whiskers. Only significant P-values (< 0.05) from post hoc Tukey tests are shown
The presence and type of physical structures (fences and walls) preventing access of wildlife to roads reduced the roadkill likelihood in mammals and birds. In both taxa, the number of casualties decreased as the difficulty of accessing roads increased from unfenced points, followed by barber fences and points having mesh wire fences or walls (mammals: estimate (SE) − 0.57 (0.225), Z = − 2.53, P = 0.011; birds: estimate (SE) − 0.587 (0.185), Z = − 3.16, P = 0.002; Fig. 5). Roadkill risk also decreased as vegetation cover in the median strips increased (mammals: estimate (SE) − 0.330 (0.184), Z = − 1.79, P = 0.07; birds: estimate (SE) − 0.57 (0.17), Z = − 3.345, P < 0.001; Fig. 5).

Probability of vehicle collision in birds (left) and mammals (right) according to type of fence (upper figures) and amount of vegetation cover in the median strips (lower figures). Boxplots show the extreme of the lower whisker, the lower hinge, the median, the upper hinge, and the extreme of the upper whisker. Dots are data points that lie beyond the extremes of the whiskers
Discussion
Based on a large-scale survey and accurate description of the sampling sites, we have shown that roadkill risk in small and medium-sized birds and mammals may vary at exceedingly small spatial scales and that collision risk is group-specific. A fine-scale description of the road attributes at both roadkill and random points allowed us to unravel the road characteristics (e.g. steep embankments at roadsides and fences) determining the risk of WCV in birds and mammals. Other factors like the adjacent landscape matrix, the roadside vegetation, and vegetation density in center medians also contributed to determine roadkill probability.
Road related features
Traffic density is one of the most important predictors of roadkills (e.g. Clevenger et al. 2003; Seiler 2005; Barrientos and Bolonio 2009; Zuberogoitia et al. 2014; Gagné et al. 2015), although its influence on mortality is often non-linear; i.e. mortality peaks occur at intermediate levels of traffic density because animals are reluctant to cross highly transited roads (Madsen et al. 2002; Seiler 2005; Zuberogoitia et al. 2014). In our survey, the number of roadkills was associated with traffic density, but we did not found the expected reduction in mortality at high traffic density, even when we surveyed roads with enormous levels of traffic. At least two factors might explain the lack of a non-linear relationship between WVCs and traffic density. First, there might be a mismatch between the levels of traffic density and animal activity, since traffic density on the surveyed roads may be high only during the day, and many roadkilled species, especially mammals (see Table 2 in the Supplementary material), are most active during the night. Second, although often having a deterrent effect, traffic noise or lighting may also attract some bird species to roadsides increasing their mortality rates (Blackwell et al. 2015; Kociolek et al. 2015).
The influence of road topography in the WVC risk was in agreement with our predictions. For birds, collision risk decreased in road sections with steep buried or elevated roadside embankments as opposed to those at the ground level or with soft slopes. Possibly, flat roads enable birds to fly close to the ground while crossing, thereby increasing collision risk, whereas the reverse is likely true if steep embankments are present (Clevenger et al. 2003; Kociolek et al. 2015). Note, however, that the effect of topography (as well as that of road characteristics; see below) may be species-specific and/or conditional on other factors. For example, car lights may dazzle nocturnal birds and increase their susceptibility to WVC or predators (Blackwell et al. 2015; Kociolek et al. 2015). Further, scavengers (e.g. raptors) attracted to roads for foraging on roadkilled animals or species typically showing low-flight behaviors (e.g. owls; Massemin and Zorn 1998) may be particularly vulnerable to traffic mortality, and such susceptibility may in turn be increased or diminished by the type of road embankments. For mammals, raised road points and those at the ground level showed, respectively, the highest and the lowest roadkill rates. These results suggest that roads with steep slopes at the roadside may discourage mammals from crossing (Alexander and Waters 2000; Clevenger et al. 2003; Malo et al. 2004; Gunson et al. 2011). Elevating roads may therefore be a good option to mitigate roadkills of small- and medium-sized mammals, especially when combined with other elements such as fences or crossing structures (Clevenger et al. 2003; Malo et al. 2004; Glista et al. 2009; Gunson et al. 2011).
The distance to the nearest curve, as determining the trade-off between improved visibility (reduced WVC risk) and increased velocity (increased WVC risk), was another important predictor of roadkill in mammals and birds. Given that vehicles must decelerate as approaching a curve, the quadratic effect of the distance to the curve on roadkill probability found in our study can be reasonably expected. Non-linear relationship between proximity to the nearest curve and roadkill risk has been previously reported in other studies (Table 1), although the distance with the highest risk of roadkill varies widely among them, possibly due to a number of additional factors (e.g. presence of dense roadside vegetation, type of road and focal species) influencing the likelihood of roadkill (Malo et al. 2004; Ramp et al. 2005; Zuberogoitia et al. 2006; Grilo et al. 2009; Gunson et al. 2011).
The presence of roadside barriers (fences with varying mesh sizes and walls) also shaped mortality risk in birds and mammal; so the presence of walls or fences with small mesh size that difficult the access to roads minimized roadkill risk. Our findings are in agreement with previous works suggesting that, overall, these mitigation measures may be effective in reducing roadkills (Gunson et al. 2011; van der Grift et al. 2013; van der Ree et al. 2015a), but at least two considerations should be taken into account. First, in the case of terrestrial vertebrates (mammals, amphibians and reptiles) fences may act as barriers hampering wildlife (pre-breeding and/or dispersal) movements, thus reducing connectivity between populations (Trombulak and Frissell 2000; Forman et al. 2003; Coffin 2007). Second, as reiterated in the literature (Glista et al. 2009; van der Grift et al. 2013; D’Amico et al. 2015), the use of barriers as a mitigate measure to prevent wildlife access to roads should ideally be combined with other measures, such as underpasses and scape structures, to keep permeability between populations and thus avoid the fatal consequences of trap-effects (Colino-Rabanal et al. 2011; Cserkész et al. 2013; Zuberogoitia et al. 2014; van der Ree et al. 2015a).
The influence of median strips on roadkill risk has been scarcely assessed (Bellis and Graves 1971; Clevenger et al. 2003; Clevenger and Kociolek 2013), even when these structures may have a critical effect on WVC (reviewed in Clevenger and Kociolek 2013). Medians are usually covered by dense vegetation that may provide relatively undisturbed breeding habitat, food resources (depending on the vegetation composition; Kociolek et al. 2015), and concealment from predators and can therefore attract many animals (Adams 1984; Clevenger and Kociolek 2013). Medians may thus increase roadkill risk by increasing wildlife presence and movements around roads (Bellis and Graves 1971; Clevenger et al. 2003; Clevenger and Kociolek 2013). By contrast, we have found that the roadkill rate of birds and mammals (except lagomorphs; see Supplementary material) decreased as the amount of vegetation cover in the medians increased. Several factors may explain these results. Perhaps, in our study area, the composition and/or structure of vegetation in the median strips are not suitable as a foraging or breeding site. Densely vegetated medians might also function as an obstacle for crossing birds, encouraging them to fly high (see above) and thus avoid potential collisions (Kociolek et al. 2015). In addition, the specific requirements of the species affected and the synergistic effect of medians and microhabitat attributes might explain the apparent discrepancies between studies. For example, rabbits (the lagomorph most frequently found roadkilled during surveys) predominantly use the roadside vegetation and embankments as a refuge (Planillo and Malo 2013, 2018), which might explain the lack of relationship between vegetation cover in the medians and the probability of roadkill in this group. Regardless of the determinants of collision risk, our findings provide invaluable information about the effects of medians on WVC given the limited knowledge on this topic (Clevenger and Kociolek 2013). Further research (e.g. testing the impact of continuous and discontinuous strips of median cover on different vertebrate groups and/or their effect if combined with other crossing structures) is needed to better understand the effect of these linear developments on animal movement and mortality (Clevenger and Kociolek 2013).
Landscape features
Bird roadkills were more likely to occur on roads with adjacent wooded areas, perhaps because wooded sites offer lower visibility in relation to more open habitats, as scrubland and farmlands (Clevenger et al. 2003). Dense tree cover may at the same time increases bird abundance around roads, as they often use trees as foraging and nesting sites. Indeed, the abundance of a species in the road surroundings was likely a major determinant of their roadkill rate since, although no data on local bird abundances are available (see below), top ranked species recorded in our study are among the most common species in Andalusia (e.g. Passer domesticus, Turdus merula, Sylvia atricapilla or Erithacus rubecula). In mammals, the highest rates of fatalities occurred in forested areas, but also in points adjacent to farm areas. These findings are in agreement with other reports showing that mammal casualties increased in forested areas (Clevenger et al. 2003; Malo et al. 2004; Ramp et al. 2005; Seiler 2005; Grilo et al. 2009; Gunson et al. 2011). Moreover, the influence of landscape on the roadkill risk may depend on species-specific habitat preferences (Gunson et al. 2011) and, in our survey, mammal mortality was dominated by wild rabbits and European hares (see Canal et al. 2018), typically associated to open and/or farm areas.
For birds and mammals, roadkill risk increased when the roadside vegetation was either very tall or very short. Short roadside vegetation -or lack thereof- may reduce WVC by increasing the reaction capacity of drivers and animals to dodge the collisions. On the contrary, by providing protection or food, medium-sized vegetation at the roadside such as small trees and shrubs may attract individuals to roads and, subsequently, influence the probability of roadkill (Barrientos and Bolonio 2009). Small trees and shrubs may also increase collision rates by, for example, favoring low-to-ground-level flights while crossing roads (Clevenger et al. 2003; Ramp et al. 2006), especially in narrow roads (personal observation) or when central median with scarce or no vegetation are present (see above). Furthermore, the roadside cover in the study area consists of lush plants, such as Pistacia lentiscus, Rubus ulmifolius, Arbutus unedo, which contribute to reduce visibility and, consequently, increases roadkill probabilities. In fact, roadside management (e.g. regular cutting and removal of dense vegetation) has proven an effective mitigation measure in diverse carnivores (Trombulak and Frissell 2000; Grilo et al. 2009) and birds (Kociolek et al. 2015). For non-flying mammals, it is not surprising that the influence of vegetation height on collision risk decreased, after a threshold. In birds, however, tall vegetation should encourage high flights to cross roads, thereby reducing the probability of vehicle collision as vegetation height increases (Clevenger et al. 2003; Ramp et al. 2006). Not surprisingly, adding fences/walls adjacent to dense vegetation sites has proven an effective mitigation measures in birds and bats (Kociolek et al. 2015). However, as discussed above (see “road related features”), it is important to ensure that those barriers do not entail additional, undesirable impacts on wildlife, such as collision (as may occur if walls are made of clear glass) or insurmountable barriers to movement (Kociolek et al. 2015).
Potential limitations of the study
Field effort in terms of road distance covered and sampling frequency can strongly influence the accuracy of roadkill counts, because roadkills may be clustered in time and space and several biotic (scavengers) and abiotic (rainfall) factors may affect carcass persistence (Guinard et al. 2012; Teixeira et al. 2013; Barrientos et al. 2018). Thus, it is possible that the total number of casualties for some species is underestimated by monthly sampling (Teixeira et al. 2013). However, this should not be an issue because an accurate estimate of the number of road casualties is not the primary aim of this study; rather, our goal was to investigate the landscape and road features underlying the probability of roadkill. Our conclusions concerning the determinants of roadkill probability are unlikely to be biased by the sampling strategy, since we randomly alternated the road surveys (i.e. were randomly conducted in relation to the ecoregion, weather, type of road, and their fine-scale characteristics) and, therefore, there is no reason to think that roadkills passed systematically unnoticed at the most risky points, and viceversa.
During the first year of study, survey effort during autumn months was comparatively reduced due to logistic issues. This might have affected the roadkill estimates, for example, by decreasing the detection probability of those species that are most active or abundant during autumn, such as migratory birds. However, despite this potential inaccuracy, autumn peaks of mortality for birds and mammals clearly emerged from our survey and, importantly, the composition and temporal distribution of roadkills (see Canal et al. 2018) are in line with those found in surveys conducted at a shorter sampling periodicity (weekly or fortnightly) in the Iberian Peninsula (e.g. Frias 1999; Grilo et al. 2009; Garriga et al. 2012; Zuberogoitia et al. 2014; D’Amico et al. 2015). Thus, we are confident that our results were not qualitatively affected by the lesser survey effort performed during the autumn months of the first year of study.
Finally, due to the large spatial scale and range of taxa covered by our survey, local estimates of animal abundance and movements could not be obtained for the entire study region, and their potential effects could not be accounted for as suggested by many authors (Fahrig and Rytwinski 2009; Gunson et al. 2011; van der Ree et al. 2015b; D’Amico et al. 2015). Nonetheless, to partially control for this limitation, the characteristics of the landscape (e.g. land use) adjacent to roads were considered in our analyses, as they often influence animal distribution, abundance, and movements (D’Amico et al. 2015). Future confirmatory studies should explicitly account for these variables when developing models on WVC risk (van der Ree et al. 2015b).
Conclusions
Data on wildlife roadkills were collected at an unusually large temporal (22 moths) and spatial (regional) scale, providing stronger inferences of the patterns detected. Fine-grained characterization of road and adjacent landscape characteristics allowed the identification of important factors determining collision risk in small-to medium-sized mammals and birds. Given that roadkill risk may vary at very small spatial scales, we highlight the importance of assessing collision risk based on site-specific attributes and not uniquely on geographic information systems. Overall, reduced traffic density, steep roadside embankments, and structures hampering road access substantially reduced roadkills. By contrast, the effect of other predictors, such as land use adjacent to roads or the presence of curves, varied between vertebrate groups. It was also evident from our analyses that roadkill risk actually reflects the interplay between different variables. Hence, we suggest that future studies should focus on assessing the effect of particular predictors in road sections with no or little variation in other influential factors e.g. by assessing the effect of different median designs at sites showing similar roadside and landscape characteristics. Further research addressing the impact of medians on wildlife movement and mortality is urgently required because, despite their widespread use, the actual conservation impact of medians remains unclear.
References
Abbott I, Berthinussen A, Stone M et al (2015) Bats and roads. In: van der Ree R, Smith D, Grilo C (eds) Handbook of road ecology. Wiley, Oxford, pp 290–299
Adams LW (1984) Small mammal use of an interstate highway median strip. J Appl Ecol 21:175. https://doi.org/10.2307/2403045
Alexander SM, Waters NM (2000) The effects of highway transportation corridors on wildlife: a case study of Banff National Park. Transp Res C 8:307–320. https://doi.org/10.1016/S0968-090X(00)00014-0
Barrientos R, Bolonio L (2009) The presence of rabbits adjacent to roads increases polecat road mortality. Biodivers Conserv 18:405–418. https://doi.org/10.1007/s10531-008-9499-9
Barrientos R, Martins RC, Ascensão F et al (2018) A review of searcher efficiency and carcass persistence in infrastructure-driven mortality assessment studies. Biol Conserv 222:146–153. https://doi.org/10.1016/j.biocon.2018.04.014
Bates D, Mächler M, Bolker B, Walker S (2014) Fitting linear mixed-effects models using lme4. J Stat Softw. https://doi.org/10.18637/jss.v067.i01
Bellis E, Graves H (1971) Deer mortality on a Pennsylvania interstate highway. J Wildl Manag 35:232–237
Bissonette JA, Kassar CA, Cook LJ (2008) Assessment of costs associated with deer-vehicle collisions: human death and injury, vehicle damage, and deer loss. Hum-Wildl Interact 2:17–27
Blackwell B, DeVault T, Seamans T (2015) Understanding and mitigating the negative effects of road lighting on ecosystems. In: van der Ree R, Smith D, Grilo C (eds) Handbook of road ecology. Wiley, Oxford, pp 143–150
Camacho C (2014) ‘Bodyguard’ plants: predator−escape performance influences microhabitat choice by nightjars. Behav Processes 103:145–149
Canal D, Camacho C, Martín B et al (2018) Magnitude, composition and spatiotemporal patterns of vertebrate roadkill at regional scales: a study in southern Spain. Anim Biodivers Conserv 2:281–300
Clevenger AP, Kociolek AV (2013) Potential impacts of highway median barriers on wildlife: state of the practice and gap analysis. Environ Manag 52:1299–1312. https://doi.org/10.1007/s00267-013-0155-0
Clevenger AP, Chruszcz B, Gunson KE (2003) Spatial patterns and factors influencing small vertebrate fauna road-kill aggregations. Biol Conserv 109:15–26. https://doi.org/10.1016/S0006-3207(02)00127-1
Coffin AW (2007) From roadkill to road ecology: a review of the ecological effects of roads. J Transp Geogr 15:396–406. https://doi.org/10.1016/j.jtrangeo.2006.11.006
Colino-Rabanal VJ, Lizana M (2012) Herpetofauna and roads: a review. Basic Appl Herpetol 26:5–31
Colino-Rabanal VJ, Lizana M, Peris SJ (2011) Factors influencing wolf Canis lupus roadkills in Northwest Spain. Eur J Wildl Res 57:399–409. https://doi.org/10.1007/s10344-010-0446-1
Crone SF, Finlay S (2012) Instance sampling in credit scoring: an empirical study of sample size and balancing. Int J Forecast 28:224–238. https://doi.org/10.1016/j.ijforecast.2011.07.006
Cserkész T, Ottlecz B, Cserkész-Nagy Á, Farkas J (2013) Interchange as the main factor determining wildlife–vehicle collision hotspots on the fenced highways: spatial analysis and applications. Eur J Wildl Res 59:587–597. https://doi.org/10.1007/s10344-013-0710-2
D’Amico M, Román J, de los Reyes L, Revilla E (2015) Vertebrate road-kill patterns in Mediterranean habitats: who, when and where. Biol Conserv 191:234–242. https://doi.org/10.1016/j.biocon.2015.06.010
Eddelbuettel D, François R (2011) Rcpp seamless R and C++ integration. J Stat Softw. https://doi.org/10.18637/jss.v040.i08
Erickson W, Johnson G, Young DJ (2005) A summary and comparison of bird mortality from anthropogenic causes with an emphasis on collisions. In: John RC and Rich TD (eds) Bird conservation implementation and integration in the Americas: proceedings of the third international partners in flight conference. 2002 March 20–24; Asilomar, California, Volume 2 Gen. Tech. Rep. PSW-GTR-191. Albany, CA: US Dept. of Agriculture, Forest Service, Pacific Southwest Research Station
Erritzøe J, Mazgajski TD, Rejt Ł (2003) Bird casualties on european roads—a review. Acta Ornithol 38:77–93. https://doi.org/10.3161/068.038.0204
Fahrig L, Rytwinski T (2009) Effects of roads on animal abundance: an emperical review and synthesis. Ecol Soc 14:21–41
Forman RTT, Sperling D, Bissonette JA et al (2003) Road ecology. Island Press, Washington, DC
Fox J, Weisberg S (2011) An R companion to applied regression. Sage Publications, Thousand Oaks, CA
Frías O (1999) Estacionalidad de los atropellos de aves en el centro de España: número y edad de los individuos y riqueza y diversidad de especies. Ardeola 46:23–30
Fulton L, Eads G (2004) IEA/SMP model documentation and reference case projection. International Energy Agency (IEA)/World Business Council for Sustainable Development (WBCSD), Geneva
Gagné SA, Bates JL, Bierregaard RO (2015) The effects of road and landscape characteristics on the likelihood of a Barred Owl (Strix varia)-vehicle collision. Urban Ecosyst 18:1007–1020. https://doi.org/10.1007/s11252-015-0465-5
Garriga N, Santos X, Montori A et al (2012) Are protected areas truly protected? The impact of road traffic on vertebrate fauna. Biodivers Conserv 21:2761–2774. https://doi.org/10.1007/s10531-012-0332-0
GIASA et al (2006) Libro blanco de las carreteras y los espacios naturales protegidos de Andalucía. Consejeria de Obras Públicas y Transportes, Junta de Andalucia
Gibbs JP, Shriver WG (2002) Estimating the effects of road mortality on turtle populations. Conserv Biol 16:1647–1652. https://doi.org/10.1046/j.1523-1739.2002.01215.x
Glista DJ, DeVault TL, DeWoody JA (2009) A review of mitigation measures for reducing wildlife mortality on roadways. Landsc Urban Plan 91:1–7. https://doi.org/10.1016/j.landurbplan.2008.11.001
Gomes L, Grilo C, Silva C, Mira A (2009) Identification methods and deterministic factors of owl roadkill hotspot locations in Mediterranean landscapes. Ecol Res 24:355–370. https://doi.org/10.1007/s11284-008-0515-z
Grilo C, Bissonette JA, Santos-Reis M (2009) Spatial–temporal patterns in Mediterranean carnivore road casualties: consequences for mitigation. Biol Conserv 142:301–313. https://doi.org/10.1016/j.biocon.2008.10.026
Guinard É, Julliard R, Barbraud C (2012) Motorways and bird traffic casualties: carcasses surveys and scavenging bias. Biol Conserv 147:40–51. https://doi.org/10.1016/j.biocon.2012.01.019
Gunson KE, Mountrakis G, Quackenbush LJ (2011) Spatial wildlife-vehicle collision models: a review of current work and its application to transportation mitigation projects. J Environ Manag 92:1074–1082. https://doi.org/10.1016/j.jenvman.2010.11.027
Hartig F (2018) DHARMa: residual diagnostics for hierarchical (Multi-Level/Mixed) regression models. R package version 0.2.0. https://CRAN.R-project.org/package=DHARMa
Huijser MP, Duffield JW, Clevenger AP et al (2009) Cost-benefit analyses of mitigation measures aimed at reducing collisions with large ungulates in the united states and canada: a decision support tool. Ecol Soc 14:15. https://doi.org/10.1016/j.contraception.2009.11.002
Kociolek A, Grilo C, Jacobson S (2015) Flight doesn’t solve everything: mitigation of road impacts on birds. In: van der Ree R, Smith D, Grilo C (eds) Handbook of road ecology. Wiley, Oxford, pp 281–289
Kuznetsova A, Brockhoff PB, Christensen RHB (2017) lmerTest package: tests in linear mixed effects models. J Stat Softw. https://doi.org/10.18637/jss.v082.i13
Langen TA, Gunson KE, Scheiner CA, Boulerice JT (2012) Road mortality in freshwater turtles: identifying causes of spatial patterns to optimize road planning and mitigation. Biodivers Conserv 21:3017–3034. https://doi.org/10.1007/s10531-012-0352-9
Madsen A, Strandgaard H, Prang A (2002) Factors causing traffic killings of roe deer Capreolus capreolus in Denmark. Wildl Biol 8:55–61
Malo JE, Suárez F, Díez A (2004) Can we mitigate animal–vehicle accidents using predictive models? J Appl Ecol 41:701–710
Massemin S, Zorn T (1998) Highway mortality of barn owls in Northeastern France. J Raptor Res 32:229–232
Meyer I, Kaniovski S, Scheffran J (2012) Scenarios for regional passenger car fleets and their CO2 emissions. Energy Policy 41:66–74. https://doi.org/10.1016/j.enpol.2011.01.043
Mumme RL, Schoech SJ, Woolfenden GE, Fitzpatrick JW (2000) Life and death in the fast lane: demographic consequences of road mortality in the Florida Scrub-Jay. Conserv Biol 14:501–512. https://doi.org/10.1046/j.1523-1739.2000.98370.x
Myers N, Mittermeier RA, Mittermeier CG et al (2000) Biodiversity hotspots for conservation priorities. Nature 403:853–858. https://doi.org/10.1038/35002501
Planillo A, Malo JE (2013) Motorway verges: paradise for prey species? A case study with the European rabbit. Mamm Biol 78:187–192. https://doi.org/10.1016/j.mambio.2012.11.001
Planillo A, Malo JE (2018) Infrastructure features outperform environmental variables explaining rabbit abundance around motorways. Ecol Evol 8:942–952. https://doi.org/10.1002/ece3.3709
Ramp D, Caldwell J, Edwards KA et al (2005) Modelling of wildlife fatality hotspots along the snowy mountain highway in New South Wales, Australia. Biol Conserv 126:474–490. https://doi.org/10.1016/j.biocon.2005.07.001
Ramp D, Wilson VK, Croft DB (2006) Assessing the impacts of roads in peri-urban reserves: road-based fatalities and road usage by wildlife in the Royal National Park, New South Wales, Australia. Biol Conserv 129:348–359. https://doi.org/10.1016/j.biocon.2005.11.002
R Core Team (2018) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria
Sandberg M, Eriksson L, Jonsson J et al (1998) New chemical descriptors relevant for the design of biologically active peptides. A multivariate characterization of 87 amino acids. J Med Chem 41:2481–2491. https://doi.org/10.1021/jm9700575
Seiler A (2005) Predicting locations of moose-vehicle collisions in Sweden. J Appl Ecol 42:371–382. https://doi.org/10.1111/j.1365-2664.2005.01013.x
Teixeira FZ, Coelho AVP, Esperandio IB, Kindel A (2013) Vertebrate road mortality estimates: effects of sampling methods and carcass removal. Biol Conserv 157:317–323. https://doi.org/10.1016/j.biocon.2012.09.006
Trombulak SC, Frissell CA (2000) Review of ecological effects of roads on terrestrial and aquatic communities. Conserv Biol 14:18–30
van der Grift EA, van der Ree R, Fahrig L et al (2013) Evaluating the effectiveness of road mitigation measures. Biodivers Conserv 22:425–448. https://doi.org/10.1007/s10531-012-0421-0
van der Ree R, Gagnon J, Smith D (2015a) Fencing: a valuable tool for reducing wildlife-vehicle collisions and funnelling fauna to crossing structures. In: van der Ree R, Smith D, Grilo C (eds) Handbook of road ecology. Wiley, Oxford, pp 159–171
van der Ree R, Smith D, Grilo C (2015b) Handbook of road ecology. Wiley, Hoboken
van der Ree R, Smith D, Grilo C (2015c) The ecological effects of linear infrastructure and traffic: challenges and opportunities of rapid global growth. In: van der Ree R, Smith D, Grilo C (eds) Handbook of road ecology. Wiley, Oxford, pp 1–9
Zuberogoitia I, Martínez JE, Martínez JA et al (2006) Influence of management practices on nest site habitat selection, breeding and diet of the common buzzard Buteo buteo in two different areas of Spain. Ardeola 53:83–98
Zuberogoitia I, del Real J, Torres JJ et al (2014) Ungulate vehicle collisions in a peri-urban environment: consequences of transportation infrastructures planned assuming the absence of Ungulates. PLoS ONE 9:e107713. https://doi.org/10.1371/journal.pone.0107713
Acknowledgements
We thank Miguel A. Sanza for his dedicated assistance during the road surveys. Two anonymous reviewers provided numerous comments and suggestions that substantially improved the manuscript. This project was funded by Consejería Medio Ambiente (Junta de Andalucía). CC was supported by the Spanish Ministry of Economy and Competitiveness (SVP-2013-067686).
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by David Hawksworth.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Canal, D., Camacho, C., Martín, B. et al. Fine-scale determinants of vertebrate roadkills across a biodiversity hotspot in Southern Spain. Biodivers Conserv 28, 3239–3256 (2019). https://doi.org/10.1007/s10531-019-01817-5
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10531-019-01817-5