
Abstract
Although pollen dispersal has been extensively studied in trees, parameters influencing between-population variation are still poorly understood. In this study, we conducted paternity analyses on open-pollinated seeds in four natural populations of wild cherry (Prunus avium) with contrasting density and clonal propagation, using eight microsatellite loci and one self-incompatibility system locus. We also measured four quantitative traits and spatial positions as potential correlates of reproductive success. Levels of polyandry differed among populations and 30% of the seed families exhibited unequal paternal contributions, suggesting variation in reproductive success rather than variation in mate availability. Mating occurred preferentially among neighbours in all populations, suggesting that it is a common pattern in wild cherry and probably results from pollinator behaviour. Paternal success was positively correlated with diameter at breast height, as indicated in previous studies and tree dominance only resulted in higher paternal success in low density plots. Mating patterns were thus also affected by both density and tree size. Large-scale studies are needed to disentangle relative influences of these factors on the mating system and pollination success.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The conservation of genetic resources is an important issue among current concerns about climate change, biological invasions, habitat fragmentation and plant breeding. In particular, the reproductive success of adults strongly influences the genetic structure of the following generations and thus the evolution of species. Mate diversity optimises female fitness through dilution of potential negative effects of an inferior or incompatible pollen donor (Janzen 1977; Karron and Marshall 1993). This is of particular interest in species exhibiting high rates of asexual reproduction, where clonal groups may provide high pollen loads to neighbouring pollen recipients that might be incompatible (e.g. other ramet of the same clone) or of poor quality (e.g. poor germination ability).
High genetic quality and diversity in set seed is crucial to population maintenance. Style saturation in pollen recipients with incompatible pollen might result in lower offspring diversity and fitness (Burke et al. 2000; Stoeckel et al. 2006). Seedling and sapling stages suffer from intense competition in tree species (Gonzalez-Martinez et al. 2002; Petit and Hampe 2006), resulting in high selective pressure and mortality at the younger stages. Furthermore, long-lived individuals will encounter different environmental conditions during their lifespan, thus resulting in further selective processes on the population.
Variation in phenology, tree size, compatibility systems, population size, isolation and interspecies interactions within and among populations may influence reproductive patterns and may lead to genetic erosion through lower mate availability. Studying reproductive patterns in several geographically separated populations can provide insight into the effects of local environments on the mating system. However, only a few studies considered population variation in reproductive success (Dick et al. 2003; Byrne et al. 2007; Hanson et al. 2008), probably due to the extensive sampling and laboratory work necessary for paternity analyses (Smouse and Sork 2004). Studies of pollen dispersal and male mating success within single populations indicate that pollen dispersal distances are generally shorter in animal-pollinated than in wind-pollinated species (Smouse and Sork 2004), but long-distance pollination events occur. Low pollen dispersal distance enhances spatial genetic structure within populations and could lead to higher rates of biparental inbreeding (Degen et al. 2004), at least in species in which no incompatibility system occurs.
Wild cherry (Prunus avium L.) is a rare and economically important species, used as a timber species naturally occurring throughout Europe, Western Asia and Northern Africa (Ducci and Santi 1997). It has also been used in reforestation in temperate forests for several decades (Russell 2003). P. avium is a rare and scattered species in mixed deciduous forests, lives up to 100 years and is characterised by a short juvenile period, with flowering occurring at d.b.h of 4 cm. Genetic diversity in wild cherries has been extensively studied because of its role as potential gene pool for improving cherry cultivars. Flowers are hermaphrodite but self-fertilisation is prevented by gametophytic self-incompatibility (GSI). Obligate outcrossing and complete incompatibility result when two individuals carry exactly the same two-allele genotype at the S-locus. Variability at the S-locus has been extensively studied (Granger 2004; De Cuyper et al. 2005; Schueler et al. 2006; Marchese et al. 2007; Vaughan et al. 2008). Vegetative reproduction via root suckering is very common, 9–65% of the trees in the stand (see Vaughan et al. 2007a), resulting in the occurrence of clonal clusters and aggregated distribution of the cherry trees within stands (Ducci and Santi 1997; Gomory 2004; Schueler et al. 2006). Pollen is transferred between trees by insects (honeybees and bumblebees), and seeds are dispersed by gravity and birds. In previous studies, pollen dispersal distances were short and ranged from 20 to 145 m (Granger 2004; Schueler 2005; Stoeckel 2006, Cottrell et al. 2009), while seed dispersal patterns and distances have not been addressed yet. Since these studies included only single populations and sometimes few families, the factors influencing pollen dispersal distances in P. avium are still unknown. High density of conspecifics might result in shorter pollen dispersal (Bacles and Jump 2011; Robledo-Amuncio et al. 2004; Smouse and Sork 2004), while the presence of incompatible related trees or ramets of the same clone might result in longer pollination distances. Significant spatial genetic structure is often reported in wild cherry populations and could reduce local mate availability through the occurrence of incompatible individuals (Schueler et al. 2006, Vaughan et al. 2007b). Conspecific density and clonal propagation might present opposing influences on pollen dispersal patterns at high density.
In this study, we aimed at better understanding of variation in mating system in wild cherry in relation to spatial arrangement, quantitative variation among individual trees and amount of clonal ramets. Previous studies reported the effective number of fathers sampled by pollen recipients (Schueler 2005; Stoeckel 2006); however, they did not address whether unequal paternal contributions occurred. These might indicate differential reproductive success among fathers. According to Cottrell et al. (2009) and Stoeckel (2006), diameter at breast height (d.b.h.) corresponded with better reproductive success, and clonal groups exhibited stronger male success and reduced mate availability in neighbour compatible pollen recipients. However, Cottrell et al. (2009) assessed only kinship among progenies, thus not clearly taking into account the presence of spatial genetic structure. Hence, it is unclear whether these variations in reproductive success and mate availability are common among wild cherry populations, and if they vary with density and amount of clonal propagation.
In this paper, we study sexual mating patterns in four geographically separated P. avium populations varying in conspecific density and extend of clonality. We address in particular the following questions: (1) Is there variation in spatial mating patterns and pollination distances? (2) Does maternal and paternal reproductive success vary within and among populations? (3) What are the parameters underlying the observed variation in sexual mating system?
Materials and methods
Populations and sampling
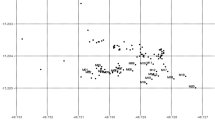
Four P. avium populations from different provenance regions (REF: German act of forest reproductive material) in northern Germany were included in this study (Fig. 1). The stands consist of mixed deciduous forests, dominated by Fagus sylvatica, Quercus robur and Carpinus betulus. Stands Chorin and Einhaus represent relatively isolated forest fragments, whereas Hildesheim and Spargründe are sites within a larger forest area. At each site, one leaf from each reproductive trees (Chorin and Einhaus) or from a continuous plot of at least 150 wild cherry trees (Hildesheim and Spargründe) was sampled. Adult trees were randomly distributed in stands Hildesheim and Spargründe, while moderate aggregation was found in stands Einhaus and Chorin (Jolivet et al. 2011). In each stand, we also sampled one leaf from 150 to 180 saplings (diameter at breast height < 4 cm). Since densities of wild cherry trees differed among stands (number of adult wild cherries in the stand divided by the stand area; Table 1), size of the plots varied (Fig. 1), resulting in a sub-optimal sampling design. Establishing a good sampling design is challenging when disturbance and management strategies cannot be controlled (Bacles and Jump 2011). However, given that density is around 40-fold higher in Spargründe than in Chorin, increasing plot size in high density stands would have resulted in an extremely high sampling effort. By contrast, reducing plot size in low density populations would have resulted in low sample size and imprecise estimation of allele frequencies.

Distribution of the four sampled wild cherry (Prunus avium) populations in the Northern part of Germany, and spatial positions in each stand of single genotype trees (empty circles), clonal individuals (full circles) and mother trees (squares, bold labels). Crosses and italic labels indicate the spatial position of the virtual clonal individual used for the analysis on the genet dataset. In highly clonal stands, we draw the limits of each clonal group
The spatial position, d.b.h., height and sociological class (Kraft 1884) of all sampled adult trees were measured. The latter was a scale from one to five representing tree dominance, from dominant tree (class one) to completely overtopped tree (class five). In further statistical analysis, we used letters in order to emphasise the categorical nature of this variable (from class A for dominant tree to E for overtopped tree). Trees with a d.b.h. of 4 cm or more were considered to be reproductive. In 2006, about 50 seeds per tree were sampled from at least 20 adults in each stand.
Genotyping
DNA from adult trees, saplings and seeds was isolated according to Dumolin et al. (1995). The DNA was quantified by spectrometry (Eppendorf Biophotometer, Hamburg, Germany) and diluted to a standardised DNA concentration of 10 ng/μL. We amplified eight microsatellite loci previously described in the literature: UDP96_001, UDP98_021, UDP98_410, UDP98_411, UDP98_412, UDP96_005 (Schueler et al. 2003; Testolin et al. 2000), BPPCT_034 and BPPCT_040 (Dirlewanger et al. 2002). For the first quadruplex (UDP98_411 (HEX), UDP98_412 (HEX), UDP96_005 (TET), BPPCT_040 (FAM)), we performed a single PCR (Polymerase Chain Reaction) using 2.5 μL 10× PCR buffer, 3 μL MgCl2 (DNA-Cloning-Service, Hamburg, Germany), 1 μL dNTP mix (Carl Roth and Co KG, Karlsruhe, Germany), 0.2 μL Taq polymerase (DNA-Cloning-Service, Hamburg), 0.2 μL (20 μM) of each primer reverse and forward (Eurofins MWG Operon, Ebersberg, Germany), except for primers UDP98_412-F (HEX) and UDP98_412-R for which we used 1.2, and 5 μL DNA template, for a total PCR volume of 25 μL. PCR amplification was conducted on a Biometra thermocycler and consisted of an initial denaturing of 4 min at 94°C, then 30 cycles of 25 s at 94°C, 25 s at 60°C, 45 s t 72°C and a final extension of 45 min at 60°C. For the second quadruplex (UDP96_001 (FAM), UDP98_021 (HEX), UDP98_410 (TET), BPPCT_034 (HEX)), we used 0.5 μL of each UDP96_001-F (FAM) and UDP96_001-R, 0.3 μL of each UDP98_021-F (HEX) and UDP98_021-R, 0.8 μL of each UDP98_410-F (TET) and UDP98_410-R and 0.2 μL of each BPPCT_034-F (HEX) and BPPCT_034-R, the other PCR reagent quantities being the same as in the first quadruplex. PCR was the same as for the first quadruplex, but consisted of 25 cycles instead of 30, and annealing temperature was 56°C instead of 60°C. Fragments were analysed on a Megabace genetic analyser (Amersham Biosciences, Buckinghamshire, England) using the Megabace ET 550-R size standard (GE Healthcare, Munich, Germany). Allele binning and scoring was performed with the software MegaBace Fragment Profiler v1.2 (Amersham Biosciences, Buckinghamshire, England). Microsatellite loci were controlled for presence of null alleles, large allele dropout and stuttering errors with MicroChecker v 2.2.3 (Van Oosterhout et al. 2004).
GSI genotypes were characterised for all adult individuals by amplification and gel visualisation of the first and second introns of the S-RNase gene, which exhibit length polymorphism (Sonneveld et al. 2003). For the first consensus primer (PaConsI), we used 60 ng DNA, 2.5 μL 10× Buffer, 2.5 mM MgCl2 (Invitrogen, Darmstadt, Germany), 0.2 mM dNTP (Carl Roth and Co KG, Karlsruhe, Germany), 0.2 mM of each forward and reverse primer (Eurofins MWG Operon, Ebersberg, Germany), 0.625 U Taq polymerase (Invitrogen, Darmstadt, Germany) for a total reaction volume of 25 μL. PCR was conducted with the following conditions: 2 min denaturing at 94°C, followed by 35 cycles of 1 min at 94°C, 1 min at 54°C and 1 min at 72°C, and a final extension of 5 min at 72°C. For the second consensus primer (PaConsII), we used 60 ng DNA, 2.5 μL 10× Buffer, 2 mM MgCl2, 0.2 mM dNTP, 0.2 mM of each forward and reverse primer, 1.25 μL W-1 buffer (Invitrogen, Darmstadt, Germany) and 1.25 U Taq polymerase for a total reaction volume of 25 μL. PCR was conducted with the following conditions: 2 min denaturing at 94°C, followed by 10 cycles of 10 s at 94°C, 2 min at 58°C and 2 min at 68°C, then 25 cycles of 10 s at 94°C, 2 min at 58°C and 2 min at 68°C, with an increment of the last step of 10 s at each cycle. Alleles were identified according to Vaughan et al. (2008).
Data analysis
Clones were first identified based on identity analysis of the genotypes at the eight microsatellite loci using CERVUS v. 3.0.3 (Marshall et al. 1998; Kalinowski et al. 2007) allowing one mismatch between pair of tested individuals. Although the microsatellite loci tested did not exhibit presence of null alleles, allele dropout or stuttering errors, mismatches were allowed. Some loci, especially UDP98_412 and BPPCT_040, exhibited crossfluorescence, resulting in higher rates of genotyping error (not estimated). Furthermore, somatic mutations may accumulate during asexual reproduction (Vaughan et al. 2007a). Clones were confirmed with the GSI genotypes. For analyses of male contribution, we removed all trees belonging to a clonal group and replaced them by a single virtual individual, to which was attributed the most frequent genotype for each locus, average spatial position and quantitative measurements corresponding to the ramet exhibiting the highest d.b.h within the clonal group.
Paternity
We first addressed the number of candidate fathers indirectly with a mating system analysis (MLTR, Ritland 2002). Multilocus correlated paternities (rp(m)) were 0.108, 0.142, 0.098 and 0.089 for Einhaus, Chorin, Hildesheim and Spargründe, respectively, yielding estimates of effective number of fathers (1/rp(m)) of 9.3, 7.04, 8.5 and 10.2. Therefore, there should be more than ten candidate fathers considered for further analysis.
We then conducted paternity analyses based on the microsatellite data with the software CERVUS v. 3.0.3. Allele frequencies were calculated only for adults. Simulations were run for 50 000 offspring, considering 20 candidate fathers. We took into account the fraction of adult trees sampled in each stand (70% in Einhaus, 50% in Hildesheim, 80% in Chorin and 10% in Spargründe), thus considering the occurrence of not sampled cherry trees close to the sampled plots. Selfing was not allowed. Fathers were assigned based on 95% confidence levels of LOD scores. Considering all individuals above the critical LOD determined by simulations, 95% of the fathers are correctly assigned. Because our stands show significant levels of spatial genetic structure, we did not use the Δ statistic. Δ represents the difference between the LOD scores of the two most likely fathers. Therefore, high relatedness between candidate fathers would result in low Δ and thus to ambiguous assignment. Assignment based on Δ would thus have underestimated local pollinations events because spatial genetic structure occurs, although preliminary analyses did not show huge differences.
We checked each assigned father for compatibility with the mother at the S-locus and excluded all offspring stemming from incompatible fathers from further analysis. However, this method does not take into account that only 50% of the pollen from the candidate fathers which exhibit one allele in common with the mother at the S-locus is compatible. Stoeckel (2006) used a model to take this into account and found similar results applying this method and CERVUS. This is probably due to the high exclusion probability obtained with eight microsatellite loci (combined exclusion probability for the parent pair > 0.999 in all four populations included in our study). In order to estimate mate availability, we report an average percentage of compatible mates based on S-genotypes, where mates sharing one S-allele counted as half.
Effective pollen dispersal was estimated as the distance between the mother tree and the assigned father, therefore representing pollen-mediated gene dispersal, as this estimate is post-fertilisation. We then estimated cryptic pollen flow, that is, the probability that a father is assigned, whereas the true father grows outside the stand. According to Dow and Ashley (1998), cryptic pollen flow is calculated as P m = 1 − (1 − x)n, where x is the non-exclusion probability of the second parent and n the number of individuals in the population participating in mating through their male function.
To address whether males showed equal paternity contributions, we calculated the unbiased estimator of the effective number of fathers in each seed family (K e) as described in Nielsen et al. (2003), eq. 16. In addition, the inverse of the correlated paternity measure within seed families (K p) was calculated with the software Kindist v. 1.0 (Robledo-Arnuncio et al. 2007). The latter represents an indirect estimator of the effective number of fathers based on the calculation of kinship coefficients among offspring sharing the same mother. We choose to present both methods because categorical paternity assignment methods (CERVUS) may result in overrepresentation of distant males, therefore overestimating the effective number of fathers. By contrast, indirect methods based on kinship coefficients assume no spatial genetic structure, and in our case, we expect underestimation of the effective number of fathers. χ2 tests were conducted in each family to test for the significance of the paternity contributions observed against the null hypothesis of equal frequency among fathers.
Quantitative traits
Prior to further analysis including quantitative traits, we used a Kruskal–Wallis test to see whether the size of the trees was different across populations. Fractional paternity analysis models taking quantitative traits into account have not yet been published on species carrying a gametophytic incompatibility system. Therefore, we tested potential correlates of paternal and maternal reproductive success (number of seeds sired, number of females sired and average number of seeds sired per female for paternal success; number of males sampled for maternal success) with generalised linear models using Poisson distributions corrected for over-dispersion. In Hildesheim and Spargründe, where many clones were identified, we tested whether belonging to a clonal group had a significant effect on male mating success by adding this as a covariate to the model. After correcting for the effect of distance among mates, each potential correlate (height, ground-crown distance, tree dominance and d.b.h.) was tested separately as a class variable to avoid miscalculation problems due to colinearity between these variables (correlation tested using an ANCOVA analysis taking into account differences between populations, all correlations highly significant) and to missing data. In fact, methods to correct for multicollinearity such as, e.g., ridge regression are not adapted to non-normally distributed data; therefore, we could not fit all covariables to a single model. We simplified the model when no significant interaction or no significant effect of clonal propagation was detected. For female reproductive success, we added the number of assigned seeds as a covariate to take into account differences in sample size.
Due to spatial arrangement among trees, we expected a significant effect of the distance between mates. Therefore, we compared the observed effective pollination events with pollination events expected under random mating. For each seed tree, we calculated the distance to all compatible pollen donors (other seed trees are also considered as possible pollen donors), including all clonal ramets. We conducted a paired t-test between frequencies of observed versus expected pollination distance between 0 and 100 m for Einhaus and Chorin, and between 0 and 50 m for Hildesheim and Spargründe. Each seed tree was considered as one replicate.
All statistical analyses were performed with the software R v 2.0.6 (R Development Core Team 2004).
Results
Molecular diversity and clonal reproduction
The number of clones varied across populations. Since the probability that two individuals drawn randomly had the same multilocus genotypes was low for seven of the eight loci (combined probability of identity of 10−6), we assumed that individuals exhibiting the same genotype at these seven loci were ramets of the same clone. We used the GSI genotype to exclude “false positives”, which represented less than one per cent of the clonal ramets. Population Chorin showed the lowest clonal proportion (proportion of individuals belonging to a clonal group within the stand) at the adult stage (6%) and Spargründe the highest (65%), while populations Einhaus and Hildesheim exhibited intermediate values (14 and 29%) (Fig. 1). Vegetative propagation was even higher in saplings, where clonal proportion was the lowest for Chorin (24%) and the highest for Spargründe (81%). Of the saplings in Hildesheim and Einhaus, 27 and 56%, respectively, were derived from asexual reproduction.
The GSI locus exhibited the highest diversity among adult trees (22 alleles), while across populations, we observed 4–14 alleles at microsatellite loci (Table 2). Variation in effective number of alleles resulted in differences in paternity non-exclusion probabilities (calculated with microsatellite data only) between loci and populations (Table 2), higher polymorphism resulting generally in lower non-exclusion probabilities.
Paternity assignment and paternal contributions
We first addressed whether it was possible to assign one father to each collected seed. Paternity assignment in CERVUS was 32–64% successful (Table 3), indicating that a high proportion of seeds could not be unambiguously assigned, although non-exclusion probabilities were small (Table 2). Of the assigned seeds, less than 1% was excluded from the analysis due to incompatibility between the parents. 53–93% of the sampled adult trees contributed pollen to the sampled seeds, and seed trees sampled up to 25 fathers (Table 4). We can thus conclude that reproduction is not restricted to a small number of adult trees and that high pollen flow occurs within P. avium populations. This was also reflected by the high compatibility among potential mating pairs (Table 3), confirming previous analyses of the same data, which showed very low biparental inbreeding (unpublished results).
Subsequently, we addressed whether the paternal contributions of males were equal. To this end, we report for each family Nielsen’s unbiased estimator (for low sample size) of the effective number of fathers (K e) and the effective number of fathers estimated from correlated paternity within pollen pools (K p) (Table 4). The number of fathers per half-sib family K was significantly correlated in all populations (linear model, all P values < 0.002) with the number of seeds genotyped. K e varied significantly among populations (ANOVA F 3,79 = 5.11, P = 0.003) but not K p (ANOVA F 3,79 = 1.08, P = 0.36). We observed significant unequal paternal contributions in 25% (Einhaus), 23% (Hildesheim), 36% (Chorin) and 31% (Spargründe) of the studied seed families (Table 4), the five most successful fathers siring, respectively, 19, 18, 23 and 47% of the assigned seeds. This suggests variation in male reproductive success.
Pollen-mediated gene flow
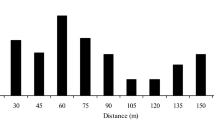
We estimated effective pollination distances from the results of the paternity analysis. Average pollen dispersion ranged from 42 to 134 m (Table 3). Interestingly, frequency of short-distance effective pollination events was higher than the expected frequency under random mating (Fig. 2), but dispersal also occurred at long distances as indicated by the maximum pollination distances observed. Furthermore, these observations are consistent with the high polyandry (K) observed. We observed a neighbour effect in all populations, as indicated by the higher frequency of observed effective pollination events in a radius of 100 m (Einhaus, Chorin) or 50 m (Hildesheim, Spargründe) around seed trees (paired t-test, all P values < 0.0001) compared with expected pollination events (Fig. 2). In the lower density populations (Einhaus and Chorin), median pollen dispersal accounted for 62 and 63% of the average pollen dispersal distances, while in the higher density stands (Hildesheim and Spargründe), it accounted for 80 and 68% of the average pollination distances. Thus, at lower densities, although pollen dispersal distances are longer, short dispersal distances seem to be relatively more frequent. However, this result could somehow be biased by the size differences among low and high density plots.

Expected and observed pollen dispersal distance in four Prunus avium populations. Expected distances (open bars) were estimated from the distance between seed trees and all compatible adult trees in the population. Observed pollen dispersal distances (full bars) were estimated directly from paternity analysis
Estimates of cryptic gene flow were very high (Table 3), but might be overestimated because our calculations do not take into account the size of the outside population and the isolation by distance to other forest fragments (Oddou-Muratorio et al. 2003).
Correlates of reproductive success
In order to understand the factors influencing variation in paternity contributions in the studied P. avium populations, we tested whether size (height, distance between the ground and the crown, and d.b.h), tree dominance and distance between mates correlate with male and female reproductive success. First, we observed very high differentiation between populations for all traits studied (Kruskal–Wallis test, all P values < 0.05). Furthermore, in Spargründe, adult trees seem to be younger, as indicated by lower height and d.b.h. (Table 1).
As expected, the number of seed sired and the number of seed trees pollinated were significantly and positively affected by the d.b.h. of pollen donors in all stands except Hildesheim (Fig. 3; Supplementary material Table S1). In populations Einhaus and Hildesheim, height also exhibited a significantly positive effect on male reproductive success. Dominant trees (sociological classes 1 and 2, class A and B in Table S1) generally had better siring success in the two low density and large plots (Einhaus and Chorin). Altogether, this suggests that larger trees are more efficient mates in the four populations.

Effect of quantitative traits on male reproductive success measured as the number of offspring sired per seed tree in four populations of Prunus avium. a Height of the pollen donor. b Height of the beginning of the crown. c Diameter at breast height (d.b.h.) d Tree dominance (kraft), 1 indicating dominant tree
Distance between mates exhibited significantly negative effects on the number of seed sired by each male across all seed trees (Table S1). When considering the number of seeds sired per seed tree, we found highly significant effects for all populations except Spargründe. Together with a higher frequency of short-distance pollinations compared with random expectations (see above), this suggests that compatible neighbouring trees are more successful, and this may have led to unequal paternal contributions. Indeed, pollination distance was smaller in families exhibiting unequal paternal contributions in three populations (Einhaus, Hildesheim, Spargründe), although this effect was only marginally significant (two-factors ANOVA, Population: F 3,75 = 48.6, P < 0.0001, Unequal Paternity (Yes/No): F 1,75 = 3.96, P = 0.05, Interaction: F 3,75 = 3.92, P = 0.01; Fig. 4a). Likewise, density of compatible individuals within 100 m for high density populations and within 200 m for low density populations tended to deflate in seed families where unequal paternity contributions were observed, although it strongly increased in Hildesheim (two-factors ANOVA, Population: F 3,75 = 179.3, P < 0.0001, Unequal Paternity (Yes/No): F 1,75 = 0.15, P = 0.7, Interaction: F 3,75 = 8.2, P < 0.001; Fig. 4b). Therefore, pollination events resulting in unequal paternity contributions stemmed mostly from neighbouring trees, while the effects of mate availability through variation in density of compatible individuals are still unclear.

Average pollination distance (a) and density of compatible individuals (b) in seed families showing equal and unequal paternity contributions in four Prunus avium stands
In populations Hildesheim and Spargründe, we additionally tested whether clonal ramets showed higher reproductive success. We observed significantly higher numbers of seeds sired and pollinated trees for clonal ramets (Supplementary material Table S1). These results suggest that clonal genotypes participate more in total population reproduction at least via their male function.
As high genetic variation between open-pollinated offspring of a single seed tree is often related to higher maternal reproductive success, we tested the effects of the size of the mother on the number of males sampled. Our results do not indicate significant effects of morphological variation among seed trees, neither does the distance to the mates influence the diversity sampled by seed trees.
Discussion
Clonal propagation
In this paper, we wanted to address whether natural P. avium populations varied in their realised mating systems. As observed in other studies (Ducci and Santi 1997; Gomory 2004; Stoeckel et al. 2006; Vaughan et al. 2007a), vegetative reproduction varied substantially across our four studied populations and ranged from 6 to 65% at the adult and from 24 to 81% in the sapling stage. The decrease in clones at the adult stage could probably result from higher mortality of the juveniles due to enhanced competition within patches of ramets. Spargründe exhibited extremely high clonal propagation. The history of the stand is unknown, but natural disturbance (storm damage) or management might have resulted in higher asexual recruitment. Vegetative propagation is more frequent in forests under historical management, i.e., coppicing by cutting down the stand at short intervals to promote light-demanding tree species able to reproduce asexually (Höltken 2005; Höltken and Gregorius 2006). In contrast, present-day management strategies (thinning of dense regeneration, clearing of understorey) have a negative effect on both sexual and asexual recruitment (Vaughan et al. 2007a). Population Chorin showed very low clonal reproduction at the adult stage, probably the result of presumed ancient planting of the stand. At this site, the higher percentages of cloned saplings might be due to the lack of management over the last decade. These results show that recruitment dynamics of sexual and asexual individuals across time still remains to be addressed, and here in particular, long-term competition effects between asexual and sexual individuals.
Pollen-mediated gene flow estimates
Mating occurred preferentially among neighbours, with most effective pollination occurring within 100 m in low density stands and within 50 m in high density stands. However, we also observed long-distance pollen flow. Our estimation of effective pollen dispersal distances was within previous estimates at similar densities for this species (Granger 2004; Schueler et al. 2006; Stoeckel 2006). However, our work is the first to consider the influence of conspecific density on effective pollen dispersal distance, as indicated by the correlation between density and pollen dispersal distance. Since the size of our high density study plots was smaller to ensure a similar sample size among populations, effective dispersal distances could be biased towards smaller sizes because long dispersal events were not detectable. However, we observed proximity effects in all stands, so things would probably not have changed dramatically if we would have sampled a much larger area in the high density stands. Based on the same data, simulations of randomly generated plots of constant size but varying density also showed shorter pollen dispersal at larger densities (Jolivet and Degen 2011). Furthermore, spatial genetic structure in these stands was stronger at high densities and resulted in shorter estimates of historical gene dispersal (Jolivet et al. 2011). Indeed, high conspecific density in continuous forests often induces reduced dispersal compared with low density fragmented forests (Bacles and Jump 2011). Higher density of flowering trees probably results in lower foraging distances of the pollinators as travelling costs are lower (Oddou-Muratorio et al. 2006) and also because pollinators tend to choose closer food sources (Pyke 1984). However, within patches, pollinator movements (Pyke 1984; Oddou-Muratorio et al. 2006) might also explain pollination among neighbours in the low density populations Einhaus and Chorin, where reproductive trees were moderately aggregated. Other studies also provided evidence for longer dispersal and sometimes higher effective numbers of fathers when density decreases (Robledo-Amuncio et al. 2004; Smouse and Sork 2004) or when the canopy opens (Smouse and Sork 2004). In fact, pollinators commonly visit flower patches distant from their nest (Chapman et al. 2003) which results in long-distance pollination events. Still, combined effects of neighbourhood density and canopy structure on pollinator behaviour are poorly understood (Oddou-Muratorio et al. 2005), as well as the relative importance of short- and long-distance pollination events.
Surprisingly, a previous study of the Einhaus stand (Schueler 2005) estimated average of pollen dispersal distance to be 84 m, whereas we observed 134 m. Although we did not use the same mother trees, this could be variation in pollen flow among years (Oddou-Muratorio et al. 2006, Smouse and Sork 2004). As quantitative traits in adults were similar between consecutive years, we could thus conclude that weather conditions or food source availability for pollinators during the flowering season could have influenced pollen dispersal. In fact, tree and pollinator phenology, pollinator activity, flowering synchrony and flowering intensity depend on temperatures. Food availability was shown to influence foraging patterns in honeybees, as a lower amount of flowers results in longer pollinator flights (Steffan-Dewenter and Kuhn 2003). Furthermore, each pollinator reacts differently to variation in temperatures. For example, honeybee activity can be reduced when weather conditions are too cold (Calzoni and Speranza 1998), and a shift to other pollinators can occur in cold years.
Polyandry and effective numbers of fathers
We observed high numbers of pollen donors in each seed family (eight to 18 on average per mother per population) and even higher effective numbers of fathers (seven to 25 on average per mother per population). The effective numbers of fathers estimated from kinship coefficients were also extremely high. This is surprising given that most studies on tree species reported only up to 11 effective fathers (Smouse and Sork 2004). P. avium should have even fewer effective fathers because it is insect pollinated. Lower levels of polyandry have been previously reported for wild cherry (eight to 16 pollen donors, three to ten effective pollen donors in Schueler 2005; 11 pollen donors, seven effective pollen donors in Stoeckel 2006). However, these studies used different methods and smaller sample sizes for the effective number of fathers. The strong correlation between the number of fathers observed and the number of seeds assigned per seed family indicated that the effective number of fathers might be sample size dependent and that we may have underestimated the true number of pollen donors.
Flowering synchrony may also affect the outcome of reproduction. Indeed, flowering time was very short in the year studied (around 16 days); most adult trees surveyed exhibited overlapping flowering (data not shown) and this could have additionally resulted in very high effective numbers of fathers (Oddou-Muratorio et al. 2006). In low density populations, more pollinator long-distance flights occur, at least when trees are not strongly isolated (Bacles and Jump 2011; Lacerda et al. 2008), thus increasing gene flow and the effective number of fathers. In the high density population Spargründe, the effective number of fathers was significantly much smaller (paired t-test, P < 0.05) when considering the unbiased estimator. Several factors could explain the results in this population, such as higher saturation of the flowers (Oddou-Muratorio et al. 2006), as a result of a higher density of flowering individuals, or a higher clonal proportion leading to a local decrease in mate availability though incompatibility. In contrast, Hildesheim did not exhibit low effective numbers of fathers. However, the high proportion of clonal propagation in this population is mainly due to three big clonal groups, and most mother trees are located outside these groups (Fig. 1), whereas in Spargründe, clonal groups overlap and density is higher. Unfortunately, the stands exhibiting high clonal propagation also showed the highest density; therefore, the results of our study fail to separate the effects of clonal propagation and density on the effective numbers of fathers. Large-scale studies including contrasting levels of both clonal propagation and local density are thus needed to unravel these mechanisms.
Unequal paternity contributions and reproductive success
High levels of polyandry were observed and overall, 30% of the seed families exhibited unequal paternal contributions, indicating variation in reproductive success among males. Advantageous quantitative traits such as high d.b.h and to a lesser extent height of the tree increased siring success probabilities compared with smaller individuals, as also reported by other studies on P. avium (Cottrell et al. 2009; Stoeckel 2006) or on other species (d.b.h: Sorbus torminalis, Oddou-Muratorio et al. 2005; height: Pinus attenuata, Burczyk et al. 1996). Our results also indicate positive effects of tree dominance, but only at lower densities.
By contrast, no effect of quantitative traits on reproductive success was reported for a wind-pollinated species (Quercus macrocarpa, Dow and Ashley 1998). Bigger trees may indeed exhibit higher flowering intensity resulting in higher attractiveness to the pollinator or higher fertilisation probability for wind-pollinated species. A few studies support this hypothesis: male flower production had a strong effect on male reproductive success (Cryptomeria japonica, Moriguchi et al. 2007; Sorbus torminalis, Oddou-Muratorio et al. 2005) or on flower visitation (in the herbaceous plant Silene latifolia, Shykoff and Bucheli 1995). However, we do not know whether d.b.h and flowering intensity are strongly correlated in P. avium. It would thus be interesting to include this quantitative trait in further studies.
Furthermore, there is high evidence that neighbouring pollen donors exhibit higher reproductive success (Burczyk 1996; Dow and Ashley 1998; Kaufman et al. 1998; van Kleunen and Burczyk 2008). Although we observed marginally significant differences among families showing equal and unequal paternity contributions in pollination distances, this still does not explain why some families were strongly pollinated by a few pollen donors. One way to unravel these effects would be to analyse our data using a model taking into account the distance among mates, quantitative traits and the incompatibility system genotype.
A further explanation for unequal paternity contributions could be pollen competition (differential germinability of the pollen: Jolivet and Bernasconi 2007, time needed to reach the ovule, i.e., “first male advantage”: Snow et al. 2000, Jolivet and Bernasconi 2007), pollen tube-style interactions (Marshall and Folsom 1991; Hormaza and Herrero1996) or post-fertilisation events such as fruit abortion (Dow and Ashley 1998). Post-pollination and post-fertilisation female selection for mates in plants are complex mechanisms which still need to be unravelled.
Unlike in pollen donors, we did not observe any effect of seed-tree quantitative traits on diversity (sampled number of fathers). This may be a consequence of other strategies in pollen recipients than size (d.b.h, height) to maximise the diversity sampled. Assuming only pollinator behaviour effects, we would have expected similar results in pollen donors and pollen recipients. One possibility would be via cryptic female mate choice (i.e. interactions between the style and pollen tubes, Marshall and Folsom 1991), as Hormaza and Herrero (1996) already suggested in P. avium. Higher diversity sampled through intense pollen competition is generally associated with higher offspring fitness, at least in plants with a mixed-mating system (Armbruster and Rogers 2004; Paschke et al. 2005). Since the selection of seedlings and saplings is extremely high in trees (Petit and Hampe 2006), in future, we should address paternal and maternal effects on offspring fitness to better understand the role of pollen diversity sampled by seed trees.
Our results provide a comparative understanding of mating processes in P. avium. Pollination events seem to be influenced by genetic and spatial processes, in particular population density, as well as by the presence of clonal individuals. We found that strong mating among neighbours occurs in all populations and that d.b.h. greatly influences reproductive success, as also suggested by former studies. Yet, it is still unclear why a few individuals showed high reproductive success through their male function, resulting in unequal paternity contributions in some seed families. More work is thus needed to understand the complex interactions between spatial processes, variation at quantitative traits and pollinator behaviour to disentangle confounding effects of spatial parameters such as density and clonal propagation, limiting the interpretation of the present results. Conservation strategies of genetic resources should consider variation in reproductive processes within and between populations, since polyandry levels vary. In future, sampling strategies should be adapted to the levels of within seed family diversity expected from population parameters, such as size, density of the trees, population structure and level of genetic diversity.
References
Armbruster WS, Rogers DG (2004) Does pollen competition reduce the cost of inbreeding? Am J Bot 91:1939–1943
Bacles CFE, Jump AS (2011) Taking a tree’s perspective on forest fragmentation genetics. Trends Plant Sci 16:13–18
Burczyk J (1996) Variance effective population size based on multilocus gamete frequencies in coniferous populations: an example of a Scots pine clonal seed orchard. Heredity 77:74–82
Burczyk J, Adams WT, Shimizu JY (1996) Mating patterns and pollen dispersal in a natural knobcone pine (Pinus attenuata Lemmon) stand. Heredity 77:251–260
Burke JM, Bulger MR, Wesselingh RA, Arnold ML (2000) Frequency and spatial patterning of clonal reproduction in Louisiana iris hybrid populations. Evolution 54:137–144
Byrne M, Elliott CP, Yates C, Coates DJ (2007) Extensive pollen dispersal in a bird-pollinated shrub, Calothamnus quadrifidus, in a fragmented landscape. Mol Ecol 16:1303–1314
Calzoni GL, Speranza A (1998) Insect controlled pollination in Japanese plum (Prunus salicina Lindl.). Sci Hortic 72:227–237
Chapman RE, Wang J, Bourke AFG (2003) Genetic analysis of spatial foraging patterns and resource sharing in bumble bee pollinators. Mol Ecol 12:2801–2808
Cottrell JE, Vaughan SP, Connolly T, Sing L, Moodley DJ, Russell K (2009) Contemporary pollen flow, characterization of the maternal ecological neighbourhood and mating patterns in wild cherry (Prunus avium L.). Heredity 103:118–128
De Cuyper B, Sonneveld T, Tobutt KR (2005) Determining self-incompatibility genotypes in Belgian wild cherries. Mol Ecol 14:945–955
Degen B, Bandou E, Caron H (2004) Limited pollen dispersal and biparental inbreeding in Symphonia globulifera in French Guiana. Heredity 93:585–591
Dick CW, Etchelecu G, Austerlitz F (2003) Pollen dispersal of tropical trees (Dinizia excelsa: Fabaceae) by native insects and African honeybees in pristine and fragmented Amazonian rainforest. Mol Ecol 12:753–764
Dirlewanger E, Cosson P, Tavaud M et al (2002) Development of microsatellite markers in peach (Prunus persica L. Batsch) and their use in genetic diversity analysis in peach and sweet cherry (Prunus avium L.). Theor Appl Genet 105:127–138
Dow BD, Ashley MV (1998) Factors influencing male mating success in bur oak, Quercus macrocarpa. New For 15:161–180
Ducci F, Santi F (1997) The distribution of clones in managed and unmanaged populations of wild cherry (Prunus avium). Can J For Res 27:1998–2004
Dumolin S, Demesure B, Petit RJ (1995) Inheritance of chloroplast and mitochondrial genomes in pedunculate oak investigated with an efficient PCR method. Theor Appl Genet 91:1253–1256
Gomory D (2004) Mutual links of demographic and genetic processes in a wild cherry population during the colonization. Biologia 59:493–500
Gonzalez-Martinez SC, Gerber S, Cervera MT et al (2002) Seed gene flow and fine-scale structure in a Mediterranean pine (Pinus pinaster Ait.) using nuclear microsatellite markers. Theor Appl Genet 104:1290–1297
Granger AR (2004) Gene flow in cherry orchards. Theorl Appl Genet 108:497–500
Hanson TR, Brunsfeld SJ, Finegan B, Waits LP (2008) Pollen dispersal and genetic structure of the tropical tree Dipteryx panamensis in a fragmented Costa Rican landscape. Mol Ecol 17:2060–2073
Höltken AM (2005). Genetische Untersuchungen zu den Vorassetzungen und Konsequenzen einer rezedenten Lebensweise am Beispiel der Vogelkirsche (Prunus avium L.). PhD Thesis, University of Göttingen
Höltken AM, Gregorius HR (2006) Detecting local establishment strategies of wild cherry (Prunus avium L.). BMC Ecol 4:13
Hormaza JI, Herrero M (1996) Dynamics of pollen tube growth under different competition regimes. Sex Plant Repro 9:153–160
Janzen DH (1977) Note on optimal mate selection by plants. Am Nat 111:365–371
Jolivet C, Bernasconi G (2007) Within/between population crosses reveal genetic basis for siring success in Silene latifolia (Caryophyllaceae). J Evol Biol 20:1361–1374
Jolivet C, Degen B (2011) Spatial genetic structure in wild cherry (Prunus avium L.): II. Effect of density and clonal propagation on spatial genetic structure based on simulation studies. Tree Genet Genomes 7:541–552
Jolivet C, Höltken AM, Liesebach H, Steiner W, Degen B (2011) Spatial genetic structure in wild cherry (Prunus avium L.): I. variation among natural populations of different density. Tree Genet Genomes 7:271–283
Kalinowski ST, Taper ML, Marshall TC (2007) Revising how the computer program CERVUS accommodates genotyping error increases success in paternity assignment. Mol Ecol 16:1099–1106
Karron JD, Marshall DL (1993) Effects of environmental variation on fitness of singly and multiply sired progenies of Raphanus sativus (Brassicaceae). Am J Bot 80:1407–1412
Kaufman SR, Smouse PE, Alvarez-Buylla ER (1998) Pollen-mediated gene flow and differential male reproductive success in a tropical pioneer tree, Cecropia obtusifolia (Bertol. Moraceae): a paternity analysis. Heredity 81:164–173
Kraft G (1884) Beiträge zur Lehre von den Durchforstungen, Schlagstellungen und Lichtungshieben. Klindworth, Hannover
Lacerda AEB, Kanashiro M, Sebbenn AM (2008) Long-pollen movement and deviation of random mating in a low-density continuous population of a tropical tree Hymenaea courbaril in the Brazilian Amazon. Biotropica 40:462–470
Marchese A, Tobutt KR, Raimondo A et al (2007) Morphological characteristics, microsatellite fingerprinting and determination of incompatibility genotypes of Sicilian sweet cherry cultivars. J Hortic Sci Biotechnol 82:41–48
Marshall DL, Folsom MW (1991) Mate choice in plants—an anatomical to population perspective. Annu Rev Ecol Syst 22:37–63
Marshall TC, Slate J, Kruuk LEB, Pemberton JM (1998) Statistical confidence for likelihood-based paternity inference in natural populations. Mol Ecol 7:639–655
Moriguchi Y, Tsuchiya S, Iwata H et al (2007) Factors influencing male reproductive success in a Cryptomeria japonica seed orchard revealed by microsatellite marker analysis. Silvae Genet 56:207–214
Nielsen R, Tarpy DR, Reeve HK (2003) Estimating effective paternity number in social insects and the effective number of alleles in a population. Mol Ecol 12:3157–3164
Oddou-Muratorio S, Houot ML, Demesure-Musch B, Austerlitz F (2003) Pollen flow in the wildservice tree, Sorbus torminalis (L.) Crantz. I. Evaluating the paternity analysis procedure in continuous populations. Mol Ecol 12:3427–3439
Oddou-Muratorio S, Klein EK, Austerlitz F (2005) Pollen flow in the wildservice tree, Sorbus torminalis (L.) Crantz. II. Pollen dispersal and heterogeneity in mating success inferred from parent-offspring analysis. Mol Ecol 14:4441–4452
Oddou-Muratorio S, Klein EK, Demesure-Musch B, Austerlitz F (2006) Real-time patterns of pollen flow in the wild-service tree, Sorbus torminalis (Rosaceae). III. Mating patterns and the ecological maternal neighbourhood. Am J Bot 93:1650–1659
Paschke M, Bernasconi G, Schmid B (2005) Effects of inbreeding and pollen donor provenance and diversity on offspring performance under environmental stress in the rare plant Cochlearia bavarica. Basic Appl Ecol 6:325–338
Petit RJ, Hampe A (2006) Some evolutionary consequences of being a tree. Ann Rev Ecol Evol Syst 37:187–214
Pyke GH (1984) Optimal foraging theory—a critical-review. Annl Rev Ecol Syst 15:523–575
R Development Core Team (2004) R: a language and environment for statistical computing, Vienna. ISBN 3-900051-07-0. http://www.R-project.org
Ritland K (2002) Extensions of models for the estimation of mating systems using n independent loci. Heredity 88:221–228
Robledo-Amuncio JJ, Smouse PE, Gil L, Alia R (2004) Pollen movement under alternative silvicultural practices in native populations of Scots pine (Pinus sylvestris L.) in central Spain. For Ecol Manag 197:245–255
Robledo-Arnuncio JJ, Austerlitz F, Smouse PE (2007) POLDISP: a software package for indirect estimation of contemporary pollen dispersal. Mol Ecol Notes 7:763–766
Russell K (2003) EUFORGEN Technical guidelines for genetic conservation and use for wild cherry (Prunus avium). International Plant Genetics Ressources Institute, Rome
Schueler S (2005) Pollen-mediated gene flow of trees in the temperate zone. PhD Thesis, University of Hamburg, Germany
Schueler S, Tusch A, Schuster M, Ziegenhagen B (2003) Characterization of microsatellites in wild and sweet cherry (Prunus avium L.)—markers for individual identification and reproductive processes. Genome 46:95–102
Schueler S, Tusch A, Scholz F (2006) Comparative analysis of the within-population genetic structure in wild cherry (Prunus avium L.) at the self-incompatibility locus and nuclear microsatellites. Mol Ecol 15:3231–3243
Shykoff JA, Bucheli E (1995) Pollinator visitation patterns, floral rewards and the probability of transmission of Microbotryum-Violaceum, a venereal-disease of plants. J Ecol 83:189–198
Smouse PE, Sork VL (2004) Measuring pollen flow in forest trees: an exposition of alternative approaches. For Ecol Manag 197:21–38
Snow AA, Spira TP, Liu H (2000) Effects of sequential pollination on the success of “fast” and “slow” pollen donors in Hibiscus moscheutos (Malvaceae). Am J Bot 87:1656–1659
Sonneveld T, Tobutt KR, Robbins TP (2003) Allele-specific PCR detection of sweet cherry self-incompatibility (S) alleles S1 to S16 unsing consensus and allele-specific primers. Theor Appl Gent 107:1059–1070
Steffan-Dewenter I, Kuhn A (2003) Honeybee foraging in differentially structured landscapes. Proc R Soc Lond B 270:569–575
Stoeckel S (2006) Impact de la propagation aséxuée et du système d’auto-incompatibilité gamétophytique sur la structuration et l’évolution de la diversité génétique d’une essence forestière entomophile et disséminée, Prunus avium L. PhD Thesis, Cemagref, France
Stoeckel S, Grange J, Fernandez-Manjarres JF et al (2006) Heterozygote excess in a self-incompatible and partially clonal forest tree species—Prunus avium L. Mol Ecol 15:2109–2118
Testolin R, Marrazzo T, Cipriani G et al (2000) Microsatellite DNA in peach (Prunus persica L. Batsch) and its use in fingerprinting and testing the genetic origin of cultivars. Genome 43:512–520
van Kleunen M, Burczyk J (2008) Selection on floral traits through male fertility in a natural plant population. Evol Ecol 22:39–54
Van Oosterhout C, Hutchinson WF, Wills DPM, Shipley P (2004) MICRO-CHECKER: software for identifying and correcting genotyping errors in microsatellite data. Mol Ecol Notes 4:535–538
Vaughan SP, Cottrell JE, Moodley DJ, Connolly T, Russell K (2007a) Clonal structure and recruitment in British wild cherry (Prunus avium L.). For Ecol Manag 242:419–430
Vaughan SP, Cottrell JE, Moodley DJ, Connolly T, Russell K (2007b) Distribution and fine-scale spatial-genetic structure in British wild cherry (Prunus avium L.). Heredity 98:274–283
Vaughan SP, Boskovic RI, Gisbert-Climent A, Russell K, Tobutt KR (2008) Characterisation of novel S-alleles from cherry (Prunus avium L.). Tree Genet Genomes 4:531–541
Acknowledgments
We thank Alexandra Meier, Volker Schneck and Thomas Stauber for technical assistance, Dina Führmann for editing, Alexandre Sebbenn, Sheila Ward, Jutta Buschbom and two anonymous reviewers for constructive comments on the manuscript. This work was supported by the German Ministry of Food, Agriculture and Consumer Protection (BMELV) [grant 05/BE003/2 “Erfassung der genetischen Struktur der Vogelkirsche Prunus avium als Grundlage für ein genetisches Monitoring wichtiger Waldbaumarten in Deutschland”].
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by R. Matyssek.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article
Cite this article
Jolivet, C., Höltken, A.M., Liesebach, H. et al. Mating patterns and pollen dispersal in four contrasting wild cherry populations (Prunus avium L.). Eur J Forest Res 131, 1055–1069 (2012). https://doi.org/10.1007/s10342-011-0576-3
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10342-011-0576-3