
Abstract
Recently the recruitment/migration of myeloid derived suppressor cells (MDSCs) to tumor microenvironment after chemotherapy has attracted much attention. To determine the detailed mechanism for the responses of MDSCs to these chemotherapies, we investigated the changes of galectin-3 and MDSCs in response to cisplatin(0.4 mg/kg, 4 mg/kg) treatment both in vivo and ex vivo. In the process of cisplatin, we assessed levels of galectin-3 and MDSCs in the Lewis lung cancer (LLC) bearing mice using immunohistochemistry, enzyme-linked immunosorbent assay (ELISA), immunofluorence and flow cytometry (FCM). The expression and changes of galectin-3 in the LLC cell line were detected by western blot, immunofluorence and ELISA. The ligand for galectin-3 on MDSCs and the chemotaxis of galectin-3 to MDSCs were confirmed using FCM and transwell. Parallel increased level of galectin-3 with the number of MDSCs in vivo was detected after cisplatin treatment. LLC cells expressed galectin-3 and cisplatin increased galectin-3 level in the culture medium. Furthermore, MDSCs were detected to express CD98, the ligand of galectin-3, and could be recruited by galectin-3. Our results suggested that the elevated expression of gelectin-3 in LLC tumor cells may contribute to the migration of MDSCs to the tumor microenvironment in response to cisplatin.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Chemotherapy remains the primary treatment for most advanced and metastases cancers. However, complete elimination of cancer cells remains a great challenge for most current anti-tumor drugs. Tumors could develop complex chemotherapy resistance mechanisms in vivo that were crucially dependent on their interaction with host factors [1]. Indeed, the microenvironment has been recognized as a major factor influencing the growth as well as metastatic progression of cancer and impacting the outcome of therapy [2, 3].
MDSCs are a heterogeneous population of cells that are composed of myeloid progenitor cells and immature myeloid cells (IMC). The expansion of MDSCs originates from a block in the differentiation of IMCs into mature myeloid cells under pathological conditions [4]. It is believed that MDSCs are major contributors mediating tumor escape from antitumor immunity. Additionally, MDSCs accumulate in most tumor-bearing animal models. And preclinical data have indicated that MDSC levels appear to correlate proportionally with tumor burden [5, 6].
Given the important immunosuppressive role of MDSCs on individuals during tumor progression, a better understanding of the local tumor microenvironment and the exact mechanisms of induction and/or expansion of MDSCs in the tumor milieu should provide opportunities for new treatments that target these cells and alter the balance in favor of more effective antitumour immune responses. Most type of tumor cells can release nanovesicles called exosomes which could represent versatile functions in modulating the anti-tumor immune response by affecting effective immune cells [7, 8].Recently Umansky et al. [9] has reviewed that various cytokines and chemokines accumulated in the tumor microenvironment were demonstrated to stimulate the migration of MDSCs towards tumor lesions.
Evidence showed that tumor derived chronic inflammatory factors such as granulocyte–macrophage colony stimulating factor (GM-CSF) [10], IL-1 beta [11], IL-6 [12], vascular endothelial growth factor (VEGF) [13], PGE2 [14] could act as a vehicle to induce the expansion of MDSCs. In addition, Sevko et al. [15] has revealed that low-dose cyclophosphamide therapy enhanced the production of inflammatory mediators in melanoma lesions associated with increased accumulation of MDSCs. However, docetaxel reduced the number of MDSCs and directly inhibited the immunosuppressive effect of them in a STAT3 signaling dependent manner [16]. Evidence also showed that the accumulation of MDSCs in renal cell carcinoma (RCC) patients could be reversed by sunitinib, the tyrosine kinase inhibitor, without inducing the MDSC maturation [17]. This suggested, apart from chronic inflammatory factors, anti-tumor drugs may affect the recruitment of MDSCs through distinct signaling pathways in tumor lesions.
Galectin-3, a member of the β-galactoside-binding protein family, is an intracellular and extracellular lectin consisting of three structural domains and is showed to interact with laminin and glycoproteins which are involved in cell adhesion [18]. Interestingly, galectin-3 has been found to act as a chemoattractant for monocytes and macrophages [19]. We herein tested the possible correlation between galectin-3 and MDSCs after cisplatin treatment in the tumor microenvironment of a LLCs mice model.
Materials and methods
In vivo experiment
Mice and cell line
Six- to eight-week-old C57BL/6 mice were purchased from the Department of Laboratory Animal Science, School of Medicine, Fudan University. Animals were housed under specific pathogen-free conditions in a temperature and humidity controlled environment. All animal experiments were conducted within guidelines established by the Institutional Animal Care and Use Committee of Fudan University. The LLC cell line, originated spontaneously from the lung carcinoma of the C57BL/6 mouse, was purchased from the Cell Library of Chinese Academy of Sciences and maintained at 37 °C in a complete medium(DMEM with 10 % FBS) humidified with 5 % CO2 atmosphere.
Cisplatin intervention
A total of 33 C57BL/6 mice were involved in this study. Among them, 18 Mice were inoculated subcutaneously in the flank with 2 × 106 LLCs and randomly divided into three groups with each including six mice. Then cisplatin (QiLu Pharmaceutical, China) was injected intraperitoneally at a dose of 0.4 mg/kg or 4 mg/kg daily from day 14 to day 18 after the LLCs inoculation. The control group was given an equivalent amount of normal saline (NS). Meanwhile, the other 15 mice without LLCs inoculation were also divided into three groups with each having five samples and were treated with NS, cisplatin (0.4 mg/kg) and cisplatin (4 mg/kg) for 5 days respectively. Mice were mercy killed 72 h after the last injection of cisplatin. And the samples were collected and storaged according to each procedure involved.
Estimation of Gr1+CD11b+cells by immunofluorescence and FCM
After 5 days of cisplatin treatment, spleens and tumor tissues from LLCs mice were obtained on day 21 after inoculation and dissociated to single-cell suspensions. And the RBC was removed by lysis, as described for splenocytes [20]. Cells were stained with mAbs to MDSC markers Gr1 and CD11b and analyzed by FCM, as described [20].
The paraffin embedded tumor sections were deparaffined using a graded ethanol series, and endogenous peroxidase activity was blocked by soaking in 0.3 % hydrogen peroxide. Then they were stained with anti-mouse Abs to CD11b and Gr-1(BD Pharmingen, USA), followed by Alexa 594- or Alexa 488-conjugated secondary Abs (Invitrogen, USA), and examined using an inverted 80i Nikon wide field fluorescence microscope and photographed with a CCD SPOT RT Camera. Cells in three high power fields were counted for quantitation.
Determination of galectin-3 by immunohistochemistry and ELISA
Sections of formalin-fixed, paraffin embedded tumor tissues from LLC mice were collected. A monoclonal rat anti mouse galectin-3 (BioLegend, San Diego, USA) was used. Slides were incubated with the primary antibody diluted 1:1000 for 1 h at room temperature. Secondary HRP conjugated goat anti-rat IgG(H+L) (1:500) (sigma, St. Louis, USA) was added and incubated for 1 h at 37 °C. Negative control slides were immunostained under identical conditions, substituting the primary antibody with buffer solution. Sections were assessed by standard light microscopy by three persons. Immunoreactivity for galectin-3 was scored semiquantitatively in areas of tumour as previously described [21]: (i) for strength of staining (absent = 0; weak = 1; moderate = 2; strong = 3) and (ii) for percentage of cells staining (absent = 0; <20 % = 1; 20–80 % = 2; >80 % = 3).
The level of galectin-3 in the plasma of mice from each group was measured by a galectin-3 ELISA kit (R&D Company, USA) according to the manufacturer’s protocol. Plates were read at 420 nm on a Themo-sci 1510 microplate reader and quantified using a standard curve. Data represented the mean ± SD of triplicate wells.
Ex vivo experiment
Detection of galectin-3 in LLC cells and the conditioned media(CM)
Immunofluorescence
The localization of galectin-3 on LLC cells was determined by immunofluorescence as described earlier [18].
Western blot
LLC cells were collected, washed with ice-cold PBS, and lysed in 30 mmol/L HEPES (pH 7.5), 10 mmol/L NaCl, 5 mmol/L MgCl2, 25 mmol/L NaF, 1 mmol/L EGTA, 1 % Triton X-100, 10 % glycerol, and protease and phosphatase inhibitor cocktails (Sigma). To detect galectin-3 expression, Cell lysates were centrifuged at 12,000×g for 5 min to remove nuclei and cell debris. The proteins of the soluble extracts were collected. Western blotting was done as described earlier [22]
ELISA
The LLC cells were cultured with or without 2 μM cisplatin for 48 h and the supernatants were collected for galectin-3 detection. The levels of galectin-3 in the supernatants of the LLC cell line were detected by a galectin-3 ELISA kit (R&D Company, USA) according to the manufacturer’s protocol. Plates were read at 420 nm on a Themo-sci 1510 microplate reader and quantified using a standard curve. Data represented the mean ± SD of triplicate wells.
Chemotaxis of galectin-3 to MDSCs
Spleens from LLCs tumor-bearing mice were obtained on day 21 after inoculation when the tumors were about 1.5 cm in diameter and dissociated to single-cell suspensions. Splenic lymphocytes were detached using lymphocyte separation medium (BioLegend, USA). Thereafter, red blood cells were depleted by the RBC lysing buffer, and MDSCs were isolated by magnetic bead sorting using CD11b mAb and MS columns according to the directions of the manufacturer (Miltenyi Biotec, Germany). Purified splenic MDSCs contained more than 90 % Gr1+CD11b+ cells. CD98, one of the ligands for galectin-3, was also tested by FCM on the surface of MDSCs.
In vitro migration of MDSCs was evaluated in 24-well plates with transwell polycarbonate permeable supports (8.0 μm) (Costar Corning). 5 × 106 MDSCs were plated in 100 μl of serum-free IMDM in the upper compartment and 500 ul of chemoattractant [LLC cell CM ± 10 μg of Abs to galectin-3, or IgG control Ab (BioLegend, USA)] were added to the lower compartment. Plates were incubated at 37 °C with 5 % CO2 for 3 h, and the number of MDSCs in the bottom compartment was counted. For CM, supernatants were harvested from confluent cultures of LLCs cultured in medium IMDM containing 3 % serum. And CM was filter sterilized and stored at -80 °C as single-use aliquots.
Statistical analysis
Data was analyzed with student’s two-tailed t test by using SPSS16.0. Differences were considered significant if p < 0.05.
Results
Parallel elevated level of galectin-3 with MDSCs were detected after cisplatin therapy in LLC mice
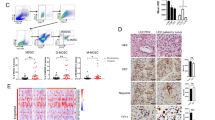
Figure 1a showed that cisplatin significantly increased the expression of galectin-3 in the tumor site, and this protein mainly localized in the cytoplasm. Meanwhile, the galectin-3 level in the plasma was increased by 125 % in the low cisplatin (0.4 mg/kg) group and 200 % in the high cisplatin (4 mg/kg) group as compared with the NS group. However, galectin-3 was not affected by cisplatin in mice without LLC cell inoculation (Fig. 1b).

Cisplatin increased the expression of galectin-3 both in the tumor tissue and the peripheral blood. a Galectin-3 protein in tumor tissues determined by immunoperoxidase staining: (a), negative control; (b), Weak staining without cisplatin treatment; (c), Moderate staining with 0.4 mg/kg of cisplatin; (d), Strong staining with 4 mg/kg of cisplatin. b Galectin-3 level in peripheral blood measured by ELISA. Compared to the NS group, the level of galectin-3 in blood plasma of the higher cisplatin level group (4 mg/kg) significantly increased (n = 6; *, P < 0.05). In contrast, in LLC free mice, cisplatin treatment could not change the galectin-3 level (n = 5)
The immunofluorescence result showed that compared to the NS group, the cisplatin group of both dose had higher number of MDSCs in the tumor site (Fig. 2a, b). Meanwhile, the Flow Cemetry results indicated that, cisplatin increased the number of MDSCs from both tumor sites and spleen in tumor-bearing mice (Fig. 2c, d).

Cisplatin induced MDSCs accumulation both in tumor tissue and spleen. a Immunofluorescence showed CD11b+Gr-1+cells accumulated in the spleen after cisplatin treatment. b Average number of Gr1+CD11b+cells counted in three high power fields of tumor tissue in different groups. c, d The proportion of Gr-1+CD11b+MDSC was evaluated by flow cytometry using anti-Gr-1 and anti-CD11b mAbs in spleens and tumor tissues with or without cisplatin treatment. Each group included six mice (*, P < 0.05)
Cisplatin treatment elevated the galectin-3 level in LLC cell lines
To confirm that the elevated galectin-3 level in the LLC mice mainly originated from LLC tumor cells, we observed the expression of galectin-3 in the LLC cells. The results showed that galectin-3 mainly located in the cytoplasm of LLC cells (Fig. 3). Western blot results also showed positive expression of galectin-3 in the LLC cells (Fig. 4c). Moreover, we compared the galectin-3 levels in each group of cultural supernatant of LLC cell line. All specimens tested showed positive expression of galectin-3. Moreover, the galectin-3 level of the culture supernatant with cisplatin treatment (14.26 ± 2.16 ng/ml) was higher than that without cisplatin intervention (9.23 ± 2.54 ng/ml), although not to statistical significance (p > 0.05).

The expression of galectin-3 in LLC cells was detected by immunofluorescence. LLCs of climbs were incubated with galectin-3-PE and examined using an inverted 80i Nikon wide field fluorescence microscope and photographed with a CCD SPOT RT Camera. (a), The LLC cells under normal light; (b), The nucleuses were dyed with DAPI; (c, d), Galectin-3 located mainly in the cytoplasm

LLCs derived galectin-3 proteins were chemotactic for MDSCs. a Splenic MDSCs were isolated from the LLC mice and the Gr-1 and CD11b were detected by FCM. b Splenic MDSCs were tested by chemotaxis assay for their migration in response to LLC culture medium (CM). Data are pooled from two independent experiments. c The western blot confirmed the expression of galectin-3 in LLC cells. d The galectin-3 ligand CD98 was detected by flow cytometer on the cell membrane of MDSCs
MDSCs expressed CD98 and were recruited by galectin-3
To further estimate the possible role that galectin-3 may play on MDSCs. We tested the chemotactic effect of galectin-3 to MDSCs from LLCs mice in vitro. Supernatant from LLCs CM which had been verified to containing galectin-3 was collected as the chemoattractant, and the migration of MDSCs was tested in response to this CM using transwell chambers. We observed that the number of MDSCs in the group added with Abs of galectin-3 to the CM obviously decreased compared to that without galectin-3 Abs or IgG control Ab (Fig. 4b). This result suggested that galectin-3 may be one of the pivotal chemoattractants for MDSCs.
To confirm that galectin-3 could interact directly with MDSCs, we tried to make clear whether MDSCs have receptors for galectin-3. It is reported that more than one ligands exist that have high affinity to galectin-3 [23, 24]. CD98, which was commonly recognized as one of the receptors for galectin-3, was detected to be located on the surface of isolated MDSCs (Fig. 4d).
Discussion
In the context of cisplatin, along with the increased level of galectin-3, the number of MDSCs both in the tumor tissue (Fig. 2a, b) as well as the spleen (Fig. 2c, d) was detected to be increased in our test. This is in accordance with previous observations revealing that chemotherapy could increase the level of MDSCs in circulation [5] as well as the tumor site [25]. Furthermore, we detected CD98, one of the ligand for galectin-3 on the surface of MDSCs. Considering that the LLC cell line could express galectin-3 which was confirmed in our in vitro experiment to be chemotactic to MDSCs, we propose that galectin-3 may contribute at least partially to the accumulation of MDSCs in the LLCs bearing mice in response to cisplatin therapy.
The progression of various tumors was considered to correlate with abnormal levels of galectin-3 [26–28]. Moreover, preclinical data has shown that higher level of serum galectin-3 was related to lymph node metastasis [29, 30]. In this study, the increased accumulation of galectin-3 in the cytoplasm of the LLC tumor as well as in the circulation (Fig. 1) as detected in our study strongly suggested the complicated mechanism by which LLC tumor cells respond to cisplatin. Evidence showed that tumor cells were able to expel cisplatin through the enhanced release of exosomes [31], this was in accordance with our results that increased level of peripheral galectin-3 may be the response of the LLC tumor microenvironment to the stimulation of cisplatin.
It should be noted that cisplatin was not detected to increase the galectin-3 level of the LLC supernatant statistically in our ex vivo experiment(14.26 ± 2.16 ng/ml vs 9.23 ± 2.54 ng/ml). Nevertheless, our in vivo results clearly showed that galectin-3 mostly localized in the cytoplasm of the LLC tumor cells, and the immunostaining of galectin-3 increased significantly after cisplatin intervention (Figs. 1a, 3). It was reported that intracellular galectin-3 shifts dynamically depending on the cell status [32]. Evidence showed that levels of nuclear galectin-3 decreased during the progression from normal to cancerous states. Concomitantly, cytoplasmic expression was increased [33]. On this basis, the elevated of cytoplasmic galectin-3 expression after cisplatin in our study may indicate the harmful reactions of tumor cells. Generally, we demonstrated that LLC tumor cells could express galectin-3 and that the elevated galectin-3 level was mostly the response of this tumor to cisplatin therapy.
Additionally, our immunofluorescence and FCM results showed that, besides the elevated number of MDSCs in spleen, the accumulation of MDSCs also increased significantly in the tumor site after cisplatin intervention (Fig. 2). It was reported that the MDSCs infiltrated to the tumor tissue could mediate the tumor cell resistance to cytotoxic T cells [34]. Therefore, it is likely that the increased accumulation of MDSCs in the tumor site may help to be more competitive to the immunosurveillance of the host. Various factors including GM-CSF [10], PGE2 [35], stem cell factor (SCF) [36] have been found to induce the expansion of MDSCs. And the signaling pathways triggered by most of these factors are reported to converge on Janus kinase (JAK) protein family members and signal transducer and activator of transcription 3 (STAT3) [4]. It was reported that galectin-3 regulates the adhesion of epithelial cells to collagens and laminins, and promotes keratinocyte migration over wound re-epithelialization in mice [37, 38].Furthermore, galectin-3 stimulates neutrophil adhesion and migration, as well as eosinophil adhesion [39, 40].Interestingly, galectin-3 has recently been revealed to exert regulatory actions through the JAK-STAT pathway under pathological conditions [41]. If that is true, our results that galectin-3 induced accumulation of MDSCs in tumor tissue as well as the spleen in the LLCs bearing mice may be dependent on the JAK-STAT pathway.
Recently, the response of the tumor microenvironment to chemotherapy has attracted much attention for their potentials in drug resistance [42]. Our results strongly suggested that cisplatin-induced galectin-3 might act as a chemotactic factor to MDSCs. Interestingly, a recent study has indicated that common cytotoxic drugs could induce neoplastic cells to produce monocyte/macrophage recruitment factors, which in turn enhance macrophage infiltration into the tumors [43]. Nakasone et al. [25] further proposed that the myeloid cell infiltration occurred after chemotherapy impedes the response to therapy, and this could be regulated by the tumor immune microenvironment. Our observation was in accordance with these studies, and we propose that galectin-3 may mediate the cisplatin induced expansion of MDSCs, which could exert adverse effects to cisplatin therapy. Further researches are needed to confirm that.
Conclusions
In summary, our results suggested that galectin-3 was chemotactic to MDSCs. And the accumulation of MDSCs in the tumor site as well as the peripheral blood of the LLCs mice in response to cisplatin therapy may be due to the tumor derived gelectin-3. This primary data provided new clues that targeting MDSCs recruitment, in combination with cytotoxic therapy, may have generally improved outcomes in treating cancers.
References
Teicher BA, Herman TS, Holden SA, Wang YY, Pfeffer MR, Crawford JW, Frei E 3rd (1990) Tumor resistance to alkylating agents conferred by mechanisms operative only in vivo. Science 247:1457–1461
Basak GW, Srivastava AS, Malhotra R, Carrier E (2009) Multiple myeloma bone marrow niche. Curr Pharm Biotechnol 10:345–346
Joyce JA, Pollard JW (2009) Microenvironmental regulation of metastasis. Nat Rev Cancer 9:239–252. doi:10.1038/nrc2618
Gabrilovich DI, Nagaraj S (2009) Myeloid-derived suppressor cells as regulators of the immune system. Nat Rev Immunol 9:162–174. doi:10.1038/nri2506
Diaz-Montero CM, Salem ML, Nishimura MI, Garrett-Mayer E, Cole DJ, Montero AJ (2009) Increased circulating myeloid-derived suppressor cells correlate with clinical cancer stage, metastatic tumor burden, and doxorubicin–cyclophosphamide chemotherapy. Cancer Immunol Immunother 58:49–59. doi:10.1007/s00262-008-0523-4
Gabitass RF, Annels NE, Stocken DD, Pandha HA, Middleton GW (2011) Elevated myeloid-derived suppressor cells in pancreatic, esophageal and gastric cancer are an independent prognostic factor and are associated with significant elevation of the Th2 cytokine interleukin-13. Cancer Immunol Immunother 60:1419–1430. doi:10.1007/s00262-011-1028-0
Iero M, Valenti R, Huber V, Filipazzi P, Parmiani G, Fais S, Rivoltini L (2008) Tumour-released exosomes and their implications in cancer immunity. Cell Death Differ 15:80–88. doi:10.1038/sj.cdd.4402237
Filipazzi P, Burdek M, Villa A, Rivoltini L, Huber V (2012) Recent advances on the role of tumor exosomes in immunosuppression and disease progression. Semin Cancer Biol 22:342–349. doi:10.1016/j.semcancer.2012.02.005
Umansky V, Sevko A (2012) Tumor microenvironment and myeloid-derived suppressor cells. Cancer Microenviron. doi:10.1007/s12307-012-0126-7
Morales JK, Kmieciak M, Knutson KL, Bear HD, Manjili MH (2010) GM-CSF is one of the main breast tumor-derived soluble factors involved in the differentiation of CD11b-Gr1-bone marrow progenitor cells into myeloid-derived suppressor cells. Breast Cancer Res Treat 123:39–49. doi:10.1007/s10549-009-0622-8
Bunt SK, Sinha P, Clements VK, Leips J, Ostrand-Rosenberg S (2006) Inflammation induces myeloid-derived suppressor cells that facilitate tumor progression. J Immunol 176:284–290
Wu CT, Hsieh CC, Lin CC, Chen WC, Hong JH, Chen MF (2012) Significance of IL-6 in the transition of hormone-resistant prostate cancer and the induction of myeloid-derived suppressor cells. J Mol Med (Berl) 90:1343–1355. doi:10.1007/s00109-012-0916-x
Jayaraman P, Parikh F, Lopez-Rivera E, Hailemichael Y, Clark A, Ma G, Cannan D, Ramacher M, Kato M, Overwijk WW, Chen SH, Umansky VY, Sikora AG (2012) Tumor-expressed inducible nitric oxide synthase controls induction of functional myeloid-derived suppressor cells through modulation of vascular endothelial growth factor release. J Immunol 188:5365–5376. doi:10.4049/jimmunol.1103553
Obermajer N, Muthuswamy R, Odunsi K, Edwards RP, Kalinski P (2011) PGE(2)-induced CXCL12 production and CXCR4 expression controls the accumulation of human MDSCs in ovarian cancer environment. Cancer Res 71:7463–7470. doi:10.1158/0008-5472.can-11-2449
Sevko A, Sade-Feldman M, Kanterman J, Michels T, Falk CS, Umansky L, Ramacher M, Kato M, Schadendorf D, Baniyash M, Umansky V (2013) Cyclophosphamide promotes chronic inflammation-dependent immunosuppression and prevents antitumor response in melanoma. J Invest Dermatol 133:1610–1619. doi:10.1038/jid.2012.444
Kodumudi KN, Woan K, Gilvary DL, Sahakian E, Wei S, Djeu JY (2010) A novel chemoimmunomodulating property of docetaxel: suppression of myeloid-derived suppressor cells in tumor bearers. Clin Cancer Res 16:4583–4594. doi:10.1158/1078-0432.CCR-10-0733
Ko JS, Zea AH, Rini BI, Ireland JL, Elson P, Cohen P, Golshayan A, Rayman PA, Wood L, Garcia J, Dreicer R, Bukowski R, Finke JH (2009) Sunitinib mediates reversal of myeloid-derived suppressor cell accumulation in renal cell carcinoma patients. Clin Cancer Res 15:2148–2157. doi:10.1158/1078-0432.CCR-08-1332
Ohannesian DW, Lotan D, Thomas P, Jessup JM, Fukuda M, Gabius HJ, Lotan R (1995) Carcinoembryonic antigen and other glycoconjugates act as ligands for galectin-3 in human colon carcinoma cells. Cancer Res 55:2191–2199
Sano H, Hsu DK, Yu L, Apgar JR, Kuwabara I, Yamanaka T, Hirashima M, Liu FT (2000) Human galectin-3 is a novel chemoattractant for monocytes and macrophages. J Immunol 165:2156–2164
Sinha P, Clements VK, Ostrand-Rosenberg S (2005) Reduction of myeloid-derived suppressor cells and induction of M1 macrophages facilitate the rejection of established metastatic disease. J Immunol 174:636–645
Buttery R, Monaghan H, Salter DM, Sethi T (2004) Galectin-3: differential expression between small-cell and non-small-cell lung cancer. Histopathology 44:339–344. doi:10.1111/j.1365-2559.2004.01815.x
Nefedova Y, Huang M, Kusmartsev S, Bhattacharya R, Cheng P, Salup R, Jove R, Gabrilovich D (2004) Hyperactivation of STAT3 is involved in abnormal differentiation of dendritic cells in cancer. J Immunol 172:464–474
Dong S, Hughes RC (1997) Macrophage surface glycoproteins binding to galectin-3 (Mac-2-antigen). Glycoconj J 14:267–274
MacKinnon AC, Farnworth SL, Hodkinson PS, Henderson NC, Atkinson KM, Leffler H, Nilsson UJ, Haslett C, Forbes SJ, Sethi T (2008) Regulation of alternative macrophage activation by galectin-3. J Immunol 180:2650–2658
Nakasone ES, Askautrud HA, Kees T, Park JH, Plaks V, Ewald AJ, Fein M, Rasch MG, Tan YX, Qiu J, Park J, Sinha P, Bissell MJ, Frengen E, Werb Z, Egeblad M (2012) Imaging tumor-stroma interactions during chemotherapy reveals contributions of the microenvironment to resistance. Cancer Cell 21:488–503. doi:10.1016/j.ccr.2012.02.017
Matsuda Y, Yamagiwa Y, Fukushima K, Ueno Y, Shimosegawa T (2008) Expression of galectin-3 involved in prognosis of patients with hepatocellular carcinoma. Hepatol Res 38:1098–1111. doi:10.1111/j.1872-034X.2008.00387.x
Baptiste TA, James A, Saria M, Ochieng J (2007) Mechano-transduction mediated secretion and uptake of galectin-3 in breast carcinoma cells: implications in the extracellular functions of the lectin. Exp Cell Res 313:652–664. doi:10.1016/j.yexcr.2006.11.005
Cay T (2012) Immunhistochemical expression of galectin-3 in cancer: a review of the literature. Turk Patoloji Derg 28:1–10. doi:10.5146/tjpath.2012.01090
Iacovazzi PA, Notarnicola M, Caruso MG, Guerra V, Frisullo S, Altomare DF (2010) Serum levels of galectin-3 and its ligand 90 k/mac-2 bp in colorectal cancer patients. Immunopharmacol Immunotoxicol 32:160–164. doi:10.1080/08923970902936880
Iurisci I, Tinari N, Natoli C, Angelucci D, Cianchetti E, Iacobelli S (2000) Concentrations of galectin-3 in the sera of normal controls and cancer patients. Clin Cancer Res 6:1389–1393
Safaei R, Larson BJ, Cheng TC, Gibson MA, Otani S, Naerdemann W, Howell SB (2005) Abnormal lysosomal trafficking and enhanced exosomal export of cisplatin in drug-resistant human ovarian carcinoma cells. Mol Cancer Ther 4:1595–1604. doi:10.1158/1535-7163.mct-05-0102
Haudek KC, Spronk KJ, Voss PG, Patterson RJ, Wang JL, Arnoys EJ (2010) Dynamics of galectin-3 in the nucleus and cytoplasm. Biochim Biophys Acta 1800:181–189. doi:10.1016/j.bbagen.2009.07.005
Honjo Y, Inohara H, Akahani S, Yoshii T, Takenaka Y, Yoshida J, Hattori K, Tomiyama Y, Raz A, Kubo T (2000) Expression of cytoplasmic galectin-3 as a prognostic marker in tongue carcinoma. Clin Cancer Res 6:4635–4640
Lu T, Ramakrishnan R, Altiok S, Youn JI, Cheng P, Celis E, Pisarev V, Sherman S, Sporn MB, Gabrilovich D (2011) Tumor-infiltrating myeloid cells induce tumor cell resistance to cytotoxic T cells in mice. J Clin Invest 121:4015–4029. doi:10.1172/jci45862
Sinha P, Clements VK, Fulton AM, Ostrand-Rosenberg S (2007) Prostaglandin E2 promotes tumor progression by inducing myeloid-derived suppressor cells. Cancer Res 67:4507–4513. doi:10.1158/0008-5472.CAN-06-4174
Pan PY, Wang GX, Yin B, Ozao J, Ku T, Divino CM, Chen SH (2008) Reversion of immune tolerance in advanced malignancy: modulation of myeloid-derived suppressor cell development by blockade of stem-cell factor function. Blood 111:219–228. doi:10.1182/blood-2007-04-086835
Cao Z, Said N, Amin S, Wu HK, Bruce A, Garate M, Hsu DK, Kuwabara I, Liu FT, Panjwani N (2002) Galectins-3 and -7, but not galectin-1, play a role in re-epithelialization of wounds. J Biol Chem 277:42299–42305. doi:10.1074/jbc.M200981200
Friedrichs J, Manninen A, Muller DJ, Helenius J (2008) Galectin-3 regulates integrin alpha2beta1-mediated adhesion to collagen-I and -IV. J Biol Chem 283:32264–32272. doi:10.1074/jbc.M803634200
Kuwabara I, Liu FT (1996) Galectin-3 promotes adhesion of human neutrophils to laminin. J Immunol 156:3939–3944
Rao SP, Wang Z, Zuberi RI, Sikora L, Bahaie NS, Zuraw BL, Liu FT, Sriramarao P (2007) Galectin-3 functions as an adhesion molecule to support eosinophil rolling and adhesion under conditions of flow. J Immunol 179:7800–7807
Jeon SB, Yoon HJ, Chang CY, Koh HS, Jeon SH, Park EJ (2010) Galectin-3 exerts cytokine-like regulatory actions through the JAK-STAT pathway. J Immunol 185:7037–7046. doi:10.4049/jimmunol.1000154
Correia AL, Bissell MJ (2012) The tumor microenvironment is a dominant force in multidrug resistance. Drug Resist Updat 15:39–49. doi:10.1016/j.drup.2012.01.006
DeNardo DG, Brennan DJ, Rexhepaj E, Ruffell B, Shiao SL, Madden SF, Gallagher WM, Wadhwani N, Keil SD, Junaid SA, Rugo HS, Hwang ES, Jirstrom K, West BL, Coussens LM (2011) Leukocyte complexity predicts breast cancer survival and functionally regulates response to chemotherapy. Cancer Discov 1:54–67. doi:10.1158/2159-8274.cd-10-0028
Acknowledgments
We would like to express special thanks to Dr. Tengfang Zhu for his assistance with sectioning and staining during the immunochemistry experiment. This work was supported by National Natural Science Foundation of China (No. 81000910, 81101551).
Conflict of interest
The authors declare no competing interests.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Wang, T., Chu, Z., Lin, H. et al. Galectin-3 contributes to cisplatin-induced myeloid derived suppressor cells (MDSCs) recruitment in Lewis lung cancer-bearing mice. Mol Biol Rep 41, 4069–4076 (2014). https://doi.org/10.1007/s11033-014-3276-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11033-014-3276-5