
Abstract
The two-line system of hybrid rice seed production based on thermo-sensitive genic male-sterile (TGMS) lines is much more cost-effective, simple and efficient than the three-line system with cytoplasmic male sterility. In this study, we report the introgression of a rice blast resistance gene Pi2 from VE6219 and a bacterial blight (BB) resistance gene Xa23 from HBQ810 into Guangzhan63-4S (GZ63-4S), an elite TGMS rice line, through marker-assisted pyramiding, leading to the development of Hua1015S, a blast- and BB-resistant pyramided line. The newly developed TGMS line Hua1015S and its derived hybrids showed resistance to rice blast and BB. Hua1015S had a similar critical temperature point of fertility–sterility alteration to GZ63-4S and similar key agronomic and grain quality traits. However, Hua1015S gave higher hybrid seed yields on account of increased outcrossing rates due to better floral traits, e.g. spikelet dehiscence, angle of glume opening, and stigma exsertion. The newly developed TGMS parental line Hua1015S with Pi2 and Xa23 is currently being utilized for heterosis breeding of broad-spectrum blast- and BB-resistant two-line rice hybrids and as an improved disease donor source for further TGMS parental line improvement.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Rice blast and bacterial blight (BB) caused by Magnaporthe grisea and Xanthomonas oryzae pv. oryzae (Xoo), respectively, are the two most prevalent and destructive diseases causing significant reduction in rice (Oryza sativa L.) production and grain quality losses (Yi et al. 2004). Rice blast disease has been a major problem in hybrid rice production on account of the narrow genetic base of parents and the high pathogenic diversity over the past 40 years (Liu et al. 2010). Rice blast alone can cause annual yield losses of between 10 and 30 % of the total harvest. Even a yield loss level of 10 % is significant and is sufficient enough to feed 60 million people for 1 year (Skamnioti and Gurr 2009). Blast disease occurrence was reported by the Ministry of Agriculture of China to be as high as 20 % of the hybrid rice fields cultivated in 2006 (Jiang et al. 2012). Significant grain yield losses by BB, especially in hybrid rice, have been reported worldwide (Chen et al. 2000). A grain yield reduction of 20 % has been observed for BB at maximum tillering stage and in severe cases is reduced to as much as 50 % (Ou 1985). Thus it is now imperative to breed rice cultivars with resistance to both blast and BB diseases and this is considered as a priority in most rice improvement programs.
An isogenic line C101A51 was developed in the background of susceptible cultivar CO39 carrying rice blast resistance gene Pi2 at the International Rice Research Institute from a resistant indica rice cultivar 5173 (Mackill and Bonman 1992). As many as six rice blast resistance genes (Pi2, Pi9, Piz-t, Piz, Pigm and Pi50) were identified on the same locus on chromosome 6 short arm close to the centromere (Deng et al. 2006; Hayashi et al. 2004; Liu et al. 2002; Zhu et al. 2012). Pi2 conferred broad-spectrum resistance to diverse M. oryzae isolates as it was found to be resistant to 36 out of 43 blast isolates inoculated and collected from 13 countries (Liu et al. 2002). The cloned Pi2 gene encoded a protein that was associated with a nucleotide-binding site–leucine-rich repeat domain (Zhou et al. 2006). Likewise, another broad-spectrum BB resistance gene, Xa23, was first identified in Oryza rufipogon and was mapped on chromosome 11 (Zhang et al. 1998). In the past, most rice breeding programs widely utilized Xa3, Xa4 and Xa21 to control the damage from BB (Song et al. 1995), but these genes were overcome by the new virulent BB strains in Southern and Yangtze River Valley in China (Ji et al. 2003; Zeng et al. 2002; Zhang 2009). The resistance gene Xa23 is a single dominant resistance gene that was found to be effective at all growth stages, and it conferred broad-spectrum resistance to 20 races representing ten, three and seven from the Philippines, Japan, and China, respectively (Wang et al. 2015; Zhang et al. 2001).
The thermo-sensitive genetic male sterility (TGMS)-based two-line system has comparative advantages over the three-line system, especially on the wider choice of pollen parents for developing sub-specific hybrids, besides simpler breeding and hybrid seed production techniques (Yang et al. 2007). Two-line heterosis breeding is an effective approach for increasing rice grain yields over three-line hybrids in China and tropical Asia; however, there is an urgent need for developing disease-resistant hybrids. Here we report the research work carried out in developing an elite TGMS line with broad-spectrum resistance to blast and BB by molecular pyramiding of Pi2 and Xa23 genes.
Materials and methods
Plant materials
GZ63-4S, a widely used female parent in two-line hybrid rice breeding in China, but susceptible to blast and BB, was used as the TGMS gene donor and recurrent parent. VE6219, a breeding line possessing the broad-spectrum Pi2 gene resistant against rice blast, was used for the blast resistance gene donor parent. HBQ810, a breeding line carrying the Xa23 gene resistant to BB, was used for the BB resistance gene donor parent (Huang et al. 2012).
Molecular markers and PCR amplification
The adoption of marker-assisted selection (MAS) was facilitated by the use of simple sequence repeat (SSR) marker RM527 for Pi2 (Chen et al. 2008), and the Xa23 gene was detected by using insertion/deletion (indel) marker M-Xa23 (Gao et al. 2010). Total DNA was extracted according to the procedure of Dellaporta et al. (1983). PCR reactions were performed on a MyCycler™ thermal cycler (Bio-Rad, USA). Each 20-μl PCR reaction mixture contained 20 ng genomic DNA, 10 mM Tris–HCl (pH 9.0), 50 mM KCl, 2.5 mM MgCl2, 2 mM dNTPs, 10 μM each of the primer pair and 1 U Taq DNA polymerase. Template DNA was initially denatured at 94 °C for 5 min, prior to 35 cycles of denaturation at 94 °C for 30 s, annealing at 55 °C for 30 s and extension at 72 °C for 45 s. In the final step, the reaction mixture was incubated at 72 °C for 5 min before completion. The amplified products were then electrophoretically resolved on 2 % agarose gel for marker M-Xa23, and 4 % polyacrylamide gel for marker RM527 in 1× TBE buffer.
Pyramiding blast and BB resistance genes by MAS
F1 plants with heterozygous alleles of the blast resistance gene (Pi2) and BB resistance gene (Xa23) were obtained from the crosses of GZ63-4S/VE6219 and GZ63-4S/HBQ810, respectively. Molecular markers linked with two resistance genes confirmed heterozygosity and F1s were backcrossed with GZ63-4S. The 40 individual plants in each BC1F1 population were identified to confirm the presence of target genes. Five sterile plants heterozygous at the Xa23 locus in BC1F1 of GZ63-4S*2/HBQ810 and five fertile plants heterozygous at the Pi2 locus in BC1F1 of GZ63-4S*2/VE6219 were selected based on the PCR analysis of linked markers and phenotype during the heading stage. The five sterile plants selected as the female recipients were crossed (multi-cross, MF) with the five selected fertile plants. One hundred plants in MF1 progenies were transplanted. Two sterile plants heterozygous at the Pi2 and Xa23 loci were selected based on the PCR analysis of linked markers and phenotype. The two selected plants were then ratooned and the stubbles were placed under low-temperature conditions (about 21 °C for more than 12 days during the panicle initiation primordial stage). The selfed seeds were harvested from the ratooned stubbles. Furthermore, three sterile plants that were homozygous at the Pi2 and Xa23 loci from 200 MF2 progenies were selected and their selfed seeds were obtained by placing stubble cuts under low-temperature conditions as mentioned earlier. Three family lines of MF3 were planted and 20 plants from each line were identified to confirm them as homozygous at the Pi2 and Xa23 loci (Fig. 1; Supplementary Fig. S1). From these, one promising line closely resembling the recipient parent phenotype was selected and designated as Hua1015S.

The molecular pyramiding scheme for the development of BB- and blast-resistant TGMS lines by the introgression of broad-spectrum BB resistance gene Xa23 (donor parent: HBQ810) and blast resistance gene Pi2 (donor parent: VE6219) into a popular TGMS recipient parent, GZ63-4S, susceptible to both diseases, through MAS strategy (MAS marker-assisted selection, RE resistance evaluation, SE sterility evaluation, FPH field phenotype of hybrids)
Plant materials and blast resistance evaluation
Hua1015S, GZ63-4S (TGMS gene donor), VE6219 (blast-resistance gene donor) and CO39 (blast-susceptible control) were inoculated with 62 isolates of M. oryzae from Zhejiang and Guangdong provinces, China. The leaf resistance levels were identified in greenhouse conditions. Leaf and neck blast resistance were identified under natural conditions by planting Hua1015S, GZ63-4S, VE6219 and CO39 in two rice blast disease hotspot locations, Wangjia village of Yuan-An county and Lianghe village of Enshi city, Hubei province, China, where rice blast diseases are epidemic every year, but the blast-susceptible male parents, C60910, R1005, XH68, FXH1 and Hua15, and their hybrids were planted only in Wangjia village. The disease resistance screening and evaluation standards, both in greenhouse and in natural fields, were carried out according to the standard evaluation system for rice (IRRI 2002).
Plant materials and BB resistance evaluation
Hua1015S was crossed with three BB-susceptible varieties, IR24, Xin613 and Xianhui207. Hua1015S and its derived hybrid combinations were evaluated for BB resistance against seven races of Xoo from China and the Philippines. Of the seven races, ZHE173 (C4) and GD1358 (C5) pathotypes were causing severe BB epidemics in the rice-growing regions of southern China (Huang et al. 2012). PXO61 (Race 1) and PXO99 (Race 6) were from the Philippines, and the other three races, YN24 (R9), FuJ (R8) and HeN11 (R4), were collected from different provinces in southwest China (Liu et al. 2007). All the seven Xoo races were provided by the Plant Protection Department of Huazhong Agricultural University (HAU). Bacterial suspension was prepared following the method described previously (Maruthasalam et al. 2007). Lesion length (LL) was measured 3 weeks after inoculation in the field. The disease symptoms were scored according to the resistance level of Huang et al. (2012).
Characterization of TGMS lines for fertility–sterility alteration in growth chamber
One hundred seeds per line were sown at the nursery plots of the experimental farm of HAU on 10 May 2012. Uniform and healthy rice seedlings at the five-leaf stage (about 25 days after sowing) were selected for transplanting five single plants per hill per plastic pot and each plant was labeled with plastic tags. Care was taken with proper crop management to allow the plants in each plot to grow well and be uniform. Five growth chambers (Model ZSX1500GS; The Shanghai Jing Wins and Scientific Equipment Co., Ltd., China) were adjusted for dry runs, 1 week prior to their actual use for the experiments. Light duration was set to 14 h and relative humidity level to 75 % uniformly in all the five growth chambers while the daily mean temperatures were set at 21, 22, 23, 24 and 25 °C, respectively. The temperature and light duration were carefully programmed to follow diurnal patterns while at the same time achieving the required daily mean temperature and daylight conditions. The plants were placed in growth chambers at stage IV, pistil/stamen primordia-forming stages (Virmani et al. 2003). The plants were moved out of the growth chambers after 12 days of temperature treatment. The pollen grains collected from the top five spikelets of each panicle per plant that headed 516 days after the end of controlled growth chamber environment treatment were observed under the microscope. The pollen sterility of each panicle was recorded according to the classification of pollen morphology on IK-I staining of pollen grains (Virmani et al. 2003). Lines with more than 99.5 % pollen sterility on average were considered completely sterile.
Dynamic observation of pollen fertility–sterility alteration in the field
One hundred and twenty seeds in each line were sown at 15-day intervals from 1 April to 1 July 2012 at the experimental farm of HAU. Forty uniform and healthy rice seedlings at the five-leaf stage were transplanted in fields with a spacing of 20 cm between plants and 25 cm between rows. The pollen fertilities in each line from the top five spikelets of primary panicles were investigated dynamically at 2-day intervals under the microscope from 8 July to 1 October. The pollen sterility data were recorded as the average of five panicles from each line. The daily mean temperature data was provided by the Agricultural Meteorology Department of HAU. Analysis of pollen sterility data in relation to temperature weather charts was carried out to determine the critical temperature point (CTP) of fertility–sterility alteration under natural conditions.
Evaluation of agronomic and rice grain quality traits
Thirty plants each of the newly developed TGMS line Hua1015S and its recipient parent GZ63-4S were transplanted in the field during the winter season of 2011 at the Rice Breeding Station of HAU, Lingshui county, Hainan province, China and the summer season of 2012 at the experimental farm of HAU, Wuhan city, Hubei province, China. Each plot comprised three rows with 10 plants per row at a planting density of 17 cm between plants and 20 cm between rows. The agronomic and rice quality traits were measured according to the standard evaluation system for rice (IRRI 2002). Five individuals in the middle of the second row in each plot were taken for measurements of agronomic traits, including days from sowing to heading, plant height, panicles per plant, panicle length, spikelets per panicle, spikelet fertility and 1000-grain weight. One-way ANOVA was performed to detect statistical differences. A P value of less than 0.05 was considered to be statistically significant. Harvested bulk seeds from each plot were used for analyzing rice grain quality traits that included percentages of brown, milled, head and chalky rice. Rice grain length, grain length-to-width ratio, amylose content (%), gel consistency (mm) and alkali spreading value were also measured.
Evaluation of flowering traits
One hundred seeds of Hua1015S and GZ63-4S were sown in the seedling nursery on 25 April and 20 June 2013, respectively. Thirty rice seedlings at the five-leaf stage were transplanted in fields with a spacing of 17 cm between plants and 20 cm between rows. The first batch was observed for six flowering traits: number of flowering spikelets in a day per panicle, indehiscent spikelet rate, angle of glume opening, stigma exertion rate, unopened panicle portion within flag leaf sheath, and extent of outcrossing (IRRI 2002). The first batch was observed for these six floral traits around the last week of July with the mean temperature above 33 °C, while the second batch was observed at a mean temperature around 28 °C except for extent of outcrossing trait.
Agronomic traits of two-line hybrids
Fourteen hybrids, derived from Hua1015S and GZ63-4S, were planted in three-row plots with ten plants per row and a spacing of 17 × 20 cm in a randomized complete block design with three replications to evaluate agronomic traits. Three individuals in the middle row in each plot were taken for measurements of agronomic traits: heading date (days), plant height (cm), panicles per plant, panicle length (cm), number of grains per panicle, spikelet fertility (%), 1000-grain weight (g) and single-plant yield (g).
Results
Evaluation of blast resistance
Leaf blast resistances of the test materials revealed that Hua1015S was resistant to 56 isolates with a resistance frequency of 90.3 %, similar to the resistance frequency of VE6219 (donor parent). However, the recipient TGMS parent GZ63-4S was resistant to only 26 isolates with a resistance frequency of 41.9 %. The resistance frequency of CO39 (susceptible control) was very low, only 4.8 %. The results showed that the leaf blast resistance levels of the newly developed TGMS line Hua1015S were significantly improved (Supplementary Table S1).
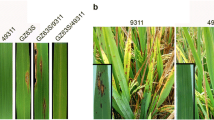
The results of blast evaluations in the rice blast epidemic area in 2 years showed that the resistance scores of leaf blast in Hua1015S were 3 and 4 at Wangjia village of Yuan-An county, Hubei province, China, and 2 and 5 at Lianghe village of Enshi city, Hubei province, China, in 2012 and 2013, respectively, compared to GZ63-4S with scores of 7 and 8 at the same two locations over 2 years. The leaf blast evaluation scores of the six hybrids with Hua 1015S as female parent gave non-significant or similar resistance reactions to Hua1015S, and they also had better resistant reactions than the same set of hybrids with the same male parents but with GZ63-4S used as the female parent. A neck blast incidence of 20 % was observed in the newly developed TGMS line Hua1015S as compared with GZ63-4S, which recorded higher than 90 % at Wangjia village of Yuan-An county, Hubei province, China, in 2012 and 2013. However, the neck blast incidence of Hua1015S was lower than 60 % at Lianghe village of Enshi city, a hotspot for neck blast in Hubei province, as compared to GZ63-4S with 100 % over 2 years (Supplementary Table S2). Overall, the leaf and neck blast resistance levels of the newly developed TGMS line Hua1015S and its derived hybrids in the rice blast epidemic area were significantly improved in comparison to the recipient TGMS parent GZ63-4S and its derived hybrids.
Evaluation of BB resistance
The newly developed TGMS line Hua1015S and its derived hybrids were evaluated for their resistance to BB in the field by inoculating seven strains of Xoo prevalent in China and the Philippines. Two of these strains, ZHE173 and GD1358, were severe epidemic pathotypes in the southern China rice-growing regions. The results of the disease reaction on inoculation with seven Xoo strains showed that GZ63-4S (recipient parent) was susceptible to six Xoo strains, with lesion lengths ranging from 5.8 to 17.9 cm, whereas the donor line HBQ810 with the resistance gene Xa23 was resistant to all strains, with lesion length less than 2.0 cm. However, pyramiding line Hua1015S with Xa23 and Pi2 genes exhibited short lesion lengths, rendering resistance against all seven Xoo strains, with lesion lengths ranging from 0.5 to 3.6 cm.
Two inbred lines, IR24 and Xianhui207, were found to be susceptible to all of the seven Xoo strains tested, with average lesion lengths being 14.6 and 18.8 cm, respectively. As expected, the two derived hybrids with GZ63-4S (as female parent) and IR24 and Xianhui207 (male parents) were susceptible to all seven Xoo strains, with average lesion lengths of 13.1 and 15.7 cm, respectively. However, the two derived hybrids with Hua1015S (as female parent) and IR24 and Xianhui207 (male parents) were resistant or moderately resistant to all seven Xoo strains, with average lesion lengths of 2.6 and 2.1 cm, respectively. Another inbred line, Xin613, was susceptible to five Xoo strains and moderately resistant to two Xoo strains tested, with average lesion length of 10.0 cm. The derived hybrid between GZ63-4S and Xin613 was susceptible to six Xoo strains and moderately resistant to one Xoo strain, with an average lesion length of 13.1 cm. However, the hybrid between Hua1015S and Xin613 was resistant to six Xoo strains and moderately susceptible to one Xoo strain, with average lesion length of 2.3 cm (Table 1). The newly developed TGMS line Hua1015S and its derived hybrids showed significant improvement in BB resistances as compared with its recipient TGMS line GZ63-4S and its derived hybrids.
Based on these results, the newly developed TGMS pyramiding line Hua1015S with Pi2 and Xa23 genes could be fully exploited for developing both blast- and BB-resistant two-line hybrids.
Characterization of fertility–sterility alteration
The CTP of fertility–sterility alteration of the TGMS line has a practical application to safe seed production. The sensitive stage of pollen fertility–sterility alteration to temperature lies between 5 and 16 days after panicle initiation (Virmani et al. 2003). Hua1015S and GZ63-4S were test-verified in a growth chamber for 12 days coinciding with the temperature-sensitive stage during the summer season of 2012. The results clearly showed that GZ63-4S and Hua1015S had similar CTPs of fertility–sterility alteration. Partial fertility was observed in both lines with daily mean temperature (DMT) below 21 °C, but upon exceeding 22 °C the lines became completely male-sterile (pollen sterility was more than 99.5 %) (Table 2). The CTP of Hua1015S was thus determined to be located at 22 °C of DMT. These results clearly demonstrated that Hua1015S could be widely and safely used in two-line hybrid seed production and heterosis breeding programs in China.
The dynamic pollen grain fertility expressions of Hua1015S and GZ63-4S were observed from 8 July to 1 October 2012 under natural field conditions at the experimental farm of HAU, Wuhan, China. Hua1015S followed a similar pattern to GZ63-4S, in being completely male-sterile when flowering from July to the middle of September at Wuhan; however, its duration of complete pollen sterility was 81 days, which was longer than GZ63-4S by 4 days. We also noted that when the DMT during 13–16 September declined to below 22 °C, the pollen grains of Hua1015S and GZ63-4S showed reversal to partial fertility on 27 September and 25 September, respectively (Supplementary Fig. S2). These results indicated that the CTPs of Hua1015S and GZ63-4S were the same under both growth chamber and natural field conditions.
Evaluation of agronomic, rice quality and flowering traits of Hua1015S
Effective comparison between Hua1015S and GZ63-4S for key agronomic and grain quality traits both in the field and laboratory showed non-significant differences for most of the traits, including plant type and grain quality (Table 3). Both the TGMS lines were fertile in the winter season at Hainan and were sterile during the summer season at Wuhan. Likewise, during the winter under fertile phase at Hainan both the TGMS lines had similar traits such as days to heading, plant height, panicle number per plant, number of grains per panicle, 1000-grain weight and four grain quality traits. However, Hua1015S had a shorter panicle length by 1.9 cm but with higher spikelet fertility of 18.8 % over GZ63-4S. Higher spikelet fertility is a desirable trait for increasing TGMS seed multiplication yield. In the summer season at Wuhan, the plant height of Hua1015S was higher than that of GZ63-4S by 4.1 cm. This is an undesirable trait for seed production and reduces lodging resistance. However, Hua1015S gave 8 % more grains per panicle than GZ63-4S, which is a desirable trait for increasing seed production. Desirable floral traits promoting outcrossing in the male-sterile line are important for enhancing the hybrid seed yields. Floral traits of Hua1015S and GZ63-4S were determined by screening them separately both under high (DMT > 33 °C) and normal temperature (DMT ~ 28 °C) conditions in the natural field conditions. Under both temperature conditions, the number of flowering spikelets in a day per panicle of Hua1015S was greater than GZ63-4S, and the indehiscent spikelet rate of Hua1015S was lower than GZ63-4S (Table 4). Hua1015S also had a wider angle of glume opening than GZ63-4S under both high and normal mean temperatures. Under higher temperature conditions, Hua1015S had higher stigma exsertion rate and lower unopened panicle portion within flag leaf sheath than GZ63-4S. With these excellent flowering traits, the outcrossing seed-setting rate with pollen parents was significantly higher in Hua1015S, with 40.2 %, than GZ63-4S, with only 1.0 %. The relative adaptation of Hua1015S is based on its higher hybrid rice seed yield, which is influenced by earlier heading and better floral features than GZ63-4S, despite their similar CTPs.
Evaluation of agronomic traits in hybrid rice
Fourteen hybrid rice combinations were derived from the two TGMS lines, Hua1015S and GZ63-4S, and crossed with the seven pollen parents, C60910, R1005, HH651, YD4, XH68, FXH1 and HH23. In all seven groups compared, the heading dates of Hua1015S hybrid combinations were 0.5–2.5 days shorter than the GZ63-4S hybrid combinations (Supplementary Table S3). Hua1015S/YD4, Hua1015S/FXH1 and Hua1015S/HH23 were slightly taller than the controls, GZ63-4S/YD4, GZ63-4S/FXH1 and GZ63-4S/HH23, respectively (Supplementary Table S3). The spikelet fertility of Hua1015S hybrid combinations did not show any significant difference from the GZ63-4S hybrid combinations (Supplementary Table S3). For the other four agronomic traits, panicles per plant, panicle length, number of grains per panicle and 1000-grain weight, only one or two hybrid combinations showed differences (Supplementary Table S3). Interestingly, two Hua1015S hybrid combinations, Hua1015S/R1005 and Hua1015S/XH68, produced higher single-plant yields than the GZ63-4S-derived hybrid combinations, respectively (Supplementary Table S3).
Discussion
Hybrid rice technologies based on the three-line and two-line systems have greatly contributed to the global increase of rice productivity and production (Khush 2001; Yuan 2004). The three-line system of hybrid rice seed production is cumbersome as it uses cytoplasmic male sterility (CMS), maintainer and restorer lines for developing hybrids (Wang et al. 2006). In the two-line system using a TGMS line the expression of male sterility is governed by a nuclear recessive gene and this in turn is influenced by temperature alterations. Thus, the TGMS line could be used in a dual manner by facilitating the conducive temperature conditions at early panicle initiation stages to become selectively male-sterile, suitable for production of hybrid seeds, and yet in another set of conditions the same line could become male-fertile to produce self-seed multiplication. The two-line system offers simpler, cheaper and more efficient hybrid rice seed production in comparison to the three-line system. Two-line-based hybrids are relatively more diverse, as any non-TGMS variety could be used as a pollen parent, unlike the three-line system, which requires a restorer system. In recent years, two-line hybrid rice is being strengthened for large-scale seed production in China. GZ63-4S had good agronomic traits, high combining ability and heterosis. There are more than 20 two-line hybrids derived from GZ63-4S released in China, and one of them, Yangliangyou 6, was used as the check for national variety traits (Dai et al. 2005), thus leaving most these derived hybrids highly susceptible to rice blast and BB and also hindering their large-scale deployment in this region. This study on MAS-based pyramiding of multiple resistance genes into parental lines of two-line hybrids appears to be a feasible strategy for preventing the disease damage, and several researchers have demonstrated the success of this approach in developing rice restorer lines, maintainer lines or inbred varieties with resistance to rice blast (Hittalmani et al. 2000; Jiang et al. 2012) and BB (Chen et al. 2000; Huang et al. 2012; Luo et al. 2012). Ours is the first report on the pyramiding of two broad-spectrum disease resistance genes for rice blast (resistance gene Pi2) and BB (resistance gene Xa23) into an elite TGMS line GZ63-4S, resulting in the development of an improved TGMS line, Hua1015S, with broad-spectrum resistance to both rice blast and BB. This study also showed the compatibility of pyramiding Pi2 and Xa23 genes without any reduction in resistance reaction levels against both the diseases. It also proved valuable for the improvement of rice blast and BB resistance in hybrid rice seed production, and paved the way for developing blast- and BB-resistant two-line rice hybrids in the future.
Incorporation of rice blast and BB broad-spectrum resistance genes into an elite TGMS line will improve rice blast and BB resistance of the newly derived rice hybrids. Although more than 31 resistance genes to BB have been reported to date, only a few of them have been used in breeding programs, primarily on account of their lower resistance level or narrow resistance spectrum. Of them, Xa4 has been widely used in the development of BB-resistant rice varieties. However, rice hybrids carrying Xa4 for defense against Xoo had become susceptible because of their incompletely dominant nature. Conversely, the Xa23 gene shows broad-spectrum resistance to different BB races from several countries (Zhang et al. 2001), and has been successfully used in breeding programs (Huang et al. 2012; Zhou et al. 2009, 2011). The rice blast resistance gene Pi2 renders broader spectrum resistance to different rice blast races, and was earlier proved to be successful for developing the blast-resistant rice restorer lines or maintainer lines (Hittalmani et al. 2000; Jiang et al. 2012). However, none of these earlier studies had shown MAS for improving the rice hybrids for disease resistance by incorporation of Xa23 and Pi2. We report the newly developed TGMS line Hua1015S with Xa23 and Pi2, and its derived hybrid combinations were resistant to BB races and rice blast. This also indicated that the BB resistance gene Xa23 and rice blast resistance gene Pi2 were completely dominant in expressing resistance in hybrids without showing any influence of a gene dosage effect in the heterozygous state, and thus Xa23 and Pi2 could be considered an ideal resistance gene combination for developing BB- and blast-resistant two-line rice hybrids. Interestingly, Pi2 also conferred resistance to neck blast in our study on TGMS improvement, which was previously observed in maintainer line development (Jiang et al. 2012).
The genetic background may influence R-gene-mediated resistance to Xoo in rice. Among all BB resistance genes, the Xa23 gene derived from wild rice, Oryza rufipogan, showed broad-spectrum resistance and was effective at all growth stages and was also successfully introduced into several rice inbred varieties (Huang et al. 2012; Zhou et al. 2009, 2011), and was resistant to the virulent BB strain GD1358. However, in this study, Hua1015S with Xa23 was resistant or highly resistant to six BB strains and only moderately resistant to GD1358. Likewise, Hua1015S with Pi2 possessed slightly lower levels of resistance to rice blast compared to the donor parent, especially at the two natural hotspot sites.
In a TGMS-based two-line breeding program, the performances of the rice hybrids are based on the level of heterosis, which also includes superior plant type and grain yield. Non-significant differences were observed in five out of seven GZ63-4S- and Hua1015S-derived hybrid combinations for agronomic traits including plant height, panicles per plant and panicle length. However, in six of them Hua1015S-derived hybrids showed increased grain yield while for two hybrid combinations it was significant (Supplementary Table S3). The newly developed Hua1015S is genetically different from GZ63-4S on account of numerous improved traits and therefore possibly shows different levels of specific combining ability, and the observed variations between Hua1015S and GZ63-4S were due to intentional selection for breeding a better TGMS line than its recurrent parent (Luo and Yin 2013; Sundaram et al. 2008). Detailed analysis showed Hua1015S to be an ideal TGMS line that could be immediately used commercially.
The ptgms2-1 gene present in GZ63-4S was derived from GZ63S, which in turn was derived from a Nonken58S source. Xu et al. (2011) discovered the gene as LOC_Os02g12290, responsible for the nuclear ribonuclease Z (RNase Z), and SNP1 at site 71 created a premature stop codon in the Guangzhan63S allele, resulting in sterility. Recently, the tms5 gene was cloned and researchers were able to understand the function of this important locus on chromosome 2 (Fan and Zhang 2014; Zhou et al. 2014).
MAS provides clear insight into the movement of disease resistance genes from both cultivated and exotic germplasm into adaptive genetic backgrounds across generations, which otherwise is often laborious and time-consuming through conventional disease breeding approaches. The efficiency of MAS could be considerably enhanced in combination with phenotype-based selection approaches, including disease resistance evaluation and agro-morphological characterization of the newly developed lines. However, in MAS for parental-line improvement programs, especially concerning heterosis breeding, one should also consider the results of the derived hybrid combinations for maximizing the gains. MAS could be effectively used for corrective plant breeding and be carried out in a routine manner considering its reduced costs and turn-around time. However, there is a word of caution in that MAS should be used for corrective plant breeding purposes with few major genes and should not in any way substitute for conventional rice breeding programs. Many of the MAS products could be ploughed back into conventional breeding programs through crossing programs and by exercising careful selection. The MAS-based pyramiding approach for disease resistance improvement of TGMS lines is very different from that of conventional inbred varieties, as both the parental TGMS lines and their derived hybrids with resistance genes need to demonstrate a higher level of resistance to rice blast and BB. Recessive and incomplete dominance genes are relatively less useful in improving the TGMS lines through conventional MAS approaches, e.g., xa13 (Chu et al. 2006; Li et al. 2012) and pi21 (Fukuoka et al. 2009). Firstly, this study clearly indicated that Pi2 and Xa23 genes are valuable for the improvement of blast and BB resistance in hybrid rice production, respectively. Secondly, the CTP that determines the fertility–sterility alteration of a TGMS line is an important factor for safe hybrid seed production, as any sudden temperature fluctuations stable enough for over 3 consecutive days coinciding with the sensitive stage could jeopardize hybrid seed production. The fertility–sterility alteration behaviors of Hua1015S and GZ63-4S were the same, with a low CTP of 22 °C DMT under growth chamber and natural field conditions. Therefore, Hua1015S could safely replace GZ63-4S for commercial seed production in temperate zones, such as central China. Interestingly, Hua1015S gave better pollen fertility reversion at Hainan with higher self-seed production than GZ63-4S. Thirdly, floral traits are extremely important for safe hybrid seed production, and improper spikelet opening of some of TGMS lines, especially under high-temperature conditions during the summer months in China, could reduce outcrossing rates and thereby give poor seed production. Fortunately, Hua1015S had better floral traits and a higher outcrossing rate than GZ63-4S, especially under high temperatures. In conclusion, the newly developed Hua1015S incorporated with broad-spectrum Xa23 and Pi2 genes can be commercially utilized for two-line hybrid seed production and as a new source for two-line breeding programs to develop superior TGMS lines.
References
Chen S, Li XH, Xu CG, Zhang QF (2000) Improvement of bacterial blight resistance of ‘Minghui 63’, an elite restorer line of hybrid rice, by molecular marker-assisted selection. Crop Sci 40:239–244
Chen HQ, Chen ZX, Ni S, Zuo SM, Pan XB, Zhu XD (2008) Pyramiding three genes with resistance to blast by marker assisted selection to improve rice blast resistance of Jin 23B. Chin J Rice Sci 22:23–27
Chu Z, Yuan M, Yao J, Ge X, Yuan B, Xu C, Li X, Fu B, Li Z, Bennetzen JL, Zhang Q, Wang S (2006) Promoter mutations of an essential gene for pollen development result in disease resistance in rice. Gene Dev 20:1250–1255
Dai Z, Liu G, Li A, Xu M, Liu X, Zhou C, Zhang H (2005) Breeding of two-line indica hybrid rice combination, “Yangliangyou 6”, and studying on its culture characteristics. Chin Agric Sci Bull 21:114–116
Dellaporta SL, Woo J, Hicks JB (1983) A plant DNA mini preparation: version II. Plant Mol Biol Rep 1:19–21
Deng YW, Zhu XD, Shen Y, He ZH (2006) Genetic characterization and fine mapping of the blast resistance locus Pigm(t) tightly linked to Pi2 and Pi9 in a broad-spectrum resistant Chinese variety. Theor Appl Genet 113:705–713
Fan YR, Zhang QF (2014) Understanding a key gene for thermosensitive genic male sterility in rice. Sci China Life Sci 57:1241–1242
Fukuoka S, Saka N, Koga H, Ono K, Shimizu T, Ebana K, Hayashi N, Takahashi A, Hirochika H, Okuno K, Yano M (2009) Loss of function of a proline-containing protein confers durable disease resistance in rice. Science 325:998–1001
Gao LG, Gao HL, Li RB, Li DY, Zhou M, Yan Q, Zhou WY, Zhang J, Deng GF (2010) Optimization and verification of molecular for rice bacterial blight resistance gene Xa23. Mol Plant Breed 8:660–664
Hayashi K, Hashimoto N, Daigen M, Ashikawa I (2004) Development of PCR-based SNP markers for rice blast resistance genes at the Piz locus. Theor Appl Genet 108:1212–1220
Hittalmani S, Parco A, Mew TV, Zeigler RS, Huang N (2000) Fine mapping and DNA marker-assisted pyramiding of the three major genes for blast resistance in rice. Theor Appl Genet 100:1121–1128
Huang B, Xu JY, Hou MS, Ali J, Mou TM (2012) Introgression of bacterial blight resistance genes Xa7, Xa21, Xa22 and Xa23 into hybrid rice restorer lines by molecular marker-assisted selection. Euphytica 187:449–459
IRRI (2002) Standard evaluation system for rice (SES). International Rice Research Institute, Los Banos
Ji GH, Zhang SG, Qian J (2003) Preliminary analysis on races of Xanthomonas oryzae pv. oryzae in Yunnan Province. Plant Prot 29:19–21
Jiang HC, Feng YT, Bao L, Li X, Gao GJ, Zhang QL, Xiao JH, Xu CG, He YQ (2012) Improving blast resistance of Jin 23B and its hybrid rice by marker-assisted gene pyramiding. Mol Breed 30:1679–1688
Khush GS (2001) Green revolution: the way forward. Nat Rev Genet 2:815–822
Li CY, Wei J, Lin YJ, Chen H (2012) Gene silencing using the recessive rice bacterial blight resistance gene xa13 as a new paradigm in plant breeding. Plant Cell Rep 31:851–862
Liu G, Lu G, Zeng L, Wang GL (2002) Two broad-spectrum blast resistance genes, Pi9(t) and Pi2(t), are physically linked on rice chromosome 6. Mol Genet Genomics 267:472–480
Liu H, Yang W, Hu B, Liu F (2007) Virulence analysis and race classification of Xanthomonas oryzae pv. oryzae in China. J Phytopathol 155:129–135
Liu J, Wang X, Mitchell T, Hu Y, Liu X, Dai L, Wang GL (2010) Recent progress and understanding of the molecular mechanisms of the rice–Magnaporthe oryzae interaction. Mol Plant Pathol 11:419–427
Luo YC, Yin ZC (2013) Marker-assisted breeding of Thai fragrance rice for semidwarf phenotype, submergence tolerance and disease resistance to rice blast and bacterial blight. Mol Breed 32:709–721
Luo YC, Sangha JS, Wang SH, Li ZF, Yang JB, Yin ZC (2012) Marker-assisted breeding of Xa4, Xa21 and Xa27 in the restorer lines of hybrid rice for broad-spectrum and enhanced disease resistance to bacterial blight. Mol Breed 30:1601–1610
Mackill DJ, Bonman JM (1992) Inheritance of blast resistance in near-isogenic lines of rice. Phytopathology 82:746–749
Maruthasalam S, Kalpana K, Kumar KK, Loganathan M, Poovannan K, Raja JAJ, Kokiladevi E, Samiyappan R, Sudhakar D, Balasubramanian P (2007) Pyramiding transgenic resistance in elite indica rice cultivars against the sheath blight and bacterial blight. Plant Cell Rep 26:791–804
Ou SH (1985) Rice diseases, 2nd edn. Commonwealth Mycological Institute, Richmond
Skamnioti P, Gurr SJ (2009) Against the grain: safeguarding rice from rice blast disease. Trends Biotechnol 27:141–150
Song WY, Wang GL, Chen LL, Kim HS, Pi LY, Holsten T, Gardner J, Wang B, Zhai WX, Zhu LH, Fauquet C, Ronald P (1995) A receptor kinase-like protein encoded by the rice disease resistance gene, Xa21. Science 270:1804–1806
Sundaram RM, Vishnupriya RM, Biradar SK, Laha GS, Ashok Reddy G, Shobha Rani N, Sarma P, Sonti RV (2008) Marker assisted introgression of bacterial blight resistance in Samba Mahsuri, an elite indica rice variety. Euphytica 160:411–422
Virmani SS, Sun ZX, Mou TM, Ali J, Mao CX (2003) Two-line hybrid rice breeding manual. International Rice Research Institute, Los Banos
Wang ZH, Zou YJ, Li XY, Zhang QY, Chen L, Wu H, Su DH, Chen YL, Guo JX, Luo D, Long YM, Zhong Y, Liu YG (2006) Cytoplasmic male sterility of rice with boro II cytoplasm is caused by a cytotoxic peptide and is restored by two related PPR motif genes via distinct modes of mRNA silencing. Plant Cell 18:676–687
Wang CL, Zhang XP, Fan YL, Gao Y, Zhu QL, Zheng CK, Qin TF, Li YQ, Che JY, Zhang MW, Yang B, Liu YG, Zhao KJ (2015) XA23 is an executor R protein and confers broad-spectrum disease resistance in rice. Mol Plant 8:290–302. doi:10.1093/mp/ssu132
Xu JJ, Wang BH, Wu YH, Du PN, Wang J, Wang M, Yi CD, Gu MH, Liang GH (2011) Fine mapping and candidate gene analysis of ptgms2-1, the photoperiod-thermo-sensitive genic male sterile gene in rice (Oryza sativa L.). Theor Appl Genet 122:365–372
Yang QK, Liang CY, Zhuang W, Li J, Deng HB, Deng QY, Wang B (2007) Characterization and identification of the candidate gene of rice thermo-sensitive genic male sterile gene tms5 by mapping. Planta 225:321–330
Yi G, Lee SK, Hong YK, Cho YC, Nam MH, Kim SC, Han SS, Wang GL, Hahn TR, Ronald PC, Jeon JS (2004) Use of Pi5(t) markers in marker-assisted selection to screen for cultivars with resistance to Magnaporthe grisea. Theor Appl Genet 109:978–985
Yuan L (2004) Hybrid rice technology for food security in the world. Crop Res 18:185–186
Zeng LX, Huang SH, Wu SZ (2002) The resistance of IRBB21 (Xa21) against 5 races of Guangdong Province. Acta Phytopathol Sin 29:97–100
Zhang Q (2009) Genetics and improvement of bacterial blight resistance of hybrid rice in China. Rice Sci 16:83–92
Zhang Q, Lin SC, Zhao BY, Wang CL, Yang WC, Zhou YL, Li DY, Chen CB, Zhu LH (1998) Identification and tagging a new gene for resistance to bacterial blight (Xanthomonas oryzae pv. oryzae) from O. rufipogon. Rice Genet Newsl 15:138–142
Zhang Q, Wang CL, Zhao KJ, Zhao YL, Caslana VC, Zhu XD, Li DY, Jiang QX (2001) The effectiveness of advanced rice lines with new resistance gene Xa23 to rice bacterial blight. Rice Genet Newsl 18:71–72
Zhou B, Qu S, Liu G, Dolan M, Sakai H, Lu G, Bellizzi M, Wang GL (2006) The eight amino-acid differences within three leucine-rich repeats between Pi2 and Piz-t resistance proteins determine the resistance specificity to Magnaporthe grisea. Mol Plant Microbe Interact 19:1216–1228
Zhou YL, Xu JL, Zhou SC, Yu J, Xie XW, Xu MR, Sun Y, Zhu LH, Fu BY, Gao YM, Li ZK (2009) Pyramiding Xa23 and Rxo1 for resistance to two bacterial diseases into an elite indica rice variety using molecular approaches. Mol Breed 23:279–287
Zhou YL, Uzokwe VNE, Zhang CH, Cheng LR, Wang L, Chen K, Gao XQ, Sun Y, Chen JJ, Zhu LH, Zhang Q, Ali J, Xu JL, Li ZK (2011) Improvement of bacterial blight resistance of hybrid rice in China using the Xa23 gene derived from wild rice (Oryza rufipogon). Crop Prot 30:637–644
Zhou H, Zhou M, Yang YZ, Li J, Zhu LY, Jiang DG, Dong JF, Liu QJ, Gu LF, Zhou LY, Feng MJ, Qin P, Hu XC, Song CL, Shi JF, Song XW, Ni ED, Wu XJ, Deng QY, Liu ZL, Chen MS, Liu YG, Cao XF, Zhuang CX (2014) RNase ZS1 processes Ub L40 mRNAs and controls thermosensitive genic male sterility in rice. Nat Commun 5:4884–4892
Zhu X, Chen S, Yang J, Zhou S, Zeng L, Han J, Su J, Wang L, Pan Q (2012) The identification of Pi50(t), a new member of the rice blast resistance Pi2/Pi9 multigene family. Theor Appl Genet 124:1295–1304
Acknowledgments
The research work was supported by Grants from the National High Technology (863) Plan of China (2014AA10A604), Key Technology Research Project of Wuhan City, China (2014020202010136) and the Bill & Melinda Gates Foundation “Green Super Rice for the Resources-Poor of Africa and Asia”.
Author information
Authors and Affiliations
Corresponding authors
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Jiang, J., Yang, D., Ali, J. et al. Molecular marker-assisted pyramiding of broad-spectrum disease resistance genes, Pi2 and Xa23, into GZ63-4S, an elite thermo-sensitive genic male-sterile line in rice. Mol Breeding 35, 83 (2015). https://doi.org/10.1007/s11032-015-0282-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11032-015-0282-9