
Abstract
Habitat connectivity is an important element of functioning landscapes for mobile organisms. Maintenance or creation of movement corridors is one conservation strategy for reducing the negative effects of habitat fragmentation. Numerous spatial models exist to predict the location of movement corridors. Few studies, however, have investigated the effectiveness of these methods for predicting actual movement paths. We used an expert-based model and a resource selection function (RSF) to predict least-cost paths of woodland caribou. Using independent data for model evaluation, we found that the expert-based model was a poor predictor of long-distance animal movements; in comparison, the RSF model was effective at predicting habitat selection by caribou. We used the Path Deviation Index (PDI), cumulative path cost, and sinuosity to quantitatively compare the spatial differences between inferred caribou movement paths and predicted least-cost paths, and quasi-random null models of directional movement. Predicted movement paths were on average straighter than inferred movement paths for collared caribou. The PDI indicated that the least-cost paths were no better at predicting the inferred paths than either of two null models—straight line paths and randomly generated paths. We found statistically significant differences in cumulative cost scores for the main effects of model and path type; however, post-hoc comparisons were non-significant suggesting no difference among inferred, random, and predicted least cost paths. Paths generated from an expert based cost surface were more sinuous than those premised on the RSF model, but neither differed from the inferred path. Although our results are specific to one species, they highlight the importance of model evaluation when planning for habitat connectivity. We recommend that conservation planners adopt similar techniques when validating the effectiveness of movement corridors for other populations and species.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The loss of biodiversity is accelerating worldwide (Hoekstra et al. 2005). There are many local, regional, and continental causes of this ‘extinction crisis’, but the fragmentation and reduction in the connectivity of habitats is one of the major factors (Noss 1991). Consequences of fragmentation are many and include genetic deterioration and inbreeding due to restriction of gene flow; direct death of mobile and wide ranging individuals during transit between ‘suitable’ habitat patches; susceptibility to environmental and demographic perturbations and disasters; reduction in ‘interior’ habitat and subsequent loss of dependent species; and an increase in the number of nonnative species in remaining habitat patches (Noss 1991; Couvet 2002). The exact effects of fragmentation will vary across landscapes and species. We must assume, however, that maintaining natural levels of connectivity is preferable to fragmentation (Pace 1991).
Connected habitats may consist of a combination of patches that serve as movement routes or habitat corridors that also meet some of the resource requirements of the transient species (Merriam 1991; Kindall and Van Manen 2007). Harris (1984) points to the strategic location of smaller habitat patches or ‘stepping stones’ between larger habitat islands as one means of maintaining high levels of habitat connectivity. This approach is not as widely favoured as continuous corridors of habitat that physically connect larger habitat patches across the landscape (Noss 1991; Csuti 1991; Soulé et al. 2004). The ‘matrix’ of land surrounding habitat patches and corridors is now also recognized as important (Ricketts 2002; Soulé et al. 2004; Rouget et al. 2006; Beier et al. 2008). Management of the matrix to reduce movement and dispersal resistance will help improve habitat connectivity to a greater degree than the maintenance of corridors alone (Ricketts 2002).
A number of GIS methods and decision support frameworks are available for identifying corridors for animal and plant movements, but there is little consistency among applications and few studies have examined the effectiveness of these methods (Beier et al. 2008). As exceptions, Kehoe (1995) investigated the ability of collections of habitats constituting ‘linkage zones’—intact areas of land joining larger habitat patches in an otherwise fragmented landscape—to predict the movements of grizzly bears in Montana. Kehoe (1995) found that bears were no more likely to use predicted ‘linkage zones’ than any other area. Clevenger et al. (2002) tested the frequency of black bear crossings of a major road with crossing points predicted from a GIS model. They found that empirical and expert literature-based models both predicted linkage zones reasonably well, while an expert-based model was a poor predictor.
The least-cost model is another widely adopted method for predicting animal movement paths and corridors (Richard and Armstrong 2010). When applied to conservation, the technique is commonly used to identify larger zones that, if conserved, will promote animal movement within a hostile matrix (Goncalves 2010). This method uses habitat permeability, also known as ‘friction’ or ‘cost’ maps to derive the least-cost path or corridor between a pre-defined start and end point (Walker and Craighead 1997; Ray et al. 2002; Singleton et al. 2002; Kautz et al. 2006; Kindall and Van Manen 2007; Chetkiewicz and Boyce 2009). The habitat permeability map is a grid where each cell is given a value indicating the ‘cost’ for the animal to move across that cell (Walker and Craighead 1997; Singleton et al. 2002). Unlike the linkage zone model, we are unaware of any systematic studies designed to test the utility of this technique.
We propose a method, the Path Deviation Index (PDI; Jan et al. 1999), for assessing the least-cost path model’s ability to predict the spatial location of movement routes of animals. We apply the least-cost path model to the prediction of movement routes for woodland caribou (Rangifer tarandus caribou) and then use the PDI and a measure of path sinuosity to test for differences between inferred (i.e., reconstructed from measured locations of long distance movement), predicted, and random movement paths. We restrict our analysis and evaluation to defined paths not corridors. Corridors represent a more comprehensive and realistic conservation strategy, but they require additional information and a much more complex and potentially subjective decision/modelling framework (Beier et al. 2008).
Woodland caribou is an excellent species for testing the least-cost path model because these animals have predictable seasonal migrations. Caribou are of considerable conservation concern and, thus, the subject of planning efforts to maintain connectivity of critical habitats (Johnson et al. 2002a; Saher and Schmiegelow 2005; Apps and McLellan 2006). Further, caribou qualify as umbrella species as they have demanding habitat requirements during winter and large seasonal ranges throughout the year. Beier et al. (2008) recommend umbrella species as candidates for the focus of corridor design.
In this paper, we evaluate the utility of the least-cost path model for predicting the location and shape of movement paths typical of wide-ranging terrestrial mammals. Recognizing that paths may be dependent on the underlying permeability maps and that multiple sources of ecological information are available to conservation planners (Clevenger et al. 2002), we predict paths using maps generated with resource selection functions (RSF) and expert knowledge. Thus, as a component of that broader objective, we evaluate the ability of an RSF species distribution model and expert knowledge to identify habitats used by caribou during large-scale movements. We tested three hypotheses relative to the PDI, accumulated movement cost, and measures of sinuosity for inferred, predicted, random, and straight line paths: Hypothesis 1—the mean PDI for the predicted paths is less than the mean PDI for the random or straight line paths; Hypothesis 2—the cumulative cost for the predicted paths is less than the cumulative cost for the random or straight line paths; and Hypothesis 3—the mean sinuosity of the predicted paths does not differ from the mean sinuosity of the inferred paths.
Methods
Study area and animals
As part of a previous study (Johnson et al. 2002a), 16 female caribou of the Wolverine herd were captured, collared and monitored from March 1996 through March 1999. Caribou locations were collected using differentially correctable global positioning system (GPS) collars (GPS 1000, Lotek Engineering, Newmarket, ON, Canada) which recorded locations every third or fourth hour. As part of another study in the same area (S. McNay, Personal Communication), a further 11 caribou (eight female and three male) were captured, collared and monitored from January 1999 through March 2002. For these caribou, locations were collected every six or 12 h using differentially correctable GPS collars (GPS-Simplex g01-01010, Televilt, Telemetry Solutions, Concord, California, USA). During winter, female caribou travel in small groups, thus, the location data and inferred movements paths likely represent a larger proportion of the population than reflected in the collar sample size (Johnson et al. 2002a).
The area used by the Wolverine herd covers approximately 5,100 km2 and is located 250 km northwest of Prince George, British Columbia, Canada. The terrain varies from valley bottoms at 900 m to mountain tops of up to 2,050 m (Johnson et al. 2003). Below 1,100 m, dominant tree species are lodgepole pine (Pinus contorta), white spruce (Picea glauca), subalpine fir (Abies lasiocarpa) and hybrid white spruce (P. glauca × P. engelmannii). At mid elevations (1,100–1,600 m), the climate is moist and cool, with Engelmann spruce (P. engelmannii) and subalpine fir being the dominant tree species. Alpine tundra occurs above 1,600 m. Vegetation across these alpine areas consists of shrubs, herbs, bryophytes, and lichens with occasional trees in krummholz form. Approximately 7.2% of the study area was affected by forest harvesting over the last 20 years (Johnson et al. 2003).
As Chetkiewicz et al. (2006) discussed and Johnson et al. (2002b) demonstrated, animals may use different criteria when choosing movement routes relative to foraging or other types of habitat selection. Thus, we used a multi-scale movement model (Johnson et al. 2002c) to identify large-scale movement paths of caribou during winter (December 1–March 31; Johnson et al. 2002c), not a collection of foraging decisions. This allowed us to address the common assumption that resistance to movement is the simple opposite of general strength of selection for habitats (Beier et al. 2008).
Previous studies for woodland caribou reported behavioural thresholds in movement defined by distinct breaks in the frequency distribution of movement rates. The reported breakpoints corresponded with two broad scales of movement: intrapatch (short distance) and interpatch (long distance) (e.g., Johnson et al. 2002b, c; Saher and Schmiegelow 2005). Using these data (Johnson et al. 2002b), we identified locations as large-scale movements if they exceeded a movement rate that equated to a mean straight-line distance of 3,738 m within a 24-h period.
Generating least-cost paths
One common method for predicting suitable areas for conservation corridors is the least-cost path model (Chetkiewicz et al. 2006). Accurate weights are the most important component of the habitat permeability model and often originate from habitat suitability or selection maps (Clevenger et al. 2002; Chetkiewicz and Boyce 2009). Furthermore, permeability maps are premised on the assumption that animals will choose the most ‘suitable’ (permeable) habitat to make their movements, even though this habitat might not be the most suitable for other functions such as feeding, breeding or resting (Walker and Craighead 1997; Beier et al. 2008). Techniques for generating habitat suitability maps include Mahalanobis distance (Clevenger et al. 2002; Johnson and Gillingham 2005), RSFs (Johnson et al. 2002b; Chetkiewicz et al. 2006), weights-of-evidence (Kindall and Van Manen 2007), and expert models based on either literature or expert opinion (Walker and Craighead 1997; Ray et al. 2002; Singleton et al. 2002; Johnson and Gillingham 2005).
We used RSF and expert-based models to derive habitat permeability maps. Both maps were developed according to a consistent suite of explanatory variables: landcover, distance to predation risk, distance to waterbodies, distance to roads, elevation, slope, and aspect. Landcover was derived from a supervised classification of Landsat Thematic Mapper imagery (25 × 25-m resolution) that was consolidated into 10 cover types (Johnson et al. 2003). Predation risk was a continuous variable that represented the distance of each cell across the study area from high-risk landcover types identified using radiolocations from wolves (see Johnson et al. 2002b). Distance to roads and distance to water-bodies were calculated using the British Columbia Terrain Resource Information Management GIS data. Slope, aspect and elevation were continuous variables obtained from a Digital Elevation Model of the study area (25 × 25-m resolution).
Resource selection function
Resource selection functions weight the influence of each model covariate on the relative probability of occurrence of caribou across the study area (Manly et al. 2002). We used conditional fixed effects logistic regression to estimate coefficients for the RSFs. Conditional logistic regression differs from traditional logistic regression in that animal use and corresponding random locations are matched (Hosmer and Lemeshow 2000). This allows the model to control for temporal and spatial variation in resource availability. We paired each long-distance caribou movement location with a random location to represent the availability of resources. These random sites were generated from within a circle with a radius equal to the 95th percentile of the movement distance for that relocation interval and centred on the previous caribou location.
We identified a number of candidate logistic regression models that served as hypotheses to test various factors influencing the long-distance movements of woodland caribou. We then used the Akaike’s information criterion difference for small samples (ΔAICc) to identify the most parsimonious of the candidate RSF models. The model with the lowest AICc does not necessarily have good discriminatory capacity (Pearce and Ferrier 2000). We used k-fold cross validation to evaluate the predictive performance of the most parsimonious RSF model (Boyce et al. 2002). We performed the k-fold procedure five times, each time randomly withholding 20% of the data. For each iteration, we used a Spearman rank correlation to assess the relationship between predicted caribou occurrence and their frequency within 10 equally sized bins derived from the range of predicted relative probabilities. We screened each model for high levels of multicollinearity and influential cases (Menard 1995).
We used a linear transformation to normalise the RSF values between 0 and 1:
where w(x) is the relative probability of occurrence of caribou making long-distance movements, w min is the minimum, and w max, the maximum RSF values.
Expert-based model
We used the Analytical Hierarchy Process (AHP) to generate the expert-based permeability map (Saaty 1977). The AHP worked by asking experts to systematically identify relative weightings for each paired combination of explanatory variables. Individual layers were then combined to produce the habitat permeability map. When developing the matrix, the experts used a 9-point ordinal scale to rate each pairing of the categorical biophysical variables. For example, each expert was asked to compare and score the relative importance of landcover type relative to topographic slope. Because pairwise comparison matrices have multiple paths by which the importance of criteria can be assessed, it was necessary to determine the consistency ratio of the matrix. For this study, experts were asked to re-evaluate matrices when consistency ratios were >0.1 (Saaty 1977). Consistency ratios and weights for habitat variables were calculated using the WEIGHT procedure in Idrisi (Eastman 2004).
We solicited the input of ten experts. An expert was defined as any person who had published a research report or paper on the ecology of northern woodland caribou or, through professional duties, had spent >5 years contributing to management guidelines for northern woodland caribou. Experts were sent a questionnaire and asked to provide a pair-wise score of each combination of the seven explanatory variables used in the study. Each variable was multiplied by the mean weight given by the experts and then overlaid upon the other layers to produce the permeability map.
Evaluation of permeability maps
We reclassified the habitat permeability maps into 10 ranked habitat classes of approximately equal area, with rank 1 being of the highest quality habitat and rank 10 being of the lowest quality. Then, we superimposed the independent caribou locations classified as large-scale movements (S. McNay, Personal Communication) on the permeability maps and calculated the density (locations/km2) of locations for each habitat class. Finally, we used a Spearman rank correlation to assess the relationship between the density of locations associated with long-distance movements and habitat class.
Generating cost maps and least cost paths
We converted the RSF and expert-based permeability maps to least-cost maps by first normalizing their respective values between 0 and 1 (e.g., Eq. 1 for RSF). Then, the inverse of the normalized values was taken (Eq. 2) resulting in the cost maps.
The strongest measures of selection represented the lowest costs of movement, thus, the combination of resources classified as habitat for large-scale movement became low resistance habitats. These low resistance cells had values near 0 while the cells with the highest cost had values closer to 1. Once we generated the cost maps, we used the least-cost function of ArcGIS software (ESRI 2005) to predict the paths between the start and end point of inferred caribou movement paths (i.e., constructed from movement locations).
Path verification
The least-cost model predicts paths that an animal might take among important habitat patches. If the least-cost model is a useful method for selecting effective movement corridors, then the spatial locations of actual movement routes and routes predicted using the model should be similar. It is possible that paths that are distant from the single best (least-cost) path also have relatively low costs. While these paths may be suitable movement routes, an assumption of this study is that the best paths are the ones that the animals choose to follow. Conversely, in highly heterogeneous landscapes, areas that are close to the least-cost path may have high costs of movement (e.g., a path along the edge of a cliff). However, contiguous areas corresponding closely to areas that animals are using for movement are likely to be effective corridors and preferentially selected by agencies making land-use decisions. It is less likely that there will be errors in the least-cost method if the distance between actual movement paths and the least-cost path is small.
As an approximation of actual movement paths, we plotted GPS location data identified as long-distance movements (sensu Johnson et al. 2002b; Fig. 1). For each individual caribou for a given year, we chose the two most distant locations as the start and end point for the movement path. The GPS locations between these two locations were joined sequentially to create an inferred path. We then used the Path Deviation Index (PDI; Jan et al. 1999) to compare the inferred with the closest predicted least-cost movement paths generated using the RSF and expert-based permeability maps (Fig. 1). The PDI is interpreted as the average distance between two paths and is calculated as the area between paths divided by the shortest path (straight line distance) between the start and end points:

The process of reconstructing movement paths (top). The image on the left shows caribou locations (black triangles) for one movement path. The inferred (white) and least-cost (black) paths, shown on the right are generated between the two most distant points for an individual animal’s path. The remaining triangles represent short distance intrapatch movements. Generation of a random path (black) from the inferred (white) caribou path is by random redistribution of path segments (middle). Measurement of area between paths (bottom) is used to calculate PDI. Movement paths are superimposed on a permeability matrix constructed from an expert-based model for the Wolverine caribou herd of northcentral British Columbia, Canada
For comparisons among multiple movement paths (i.e. paths with differing start and end points), a normalised PDI is calculated by dividing the PDI again by the shortest path distance:
The PDI is premised on area and, thus, will not capture all aspects of the variation between inferred and predicted movement paths. Path sinuosity is a second measure for assessing differences between movement paths. We calculated path sinuosity as total path length, divided by the shortest possible distance of the path (straight line from start to finish; Beyer 2004).
The PDI is a relative measure of difference between paths, but does not serve as a formal test of the performance of predictive least-cost models. Thus, we used random paths as null models for comparison with both inferred and predicted paths. A random path would be the path chosen by an animal if it were to have no habitat preference for movement and no constraints regarding time available or distance travelled. If the PDI between the predicted and inferred path is less than the PDI between the random and inferred path, then the least-cost path is a better predictor of movements than the random path. We also generated a straight line from the start to the end location of each inferred path. This second null model is of interest because in a homogeneous landscape an animal that seeks to reach its goal with a minimum of time and distance travelled will choose the shortest straight-line path. In a heterogeneous environment where selection of movement paths provides a fitness advantage, we predicted that the PDI between the predicted least-cost and inferred paths would be less than that of the straight line path. PDI is a meaningful measure of the difference between modelled and observed movements, and can help evaluate least-cost path models including the range of parameterizations (Beier et al. 2008).
We generated random paths using the ArcView algorithm alternate animal movement routes (Jenness 2004). For each inferred caribou path, the algorithm identified route segments separating consecutive animal locations and redistributed those segments in a random order and direction. The random path was constrained to the start and end point of the paired inferred path from which the path segments originated. This method is biased as it depends on pre-existing caribou movement data. Fewer path segments and a straighter inferred path will result in a random path with less opportunity to deviate from the paired inferred path. However, a truly random path (one that is not limited in any way by characteristics of the animal) may, in theory, cover an entire landscape before reaching a predetermined end point (de Smith 2004).
We used three repeated measures ANOVAs to test for mean differences in path metrics calculated for predicted, random or inferred movement paths of caribou. Where sphericity was violated (Mauchly’s test) we used the Greenhouse–Geisser corrected F values. For all analyses, path type (inferred, linear, random, predicted) served as a within subjects effect. We used the Sidak post hoc test to determine statistical differences (α = 0.05) among path type. For the first ANOVA, we tested for statistical differences in mean PDI between predicted (RSF/expert vs. inferred) and null (random/straight vs. inferred) movement paths. This repeated measures test included cost surface method (RSF/expert) as a second within subjects effect. For the second ANOVA, the cumulative cost of the predicted or inferred movement paths, as represented on the cost surfaces, were summed and mean differences among path types were compared. Finally, we used ANOVA to test for differences in mean sinuosity between the inferred and predicted paths.
Results
Permeability maps
We used 596 locations describing long-distance movements of female caribou to construct seven candidate RSF models. The model consisting of variables for landcover, slope, distance to water, and a nonlinear elevation term was the most parsimonious (Table 1). The selection coefficients suggested that during long-distance movements caribou avoided the pine and deciduous land cover types and traversed low-elevation, flat terrain near waterways (Table 2). A mean Spearman rank correlation of 0.798 (P = 0.006) indicated good predictive accuracy; we used this model to produce the RSF permeability map.
We used 341 independent caribou locations (S. McNay, Personal Communication) to test the ability of the most parsimonious RSF model to identify habitat classes used by caribou during long-distance movements. The Spearman rank correlation between the density of caribou locations and the 10 predicted habitat classes (Fig. 2) was significant (r s = 0.867, P = 0.001) suggesting that the RSF model was a good predictor of habitats used for movement.

Ranked habitat permeability class maps derived from an expert-based model (left) and a RSF (right). Maps were developed using GPS locations from woodland caribou from the Wolverine herd of northcentral British Columbia, Canada. White dots represent independent validation locations for caribou of the same herd
Six independent experts with a total of over 75 years experience studying northern woodland caribou responded to the survey. A number of matrices from the contributing experts had high consistency ratios (i.e., >0.1) and were returned to the experts. Following the re-evaluation, four matrices provided by one expert still had high consistency ratios. Due to a small sample size, only one of the matrices (landcover, consistency ratio = 0.29) was rejected, while three with marginal consistency ratios (predation risk, consistency ratio = 0.13; slope, consistency ratio = 0.11; among variables comparison, consistency ratio = 0.10) were retained. On average, experts reported that slope was the most important landscape feature influencing the selection of movement corridors by caribou (Table 3). Unlike the RSF permeability map, the Spearman rank correlation between the density of caribou locations and the predicted habitat classes (Fig. 2) was not significant (r s = 0.455, P = 0.187; Fig. 3). This suggested that the expert’s characterization of movement habitats differed from those habitats actually used by caribou.

Predictive accuracy of habitat maps describing long-distance movements of woodland caribou of the Wolverine herd. The maps were generated using a RSF (top) and expert-based model (bottom). Habitat classes were ranked from 1 (highest habitat suitability) to 10 (lowest suitability); the normalised density of independent caribou locations within each habitat class is shown. Habitat maps were used to construct permeability maps for the least-cost path model
Caribou movements and least-cost paths
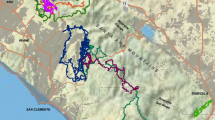
A total of 14 movement paths were created from 174 caribou locations identified as long-distance inter-patch movements. During the winters, the majority of monitored caribou moved from south to north through similar low-elevation valley bottoms (Fig. 4). Although the study area was large, movement paths were confined to relatively well defined corridors.

The study area indicating the home range of the Wolverine caribou herd, northcentral British Columbia, Canada, and the movement paths used for validation of the least-cost movement path models. Movement paths were constructed with location data collected using GPS collars from 14 individual female caribou
Path Deviation Index scores varied considerably among paths and methods used to define the path (Table 4). On average, PDI scores for the expert (\( \overline{X} = 7.556\), SE = 1.248) and RSF (\( \overline{X} = 8.719\), SE = 1.331) cost surfaces were less than the mean PDI for the random paths (\( \overline{X} = 10.000 \), SE = 2.226), but greater than the straight line paths (\( \overline{X} = 6. 381\), SE = 1.137). The differences in PDI scores, however, were not statistically significant (F (2.029,26.372) = 1.530, P = 0.235). In contrast, the ANOVA revealed a statistically significant difference in mean least cost scores totalled for paths that traversed the expert or RSF-based cost surfaces (F (1,13) = 21.262, P < 0.001), among path types (F (1.005,13.064) = 5.419, P = 0.036), and the interaction of those main effects (F (1.102,14.326) = 7.880, P = 0.012). The mean cumulative cost of the predicted least-cost paths generated with the expert and RSF cost surfaces were less than the inferred paths (Fig. 5). Post-hoc analyses, however, did not reveal significant differences among average cost scores by path type (all P ≥ 0.083). The sinuosity of movement paths differed statistically among type (F (1.022,13.283) = 5.008, P = 0.042). On average, inferred paths (\( \overline{X} =1.818\), SE = 0.312) were more sinuous than predicted paths using the expert (\( \overline{X} = 1.191\), SE = 0.035) or RSF surfaces (\( \overline{X} = 1.061\), SE = 0.006). The post-hoc test revealed a statistically significant difference in sinuosity between the predicted paths only (P = 0.013).

Mean cost scores (standard error) generated for paths representing inferred caribou movements, random and straight line paths, and least cost paths predicted using a RSF and an expert-based model
Discussion
Our results suggest that the least-cost path model is a poor predictor of the precise spatial location of movement paths generated by woodland caribou. We must caution, however, that application of this technique to different species and landscapes may yield more useful results. Furthermore, application of the least-cost logic to wider, more general corridors, rather than the ‘single-best’ (1-pixel wide) least-cost path, might result in acceptable results for conservation planning. In an effort to generalize findings and explore limitations of the technique, we pursued a number of variants in method and evaluation metrics. Regardless of the cost map, however, a straight line connecting a source and target patch was just as effective at predicting the movement corridors of caribou as the more sophisticated least-cost approach. Such counterintuitive results demand a careful evaluation of source data and constituent methods.
Independent validation data revealed that the RSF model was a good predictor of caribou distribution relative to expected long-distance movements (sensu Johnson et al. 2002b). In comparison, the experts did not accurately describe the resources that woodland caribou would select when moving across the study landscape. Previous authors have identified shortcomings with eliciting or applying expert opinion to the description of species distribution (Clevenger et al. 2002; Johnson and Gillingham 2004). In particular, expert-based species distribution models appear to underperform compared to empirically based models (Johnson and Gillingham 2005); however, definition of expert and choice of elicitation method appear to be key decisions during study design (Hurley et al. 2009; Murray et al. 2009). For this study, we do not feel that selection of experts had a significant impact on the accuracy of habitat maps. Six experts contributed over 75 years combined experience and many had excellent knowledge of the Wolverine herd. In contrast, RSFs are generally good predictors of animal distribution (e.g., Johnson et al. 2004; Fortin et al. 2005), as was the case in the current study. That both methods resulted in poor accuracy of predicted animal movement paths suggests shortcomings in the least-cost method.
Although the results were not statistically significant, the paths predicted using the least-cost method had lower cumulative costs when compared to the inferred paths and the null models (Fig. 5). While the method was successful in identifying low-cost movement routes, the PDI revealed that these routes did not correspond to the actual animal movement paths and may be a poor choice for designing effective corridors. Of note, the cumulative costs of inferred animal movement paths were variable, but on average greater than the predicted paths. On the basis of these findings, caribou were not following a path that would be considered a ‘low-cost’ route relative to the predictions of the least-cost method.
Visual inspection of the inferred caribou movement paths suggested a tendency for some non-directional shorter distance movements as well as directional long distance movements (Fig. 1). The least-cost movement model does not consider these non-directional movements that may be due to feeding or some other relatively stationary behaviour. Reassessment of the data to identify only directed movements between seasonal ranges may improve the results; although, we fit models to large-scale movement data only. Examination of a different season may also reveal less variable, directed movements (Saher and Schmiegelow 2005). Also, larger spatiotemporal scales of movement may provide a stronger fit of observation to least-cost path models. Regardless, the woodland caribou we observed did move continuously across the landscape covering a considerable area; mean average path length was 24,752 m (SD = 17,769; range = 6,729–124,625 m). Given these distances and observed directional travel, we should expect a lower PDI between the predicted and inferred paths when compared to random. Similarly, we should expect comparable measures of path sinuosity between predicted and inferred paths.
For this analysis, and most applications of the least-cost path model, there is a mismatch between the resolution of inferred and predicted paths. The predicted paths were based on a cell resolution of 25 m, while the observed caribou movement points were taken at least 3 h apart resulting in a coarser spatial resolution. While individual caribou locations are accurate to 3–8 m, the movement vector that occurs between locations is inferred. This problem is possibly exacerbated by the exclusion of the short ‘intrapatch’ movements that connect the long-distance movements. However, removal of these short-distance locations resulted in very little change to the final inferred movement path.
We recognise a temporal disconnect between the permeability of the present, relatively undisturbed landscape and the future landscape that we assume will have a much more hostile matrix for caribou and fewer options for transit. Caribou had access to a wide range of resources during their movements suggesting the use of habitat linkage zones or corridors in contrast to more well defined travel paths (Kindall and Van Manen 2007). We suspect that the performance of the least-cost method would improve for populations forced through ‘bottlenecks’ of relatively narrow travel corridors in an otherwise disturbed landscape. Regardless of the relative lack of disturbance across the study area, our analyses demonstrated that there was a sufficient cost gradient to allow significant differences in the cumulative cost of inferred and simulated paths. In more fragmented landscapes, we would expect to find an even greater contrast in cumulative costs.
When compared to specific narrow paths that are the product of least-cost path models, corridor models likely have greater utility for identifying movement zones of value to conservation planning (Beier et al. 2008). However, the least-cost method, if effective, could help identify a single or series of adjacent paths that can be integrated into wider habitat corridors. We illustrate the utility of one least-cost path to identify the anchor point for a corridor: progressively darker areas of Fig. 6 indicate areas of value for movement that may be included as part of the larger conservation corridor. The effectiveness of the least-cost method in capturing important movement routes would depend on the width of the corridor, although for practical reasons, narrow corridors are favoured by land managers (Beier et al. 2008).

The cumulative cost surface for one movement path, showing the predicted (least-cost) movement path corresponding with the area of lowest cumulative cost, and the actual movement path taken by a caribou (inferred path)
There is uncertainty as to how well the inferred caribou paths represented actual movement corridors. We recognize these limitations in our application of the least-cost model and corresponding test, however, the resolution and frequency of the movement data are excellent when compared to other techniques for tracking animal movement (e.g., VHF/satellite collars, visual observation). Currently, these data serve as a best-case scenario for empirically monitoring and predicting animal movement over large temporal and spatial scales (e.g., Kautz et al. 2006; Kindall and Van Manen 2007).
We noted two previous studies that attempted to independently evaluate the effectiveness of GIS-based techniques designed to predict animal movement corridors (Kehoe 1995; Clevenger et al. 2002). Both of these studies were carried out on different species in areas with a higher degree of disturbance and fragmentation. Differences among studies emphasize the need for effective strategies that are species and population specific. We have presented one case that represents a population of conservation concern that still inhabits relatively intact habitats. Clearly, this is not the situation for all woodland caribou populations, most of which are found across fragmented historical ranges (Poole et al. 2000; Apps and McLellan 2006). When defining corridors or linkage networks, practitioners have a number of choices among information sources and planning strategies. Our observational data suggest that purely predictive models are ineffective for this purpose, but we worked with a population that still has many corridor options and we attempted to represent well defined movement paths. Furthermore, some groups of experts may have insufficient knowledge to predict or identify movement paths or corridors (Hurley et al. 2009). Although the Wolverine herd is well studied (Johnson et al. 2002a), the experts may not be familiar with the relatively infrequent long-distance movement behaviour.
By providing a tool for path validation, the techniques presented here will assist with the development of more effective GIS methods to predict movement corridors. Improved GIS techniques are important as poorly designed corridors may act as a population sink, a reservoir for edge species, aid in the spread of catastrophes such as fire; or squander precious financial and stakeholder support (Simberloff and Cox 1987; Simberloff et al. 1992; Beier et al. 2008). Results from the PDI suggest that the least-cost corridor method is no better at predicting actual animal movements than a straight line or random path. However, considering the sound theoretical foundation and ease of application of the least-cost technique and some limitations of our study design we must temper our criticisms. Without replication with different species and landscapes, we cannot definitively reject the least-cost method. While we await further evaluation of the technique, conservation planners and land managers should approach the use of the least-cost corridor method with caution, especially if derived from expert-based models.
References
Apps CD, McLellan BN (2006) Factors influencing the dispersion and fragmentation of endangered mountain caribou populations. Biol Conserv 130:84–97
Beier P, Majka DR, Spencer WD (2008) Forks in the road: choices in procedures for designing wildland linkages. Conserv Biol 22:836–851
Beyer HL (2004) Hawth’s analysis tools for ArcGIS. http://www.spatialecology.com/htools
Boyce MS, Vernier PR, Nielsen SE, Schmiegelow FKA (2002) Evaluating resource selection functions. Ecol Model 157:281–300
Chetkiewicz CLB, Boyce MS (2009) Use of resource selection functions to identify conservation corridors. J Appl Ecol 46:1036–1047
Chetkiewicz CLB, St. Clair CC, Boyce MS (2006) Corridors for conservation: integrating pattern and process. Annu Rev Ecol Syst 37:317–342
Clevenger AP, Wierzchowski J, Chruszcz B, Gunson K (2002) GIS-generated expert based models for identifying wildlife habitat linkages and mitigation passage planning. Conserv Biol 16:503–514
Couvet D (2002) Deleterious effects of restricted gene flow in fragmented populations. Conserv Biol 16:369–376
Csuti B (1991) Conservation corridors: countering habitat fragmentation. In: Hudson WE (ed) Landscape linkages and biodiversity. Island Press, Washington, pp 81–90
de Smith MJ (2004) Distance and path: the development, interpretation and application of distance measurement in mapping and spatial modelling. PhD thesis, University of London, University College, Department of Geography, London
Eastman JR (2004) IDRISI Kilimanjaro, version 14.01. Clark University Laboratory, Clark University, Worcester
ESRI (2005) ArcGIS—ArcInfo, version 9.1. Environmental Systems Research Institute, Inc, Redlands
Fortin D, Beyer HL, Boyce MS, Smith DW, Duchesne T, Mao JS (2005) Wolves influence elk movements: behavior shapes a trophic cascade in Yellowstone National Park. Ecology 86:1320–1330
Goncalves AB (2010) An extension of GIS-based least cost path modelling to the location of wide paths. Int J Geogr Inf Sci 24:983–996
Harris LD (1984) The fragmented forest: island biogeography theory and the preservation of biotic diversity. The University of Chicago Press, Chicago
Hoekstra JM, Boucher TM, Ricketts TH, Roberts C (2005) Confronting a biome crisis: global disparities of habitat loss and protection. Ecol Lett 8:23–29
Hosmer DW, Lemeshow S (2000) Applied logistic regression, 2nd edn. Wiley, New York
Hurley MV, Rapaport EK, Johnson CJ (2009) Utility of expert-based knowledge for predicting wildlife-vehicle collisions. J Wild Manag 73:278–286
Jan O, Horowitz AJ, Peng Z-R (1999) Using GPS data to understand variations in path choice. Paper presented at the 78th meeting of the Transportation Research Board, Washington
Jenness J (2004) Alternate animal movement routes (altroutes.avx) extension for ArcView 3.x, v. 2.1. Jenness Enterprises. http://www.jennessent.com/arcview/alternate_routes.htm
Johnson CJ, Gillingham MP (2004) Mapping uncertainty: sensitivity of wildlife habitat ratings to variation in expert opinion. J Appl Ecol 41:1032–1041
Johnson CJ, Gillingham MP (2005) An evaluation of mapped species distribution models used for conservation planning. Environ Conserv 32:117–128
Johnson CJ, Heard DC, Parker KL (2002a) Expectations and realities of GPS animal location collars: results of three years in the field. Wild Biol 8:153–159
Johnson CJ, Parker KL, Heard DC, Gillingham MP (2002b) A multiscale behavioural approach to understanding the movements of woodland caribou. Ecol Appl 12:1840–1860
Johnson CJ, Parker KL, Heard DC, Gillingham MP (2002c) Movement parameters of ungulates and scale-specific responses to the environment. J Anim Ecol 71:225–235
Johnson CJ, Alexander ND, Wheate RD, Parker KL (2003) Characterizing woodland caribou habitat in sub-boreal and boreal forests. For Ecol Manag 180:241–248
Johnson CJ, Seip DR, Boyce MS (2004) A quantitative approach to conservation planning: using resource selection functions to identify important habitats for mountain caribou. J Appl Ecol 41:238–251
Kautz R, Kawula R, Hoctor T, Comiskey J, Jansen D, Jennings D, Kasbohm J, Mazzotti F, McBride R, Richardson L, Root K (2006) How much is enough? Landscape-scale conservation for the Florida panther. Biol Conserv 130:118–133
Kehoe NM (1995) Grizzly bear distribution in the north fork of the Flathead River valley: a test of the linkage zone prediction model. MS thesis. University of Montana, Missoula
Kindall JL, Van Manen FT (2007) Identifying habitat linkages for American black bears in North Carolina, USA. J Wild Manag 71:487–495
Manly BFJ, McDonald LL, Thomas DL, McDonald TL, Erickson WP (2002) Resource selection by animals: statistical design and analysis for field studies. Kluwer, Dordrecht
Menard S (1995) Applied logistic regression analysis. Quantitative applications in the social sciences series no. 07-106. Sage University, Thousand Oaks, CA
Merriam G (1991) Corridors and connectivity: animal populations in heterogeneous environments. In: Saunders DA, Hobbs RJ (eds) Nature conservation 2: the role of corridors. Surrey Beatty and Sons Pty Ltd, Chipping Norton, pp 133–142
Murray JV, Goldizen AW, O’Leary RA, McAlpine CA, Possingham HP, Choy SL (2009) How useful is expert opinion for predicting the distribution of a species within and beyond the region of expertise? A case study using brush-tailed rock-wallabies Petrogale penicillata. J Appl Ecol 46:842–851
Noss RF (1991) Landscape connectivity: different functions at difference scales. In: Hudson W (ed) Landscape linkages and biodiversity. Island Press, Washington, pp 27–38
Pace F (1991) The Klamath corridors: preserving biodiversity in the Klamath National Forest. In: Hudson W (ed) Landscape linkages and biodiversity. Island Press, Washington, pp 105–116
Pearce J, Ferrier S (2000) Evaluating the predictive performance of habitat models developed using logistic regression. Ecol Model 133:225–245
Poole KG, Heard DC, Mowat G (2000) Habitat use by woodland caribou near Takla Lake in central British Columbia. Can J Zool 78:1552–1561
Ray N, Lehmann A, Joly P (2002) Modeling spatial distribution of amphibian populations: a GIS approach based on habitat matrix permeability. Biodivers Conserv 11:2143–2165
Richard Y, Armstrong DP (2010) Cost distance modelling of landscape connectivity and gap-crossing ability using radio-tracking data. J Appl Ecol 47:603–610
Ricketts TH (2002) The matrix matters: effective isolation in fragmented landscapes. Am Nat 158:87–99
Rouget M, Cowling RM, Lombard AT, Knight AT, Graham IHK (2006) Designing large-scale conservation corridors for pattern and process. Conserv Biol 20:549–561
Saaty TL (1977) A scaling method for priorities in hierarchical structures. J Math Psychol 15:234–281
Saher DJ, Schmiegelow FKA (2005) Movement pathways and habitat selection by woodland caribou during spring migration. Rangifer 16:143–154
Simberloff DS, Cox J (1987) Consequences and costs of conservation corridors. Conserv Biol 1:63–71
Simberloff DS, Farr JA, Cox J, Mehiman DW (1992) Movement corridors: conservation bargains or poor investments? Conserv Biol 6:493–504
Singleton PH, Gaines WL, Lehmkuhl JF (2002) Landscape permeability for large carnivores in Washington: a geographic information system weighted-distance and least-cost corridor assessment. U.S. Department of Agriculture, Pacific Northwest Research Station, Portland
Soulé ME, Mackey BG, Recher HF, Williams JE, Woinarski JCZ, Driscoll D, Dennison WC, Jones ME (2004) The role of connectivity in Australian conservation. Pac Conserv Biol 10:266–279
Walker R, Craighead L (1997) Least-cost-path corridor analysis: analyzing wildlife movement corridors in Montana using GIS. In: Proceedings of the ESRI user’s conference, San Diego
Acknowledgments
We thank M. Gillingham and J. Kirkpatrick for insightful comments during the development of this paper. P. Beier and two anonymous reviewers provided a thorough review of the manuscript. Following submission, the Coordinating Editor, H. Wagner, skilfully navigated the paper through the review process and was instrumental in improving the final version. S. McNay kindly donated unpublished caribou locations that allowed us to evaluate the predictions of the RSF and expert-based models.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.

{kind=link}

{kind=link}

{kind=link}

{kind=link}
Rights and permissions
About this article
Cite this article
Pullinger, M.G., Johnson, C.J. Maintaining or restoring connectivity of modified landscapes: evaluating the least-cost path model with multiple sources of ecological information. Landscape Ecol 25, 1547–1560 (2010). https://doi.org/10.1007/s10980-010-9526-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10980-010-9526-6