
Abstract
Copper oxide nanoparticles (CuO-NPs) are frequently used for industrial or medical applications and are known for their high toxic potential. As little is known so far on the consequences of an exposure of brain cells to such particles, we applied CuO-NPs to cultured primary rat astrocytes and investigated whether such particles affect cell viability and alter their metabolic properties. Astrocytes efficiently accumulated CuO-NPs in a time- and concentration-dependent manner. The cells remained viable during a 24 h incubation with 100 µM copper in the form of CuO-NPs, while higher concentrations of CuO-NPs severely compromised the cell viability. Astrocytes that were exposed for 24 h to 100 µM CuO-NPs showed significantly enhanced extracellular lactate concentrations and increased cellular levels of glutathione and metallothioneins. The CuO-NP-induced increase in lactate release and metallothionein content were prevented by the presence of the membrane-permeable copper chelator tetrathiomolybdate, while this chelator increased already in the absence of CuO-NPs the cellular glutathione content. After removal of the CuO-NPs following a 24 h pre-incubation with 100 µM CuO-NPs, astrocytes maintained during a further 6 h incubation an elevated glycolytic lactate release and exported significantly more glutathione than control cells that had been pre-incubated without CuO-NPs. These data suggest that copper ions which are liberated from internalized CuO-NPs stimulate glycolytic flux as well as the synthesis of glutathione and metallothioneins in cultured viable astrocytes.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Copper oxide nanoparticles (CuO-NPs) have interesting physical and catalytic properties and are therefore frequently used as additives for industrial products such as paints, lubricants, heat-transfer fluids, conductive films, plastics and solid propellants [1–4]. In addition, due to their potent antimicrobial and biocidal properties CuO-NPs are used in medical products such as bandages and water filters [4–6]. The most important routes of exposure to CuO-NPs are inhalation, ingestion or skin contact [7]. The risk of occupational exposure to substantial amounts of copper-containing NPs is high for welders [8] and for workers operating universal electric motors [9].
CuO-NPs are considered as very toxic compared to other metal oxide nanoparticles [10]. Exposure of cells to CuO-NPs has been reported to cause inhibition of enzymes, expression of proinflamatory molecules and alterations in gene expression [1, 11, 12] as well as the activation of autophagic and apoptotic processes [12–15]. The generation of reactive oxygen species (ROS) and oxidative stress-mediated damage are considered to be involved in the observed cell toxicity of CuO-NPs [16, 17].
Wistar rats injected with CuO-NPs suffered from severe cognitive impairments including decreased learning and memory abilities [18]. To reach the brain, CuO-NPs have to cross the blood–brain barrier (BBB) which has been demonstrated for several types of inorganic nanoparticles including CuO-NPs [19–23]. In addition, CuO-NPs and other inorganic nanoparticles can reach the brain via nerve axons from the olfactory bulb [24–27].
The first cells in brain that encounter nanoparticles which have crossed the BBB from blood are astrocytes which are located between the endothelial cells of the vascular system and neurons [28]. Astrocytes have several essential homeostatic functions in the brain. These cells supply metabolites and gliotransmitters for neuronal function, protect the brain environment from toxins and oxidative stress and regulate the metal homeostasis of the brain [29–32]. Astrocytes have been suggested to play a key role in the copper metabolism of the brain [31, 33]. At least in culture, astrocytes efficiently accumulate, store and export ionic copper [34, 35]. However, an exposure to copper does also alter metabolic properties of astrocytes as copper accumulation in cultured astrocytes is accompanied by accelerated glycolytic flux and stimulated synthesis of glutathione (GSH) [36, 37].
Several studies have shown that cultured astrocytes efficiently accumulate silver and iron oxide nanoparticles and that these types of NPs have little toxic potential to astrocytes (for overview see: [32, 38]). In contrast, exposure of astrocytes to CuO-NPs has recently been show to acutely increase cellular ROS production and to severely damage cultured astrocytes most likely by copper-mediated oxidative stress [17]. As it is currently not known whether CuO-NPs in sub-toxic concentrations may affect metabolic properties of astrocytes, we have exposed cultured primary astrocytes to CuO-NPs and investigated the consequences of such an exposure on cell viability, glycolytic flux as well as the cellular contents of GSH and metallothioneins (MTs).
Materials and Methods
Materials
Fetal calf serum (FCS) and penicillin/streptomycin solution were obtained from Biochrom (Berlin, Germany) and Dulbecco’s modified Eagle’s medium (DMEM) from Gibco (Karlsruhe, Germany). Bovine serum albumin (BSA), sulfosalicylic acid, dithiothreitol, magnesium chloride hexahydrate, NADPH, NADP+, NAD+ and NADH were from Applichem (Darmstadt, Germany). Glutathione reductase, ATP, hexokinase, glucose-6-phosphate dehydrogenase, lactate dehydrogenase and glutamate pyruvate transaminase were purchased from Roche (Mannheim, Germany). The monoclonal mouse anti-MT antibody (clone E9; detects MTs 1 and 2) was purchased from Dako (Hamburg, Germany). The peroxidase-conjugated donkey anti-mouse antibody and the monoclonal mouse anti-actin antibody (clone Ab-5) were purchased from Dianova (Hamburg, Germany). Page ruler prestained protein ladder 10–170 kDa and hydrochloric acid were from Thermo Fisher Scientific (Fair Lawn, USA), Tween®20 from Serva (Heidelberg, Germany) and trichloroacetic acid from Jannsen (Geel, Belgium). All other chemicals were purchased at analytical grade from Merck (Darmstadt, Germany), Fluka (Buchs, Switzerland), Roth (Karlsruhe, Germany) or Sigma-Aldrich (Steinheim, Germany). Sterile 24-well cell culture plates and unsterile 96-well microtiter plates were from Sarstedt (Nümbrecht, Germany).
Primary Astrocyte Cultures
Astrocyte-rich primary cultures were prepared from brains of newborn Wistar rats as previously described [39, 40] by seeding 300,000 viable cells in wells of a 24-well culture plate in 1 mL culture medium (90 % DMEM containing 25 mM glucose, 10 % FCS, 18 U/mL penicillin G, 18 µg/mL streptomycin sulphate). The cultures were maintained in presence of 10 % CO2 in the humidified atmosphere of a Sanyo incubator (Osaka, Japan). Immunocytochemical staining for the astrocyte marker glial fibrillary acidic protein confirmed that these cultures contain predominately astrocytes [40, 41]. The culture medium was renewed every 7 days. Confluent cultures of an age between 14 and 28 days were used for the experiments described here.
Synthesis and Characterization of Copper Oxide Nanoparticles
CuO-NPs were synthesized by alkaline precipitation of copper nitrate and coated with 2,3-dimercaptosuccinic acid as described recently [17]. These particles are agglomerates of crystalline primary particles which have an average size of approximately 5 nm. Dispersed in culture medium CuO-NPs have a mean hydrodynamic diameter of 178 ± 12 nm and a ζ-potential of −10 mV [17]. A dispersion of CuO-NPs (30 mM copper) in water was diluted in culture medium to the final particle concentrations. The concentrations of CuO-NPs given here represent the concentration of total copper in a given incubation medium and not the concentration of particles.
Experimental Incubations
The cultures were washed twice with 1 mL prewarmed (37 °C) phosphate-buffered saline (PBS; 10 mM potassium phosphate buffer, pH 7.4, containing 150 mM sodium chloride) and subsequently incubated for up to 24 h at 37 °C in an incubator with 10 % CO2 in 1 mL culture medium containing CuO-NPs and/or other compounds in the concentrations given in the legends of the figures and tables. Ammonium tetrathiomolybdate (TTM) was prepared as 10 mM stock solution in water and diluted in culture medium to a final concentration of 100 µM.
To investigate the persistence of CuO-NP-induced metabolic alterations, astrocytes were preincubated for 24 h in 1 mL culture medium in the absence or presence of 100 µM CuO-NPs, washed twice with prewarmed (37 °C) PBS and subsequently incubated at 37 °C (in the humidified atmosphere of an incubator without CO2 supply) in 1 mL incubation buffer (IB; 20 mM HEPES, 5 mM glucose, 1.8 mM CaCl2, 1 mM MgCl2, 5.4 mM KCl, 145 mM NaCl, adjusted to pH 7.4) containing 100 µM of the membrane-impermeable copper chelator bathocuproine disulfonate (BCS). After a given incubation period, the incubation media were harvested and the cells were washed twice with 1 mL ice cold PBS. Dry cells were stored at −20 °C for quantification of copper and protein contents.
Copper and Protein Quantification
The copper content of CuO-NP-treated cells was determined by graphite furnace atomic absorption spectroscopy using a Varian (Darmstadt, Germany) AA-240Z spectrophotometer and a Varian GTA-120 graphite tube atomizer as recently described [17, 34]. The protein content of the cells per well was determined by the Lowry method [42] using bovine serum albumin as standard protein.
Viability and Vitality of Cultured Astrocytes
Cell vitality and functionality was assessed by quantification of the capacity of the cells to reduce 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT), by determination of the cellular activity of the enzyme lactate dehydrogenase (LDH) and by propidium iodide (PI) staining. The MTT reduction capacity was quantified as described recently [17]. Briefly, after the experimental incubation the cells were washed with prewarmed PBS and incubated for additional 90 min at 37 °C with culture medium containing 0.5 mg/mL MTT. Subsequently, the cells were lysed in DMSO and the absorbance of the generated formazan was determined at 540 nm in wells of a microtiter plate by a Sunrise microtiter plate photometer (Tecan, Crailsheim, Germany). The activity of the enzyme LDH in cell lysates was measured by following the NADH consuming reduction of pyruvate to lactate as described previously [43]. The integrity of the cell membrane was determined by application of the fluorescent and membrane-impermeable dye PI [40]. The staining procedure was performed as recently described [17] with Hoechst 33342 (H33342) for nuclear staining of all cells present and with silver nitrate as positive control for cell toxicity.
Determination of Lactate, Glucose and Glutathione
The concentrations of extracellular lactate and glucose were determined by coupled enzymatic assays as described previously [40, 44]. The amount of glucose consumed was calculated as difference between the glucose content of the incubation medium harvested after a given incubation and the medium applied to the cells. The contents of total glutathione [GSx = amount of GSH plus two times the amount of glutathione disulfide (GSSG)] and GSSG in cell lysates and in incubation media were quantified by the Tietze method using a microtiter plate assay [40, 45]. For all experiments described here, the cellular GSSG levels were in the range of the detection limit of the assay used [40] and did not significantly contribute to the GSx values determined.
Western Blot Analysis
Western blots to test for the presence of MTs in astrocytes were performed as previously described [46, 47]. Briefly, after a given treatment, the cells in a well of a 24-well plate were directly lysed in 80 μL loading buffer (62.4 mM Tris/HCl, pH 6.8, 2 % (w/v) sodium dodecylsulfate, 10 % (w/v) glycerol, 0.01 % (w/v) bromphenole blue, 4 mM dithiotreitol). Lysates from three wells were pooled and stored at −20 °C. For Western blotting, the lysates were boiled for 10 min and subsequently incubated with 12.5 mM iodoacetamide for 15 min at 50 °C to mask sulfhydryl groups. Per lane of a polyacrylamide gel (4 % stacking gel, 15 % separation gel) 12.5 µg protein were loaded and separated for 1 h at 160 V. Subsequently the gel was electroblotted onto an Amersham Hybond™-C Extra nitrocellulose membrane (GE Healthcare, Munich, Germany) for 1 h at 360 mA. The membranes were washed with TBS-T (10 mM Tris/HCl, 150 mM NaCl, 0.1 % (v/v) Tween®20, pH 7.3) and subsequently unspecific binding sites were blocked for 1 h with 5 % (w/v) milk powder in TBS-T (TBS-T-MP). The membranes were incubated with the monoclonal anti-MT antibody (1:500 in TBS-T-MP) or the monoclonal anti-actin antibody (1:2,000 in TBS-T-MP) overnight at 4 °C on a Heidolph (Schwabach, Germany) Duomax 1030 shaker. After the incubation, the membranes were washed thrice for 15 min with TBS-T and subsequently incubated for 1 h with a peroxidase-conjugated anti-mouse antibody (1:20,000 in TBS-T-MP) at room temperature. After three additional washing steps, the blots were developed with the ECL Western blot detection reagents (GE Healthcare) on a Amersham Hyperfilm ECL X-ray film (GE Healthcare). The signals for iodoacetamide-conjugated MTs (12 kDa) and actin (42 kDa) were detected at the expected molecular masses.
Presentation of Data
The data shown in figures and tables represent means ± standard deviations of values derived from n experiments that were performed on independently prepared cultures. Figures showing Western blots and microscopic images are from representative experiments that were reproduced at least twice on independently prepared cultures. Statistical analysis between multiple groups of data was performed by ANOVA followed by the Bonferroni’s post hoc test, analysis of the significance of differences between two sets of data by the paired Student’s t test. p > 0.05 was considered as not significant.
Results
Accumulation and Toxicity of CuO-NPs in Cultured Astrocytes
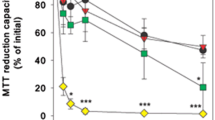
To investigate cellular uptake and toxicity of CuO-NPs, cultured astrocytes were incubated with up to 300 µM (18.9 µg/mL) of copper as CuO-NPs for up to 24 h and the MTT reduction capacity as well as the specific cellular copper contents were determined (Fig. 1). CuO-NP-treated astrocytes showed a time- and concentration-dependent increase in their specific copper contents (Fig. 1a). Within 24 h of incubation with 100, 200 and 300 µM CuO-NPs, the cells increased their specific cellular copper content from an initial value of 0.9 ± 0.4 to 19.7 ± 2.9, 61.3 ± 8.2 and 62.2 ± 7.1 nmol/mg, respectively. However, the strong increase in the copper contents of CuO-NP-treated cells was accompanied by a time- and concentration-dependent decrease in the MTT reduction capacity of the cells (Fig. 1b), which is considered as indicator for compromised cell vitality of copper-treated cells [17, 34]. While the MTT reduction capacity of astrocytes that had been incubated with 100 µM CuO-NPs did not differ significantly to that of control cells (incubation without CuO-NPs), the MTT reduction capacity of cells exposed to 200 or 300 µM CuO-NPs was found significantly impaired already after 12 and 6 h of incubation, respectively. For these conditions, the MTT reduction capacity declined further with longer incubation time and reached after 24 h of incubation values that corresponded to around 40 (200 µM) and 30 % (300 µM) of the initial values (Fig. 1b). In contrast, control astrocytes and cultures exposed to 100 µM CuO-NPs still possessed 85 % of the initial MTT reduction capacity after 24 h of incubation (Fig. 1b). Similar experiments performed with CuCl2 in micromolar concentrations revealed that exposure of astrocytes for 24 h to CuCl2 lowered the cellular MTT reduction capacity to 72 ± 3 (100 µM), 61.5 ± 4.6 (200 µM) and 49.7 ± 4.5 % (300 µM) of the control values (n = 3) and significantly increased the specific cellular copper contents to 13.7 ± 1.1 (100 µM), 17.6 ± 2.2 (200 µM) and 20.2 ± 2.1 nmol/mg (300 µM).

Effects of a treatment of astrocytes with CuO-NPs on cellular copper content, cell viability and lactate generation. The cells were incubated without or with CuO-NPs in the indicated concentrations for up to 24 h and the specific cellular copper content (a), the cellular MTT reduction capacity (b) as well as the extracellular lactate concentration (c) were determined. The data shown represent means ± standard deviations of values derived from three experiments that were performed on independently prepared cultures. Asterisks in a and b indicate the significance of differences (ANOVA) to the respective values determined for control cells (incubation in the absence of CuO-NPs) (*p < 0.05; **p < 0.01; ***p < 0.001). Hashes in c indicate the significance of differences (t test) in the extracellular lactate concentrations of astrocytes that were incubated in the absence and the presence of 100 µM CuO-NPs (# p < 0.05; ## p < 0.01; ### p < 0.001)
The concentration-dependent toxicity of CuO-NPs was confirmed by PI staining. While cultures incubated for 24 h without (Fig. 2a, b) or with 100 µM CuO-NPs (Fig. 2c, d) did hardly contain any PI-positive cells (Fig. 2a, c), a substantial number of PI-positive cells was detected after incubation of astrocytes with 200 (Fig. 2e) or 300 µM (Fig. 2g) CuO-NPs as well as after exposure to AgNO3 (Fig. 2i), which served as positive control for toxicity [48].

Cytochemical staining of CuO-NP-treated astrocytes for membrane integrity. Primary astrocyte cultures were incubated for 24 h in the absence (a, b) or the presence of CuO-NPs (c–h) in the indicated concentrations and subsequently stained for loss of membrane integrity with propidium iodide. Presence of all cell nuclei was visualized by staining with the membrane-permeable dye H33342. An incubation with silver nitrate (100 µM) for 24 h was used as positive control for impaired membrane integrity (i). The images are from a representative experiment that was reproduced twice on independently prepared cultures with similar outcome. The scale bar in a applies to all panels
Cultured astrocytes contained after 24 h of incubation with 100 µM CuO-NPs a specific cellular copper content of 19.7 ± 2.9 nmol/mg, while cells incubated in the absence of nanoparticles contained only around 1 nmol copper/mg (Table 1). Despite of this severe increase in the cellular copper content, astrocyte cultures that had been incubated for 24 h with 100 µM CuO-NPs did not significantly differ from control cells (incubated in the absence of CuO-NPs) in their cellular MTT reduction capacity (Fig. 1a; Table 1), PI-staining (Fig. 2c), in their protein content (Table 1) or their specific cellular LDH activity (Table 1). This demonstrates that cultured astrocytes are not damaged by a 24 h incubation with 100 µM CuO-NPs. Thus, this incubation paradigm was considered as suitable to investigate the consequences of an exposure to sub-toxic concentration of CuO-NPs on metabolic properties of cultured astrocytes.
CuO-NP Exposure Affects Glycolytic Flux and the Cellular Levels of GSH and Metallothioneins
To test for a potential effect of CuO-NPs on the glucose metabolism, astrocytes were incubated without or with 100 µM CuO-NPs. After 24 h of incubation, glucose consumption and lactate release by CuO-NP-treated astrocytes were found significantly increased by 23 and 24 %, respectively, compared to control cells that were incubated without CuO-NPs (Table 1). During the first hours of incubation, almost identical extracellular lactate concentrations were determined for astrocytes treated without or with CuO-NPs. However, already after 12 h of incubation the extracellular lactate concentration was significantly increased for CuO-NP-treated astrocytes compared to control cells (Fig. 1c). In contrast, the extracellular lactate concentration of astrocytes that had been incubated with toxic concentrations of 200 or 300 µM CuO-NPs for 24 h were not increased (200 µM CuO-NPs: 6.6 ± 1.2 mM) or even lowered (300 µM CuO-NPs: 4.9 ± 1.0 mM) compared to control cells (6.7 ± 1.2 mM).
Exposure of astrocytes for 24 h with 100 µM CuO-NPs significantly increased the specific cellular GSx contents by about 36 % compared to the values determined for control cells (Table 1), while the specific cellular contents of GSSG remained very low (below 2 % of the GSx values) in both control and CuO-NP-treated astrocytes (Table 1). Finally, also the cellular level of MTs was found significantly elevated compared to control cells after 24 h of incubation with 100 µM CuO-NPs (Fig. 3).

Content of metallothioneins (MTs) in astrocytes. Cultured astrocytes were incubated for 24 h without or with 100 µM CuO-NPs in the presence or the absence of 100 µM tetrathiomolybdate (TTM) and their lysates were analyzed for MTs and actin by Western blotting. a shows a representative blot, b the results of the analysis of the signal intensities for blots performed on lysates derived from experiments on four independently prepared cultures. Asterisks indicate the significance of differences (ANOVA) compared to the data of control cells (treated without CuO-NPs and TTM) (***p < 0.001)
Effects of the Copper Chelator TTM on the CuO-NP-Induced Alterations in Astrocytic Properties
To test for a potential involvement of copper ions that had been liberated from accumulated CuO-NPs in the observed effects of an CuO-NP-treatment on lactate production, glucose consumption and the cellular contents of GSx and MTs, astrocytes were incubated for 24 h with 100 µM CuO-NPs in the absence or presence of the membrane-permeable copper ion chelator TTM. Presence of TTM lowered the CuO-NP-dependent increase in specific cellular copper content by 47 % (Table 2), while the CuO-NP-induced increases in glucose consumption, lactate production and levels of MTs were completely prevented in the presence of TTM (Table 2; Fig. 3). In contrast, TTM did not affect the CuO-NP-induced increase in the specific cellular GSx contents but increased already in the absence of CuO-NPs the specific cellular GSx content (Table 2).
Test for Persistence of the CuO-NP-Induced Stimulation of Glycolytic Flux
To investigate whether the observed CuO-NP-induced alterations in astrocytic glucose metabolism depends on the presence of extracellular CuO-NPs, the cultures were pre-incubated for 24 h without or with 100 µM CuO-NPs, washed and subsequently incubated at 37 °C for 6 h in an copper-free incubation buffer containing 100 µM of the extracellular copper chelator BCS. During the 6 h main incubation the copper content of the CuO-NP-pre-incubated cells was lowered by 45 % from 19.7 ± 2.9 (Table 1) to 10.9 ± 1.1 nmol/mg (Table 3). This loss of cellular copper was accompanied by a matching increase in the extracellular copper content, as the medium harvested after the 6 h incubation contained 1.7 ± 0.3 nmol of copper which corresponds to 11.2 ± 1.1 nmol copper/mg protein. An analogous 6 h incubation of CuO-NP-pre-incubated astrocytes at 4 °C partially prevented (p < 0.05; n = 3) the loss of cellular copper (cellular content: 16.3 ± 2.1 nmol/mg) and the accumulation of extracellular copper content (5.2 ± 1.0 nmol/mg), suggesting that the reported temperature-sensitive astrocytic export of copper ions [35] is involved in the observed loss of cellular copper. However, as this copper loss was only partially prevented by lowering the incubation temperature to 4 °C, also detachment of CuO-NPs which had been extracellularly adsorbed to the cell membrane [17] is likely to contribute to this process.
The additional 6 h incubation after a 24 h pre-incubation lowered the MTT reduction capacity of CuO-NP-treated astrocytes to some extent while the cellular protein content per well was not altered compared to control cells (Table 3). In addition, PI staining of the cells after the 6 h main incubation confirmed that the membrane integrity of the CuO-NP-pre-incubated cells was not impaired (Fig. 4).

Cytochemical staining of astrocytes for membrane integrity after removal of CuO-NPs. Primary astrocyte cultures were pre-incubated without (a, b) or with 100 µM of CuO-NPs (c, d) for 24 h, washed and subsequently incubated for additional 6 h in incubation buffer containing 100 µM BCS. Subsequently the cells were stained with propidium iodide and H33342. A pre-incubation with silver nitrate (100 µM) for 24 h followed by a 6 h main incubation without silver nitrate was used as positive control for impaired membrane integrity (e, f). The images are from a representative experiment that was reproduced twice on independently prepared cultures with similar outcome. The scale bar in a applies to all panels
Astrocytes that had been pre-incubated for 24 h with 100 µM CuO-NPs maintained during a following 6 h main incubation an increased glyolytic flux. Compared to control cells that had been pre-incubated without nanoparticles, the amounts of lactate released and glucose consumed within 6 h by the cells was significantly increased by 24 and 70 %, respectively, in CuO-NP-treated cultures (Table 3) and the specific lactate release rate of these cells, as calculated from the almost linear increase in the extracellular lactate accumulation (Fig. 5a), was increased by 40 % compared to controls (Table 3).

Persistence of the stimulated lactate release and accumulation of extracellular glutathione in CuO-NP-treated astrocytes. Cultured astrocytes were pre-incubated for 24 h in the absence (Control) or the presence of 100 µM CuO-NPs. Subsequently the cells were washed and incubated for up to 6 h in copper-free incubation buffer containing 100 µM BCS. During the 6 h main incubation the extracellular contents of lactate (a) and GSx (b) were determined. The data shown represent means ± standard deviations of values derived from three experiments that were performed on independently prepared cultures. Asterisks indicate the significance of differences (t test) of data obtained for cells that had been pre-incubated without and with CuO-NPs (*p < 0.05; **p < 0.01)
Effects of a CuO-NP-Pre-incubation on GSH Export
After a 24 h pre-incubation of astrocytes with 100 µM CuO-NPs, the specific cellular GSx content was significantly increased compared to control cells (Table 1). After a subsequent 6 h main incubation without CuO-NPs the specific GSx content of cells that had been exposed to CuO-NPs remained higher than that of the respective control cells, although this difference did not reach the level of significance (Table 3). In addition, the cellular GSSG levels were very low after the 6 h main incubation for cells that had been pre-incubated without or with CuO-NPs (Table 3).
Quantification of the extracellular GSx contents revealed that GSx accumulated quicker in the incubation medium of astrocytes that had been pre-incubated with CuO-NPs compared to control cells (Fig. 5b). After 6 h of incubation the extracellular GSx content of CuO-NP-pre-incubated cultures was significantly increased by 70 % and the GSx accumulation rate of such cells was doubled compared to the respective data of control cultures (Table 3). The extracellular GSx values accounted almost exclusively for GSH, as only minute amounts of extracellular GSSG (<0.8 nmol/mg) were detectable for astrocytes that had been pre-incubated without or with CuO-NPs (data not shown).
The sum of extracellular plus cellular GSx contents determined after the 6 h main incubation without (43.6 ± 14.1 nmol/mg; n = 3) and with CuO-NPs (58.2 ± 11.9 nmol/mg; n = 3) did not differ significantly (p > 0.05) from the values determined after the respective pre-incubation without or with CuO-NPs (Table 1).
Discussion
To test for the potential of CuO-NPs to compromise the viability of astrocytes and to affect metabolic properties of these cells, we exposed primary cultured astrocytes in serum-containing culture medium to CuO-NPs for 24 h and found that astrocytes accumulated such particles in a time- and concentration-dependent manner. Astrocytes tolerated concentrations of 100 µM CuO-NPs very well, but were severely damaged after exposure to higher particle concentrations. The observed accumulation of CuO-NPs by astrocytes and the toxicity induced by elevated concentrations of CuO-NPs are in line with results obtained recently for astrocytes that had been acutely exposed for up to 6 h to CuO-NPs [17]. Compromised viability of cultured astrocytes was previously observed for conditions which elevated the specific cellular copper content within a few hours after exposure to copper salts or CuO-NPs to values of above 10 nmol/mg, while lower specific copper contents were not accompanied by a compromised viability [17, 34, 35]. This suggests that cultured astrocytes can deal well with an elevated cellular copper content as long as it does not exceed a given threshold value.
Studies on astrocytes [17] and other cell types [12, 13, 49–52] revealed that CuO-NP-treated cells suffer from an increased formation of cellular ROS. For astrocytes, elevated ROS levels and toxicity were prevented by co-application of the membrane-permeable copper chelator TTM, suggesting that copper ions which have been liberated from the accumulated CuO-NPs are responsible for the ROS-induced toxicity [17]. Indeed, TTM prevented also the impairment of cellular MTT reduction capacity and membrane integrity during a 24 h exposure of cultured astrocytes to 200 µM CuO-NPs (data not shown).
The uptake of CuO-NPs into astrocytes in serum-containing medium is likely to be mediated by endocytosis as recently described for silver and iron oxide NPs [48, 53, 54]. At least for other cell types CuO-NPs were found to be present in cellular vesicles [50, 55, 56]. Such vesicles may rapidly enter the lysosomal pathway and the CuO-NPs will then encounter an environment that facilitates the release of copper ions from the particles [52, 56]. As alternative scenario for the observed copper accumulation by CuO-NP-treated astrocytes it can currently not be exclude that a part of the cellular copper is derived from uptake of copper ions which have been liberated from the applied CuO-NPs. However, astrocytes exposed to CuCl2 contained less copper and remained a higher MTT reduction capacity compared to cells exposed to the respective concentration of copper as CuO-NPs. In addition, the size distribution curves of the CuO-NPs in culture medium were not obviously shifted to smaller sizes within a 24 h incubation (data not shown), demonstrating that CuO-NPs are not rapidly disintegrate to copper ions under the conditions used. These results support the view that the effects observed for CuO-NP-treated astrocytes are predominantly caused by copper liberated in the cells after endocytotic uptake of CuO-NPs.
A treatment of astrocytes with 100 µM CuO-NPs stimulated glycolytic lactate production, elevated the specific cellular GSx content and induced the synthesis of MTs. Similar alterations in astrocytic metabolism were observed after exposure of viable astrocytes to CuCl2 [31, 36, 37]. These results suggest that not the CuO-NPs themselves but rather copper ions which are liberated from the accumulated CuO-NPs are responsible for the observed alterations in astrocytic metabolism. This hypothesis is strongly supported by the ability of the membrane-permeable copper chelator TTM to prevent the observed stimulation of lactate production and MT synthesis in CuO-NP-treated astrocytes. However, a potential TTM-induced disintegration of extracellular CuO-NPs and extracellular chelation of the liberated copper ions would also explain the abolishing effects of TTM on the CuO-NP-induced stimulation in lactate and MT synthesis. Indeed, although the size distribution curves of the dispersed CuO-NPs were not obviously shifted to smaller sizes during a 24 h incubation with an equimolar concentration of TTM (data not shown), the strong absorbance of 100 µM copper-free TTM at 320 and 470 nm [57] was lowered by 30–40 % after incubation with 100 µM copper as CuO-NPs, but completely abolished by application of 100 µM copper ions (data not shown). These data demonstrate that in presence of TTM at least some copper ions were extracellularly chelated by TTM, while the majority of the copper applied remained during incubation in nanoparticle form. This, extracellular chelation may partly contribute to the observed lowering effect by TTM on the cellular copper content of CuO-NP-treated astrocytes.
The ability of TTM to prevent the CuO-NP-induced stimulation of lactate and MT synthesis makes also a potential contribution of the coating material on these effects unlikely, consistent with results from previous studies which revealed that even large amounts of accumulated copper-free DMSA-coated iron oxide nanoparticles (400 nmol iron/mg) did not affect cell viability, GSH and glucose metabolism of cultured astrocytes [58].
The mechanisms involved in the stimulation of glycolytic lactate production and synthesis of GSH and MTs by copper ions are currently unclear. The copper-induced stimulation of glycolytic flux was prevented by an inhibitor of protein synthesis [37]. This would explain the delayed onset of the stimulation of lactate production by CuO-NPs, which was only observed after more than 8 h of incubation, as well as the persistence of this stimulation even after removal of extracellular CuO-NPs. Also the increased astrocytic levels of MTs are likely to be a consequence of elevated concentrations of copper ions as intracellular metal ions can stimulate MT synthesis in astrocytes [31, 47] and as the chelation of such ions is known to prevent MT synthesis [59–61].
The increased GSx levels in viable astrocytes that had been exposed to CuO-NPs are in line with literature data for CuCl2-treated astrocytes [36] and are similar to elevated astrocytic GSx values found after treatment of cultured astrocytes with ammonia, formaldehyde, arsenite or arsenate [36, 62–64]. Such elevated cellular GSx contents have been discussed as consequence of a stressor-induced stimulation of de novo GSH synthesis or of an accelerated uptake of GSH precursors. Also the accelerated GSH export from CuO-NP-treated astrocytes resembles that reported for CuCl2-treated astrocytes [36] and is likely to be a direct consequence of the copper-induced increase in cellular GSH concentration. GSH export from astrocytes is predominantly mediated by the multidrug-resistance protein 1 and has a KM-value of around 100 nmol/mg [62, 64]. Thus, the elevated cellular GSH contents of CuO-NP-treated cultures will directly cause an accelerated GSH export, as previously also reported for astrocytes exposed to copper chloride or arsenicals [36, 62, 64]. A contribution of GSSG export [65, 66] in the increased extracellular GSx accumulation of CuO-NP-treated cells can be excluded, as GSSG hardly contributed to the GSx values observed for cells and media.
Presence of TTM prevented the stimulation of lactate production and MT synthesis in CuO-NP-treated astrocytes, suggesting that intracellular copper ions which are liberated from internalized CuO-NPs mediate these processes. In contrast, presence of TTM was not able to prevent the increase in cellular GSx contents in CuO-NP-treated astrocytes. However, this does not challenge the concept that copper ions liberated from internalized CuO-NPs stimulate GSH synthesis, as TTM itself even in the absence of exogenous copper caused an increase in astrocytic GSx contents. This was not unexpected as TTM has been reported to increase GSH contents in rat hepatocytes [67] and as molybdate application to rats increased GSH levels in liver and kidney [68].
In conclusion, a treatment of cultured astrocytes with sub-toxic amounts of copper in form of CuO-NPs stimulated their glycolytic lactate production and elevated the cellular levels of GSH and MTs. Such alterations in astrocytic metabolism were also reported for astrocytes that had been treated with copper ions [31, 36, 37], suggesting that in astrocytes copper ions are rapidly liberated from internalized CuO-NPs which subsequently induce the observed metabolic alterations. As cellular GSH and MTs are involved in cellular copper storage [33] the increase in the cellular levels of these compounds is likely to contribute to the relative resistance of astrocytes against copper-induced ROS formation and oxidative damage. On the other hand, the copper-induced accelerated release of GSH and lactate may help to supply additional metabolites to neighboring neurons which could serve as precursors for neuronal GSH synthesis [69] and as energy substrate [70, 71], respectively. The observation that astrocytes in culture deal well with moderate concentrations of copper in form of CuO-NPs, suggests that also astrocytes in brain have the potential to accumulate CuO-NPs and to store copper liberated from such NPs, which may help to protect neighboring cells against a toxic potential of CuO-NPs.
References
Hanagata N, Zhuang F, Connolly S, Li J, Ogawa N, Xu M (2011) Molecular responses of human lung epithelial cells to the toxicity of copper oxide nanoparticles inferred from whole genome expression analysis. ACS Nano 5:9326–9338
Li CW, Ciston J, Kanan MW (2014) Electroreduction of carbon monoxide to liquid fuel on oxide-derived nanocrystalline copper. Nature 508:504–507
Patil PR, Krishnamurthy VN, Joshi SS (2008) Effect of nano-copper oxide and copper chromite on the thermal decomposition of ammonium perchlorate. Propellants Explos Pyrotech 33:266–270
Rubilar O, Rai M, Tortella G, Diez MC, Seabra AB, Duran N (2013) Biogenic nanoparticles: copper, copper oxides, copper sulphides, complex copper nanostructures and their applications. Biotechnol Lett 35:1365–1375
Ahmad Z, Vargas-Reus MA, Bakhshi R, Ryan F, Ren GG, Oktar F, Allaker RP (2012) Antimicrobial properties of electrically formed elastomeric polyurethane–copper oxide nanocomposites for medical and dental applications. Methods Enzymol 509:87–99
Dankovich TA, Smith JA (2014) Incorporation of copper nanoparticles into paper for point-of-use water purification. Water Res 63C:245–251
Oberdörster G, Oberdörster E, Oberdörster J (2005) Nanotoxicology: an emerging discipline evolving from studies of ultrafine particles. Environ Health Perspect 113:823–839
Balkhyour MA, Goknil MK (2010) Total fume and metal concentrations during welding in selected factories in Jeddah, Saudi Arabia. Int J Environ Res Public Health 7:2978–2987
Szymczak W, Menzel N, Keck L (2007) Emission of ultrafine copper particles by universal motors controlled by phase angle modulation. J Aerosol Sci 38:520–531
Karlsson HL, Cronholm P, Gustafsson J, Moller L (2008) Copper oxide nanoparticles are highly toxic: a comparison between metal oxide nanoparticles and carbon nanotubes. Chem Res Toxicol 21:1726–1732
Ahamed M, Siddiqui MA, Akhtar MJ, Ahmad I, Pant AB, Alhadlaq HA (2010) Genotoxic potential of copper oxide nanoparticles in human lung epithelial cells. Biochem Biophys Res Commun 396:578–583
Xu J, Li Z, Xu P, Xiao L, Yang Z (2013) Nanosized copper oxide induces apoptosis through oxidative stress in podocytes. Arch Toxicol 87:1067–1073
Alarifi S, Ali D, Verma A, Alakhtani S, Ali BA (2013) Cytotoxicity and genotoxicity of copper oxide nanoparticles in human skin keratinocytes cells. Int J Toxicol 32:296–307
Laha D, Pramanik A, Maity J, Mukherjee A, Pramanik P, Laskar A, Karmakar P (2014) Interplay between autophagy and apoptosis mediated by copper oxide nanoparticles in human breast cancer cells MCF7. Biochim Biophys Acta 1840:1–9
Siddiqui MA, Alhadlaq HA, Ahmad J, Al-Khedhairy AA, Musarrat J, Ahamed M (2013) Copper oxide nanoparticles induced mitochondria mediated apoptosis in human hepatocarcinoma cells. PLoS One 8:e69534
Fu PP, Xia Q, Hwang H-M, Ray PC, Yu H (2014) Mechanisms of nanotoxicity: generation of reactive oxygen species. J Food Drug Anal 22:64–75
Bulcke F, Thiel K, Dringen R (2014) Uptake and toxicity of copper oxide nanoparticles in cultured primary brain astrocytes. Nanotoxicology 8:775–785
An L, Liu S, Yang Z, Zhang T (2012) Cognitive impairment in rats induced by nano-CuO and its possible mechanisms. Toxicol Lett 213:220–227
Kreyling WG, Semmler M, Erbe F, Mayer P, Takenaka S, Schulz H, Oberdörster G, Ziesenis A (2002) Translocation of ultrafine insoluble iridium particles from lung epithelium to extrapulmonary organs is size dependent but very low. J Toxicol Environ Health A 65:1513–1530
Lockman PR, Koziara JM, Mumper RJ, Allen DD (2004) Nanoparticle surface charges alter blood-brain barrier integrity and permeability. J Drug Target 12:635–641
Oberdörster G, Sharp Z, Atudorei V, Elder A, Gelein R, Kreyling W, Cox C (2004) Translocation of inhaled ultrafine particles to the brain. Inhal Toxicol 16:437–445
Sharma HS, Sharma A (2012) Neurotoxicity of engineered nanoparticles from metals. CNS Neurol Disord Drug Targets 11:65–80
Yim YS, Choi JS, Kim GT, Kim CH, Shin TH, Kim DG, Cheon J (2012) A facile approach for the delivery of inorganic nanoparticles into the brain by passing through the blood-brain barrier (BBB). Chem Commun (Camb) 48:61–63
Kao YY, Cheng TJ, Yang DM, Wang CT, Chiung YM, Liu PS (2012) Demonstration of an olfactory bulb–brain translocation pathway for ZnO nanoparticles in rodent cells in vitro and in vivo. J Mol Neurosci 48:464–471
Karmakar A, Zhang Q, Zhang Y (2014) Neurotoxicity of nanoscale materials. J Food Drug Anal 22:147–160
Liu Y, Gao Y, Liu Y, Li B, Chen C, Wu G (2014) Oxidative stress and acute changes in murine brain tissues after nasal instillation of copper particles with different sizes. J Nanosci Nanotechnol 14:4534–4540
Liu Y, Gao Y, Zhang L, Wang T, Wang J, Jiao F, Li W, Liu Y, Li Y, Li B, Chai Z, Wu G, Chen C (2009) Potential health impact on mice after nasal instillation of nano-sized copper particles and their translocation in mice. J Nanosci Nanotechnol 9:6335–6343
Cupaioli FA, Zucca FA, Boraschi D, Zecca L (2014) Engineered nanoparticles. How brain friendly is this new guest? Prog Neurobiol 119–120C:20–38
López-Hidalgo M, Schummers J (2014) Cortical maps: a role for astrocytes? Curr Opin Neurobiol 24:176–189
Verkhratsky A, Nedergaard M, Hertz L (2014) Why are astrocytes important? Neurochem Res (in press)
Scheiber IF, Dringen R (2013) Astrocyte functions in the copper homeostasis of the brain. Neurochem Int 62:556–565
Hohnholt MC, Geppert M, Luther EM, Petters C, Bulcke F, Dringen R (2013) Handling of iron oxide and silver nanoparticles by astrocytes. Neurochem Res 38:227–239
Scheiber IF, Mercer JF, Dringen R (2014) Metabolism and functions of copper in brain. Prog Neurobiol 116:33–57
Scheiber IF, Mercer JF, Dringen R (2010) Copper accumulation by cultured astrocytes. Neurochem Int 56:451–460
Scheiber IF, Schmidt MM, Dringen R (2012) Copper export from cultured astrocytes. Neurochem Int 60:292–300
Scheiber IF, Dringen R (2011) Copper-treatment increases the cellular GSH content and accelerates GSH export from cultured rat astrocytes. Neurosci Lett 498:42–46
Scheiber IF, Dringen R (2011) Copper accelerates glycolytic flux in cultured astrocytes. Neurochem Res 36:894–903
Petters C, Irrsack E, Koch M, Dringen R (2014) Uptake and metabolism of iron oxide nanoparticles in brain cells. Neurochem Res 39:1648–1660
Hamprecht B, Löffler F (1985) Primary glial cultures as a model for studying hormone action. Methods Enzymol 109:341–345
Tulpule K, Hohnholt MC, Hirrlinger J, Dringen R (2014) Primary cultures of rat astrocytes and neurons as model systems to study metabolism and metabolite export from brain cells. In: Hirrlinger J, Waagepetersen H (eds) Neuromethods 90: brain energy metabolism. Springer, Heidelberg, pp 45–72
Petters C, Dringen R (2014) Comparison of primary and secondary rat atrocyte cultures regarding glucose and glutathione metabolism and the accumulation of iron oxide nanoparticles. Neurochem Res 39:46–58
Lowry OH, Rosebrough NJ, Farr AL, Randall RJ (1951) Protein measurement with the Folin phenol reagent. J Biol Chem 193:265–275
Dringen R, Kussmaul L, Hamprecht B (1998) Detoxification of exogenous hydrogen peroxide and organic hydroperoxides by cultured astroglial cells assessed by microtiter plate assay. Brain Res Protoc 2:223–228
Liddell JR, Zwingmann C, Schmidt MM, Thiessen A, Leibfritz D, Robinson SR, Dringen R (2009) Sustained hydrogen peroxide stress decreases lactate production by cultured astrocytes. J Neurosci Res 87:2696–2708
Hirrlinger J, Dringen R (2005) Multidrug resistance protein 1-mediated export of glutathione and glutathione disulfide from brain astrocytes. Methods Enzymol 400:395–409
Hohnholt MC, Geppert M, Dringen R (2011) Treatment with iron oxide nanoparticles induces ferritin synthesis but not oxidative stress in oligodendroglial cells. Acta Biomater 7:3946–3954
Luther E, Schmidt M, Diendorf J, Epple M, Dringen R (2012) Upregulation of metallothioneins after exposure of cultured primary astrocytes to silver nanoparticles. Neurochem Res 37:1639–1648
Luther EM, Koehler Y, Diendorf J, Epple M, Dringen R (2011) Accumulation of silver nanoparticles by cultured primary brain astrocytes. Nanotechnology 22:375101
Akhtar MJ, Kumar S, Alhadlaq HA, Alrokayan SA, Abu-Salah KM, Ahamed M (2014) Dose-dependent genotoxicity of copper oxide nanoparticles stimulated by reactive oxygen species in human lung epithelial cells. Toxicol Ind Health (in press)
Cronholm P, Karlsson HL, Hedberg J, Lowe TA, Winnberg L, Elihn K, Wallinder IO, Moller L (2013) Intracellular uptake and toxicity of Ag and CuO nanoparticles: a comparison between nanoparticles and their corresponding metal ions. Small 9:970–982
Di Bucchianico S, Fabbrizi MR, Misra SK, Valsami-Jones E, Berhanu D, Reip P, Bergamaschi E, Migliore L (2013) Multiple cytotoxic and genotoxic effects induced in vitro by differently shaped copper oxide nanomaterials. Mutagenesis 28:287–299
Wang Z, von dem Bussche A, Kabadi PK, Kane AB, Hurt RH (2013) Biological and environmental transformations of copper-based nanomaterials. ACS Nano 7:8715–8727
Geppert M, Petters C, Thiel K, Dringen R (2013) The presence of serum alters the properties of iron oxide nanoparticles and lowers their accumulation by cultured brain astrocytes. J Nanopart Res 15:1349–1364
Lamkowsky MC, Geppert M, Schmidt MM, Dringen R (2012) Magnetic field-induced acceleration of the accumulation of magnetic iron oxide nanoparticles by cultured brain astrocytes. J Biomed Mater Res A 100A:323–334
Cuillel M, Chevallet M, Charbonnier P, Fauquant C, Pignot-Paintrand I, Arnaud J, Cassio D, Michaud-Soret I, Mintz E (2014) Interference of CuO nanoparticles with metal homeostasis in hepatocytes under sub-toxic conditions. Nanoscale 6:1707–1715
Wang Z, Li N, Zhao J, White JC, Qu P, Xing B (2012) CuO nanoparticle interaction with human epithelial cells: cellular uptake, location, export, and genotoxicity. Chem Res Toxicol 25:1512–1521
Clarke NJ, Laurie SH (1982) The copper–molybdenium antagonism in ruminants. II: interactions of thiomolybdates with copper(II) in aqueous media. Inorg Chim Acta 66:L35–L38
Geppert M, Hohnholt MC, Nurnberger S, Dringen R (2012) Ferritin up-regulation and transient ROS production in cultured brain astrocytes after loading with iron oxide nanoparticles. Acta Biomater 8:3832–3839
Czachor JD, Cherian MG, Koropatnick J (2002) Reduction of copper and metallothionein in toxic milk mice by tetrathiomolybdate, but not deferiprone. J Inorg Biochem 88:213–222
Nemec AA, Leikauf GD, Pitt BR, Wasserloos KJ, Barchowsky A (2009) Nickel mobilizes intracellular zinc to induce metallothionein in human airway epithelial cells. Am J Respir Cell Mol Biol 41:69–75
Parat M-O, Richard M-J, Meplan C, Favier A, Béani J-C (1999) Impairment of cultured cell proliferation and metallothionein expression by metal chelator NNN′ N′-tetrakis-(2-pyridylmethyl) ethylene diamine. Biol Trace Elem Res 70:51–68
Sagara J, Makino N, Bannai S (1996) Glutathione efflux from cultured astrocytes. J Neurochem 66:1876–1881
Węgrzynowicz M, Hilgier W, Dybel A, Oja SS, Saransaari P, Albrecht J (2007) Upregulation of cerebral cortical glutathione synthesis by ammonia in vivo and in cultured glial cells: the role of cystine uptake. Neurochem Int 50:883–889
Tulpule K, Schmidt MM, Boecker K, Goldbaum O, Richter-Landsberg C, Dringen R (2012) Formaldehyde induces rapid glutathione export from viable oligodendroglial OLN-93 cells. Neurochem Int 61:1302–1313
Hirrlinger J, König J, Keppler D, Lindenau J, Schulz JB, Dringen R (2001) The multidrug resistance protein MRP1 mediates the release of glutathione disulfide from rat astrocytes during oxidative stress. J Neurochem 76:627–636
Minich T, Riemer J, Schulz JB, Wielinga P, Wijnholds J, Dringen R (2006) The multidrug resistance protein 1 (Mrp1), but not Mrp5, mediates export of glutathione and glutathione disulfide from brain astrocytes. J Neurochem 97:373–384
Chao PY, Allen KG (1992) Glutathione production in copper-deficient isolated rat hepatocytes. Free Radic Biol Med 12:145–150
Panneerselvam SR, Govindasamy S (2004) Effect of sodium molybdate on the status of lipids, lipid peroxidation and antioxidant systems in alloxan-induced diabetic rats. Clin Chim Acta 345:93–98
Dringen R, Pfeiffer B, Hamprecht B (1999) Synthesis of the antioxidant glutathione in neurons: supply by astrocytes of CysGly as precursor for neuronal glutathione. J Neurosci 19:562–569
Dringen R, Gebhardt R, Hamprecht B (1993) Glycogen in astrocytes: possible function as lactate supply for neighboring cells. Brain Res 623:208–214
Bouzier-Sore AK, Pellerin L (2013) Unraveling the complex metabolic nature of astrocytes. Front Cell Neurosci 7:179
Conflict of interest
The authors declare that they have no conflict of interest.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Bulcke, F., Dringen, R. Copper Oxide Nanoparticles Stimulate Glycolytic Flux and Increase the Cellular Contents of Glutathione and Metallothioneins in Cultured Astrocytes. Neurochem Res 40, 15–26 (2015). https://doi.org/10.1007/s11064-014-1458-0
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11064-014-1458-0