
Abstract
Widespread ecosystem change has led to declines in species world-wide. The loss of pollinators in particular constitutes a problem for ecosystem function and crop production. Understanding how landscape change affects pollinator movement, effective pollen flow, and plant and pollinator survival is therefore a global priority. In this study we investigated patterns of effective pollen flow, using wild cherry tree (Prunus avium) progeny arrays, to address two questions in three case studies: Do land-use types present different resistances to pollinator movement? Which pollinator travel path best explains the pollination data (straight lines, weighted straight lines, least cost paths or pair-wise resistance)? Trees and progeny arrays were genotyped and effective pollen flow and pollinator movement were estimated using the spatially explicit mating model. We found that pollinators did modify their travel paths in response to land-use type and arrangement, but the travel path that best described pollinator movement and the resistance rank of the land uses depended on the type and size of land-use patches and the landscape context. We propose a novel theoretical framework rooted in behavioural ecology, the resource model, for interpreting pollinator behaviour in heterogeneous landscapes. We conclude by discussing the importance and practicality of conservation and management strategies in which native and non-native land-use types together provide functional habitat and support ecosystem services across economic landscapes.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Habitat degradation, fragmentation and loss are expected to result in genetic isolation of populations, loss of genetic variation, and increased extinction risk (Hagerty et al. 2011), and therefore pose serious threats to the survival of species world-wide (Lavergne et al. 2005; Doerr et al. 2011). Global declines of both wild and managed pollinators (Potts et al. 2010), associated with fragmentation and land-use change (Kremen et al. 2007), constitute a problem not only for the survival of individual plant and pollinator species, but also for continued ecosystem function and crop production (Allen-Wardell et al. 1998). Understanding and managing the ecological impacts of land-use change and fragmentation on pollinator behaviour, plant mating patterns and, ultimately, plant and pollinator survival, is a global priority (Potts et al. 2010). Although there is an increasing amount of information about pollinator movement and patterns of pollen flow through different land-use types (e.g. Lander et al. 2011; Dyer et al. 2012), the theoretical framework for interpreting this data in terms of pollinator behavioural ecology lags far behind our need.
Pollinator movement and plant mating patterns in heterogeneous landscapes
Pollinator movement in heterogeneous landscapes may be considered from two points of view. First, the pollinators move through the landscape, foraging, mating and nesting. Second, pollinators carry pollen through the landscape, shaping the effective mating patterns of plants. Importantly, depending on the distances travelled by the pollinator, the pollinator’s foraging strategy, the plant’s reproductive system and the number of both individual plants and different plant species the pollinator visits, pollinator movement and effective pollen flow may show quite different patterns (Richards et al. 2009). To add additional complexity, both pollinator movement and pollen flow patterns are likely to depend strongly on the land-use types encountered in the landscape (Ricketts 2001; Lander et al. 2011; Cranmer et al. 2012), and may be affected in quite different ways. We suggest that this triple interaction of pollinator movement, effective pollen movement, and land-use type should be the focus of a theoretical framework for understanding and managing both plants and pollinators in heterogeneous landscapes.
Generally, programs to mitigate the potential negative effects of land-use change and fragmentation focus on increasing movement of organisms between habitat patches (Tischendorf and Fahrig 2000), which is expected to increase gene flow between habitat patches, promote colonization of new patches, increase effective population size, and reduce inbreeding (Hagerty et al. 2011). In a strict Island Biogeographical approach, the probability that organisms will move between habitat patches is a function of habitat fragment size and linear distance between fragments, and the land between habitat patches is considered ecologically uniform and hostile (MacArthur and Wilson 1967; Jules and Shahani 2003). However, there is a growing perception that the land outside habitat patches may not be homogenous or hostile (Moilanen and Hanski 1998; Ricketts 2001; Vandermeer and Carvajal 2001), and that the boundaries between land-use types may not actually delimit breeding populations. Against this background, research has shifted to investigating the ease with which study organisms travel through different land-use types outside traditionally defined habitat. Measuring this ‘landscape resistance’ has been an active field of research in recent years (e.g. Spear et al. 2010; Lander et al. 2011). Importantly, although research on organism movement in heterogeneous landscapes is becoming more nuanced, and even though most of these studies focus on animal rather than plant-pollinator systems (Dyer et al. 2012), the rich body of research in animal behavioural ecology remains to be accounted for to develop an overarching theoretical framework to explain observed movement patterns and provide a foundation for predictive models.
Tools for modelling organism movement in heterogeneous landscapes
There are four main models used to characterize organism movement in heterogeneous landscapes and to derive pair-wise distances between locations in the landscape: (1) Straight line paths use the straight Euclidean distance between two points; they do not take into account the land-use types they cross so they do not use resistance values. (2) Weighted linear distances also assume a straight path but measure the distance across each land-use type the line crosses and use the resistance value for each land-use type to compute a resistance distance. Land-use resistance values for this model may be estimated empirically by fitting to observations. (3) A least cost path (LCP) is the non-linear path between two points with the lowest cost based on a defined resistance surface (for discussion of the limitations of LCP see Pullinger and Johnson 2010). (4) Pair-wise resistance is based on circuit theory which quantifies movement between two points as current flow through all possible paths (McRae et al. 2008). The dispersers are equivalent to random walkers who choose a direction for each movement based on the relative resistance of the adjacent cells. For both LCP and pair-wise resistance distances the time and computational power required for land-use resistance value optimization are prohibitive and prevent the estimation of resistance values by fitting to observed data, thus resistance values for these models are currently based on hypothesis.
Investigations of pollinator movement patterns and estimation of land-use resistance values can benefit from the recent developments in molecular tools and statistical techniques to reconstruct patterns of effective pollen dispersal based on paternity analyses. Paternity analyses typically involve genotyping progeny arrays collected on mother plants plus all of the potential father plants within a circumscribed area. The genotypes are then used to identify the putative father of each seed and map the pollen dispersal events. Refined statistical techniques such as the spatially explicit mating model (SEMM, Stoeckel et al. 2012) allow the main factors affecting patterns of effective pollen movement within plant populations to be estimated. In particular, the spatial factors such as the pollen dispersal function and spatial distribution of adults may be separated from the ecological factors affecting fecundity. The SEMM used in this study is based on the mixed effects mating model (http://ciam.inra.fr/biosp/node/120; Klein et al. 2011), modified to account for self-incompatibility (Stoeckel et al. 2012). This modified version is available on request from the authors. Recent studies have used parentage-based tools to investigate patterns of connectivity among individuals (Fortuna et al. 2008) and how different land uses affect inter-individual connectivity (Lander et al. 2011; Dyer et al. 2012). However, to our knowledge, this is the first time these tools have been used to test specific hypotheses about pollinator movement patterns in response to land-use type, and how those patterns affect pollen flow.
Hypotheses about pollinator movement and pollen flow in heterogeneous landscapes
Here we apply existing animal behavioural theory to develop a framework for understanding the triple interaction of pollinator movement, effective pollen movement, and land-use type which we call the resource model (Box 1). The resource model is not concerned with the designation of parts of the landscape as habitat or non-habitat, but rather focuses on the quantity and accessibility of resources across the landscape. Under the resource model the entire landscape becomes a patchwork of ‘partial habitats’ of varying quality (e.g. Kremen et al. 2007). Within the resource model there are four hypotheses about pollinator movement between two points A and B: (H1) Trap-line: the presence of resources encourages pollinator movement from A to B and effective pollen movement may be low or high depending on the numbers and species of plants visited. Pollinator movement is best modelled by straight lines or weighted linear distances because this foraging behaviour is defined by directional movement. (H2) Circe principle: the presence of resources discourages pollinator movement because foraging takes place locally and effective pollen movement is low. Pollinator movement is best modelled by straight line or weighted linear distances because the pollinator is reacting to the land uses between A and B by restricting foraging distance. (H3) Search for optimal environments: the absence of resources encourages pollinator movement and effective pollen movement is high. Pollinator movement is best modelled by weighted linear distances because the pollinator is reacting to the land uses between A and B by searching for more optimal environments in a directed manner. (H4) Avoidance of suboptimal environments: the absence of resources discourages pollinator movement and effective pollen movement is low. Pollinator movement is best modelled by LCPs or pair-wise resistance because the pollinator is avoiding the land uses between A and B but not restricting foraging distance (Box 1).
In this study we investigated patterns of effective pollen flow, using SEMM analysis of wild cherry (Prunus avium L., Rosaceae) progeny arrays. Wild cherry is frequently found growing as a minor component of oak, ash and beech woodlands. In addition, it is used extensively in Europe for the afforestation of agricultural lands and it is also valued for wildlife and amenity plantings (Russell 2003). Wild cherry is thus an interesting model species to study the effects of landscape heterogeneity on realized patterns of pollination. Here, we address the following two questions in three case studies:
Q1
Which travel path type best explains the pollination data?
Q2
Do land-use types present different resistances to pollinator movement?
Below we detail the hypotheses specific to questions 1 and 2 (Q1 and Q2) for each of the three case studies. The resulting analyses allowed us to test the explanatory value of the four hypotheses of the resource model (Box 1).
Case study 1: continuous forest
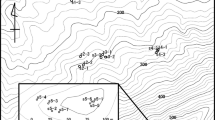
The forest at Saint Gobain was sampled in two successive years, 2002 and 2003, before and after partial clearfelling. In 2002, before clearfelling, the site was a continuous block of forest surrounded by agricultural land and forest (Fig. 1).

Top left the study sites in northern France. Top right Saint Gobain, the site for case studies 1 and 2, located north-east of Paris. In case study 1 the area shown in the top right figure as clearfell was a continuous part of the forest area. Bottom right Neuillé, the site for case study 3, located south-west of Paris. The trees indicated on the maps of the two study sites are the individual P. avium trees sampled for this study
Q1
Pollinator movement should conform to H1-Trap-line or H2-Circe Principle because there are abundant resources and no apparent suboptimal environments. Straight line travel paths or weighted linear distances should have the greatest explanatory power.
Q2
Under H1-Trap-line forest should present lower resistance to pollinator movement than the other land-use types; however under H2-Circe Principle forest should present high resistance.
Case study 2: forest divided by clearfell
In 2003, after clearfelling, the Saint Gobain site contained a ‘reverse-C’-shaped block of forest with an area of clearfell in the centre. The forest area was surrounded by agricultural land and forest (Fig. 1).
Q1
Pollinator movement should conform to H2-Circe Principle, H3-Search or H4-Avoidance. Under H2-Circe Principle, pollinators should show restricted travel distances because of abundant local resources in the forest and straight lines or weighted linear distances should have the greatest explanatory power. Under H3-Search, weighted linear distances should have greatest explanatory power because pollinators would be moving through the low-resource clearfell with higher probability than they would be moving through the forest area. Under H4-Avoidance, LCPs or pair-wise resistance should have greatest explanatory power because pollinators would be modifying their travel paths to avoid the low-resource clearfell, but not necessarily reducing foraging distances.
Q2
Under both H2-Circe Principle and H3-Search the clearfell should present low resistance to pollinator movement and the forest should present high resistance to pollinator movement; however under H4-Avoidance the clearfell should present high resistance.
Case study 3: mixed forest and agriculture
Neuillé, the second study site was dominated by agriculture but included small patches of native forest and urban land uses (Fig. 1).
Q1
If the agricultural areas offer resources, H1-Trap-line or H2-Circe Principle should provide the best explanation of pollinator behaviour, and straight line travel paths or weighted linear distances should have greatest explanatory power. However, if the agricultural areas are suboptimal environments pollinators should behave according to H3-Search or H4-Avoidance. Under H3-Search weighted linear distances have greatest explanatory power because pollinators would be moving through the agricultural areas linearly and with high probability. Under H4-Avoidance LCPs or pair-wise resistance should have greatest explanatory power because pollinators would modify their travel paths to avoid the agricultural areas.
Q2
Under H1-Trap-line or H3-Search the agricultural areas should present low resistance to pollinator movement, however under H2-Circe Principle or H4-Avoidance the agricultural areas should present high resistance.
Methods
Study species
Wild cherry (P. avium) is an early successional tree native across western Eurasia This light demanding, relatively short-lived species is frequently found on woodland edges and glades, in semi-natural and disturbed forests as well as in agricultural landscapes. The species reproduces both asexually through root-suckering and sexually through insect pollination, and has a well-documented gametophytic self-incompatibility system (Vaughan et al. 2008).
Prunus spp. have been found to be pollinated by a suite of generalist pollinators including: Hymenoptera (bees; Guitian et al. 1993), Diptera (true flies; Fortuna et al. 2008), and Lepidoptera (moths and butterflies; Jacobs et al. 2009). Although specific pollinator collection was not undertaken for this study, we expect P. avium at our study sites to be pollinated by generalist insects.
Study sites and sampling
This study was conducted at two sites: Saint Gobain, a patch of continuous forest sampled before and after clearfelling and Neuillé, a mixed forest and agricultural site (Fig. 1). Saint Gobain is a 1.5 km2 area of semi-natural forest in northern France (Aisne, 49°37′N, 3°26′E), within which a central compartment was clearfelled after P. avium flowering season in 2002. The forest contains a variety of tree species, including oaks (Quercus spp.), European beech (Fagus sylvatica), ash (Fraxinus excelsior) and maples (Acer spp.), and the understory is mainly dense blackberry (Rubus fruticosus) and bracken (Pteridium aquilinum). The forest area is bordered by agriculture. Neuillé is a 9 km2 area in northeastern France (Indre-et-Loire, 47°34′N, 0°34′E) dominated by agriculture but including small patches of semi-natural forest and urban land uses. The forest patches contain mainly European beech, hazel (Corylus avellana), ash, frisia (Robinia pseudoacacia), wild cherry and limited understory vegetation.
Land-use maps for both Saint Gobain and Neuillé (grid cell size 10 m × 10 m) were generated in ArcView v.3.2 (ESRI, Redlands, CA) based on aerial photographs. The Neuillé site was categorized into three land uses: forest, urban (roads, railways, houses and industry) and agriculture. The Saint Gobain site was also categorized into three land uses: forest, clearfell, and agriculture. Although there were roads near to the Saint Gobain site they fell outside of the area used for analysis. All forest patches at both sites were subdivided into core-forest and forest-edge (50 m inside the forest edge; Ries et al. 2004).
At Saint Gobain in June 2002, before clearfelling, we sampled 249 adult trees with diameter at 1.3 m above the ground (dbh) of 7–78 cm (median = 42 cm) and 937 seeds from 24 mother trees (18–74 seeds per tree, median = 35). In June 2003, after clearfelling, we made a second collection at Saint Gobain of 877 seeds from 26 mother trees (1–129 seeds per tree, median = 31). At Neuillé in June 2003 we sampled 324 adult trees with dbh 4–49 cm (median = 16.6 cm) and 631 seeds from 31 mother trees (19–32 seeds per tree, median = 20). All adult trees’ locations were taken using a Global Positioning System (Trimble, Sunnyvale, CA) (see Stoeckel et al. 2012).
Genotyping
Total genomic DNA was extracted from frozen leaves (adults) and seeds using the QIAGEN DNeasy 96 plant kit. Both seeds and adults were genotyped at seven microsatellite loci: UDP96-005 (Cipriani et al. 1999); BPPCT034, BPPCT040 (Dirlewanger et al. 2008); PCEGA34 (Downey and Iezzoni 2000); UDP98-021, UDP98-411 (Testolin et al. 2000); and PS12A02 (Joobeur et al. 2000) (see Stoeckel et al. 2006). Self-incompatibility genotypes were determined using three pairs of consensus primers (Sonneveld et al. 2003). Probabilities of identity within both populations provided sufficient power to discriminate individuals (Pid sib (Neuillé) = 2 × 10−3; Pid sib (Saint Gobain) = 1.5 × 10−3).
The spatially explicit mating model
The SEMM (Stoeckel et al. 2012) was used to estimate effective pollen movement based on the mapped locations of all reproductively mature trees in the study populations, the multilocus genotypes of the seeds and adults, and the allele frequencies of P. avium in surrounding populations, which were assumed to be similar to that of the study populations. As in all studies which use effective pollination as a proxy for pollinator movement, some pollinator journeys through the landscape will not be represented in the seed-derived data. If the pollinator is travelling for reasons other than foraging, if the pollinator forages but does not effectively pollinate the flower(s) visited, or, in the case of plant species with self-incompatibility mechanisms such as P. avium, if the pollinator travels between individuals which are not compatible, no record of the pollinator’s movement will be recovered in the analysis of seed parentage. However, because the technology currently available for direct pollinator tracking is limited both to larger pollinating species and in the number of individuals which may be tracked (Zeller et al. 2012); indirect estimates of pollinator movement are currently the best available option for this type of study.
The SEMM considers that a given seed i collected on mother tree (seed tree) j may be the result of pollen flow from outside the neighbourhood N with probability m, or of pollination by any sampled father tree (pollen donor) k with probability 1−m. The contribution of each sampled father tree k to the pollen cloud above mother tree j, π jk , is modelled as:
where F k is is the fecundity of the father k modelled as exp(β1 × DBHk), where DBHk is the diameter of father tree k and β1 is a parameter to be estimated. SI jk is the probability that a pollen grain from father tree k is compatible with the mother tree j at the SI locus. The value of SI jk is 0.0, 0.5 or 1.0 if k and j have 2, 1 or 0 alleles in common at the S-locus respectively. θ jk is derived directly from the pollen dispersal kernel, the probability that a pollen grain disperses from father k to mother j. We chose an exponential power function to model the pollen dispersal kernel, reflecting the higher probability of short distance compared to long distance pollen dispersal (Cottrell et al. 2009; Jolivet et al. 2012; Stoeckel et al. 2012):
where a is a scale parameter homologous to distance, b is a shape parameter affecting the tail of the dispersal kernel, d jk is the distance between father k and mother j, and Γ is the Gamma function.
All of the parameters (complete list in Table 1) were estimated by maximising the following Log likelihood over all of the sampled offspring:
where T is the Mendelian likelihood of a given genotype g o for an offspring given the genotypes g jo of the mother j o and g k of the candidate fathers k (transition probabilities following Meagher (1986) and modified in Stoeckel et al. (2012) to account for the SI). BAF stands for Background Allelic Frequencies, estimated as 75 % of the allelic frequencies of the adults in the dataset (keeping only one tree per clone) and 25 % of the allelic frequencies in the paternal contribution for the genotyped seeds. The likelihood was numerically maximised following a quasi-Newton algorithm in Mathematica 8.1 (Wolfram Research, Champaign, USA). The results of the SEMM analyses were compared based on Akaike’s information criterion (AIC), where a difference of more than two indicated substantially more support for the superior model (Burnham and Anderson 1998). The second-derivatives of the log-likelihood function were used to compute asymptotic Gaussian confidence intervals at the 95 % level.
Q1: Which travel path type best explains the pollination data?
To investigate which travel path had the greatest explanatory power for the pollination data, we used AIC to compare Models I, II, III and IV (Table 1):
-
Model I where d jk in Eq. 2 was computed as the straight line distance d SLD . Straight line distance is the classical Euclidian distance and was measured as the straight line paths between all pairs of seed tree plus possible pollen donor tree using the distance/bearing extension for ArcViewGIS v.3.2.
-
Model II where d jk was computed as the weighted linear distance d WSLD (wA, wF, wE, wU or wC) where wA was the weight (probability of pollinator travel) associated with agricultural land, wF for core-forest, wE for forest-edge, wU for urban land uses at the Neuillé site and wC for clearfell at the Saint Gobain site (Table 1). For Saint Gobain in 2002, before clearfelling, we computed the weight of the straight line across the area to be clearfelled, although at the time it was part of the continuous forest, to compare with the estimates made after the area was clearfelled in 2003. In the text this area is referred to as ‘future clearfell’. The weighted linear distance was obtained by intersecting the straight line paths with the underlying land-use map using the X-Tools extension for ArcViewGIS v.3.2. This process subdivided the straight line path into discrete sections of each land-use type crossed and a weighted sum of the lengths of these sections provided d WSLD (wA, wF, wE, wU or wC). The weights/resistances associated with each land-use type (wA, wF, wE, wU or wC) were parameters of the SEMM model II and were estimated jointly with other parameters (Tables 1, 2). The core-forest resistance was fixed at 1.0 to act as the reference land use. Resistance values <1.0 indicate land uses where effective pollen movement is favoured compared to movement through core-forest.
Table 2 The resistance surfaces used in the least cost path and pair-wise resistance analyses (Models III and IV in Table 1) -
Model III where d jk in Eq. 2 was computed as the LCD d LCD (wA, wF, wE, wU or wC). The LCDs for all mother-father pairs were calculated using Pathmatrix 1.1 (Ray 2005) which provides the distance along the LCP. We also calculated the LCP length weighted by the land-use resistance values (weighted LCP) using the gdistance package for R (van Etten 2012), but these data provided a worse fit for the pollination data in the SEMM than did the LCP length (data not shown) and so are not presented.
-
Model IV where d jk was computed as resistance distance d RD (wA, wF, wE, wU or wC). The pair-wise resistance distances for all mother-father pairs were calculated in Circuitscape v.3.5 (McRae et al. 2008).
For both the LCD and resistance distance analyses the computer processing power and time necessary to estimate the resistance coefficients for the four land uses was prohibitive. SEMM Models III and IV were therefore fitted using three resistance surfaces whose values were estimated based on: (1) the results of the weighted linear distance analysis, (2) strict island biogeography, and (3) modified island biogeography (Table 2).
-
(1)
Based on the results of the weighted linear distance analysis The land-use resistance values estimated in the SEMM weighted linear distance analysis (Model II above) were used to rank the land uses in terms of relative resistance to pollen movement (Table 2). The least resistant land use was assigned a weight of 1 and the most resistant was assigned a weight of 4 (Table 2). Relative resistance values were used because previous work has shown that LCP modelling is robust to uncertainty in resistance values if the rank is correct (Richard and Armstrong 2010).
-
(2)
Strict island biogeography Core-forest was low resistance (weight = 1) and all other land uses were equal and high resistance (weight = 4) (Table 2).
-
(3)
Modified island biogeography The more apparently ecologically similar a land use was to forest, the less resistant it was to pollinator movement. The least resistant land use was assigned a weight of 1 and the most resistant was assigned a weight of 4 (Table 2).
Q2: Do pollinators perceive differences in land-use types?
To investigate whether pollinators perceive differences in land-use types we used the SEMM to optimize the land-use resistance values for the weighted linear distances in Model II or used the LCP resistance surface with the best AIC score from Model III, depending on whether weighted linear distances or LCP had greater support in the SEMM.
Results
Question 1: Which travel path type best explains the pollination data?
Case study 1: continuous forest (Saint Gobain 2002)
The explanatory powers of straight line and weighted linear distance paths were not significantly different from each other, but both out-performed LCP and pair-wise resistance (Table 2).
Case study 2: forest divided by clearfell (Saint Gobain 2003)
The explanatory power of straight line paths was greater than that of the weighted linear distances, LCP and pair-wise resistance (Table 3).
Case study 3: mixed forest and agriculture (Neuillé)
The explanatory power of weighted linear distance was greater than that of the straight line, LCP and pair-wise resistance (Table 3). It should be noted that although the AIC suggests that the weighted linear distance provides more explanatory power than the straight line distance, the confidence intervals do not detect any significant difference between the resistance values. This is likely to be because the confidence intervals are constructed for each parameter separately, whereas the estimators can be highly correlated.
Question 2: Do pollinators perceive differences in land-use types?
Case study 1: continuous forest (Saint Gobain 2002)
The weighted linear distance analysis found that future-clearfell (part of the continuous forest area in this case study) was least resistant, followed by core-forest and forest-edge, and agriculture was most resistant to pollinator travel (Table 4). Because the explanatory power of the straight line and weighted linear distances were not significantly different from each other, the differences in resistance between land-use types may not have been significant.
Case study 2: forest divided by clearfell (Saint Gobain 2003)
The success of the straight line paths suggests that in this case study pollinators did not modify their travel paths in response to land-use type.
Case study 3: mixed forest and agriculture (Neuillé)
The weighted linear distance analysis found that agriculture was the land use least resistant to pollinator movement, followed by forest-edge, and core-forest, and urban land was most resistant to pollinator movement (Table 4).
It should be noted that although the effects of land-use type were found to be significant in the SEMM, the other factors included in the model, such as tree dbh and SI genotype, also had a strong influence on pollination probability (see Stoeckel et al. 2012).
Discussion
In this study we used three case study landscapes to explore how pollinator behaviour may vary in response to landscape composition and pattern, and how this affects pollen flow. We found that both the travel path that best described pollinator movement and the resistance rank of the land-use types in the landscape depended on the type and size of land-use patches in the study area and the landscape context. Based on our results we developed the Resource Model, a novel theoretical framework rooted in animal behavioural ecology, to explain pollinator and pollen movement patterns in heterogeneous landscapes.
Question 1: Which travel path type best explains the pollination data?
In response to Question 1 we found that in the resource-rich, ecologically rather homogeneous continuous forest of Case study 1 and in the highly fragmented, agriculture-dominated landscape of Case study 3 straight lines and weighted linear distances best described pollinator movement. These straight paths are shorter than either LCPs or pair-wise resistance distances and so energetically more efficient for the pollinators. We would therefore expect straight paths to be the default movement pattern unless there is a specific driver for deviation from the straight path. In the continuous forest of Case study 1 there was apparently no ecological driver for deviation from straight paths. In Case study 3 the patches of forest and urban areas were small and scattered across the large area of agriculture, so pollinators that travelled outside of forest typically only crossed agriculture until they re-entered forest. Apparently differences in crop types or management regimes between the agricultural fields in the study area were not sufficient to cause pollinators to make detectable deviations from their straight paths between forest patches. Thus, we find from Case studies 1 and 3 that landscapes dominated by a single land use, even if they include small patches of other land uses, are most frequently traversed by straight paths, even when land uses vary in their resistance to pollinator movement.
In contrast, in Case study 2 unweighted straight lines best described pollinator behaviour. We suggest that although the clearfell in the centre of the study area may not have offered many of the habitat services available in the core-forest, there are likely to have been annual flowering plants which would have provided foraging opportunities. In this case pollinators would have been foraging in both core-forest and clearfell, and any difference in the use of these two land uses was too small to be detected in our analyses. In addition, only 33 % of the 6,449 possible travel paths connecting potential parental tree pairs included the clearfell (note that this is not detected pollination events but rather lines connecting all potential mating pairs), thus there was relatively low statistical power to detect a significant difference in pollinator movement through these land uses. Overall this result suggests that various land-use types may offer foraging opportunities or other habitat services, and pollinators may use these different land uses equally, or for different purposes, but in a way that does not allow us to detect variation in land-use resistance.
Question 2: Do land-use types present different resistances to pollinator movement?
In response to Question 2 we found that in Case studies 1 and 3 path types which depended on differences in resistance between land-use types, specifically weighted straight lines, provided either equal or superior description of observed pollen movement compared to un-weighted straight line paths. Although it is inevitable that the performance of weighted straight lines would be equal or superior to straight lines in terms of description of the observations, because straight lines are a particular case of weighted straight lines where all weights are equal, this result is not inevitable when applying AIC as a method for model selection because AIC measures both the quality of description of the observations and the simplicity of the model in terms of parameters used. This result therefore suggests that land uses do differ in their resistance to pollinator movement. Importantly, the relative resistances of the different land uses varied between case studies (Table 4). We suggest that differences in land-use resistances between years and sites were due to: (1) the size and distribution of patches of land uses; specifically whether there was a dominant land use covering the majority of the study area, even if there were scattered small patches of other land uses, such that pollinators’ travel paths generally crossed only one land use (Case studies 1 and 3). (2) Similar resources may have been available in the land uses across the study area which may mean that land uses that appeared different had functional similarities from the pollinator’s point of view (Case study 2). (3) There may have been differences in the ecology of patches that were grouped together into broad land-use categories. This would have been particularly relevant in relation to agricultural areas, which are likely to change annually in terms of crops and management regimes, and ecological differences between areas broadly categorized as core-forest. (4) Finally, we suggest that pollinator behaviour is a response to the landscape as a whole rather than individual patches in isolation; for example, a pollinator may have avoided entering or moving through agricultural areas surrounding a large central patch of core-forest in Case study 1, but moved through agricultural areas quickly to arrive at small core-forest patches in Case study 3.
Value of the resource model as a theoretical framework
Based on the results of the analyses for Questions 1 and 2 we were able to test the explanatory value of the four hypotheses of the resource model (Box 1) in each of the three case studies:
Case study 1: This study site contained a large central area of resource-rich core-forest. We predicted that pollinators would behave according to H1-Trap-line, with pollinators travelling linearly through the core-forest to collect resources, or H2-Circe Principle, with pollinators showing restricted travel distances because of abundant local resources. The success of the straight line and weighted linear distances, the low resistance of core-forest, and the high resistance of agriculture were consistent with H1-Trap-line behaviour within the core-forest. Under H2-Circe Principle we would have expected core-forest to show high resistance because of reduced travel distances through the high resource area.
Case study 2: This study site contained a ‘reverse-C’-shaped block of core-forest around a central area of clearfell. We predicted that pollinators would behave according to H2-Circe Principle, with pollinators showing restricted travel distances because of abundant resources in the core-forest; H3-Search, with pollinators moving quickly through the clearfell area in search of resources; or H4-Avoidance, with pollinators actively avoiding the clearfell area. The success of the straight lines was somewhat consistent with H3-Search; although rather than clearfell showing low resistance we find that pollinators were ‘searching’ for resources across the study landscape and apparently did not distinguish between core-forest and cleafell. Under H2-Circe Principle or H4-Avoidance we would expect a significant effect of land-use type on pollinator paths, and specifically high resistance of core-forest or clearfell respectively.
Case study 3: This study area was dominated by agriculture but included small patches of core-forest and urban land uses. We predicted that if the agricultural areas offered abundant resources pollinators would behave according to H1-Trap-line or H2-Circe Principle, but if the agricultural areas were suboptimal environments pollinators would behave according to H3-Search or H4-Avoidance. The success of the weighted linear distance, the low resistance of agriculture, and the high resistance of core-forest were consistent with H2-Circe Principle, assuming core-forest was resource-rich, as well as H3-Search, if agriculture was resource-poor, or H1-Trap-line if agriculture was resource-rich. Under H4-Avoidance we would have expected either LCPs or pair-wise resistance to provide the best explanation of the observed pollen movement. Our inability to clearly identify the drivers of pollinator movement patterns in the agricultural areas in this case study is due to our lack of information about resource availability in those areas. Depending on the crop choice and management regime agricultural areas can be extremely rich foraging areas, if temporally limited, but those same areas can also be resource poor and hazardous. This suggests that researchers or conservation and management practitioners wishing to identify areas of high resistance to pollinator movement cannot rely on broad land-use categories such as ‘agriculture’, and must instead specifically investigate the resource availability and the potential threats in the landscape of interest.
It is important to note that wild cherry at our study sites is likely to be pollinated by a suite of generalist insects. Thus the pollen movement we observed is probably the result of movement of different insect species across the study landscape. Dyer et al. (2012) notes that different pollinating species, and individuals within a species, may react to the landscape in different ways. However there are likely to be particular paths which are used disproportionately often, and thus studies that include a large sample of pollination events are likely to capture the trends in pollinator use of the landscape, if not the behaviours unique to certain species or individuals. Our study therefore is likely to represent the results of the majority of pollination events, rather than the entirety or the extremes.
Implications of our results for management and conservation
In this study we found that enough detected pollination events occur between trees separated by land uses other than core-forest that overall, based on our model, the land-use types in a pollinator’s travel path can significantly affect the probability of pollination. Importantly, it cannot be assumed that even nearest-neighbour mating occurs within core-forest. Depending on population density, the shape of the forest patch, the presence of individuals outside of the forest in agricultural features such as hedgerows, and for this and many other species, the pattern of clonal spread; the nearest compatible neighbour may be accessed across a variety of non-core-forest land uses. This result is of clear interest to conservation and management professionals concerned with pollination between small and spatially isolated patches of forest.
Putting our results in the broader context, few studies to date have investigated how the interaction of pollinator and land-use type in heterogeneous landscapes might affect effective pollen movement. Moreover, of the existing studies, none that we know of has been conducted at more than one study site. Thus, the additional complexities of ecological variation between patches that are grouped together in broad land-use categories, and the effect of landscape context on pollinator response to a particular land-use patch, have not been captured before. We suggest therefore that current models of pollen and pollinator movement in heterogeneous landscapes are oversimplified, in part because of the paucity of data, but also because until now there has not been an attempt to set our understanding of pollinator use of heterogeneous landscapes within the context of behavioural theory.
Given the rapid pace of land-use change we need to develop and apply meaningful models and practical management strategies to ensure the survival of plants, pollinators and the organisms that depend upon them. Although native habitat is clearly important and is likely to provide some resources not available in other land-use types (Rands and Whitney 2011), depending exclusively on native habitat to provide ecosystem services is impractical. Understanding how economic landscapes may be designed and managed so that the native and non-native land-use types together provide functional habitat and support ecosystem services may be a useful basis for developing landscape management strategies which will be more likely to be implemented and maintained, more effective biologically, and less expensive than traditional conservation based on nature reserves.
References
Allen-Wardell G, Bernhardt P, Bitner R, Burquez A, Buchmann S, Cane J, Cox PA, Dalton V, Feinsinger P, Ingram M, Inouye D, Jones EC, Kennedy K, Kevan P, Koopowitz H, Medellin R, Medellin-Morales S, Nabhan GP, Pavlik B, Tepedino V, Torchio P, Walker S (1998) The potential consequences of pollinator declines on the conservation of biodiversity and stability of food crop yields. Conserv Biol 12(1):8–17
Bell W (1990) Searching behaviour patterns in insects. Annu Rev Entomol 35:447–467
Burnham KP, Anderson DR (1998) Model selection and inference: a practical information-theoretic approach. Springer-Verlag, New York
Carvell C, Jordan WC, Bourke AFG, Pickles R, Redhead JW, Heard MS (2012) Molecular and spatial analyses reveal links between colony-specific foraging distance and landscape-level resource availability in two bumblebee species. Oikos 121(5):734–742
Cipriani G, Lot G, Huang WG, Marrazzo MT, Peterlunger E, Testolin R (1999) AC/GT and AG/CT micosatellite repeats in peach (Prunus persica (L.) Batsch): isolation, characterization and cross-species amplification in Prunus. Theor Appl Genet 99:65–72
Cottrell JE, Vaughan SP, Connolly T, Sing L, Moodley DJ, Russell K (2009) Contemporary pollen flow, characterization of the maternal ecological neighbourhood and mating patterns in wild cherry (Prunus avium L.). Heredity 103:118–128
Cranmer L, McCollin D, Ollerton J (2012) Landscape structure influences pollinator movements and directly affects plant reproductive success. Oikos 121(4):562–568
Dirlewanger E, Capdeville G, Tauzin Y, Cosson P, Claverie J (2008) A Sweet Cherry (Prunus avium L.) linkage map and its comparison to other Prunus species. In: Burak M, Lang GA, Gulen H, Ipek A (eds) Proceedings of the Vth international cherry symposium, vols 1 and 2, Acta Horticulturae. pp 115–126
Doerr VAJ, Barrett T, Doerr ED (2011) Connectivity, dispersal behaviour and conservation under climate change: a response to Hodgson et al. J Appl Ecol 48(1):143–147
Downey S, Iezzoni A (2000) Polymorphic DNA markers in black cherry (Prunus serotina) are identified using sequences from sweet cherry, peach, and sour cherry. J Am Soc Hortic Sci 125:76–80
Dyer RJ, Chan DM, Gardiakos VA, Meadows CA (2012) Pollination graphs: quantifying pollen pool covariance networks and the influence of intervening landscape on genetic connectivity in the North American understory tree, Cornus florida L. Landsc Ecol 27:239–251
Fortuna MA, Garcia C, Guimaraes PR, Bascompte J (2008) Spatial mating networks in insect-pollinated plants. Ecol Lett 11(5):490–498
Guitian P, Guitian J, Navarro L (1993) Pollen transfer and diurnal versus nocturnal pollination in Lonicera etrusca. Acta Oecol Int J Ecol 14(2):219–227
Hagerty BE, Nussear KE, Esque TC, Tracy CR (2011) Making molehills out of mountains: landscape genetics of the Mojave Desert tortoise. Landsc Ecol 26(2):267–280
Hodges C, Miller R (1981) Pollinator flight directionality and the assessment of pollen returns. Oecologia 50:376–379
Jacobs JH, Clark SJ, Denholm I, Goulson D, Stoate C, Osborne JL (2009) Pollination biology of fruit-bearing hedgerow plants and the role of flower-visiting insects in fruit-set. Ann Bot 104(7):1397–1404
Jolivet C, Holtken AM, Liesebach H, Steiner W, Degen B (2012) Mating patterns and pollen dispersal in four contrasting wild cherry populations (Prunus avium L.). Eur J For Res 131:1055–1069
Jones E, Dornhaus A (2011) Predation risk makes bees reject rewarding flowers and reduce foraging activity. Behav Ecol Sociobiol 65:1505–1511
Joobeur T, Periam N, Vicente M, King G, Arus P (2000) Development of a second generation linkage map for almond using RAPD and SSR markers. Genome 43:649–655
Jules ES, Shahani P (2003) A broader ecological context to habitat fragmentation: why matrix habitat is more important than we thought. J Veg Sci 14(3):459–464
Klein EK, Carpentier FH, Oddou-Muratorio S (2011) Estimation of the whole variance of male fecundity from genotypes of progeny arrays: accuracy and limits of the Bayesian forward approach. Methods Ecol Evol 2:349–361
Kremen C, Williams NM, Aizen MA, Gemmill-Herren B, LeBuhn G, Minckley R, Packer L, Potts SG, Roulston T, Steffan-Dewenter I, Vázquez DP, Winfree R, Adams L, Crone EE, Greenleaf SS, Keitt TH, Klein AM, Regetz J, Ricketts TH (2007) Pollination and other ecosystem services produced by mobile organisms: a conceptual framework for the effects of land-use change. Ecol Lett 10(4):299–314
Lander TA, Bebber DP, Choy CTL, Harris SA, Boshier DH (2011) The circe principle explains how resource-rich land can waylay pollinators in fragmented landscapes. Curr Biol 21(15):1302–1307
Lavergne S, Thuiller W, Molina J, Debussche M (2005) Environmental and human factors influencing rare plant local occurrence, extinction and persistence: a 115-year study in the Mediterranean region. J Biogeogr 32:799–811
MacArthur RH, Pianka ER (1966) On the optimal use of a patchy environment. Am Nat 100:603–609
MacArthur RH, Wilson EO (1967) The theory of Island biogeography. Princeton University Press, Princeton
McRae BH, Dickson BG, Keitt TH, Shah VB (2008) Using circuit theory to model connectivity in ecology, evolution and conservation. Ecology 89(10):2712–2724
Meagher T (1986) Analysis of paternity within natural population of Chamaelirium luteum 1. Identification of most-likely male parent. Am Nat 128:199–215
Moilanen A, Hanski I (1998) Metapopulation dynamics: effects of habitat quality and landscape structure. Ecology 79:2503–2515
Ohashi K, Thomson J (2005) Efficient harvesting of renewing resources. Behav Ecol 16:592–605
Potts SG, Biesmeijer JC, Kremen C, Neumann P, Schweiger O, Kunin WE (2010) Global pollinator declines: trends, impacts and drivers. Trends Ecol Evol 25:345–353
Pullinger MG, Johnson CJ (2010) Maintaining or restoring connectivity of modified landscapes: evaluating the least-cost path model with multiple sources of ecological information. Landsc Ecol 25(10):1547–1560
Rands S, Whitney H (2011) Field margins, foraging distances and their impacts on nesting pollinator success. PLoS One 6(10):e25971
Ray N (2005) PATHMATRIX: a geographical information system tool to compute effective distances among samples. Mol Ecol Notes 5(1):177–180
Richard Y, Armstrong DP (2010) Cost distance modelling of landscape connectivity and gap-crossing ability using radio-tracking data. J Appl Ecol 47(3):603–610
Richards S, Williams N, Harder L (2009) Variation in pollination: causes and consequences for plant reproduction. Am Nat 174:382–398
Ricketts TH (2001) The matrix matters: effective isolation in fragmented landscapes. Am Nat 158(1):87–99
Ries L, Fletcher RJ, Battin J, Sisk TD (2004) Ecological responses to habitat edges: mechanisms, models, and variability explained. Annu Rev Ecol Evol Syst 35:491–522
Romero G, Antiqueira P, Koricheva J (2011) A meta-analysis of predation risk effects on pollinator behaviour. PLoS One 6(6):e20689
Russell K (2003) EUFORGEN technical guidelines for genetic conservation and use for wild cherry (Prunus avium). International Plant Genetic Resources Institute, Rome, p 6
Sonneveld T, Tobutt KR, Robbins TP (2003) Allele-specific PCR detection of sweet cherry self-incompatibility (S) alleles S1 to S16 using consensus and allele-specific primers. Theor Appl Genet 107(6):1059–1070
Spear SF, Balkenhol N, Fortin MJ, McRae BH, Scribner K (2010) Use of resistance surfaces for landscape genetic studies: considerations for parameterization and analysis. Mol Ecol 19(17):3576–3591
Stoeckel S, Grange J, Fernandez-Manjarres JF, Bilger I, Frascaria-Lacoste N, Mariette S (2006) Heterozygote excess in a self-incompatible and partially clonal forest tree species—Prunus avium L. Mol Ecol 15(8):2109–2118
Stoeckel S, Klein E, Oddou-Muratorio S, Musch B, Mariette S (2012) Microevolution of S-allele frequencies in wild cherry populations: respective impacts of negative frequency dependent selection and genetic drift. Evolution 66:486–504
Testolin R, Marrazzo T, Cipriani G, Quarta R, Verde I, Dettori MT, Pancaldi M, Sansavini S (2000) Microsatellite DNA in peach (Prunus persica L. Batsch) and its use in fingerprinting and testing the genetic origin of cultivars. Genome 43:512–520
Tischendorf L, Fahrig L (2000) How should we measure landscape connectivity? Landsc Ecol 15(7):633–641
van Etten J (2012) Gdistance: distances and movements on geographical grids. http://r-forge.r-project.org/projects/gdistance
Vandermeer J, Carvajal R (2001) Metapopulation dynamics and the quality of the matrix. Am Nat 158(3):211–220
Vaughan SP, Boskovic RI, Gisbert-Climent A, Russell K, Tobutt KR (2008) Characterisation of novel S-alleles from cherry (Prunus avium L.). Tree Genet Genomes 4(3):531–541
Winker K, Rappole J, Ramos M (1995) The use of movement data as an assay of habitat quality. Oecologia 101:211–216
Wolf S, Moritz RFA (2008) Foraging distance in Bombus terrestris L. (Hymenoptera: Apidae). Apidologie 39:419–427
Zeller KA, McGarigal K, Whiteley AR (2012) Estimating landscape resistance to movement: a review. Landsc Ecol 27(6):777–797
Acknowledgments
We thank Laurent Lévèque and Bénédicte Le Guérroué for field sampling and lab genotyping for the Neuillé population. Isabelle Bilger, Stéphane Matz and the French Office National des Forêts sampled at St-Gobain, and Jérôme Grange and Sandrine Toussaint helped to acquire genetic data for this population. This work was funded by grants to T. L. by INRA (EFPA department) and to S.M. and S.S. from Cemagref, and the Office National des Forêts.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Lander, T.A., Klein, E.K., Stoeckel, S. et al. Interpreting realized pollen flow in terms of pollinator travel paths and land-use resistance in heterogeneous landscapes. Landscape Ecol 28, 1769–1783 (2013). https://doi.org/10.1007/s10980-013-9920-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10980-013-9920-y