
Abstract
Odonata species can be used to indicate the effects of anthropogenic disturbance on agrobiodiversity. In Japan, populations of some odonate species inhabiting rice paddy fields have decreased strongly owing to their high susceptibility to insecticides, and they therefore need to be conserved. To identify effective conservation strategies, we investigated how agricultural practices (conventional vs. environmentally friendly: EF) and surrounding landscape (area of forest at three spatial scales, namely 50, 100 and 200 m from the edge of the paddy field) influenced the abundance of odonates in rice paddies in Tochigi Prefecture, Japan. Generalized linear mixed-effect models revealed that EF farming increased the abundance of odonate adults and the number of nymphal exuviae, suggesting a strong negative effect of insecticides. The influence of forest area at the local scale on the numbers of odonate adults and nymphal exuviae was small, with the exception of a strong negative influence on adult Sympetrum infuscatum: the presence of a large area of forest within 200 m of the edge of the paddy field reduced their abundance, probably reflecting this dragonfly’s oviposition site preference. Our results suggest that EF farming could generally support the conservation and recovery of local populations of odonates, but its effectiveness potentially varies depending on the ecological aspects of each species.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Conservation of biodiversity in agroecosystems is important for sustainable agriculture. To this end, environmentally friendly (EF) farming, in which the use of agrochemicals and inorganic fertilizers is reduced, is practiced around the world (Willer et al. 2011). EF farming has proved effective at enhancing biodiversity in various taxa in Europe (Hole et al. 2005; Tuck et al. 2014; but see Kleijin and Sutherland 2003; Kleijn et al. 2006). However, as its effectiveness depends on the surrounding landscape as a source habitat (Tscharntke et al. 2005; Batáry et al. 2010; Concepción et al. 2012; Merckx et al. 2009), it is necessary to evaluate the effects of both agricultural practices and land cover within the surrounding landscape to establish effective strategies for conserving agrobiodiversity.
Asian rice paddy ecosystems are hotspots of biodiversity because they function as alternative wetland habitats for many aquatic and semi-aquatic organisms that rely on temporary water bodies (Kiritani 2000; Lawler 2001; Bambaradeniya and Amerasinghe 2004; Usio and Miyashita 2015). However, modern intensive agriculture and farmland consolidation have harmed many species, for example through reduced survival and loss of oviposition sites as a result of changes in paddy field irrigation (Katayama et al. 2015). Consequently, many species have declined and are now endangered (Ministry of the Environment, Japan 2018). As diverse species provide pest control services, a decline of predator diversity could lead to pest outbreaks (Kiritani 2000). Therefore, EF farming would seem to be important in maintaining both biodiversity and ecosystem services in paddy field ecosystems. Several studies have investigated the effects of EF farming on paddy field organisms in Asian countries. However, they are limited to anurans (Kato et al. 2010; Uruma et al. 2012) and spiders (Amano et al. 2011; Takada et al. 2014; Tsutsui et al. 2016; Baba et al. 2018). Furthermore, very few studies have tested the interactive effects of agricultural practices and landscape factors on organisms (Uruma et al. 2012; Tsutsui et al. 2016).
Odonate species such as Sympetrum dragonflies and coenagrionid and lestid damselflies are representative of organisms that depend on paddy fields as habitat. These species use paddy fields as oviposition sites and nymphal habitats, with varying degrees of dependence on these sites. Sympetrum dragonflies, in particular, show strong dependence on rice fields as nymphal habitats, oviposition sites, and overwintering sites, although emerged adults leave the paddy fields for nearby forests or for mountainous areas (Ueda 1998a). Coenagrionids use both paddy fields and stable water bodies, such as ponds, as habitats. Their nymphs develop in paddies mainly in the first generation, because paddy fields are usually drained intermittently from summer and the nymphs require stable water for overwintering (Sugimura et al. 1999; Ozono et al. 2012). Some species of lestids overwinter as eggs and others as adults. In spring, overwintering eggs hatch and overwintering adults migrate into paddies to reproduce (Sugimura et al. 1999; Ozono et al. 2012).
Some odonates play an important role in regulating pest insects in paddy fields by preying on mosquito larvae and rice pests (Urabe et al. 1990). Unfortunately, populations of paddy-dwelling odonates—particularly Sympetrum species—have declined dramatically in recent years owing to changes in the agricultural environment. Among potential factors, synthetic insecticides such as fipronil and neonicotinoids are considered to be particularly detrimental (Jinguji et al. 2009, 2010, 2018; Aoda et al. 2013; Hayasaka et al. 2013; Ueda and Jinguji 2013). For example, as the use of these insecticides has increased, numbers of Sympetrum frequens have decreased (Futahashi 2012; Nakanishi et al. 2018); these and some damselfly species (e.g., Mortonagrion selenion, Ischnura asiatica, and Indolestes peregrinus) are listed as endangered in several prefectures (Ministry of the Environment, Japan; https://ikilog.biodic.go.jp/Rdb/pref). Therefore, EF farming, which minimizes the use of insecticides, may mitigate the decline of these populations. However, most previous studies were based on small-scale mesocosm experiments (Oyama and Kidokoro 2003; Hayasaka et al. 2013), and the few field studies (e.g., Aoda et al. 2013) did not consider the effect of the surrounding landscape. Because adult odonates move to different habitats after emergence, the combination of paddy fields and other habitats should be important for maintaining populations. Particularly in Japan, paddy fields are small and the effect of the surrounding landscape can be relatively strong (Washitani 2007), so to establish effective conservation practices it is important to identify landscape effects.
Here, we investigated the effects of EF farming, the surrounding landscape, and their interaction on several odonate species in Japanese paddy fields. Because the forest edge is considered to be an important habitat for some dragonfly species (e.g., Sympetrum infuscatum; Ueda 1998b; Ozono et al. 2012), we focused on forest area as a landscape factor. We posed the following questions: (1) Does the effect of EF farming on odonate assemblages depend on the surrounding landscape? (2) Do the effects of farming practices, landscape, and their interaction differ among odonate species? (3) Do landscape effects and their effective spatial scale differ among species, reflecting different odonate mobilities and habitat requirements (e.g., Hamasaki et al. 2011; Raebel et al. 2012a).
Methods
Study area
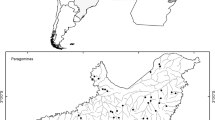
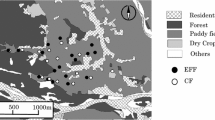
We conducted a field survey in 2012 and 2013 at Shioya-cho, Tochigi Prefecture, in the northern part of the Kanto region of Japan, in an area about 1.5 km × 1.5 km square. We surveyed 10 EF paddy fields in both years, and seven conventional farming (CF) paddy fields in 2012 and 10 in 2013 (Fig. 1). The size of the fields ranged from 1366 to 8586 m2. The location of the fields along a geographic gradient from simple arable fields to forest-rich landscape allowed us to test the influence of land cover within the surrounding landscape. Paddy-dwelling spiders were previously surveyed in the same fields (Baba et al. 2018). The major difference between the EF and CF practices in the survey area was in insecticide use (Table 1). No insecticides had been used in the EF fields since 2010. However, neonicotinoid, phenylpyrazole, or synthetic pyrethroid insecticides, or combinations of these, had been applied to the seedling nursery boxes and paddy fields in the CF fields. In addition, the fungicides and herbicides applied to the EF fields had lower toxicity to organisms than those applied to the CF fields. Insecticides applied to the nursery boxes are considered to have a great impact on the life cycles of paddy-dwelling odonates (Jinguji et al. 2010). These insecticides are applied to nursery boxes usually just before transplanting and are transferred to the paddy fields in the soil during transplanting. Sympetrum and some lestid species overwinter at the egg stage, and the eggs hatch after the fields are irrigated in spring. Thus, the hatching nymphs are directly exposed to the insecticides. Coenagrionid and some lestid eggs are deposited into irrigated paddy fields by adults that have emerged from the stable water bodies or that overwintered. The deposited eggs hatch within a few weeks and may also be exposed to the insecticides.

Study area in Shioya-cho, Tochigi Prefecture, Japan. Open circle Environmentally friendly, Filled circle conventionally farmed paddy fields. *Paddy fields surveyed only in 2013
Target organisms and sampling protocol
The Odonata are classified into two suborders: Anisoptera (dragonflies) and Zygoptera (damselflies). The dragonflies were dominated by two Sympetrum species, S. frequens and S. infuscatum. These species have a univoltine life cycle in which the adults lay eggs in paddy fields with shallow water during autumn and the eggs hatch in the flooded fields during the next spring. However, their oviposition behaviors and habitat use differ. In S. frequens, newly emerged adults move to distant highlands during summer and return during autumn to lay eggs in wet mud. In S. infuscatum, in contrast, newly emerged adults move to nearby woodlands and forest edges where they feed in order to mature; during autumn they drop their eggs while flying above rice plants before harvest, or above stubble or regrowth after harvest. The damselflies in our study area consisted of four main species: M. selenion and I. asiatica (Coenagrionidae) and I. peregrinus and Lestes temporalis (Lestidae). These damselflies overwinter at the nymphal, adult, or egg stages, and the deposited or overwintering eggs hatch in the paddy fields during spring. The species’ oviposition habits and habitat requirements differ: M. selenion, I. asiatica, and I. peregrinus lay eggs within aquatic plants near the surface of the water after the field is flooded, whereas L. temporalis lays eggs during autumn beneath the bark into shoots of trees overhanging the paddy field.
We surveyed adults of six target odonate species (S. frequens, S. infuscatum, M. selenion, I. asiatica, I. peregrinus, and L. temporalis) during the emergence period, on 21, 22, 27, 28 June 2012 and 28 June 2013. From past investigations, we knew that Sympetrum dragonflies in this area emerged over a short period in late June. We observed three rows of rice plants alongside 20 m of levee at four locations in each field and counted newly emerged adults by eye. Newly emerged adults were differentiated from aged individuals: the former were observed to be still hanging on the nymphal exuviae (Fig. 2, center left panel). we counted only newly emerged adults (referred to here as “adults”). We conducted the surveys on dry sunny or cloudy days. The time required to survey each paddy field was about 15 min. One investigator surveyed EF and CF paddy fields alternately from 08:00 to 14:30. This survey can be completed in a day. However, in 2012, we also conducted a survey of organisms other than odonates at the same time, so we took 2 days for a survey. Therefore, in 2012, it took 4 days to investigate twice. The range in survey timing may have affected the counts of newly emerged individuals, because those that emerged in the morning may have flown away in the afternoon. Additionally, because the survey period was short, it was possible that the overall numbers of odonates were inaccurate. To compensate for these weaknesses, we also counted the numbers of nymphal exuviae of Sympetrum remaining on the rice plants, because the number of nymphal exuviae is a good indicator of the total number of nymphs that developed in a paddy field (Raebel et al. 2010).

Numbers of Sympetrum nymphal exuviae and odonate adults in paddy fields in environmentally friendly (EF) and conventionally farmed (CF) paddies. In box plots, the band inside the box is the median, the bottom and top of the box are the lower and upper quartiles, and the lower and upper whiskers mark ± 1.5 × the interquartile range. For adults of S. infuscatum, the solid line represents the line of fit for EF-farmed paddies and the dashed line represents the line of fit for CF paddies. “Abundance” indicates the number of odonates observed in each field per survey per year. Panels at left illustrate Sympetrum exuvia (top), newly emerged Sympetrum adult (middle), and damselfly adult (bottom)
Landscape metrics
We derived polygon data of land uses in the study area (arable field, open water, forest, road, buildings, and abandoned fields) from a national basic map (1:10 000) and a high-resolution aerial chart in ERDAS Imagine remote-sensing software (Hexagon Geospatial, Madison, AL, USA) and ArcGIS v. 10.1 software (ESRI, Redlands, CA, USA). We measured landscape metrics in ArcView v. 3.10 software (ESRI) and evaluated the area of land dedicated to each use within 50-, 100-, and 200-m buffers from the edges of the study fields. These procedures are the same as those used by Baba et al. (2018). We used the area of forest as the land cover potentially influencing odonate abundance (Hamasaki et al. 2009, 2011; Raebel et al. 2012a). Surrounding paddy fields also influence odonate communities, but there was a strong negative correlation between forest and paddy areas in the study area (arable field vs. forest: 50 m, r = − 0.38, P = 0.10; 100 m, r = − 0.64, P = 0.002; 200 m, r = − 0.84, P < 0.001), so we considered only forest area.
Statistical analysis
To analyze the effects of local agricultural practice (EF vs. CF) and landscape cover (forest area) on the abundance of each odonate species, we used generalized linear mixed-effect models including year and individual paddy field as random factors with a negative binomial distribution (log-link function); the models were implemented in the R package lme4 (Bates et al. 2018). Because we surveyed on 4 days in 2012 (two surveys for each paddy) but on only 1 day each paddy in 2013, we adjusted the survey effort by including the number of surveys in the offset term. The models used the number of odonates or of exuviae of Sympetrum as the response variable and included forest area, agricultural practice, and their interaction as explanatory variables. Owing to the need for re-scaling, forest area was log-transformed. Because the abundance of damselfly species was much less than that of Sympetrum species (see Table 2), we used the total number of all damselfly species and the number of M. selenion, which was the most abundant among them, in the analysis. The model with the lowest Akaike’s Information Criteria (AIC) value was selected as the best model explaining the variability of abundance. First, to specify the effective spatial scale of the landscape, we applied the model to all possible combinations of explanatory variables for each buffer size and compared model performance by AIC. The buffer size producing the lowest AIC value was regarded as the most effective spatial scale. If a model without forest area was selected as the best model, we decided that landscape was not important. Next, to identify explanatory variables influencing odonate abundance, we created models with all variable combinations at the effective spatial scale and calculated the difference from the lowest AIC value (ΔAIC = model AIC − lowest AIC). We considered the model in which ΔAIC < 2 gave the same performance as the best model. When multiple models were identified, we averaged models by using values of Akaike weight (w) (Burnham and Anderson 2002). We determined influential variables from the z-values of their coefficients (estimate/SEM): variables with z > 1.96 were considered to be influential (Burnham and Anderson 2002). All procedures were conducted in the MASS and MuMIn packages (Barton 2018) of R v. 2.3.4 software (R Development Core Team 2014).
Results
We observed a total of 768 nymphal exuviae of Sympetrum dragonflies, 575 adult dragonflies (S. infuscatum and S. frequens), and 333 damselflies (M. selenion, I. asiatica, I. peregrinus, and L. temporalis). The average numbers of individuals in each species appeared to be larger in the EF fields than in the CF fields, and numbers were very small in the CF fields (Table 2; Fig. 2).
For all odonate species, the best models included the effects of EF farming practice (Table 3). All effects of EF farming were positive and influential (z > 1.96), suggesting that EF farming increases the abundances of dragonflies and damselflies. For Sympetrum nymphal exuviae and damselflies, the best model included only agricultural practice. For S. infuscatum, S. frequens, and M. selenion, on the other hand, the best model also included the surrounding forest area, but the effects differed among species. In S. infuscatum, forest area within 200 m of the edge of paddy fields had a strong negative influence (z = − 6.63). In S. frequens, in contrast, the best model included forest area within 50 m and EF farming, but other models with ∆AIC < 2 (a full model and a model including only EF farming) were also selected. The z-values of the averaged coefficients show that only EF farming was influential. In M. selenion, the best model included forest area within 200 m and EF farming, but another model including only EF farming with ∆AIC < 2 was also selected. The z-values of the averaged coefficients show that only EF farming influenced the abundance of this species.
Discussion
EF farming had a positive effect on all odonates; few odonates emerged in CF fields (Table 2). The main difference in management between CF and EF fields is insecticide use. Although in CF, insecticides are applied both in the nursery boxes before transplanting and in the paddy fields at flowering, odonate nymphs are exposed only, or primarily, to the earlier insecticide. Therefore, nursery-box application clearly caused the decrease in the abundance of odonates in the CF fields. We did not directly measure the insecticide concentrations in paddy water, but this result is highly consistent with those of studies of the relationship between insecticide concentrations in water and the emergence rate of Sympetrum dragonflies (Jinguji et al. 2009, 2010, 2013). As coenagrionid and lestid (I. peregrinus) eggs hatch a few weeks after transplantation, insecticides applied in CF could continue to harm damselflies even after transplantation, through possible persistence or indirect effects of reduced prey availability (e.g., Hayasaka et al. 2013). One recent study suggested that the direct effects of fipronil on the damselfly species Ischnura senegalensis are high even at low concentrations (Sugita et al. 2018), so the former possibility is also conceivable.
Forest area had no interactive effect with agricultural practice on any of the odonate species, and its effect varied among the species. There was no effect of forest area on the nymphal exuviae of Sympetrum or on the total number of damselflies. Because these numbers each reflected the abundance of two or more species, any effect of the area of forest may have been offset by differences in ecological properties among the species. Additionally, landscape elements other than forest may be more important for damselflies than for Sympetrum dragonflies, because the former prefer stable water bodies such as high-quality and nearby ponds as alternative habitat (Sugimura et al. 1999; Ozono et al. 2012; Raebel et al. 2012b); coenagrionids, in particular, require these water bodies for overwintering. We could not evaluate the effect of surrounding water habitat because there were few water bodies (other than a river) in the study area. Raebel et al. (2012a) reported that the species richness of dragonflies was influenced by landscape variables at a spatial scale of 1600 m; therefore, to detect landscape effects on odonates, we would need to conduct field surveys across a wider area than our survey sites (Fig. 1).
The surrounding forest area had different effects on adults of the two Sympetrum dragonfly species. The abundance of S. frequens was not influenced by forest area in the averaging model. This species seems to be affected by environmental factors at larger spatial scale because of its high mobility (Ueda 1998a). In contrast, although adults of S. infuscatum use nearby forest edges as habitat (e.g., Ueda 1998b), our results suggest that increased forest area within 200 m decreased the abundance of S. infuscatum in paddy fields. This result might mean that this species prefers open paddy fields for oviposition rather than fields enclosed by forests. Therefore, to increase the abundance of S. infuscatum, it would be more effective to conduct EF farming in paddy fields away from forests. This result is supported by previous evidence of the negative effects of woodlands on odonate assemblages (Pither and Taylor 1998; Raebel et al. 2012a). In addition, the abundance of M. selenion was not influenced by forest area in the averaging model. This species inhabits short vegetation in wetlands on plains and in hilly areas (Sugimura et al. 1999), so forests may not be so important.
Our results suggest that EF farming is generally effective for increasing the abundance of dragonflies and damselflies (e.g., an 87 × increase in the abundance of nymphal exuviae of Sympetrum and a 9.9 × increase in the adult damselflies, Fig. 2). However, its effectiveness varies with odonate species and the surrounding landscape, likely reflecting differences in the life history features among species. Therefore, it is important to understand the life history features of species to be conserved. Because the decrease in odonate populations can be attributed mainly to changes in the local agricultural environment, such as loss of oviposition sites and reduced nymphal survival due to early drainage of paddy water and insecticide application (Aoda et al. 2013), implementing EF farming practices at the regional level should mitigate population declines at that same level. In addition, dragonfly nymphs are important predators of mosquito larvae (Urabe et al. 1990). Anopheles sinensis, a vector of tertian malaria, which was epidemic throughout Japan until the 1950s, and Culex tritaeniorhynchus, a vector of Japanese encephalitis, are distributed throughout Japan and breed mainly in paddy fields (Umenai et al. 1985). Although malaria is not prevalent now in Japan, and the occurrence of Japanese encephalitis is decreasing, anticipated temperature rise and altered rainfall and increasing immigration of infected travelers and immigrants may increase the occurrence, distribution, and intensity of disease transmission (e.g., Tada et al. 2008; Miller et al. 2012; Caminade et al. 2014). On the other hand, mosquito populations have probably also been reduced in rice paddy fields by the insecticides applied to control insect pests. Therefore, the reduced use of insecticides in EF farming practice may increase vector mosquito populations. The maintenance of paddy odonate communities would help to prevent the spread of these infectious diseases.
References
Amano T, Kusumoto Y, Okamura H et al (2011) A macro scale perspective on within farm management: how climate and topography alter the effect of farming practices. Ecol Lett 14:1263–1272
Aoda T, Katano K, Toyama K, Jinguji H (2013) Assessment of paddy environment using emergence husks of red-dragonflies with civic participation. Bull Facul Agric Niigata Univ 65:131–135. (in Japanese)
Baba YG, Kusumoto Y, Tanaka K (2018) Effects of agricultural practice and fine-scale landscape factors on spiders and a pest insect in Japanese rice paddy ecosystems. Biocontrol 63:265–275
Bambaradeniya CN, Amerasinghe FP (2004) Biodiversity associated with the rice field agroecosystem in Asian countries: a brief review. 63. IWMI, Colombo
Barton K (2018) MuMIn: multi-model inference, R package version 1.42.1. http://r-forge.r-project.org/projects/mumin/
Batáry P, Báldi A, Kleijn D, Tscharntke T (2010) Landscape-moderated biodiversity effects of agri-environmental management: a meta-analysis. Proc Roy Soc London B, rspb20101923
Bates Dl, Maechler M, Bolker B et al (2018) lme4: linear mixed-effects models using Eigen and S4. R package Version 1–19. https://cran.r-project.org/web/packages/lme4/
Burnham KP, Anderson DR (2002) Model selection and multimodel inference: a practical information—theoretic approach. Springer, New York
Caminade C, Kovats S, Rocklov J et al (2014) Impact of climate change on global malaria distribution. Proc Nat Acad Sci 111:3286–3291
Concepción ED, Díaz M, Kleijn D et al (2012) Interactive effects of landscape context constrain the effectiveness of local agri-environmental management. J Appl Ecol 49:695–705
Futahashi R (2012) Recent decline of red dragonflies in Toyama Prefecture. Insect Nature 47:10–15. (In Japanese)
Hamasaki K, Yamanaka T, Tanaka K et al (2009) Relative importance of within-habitat environment, land use and spatial autocorrelations for determining odonate assemblages in rural reservoir ponds in Japan. Ecol Res 24:597–605
Hamasaki K, Yamanaka T, Tanaka K et al (2011) Environmental characteristics accounting for Odonate assemblages in rural reservoir ponds in Japan. JARQ 45:187–196
Hayasaka D, Suzuki K, Nomura T et al (2013) Comparison of acute toxicity of two neonicotinoid insecticides, imidacloprid and clothianidin, to five cladoceran species. J Pestic Sci 38:44–47
Hole DG, Perkins AJ, Wilson JD et al (2005) Does organic farming benefit biodiversity? Biol Conserv 122:113–130
Jinguji H, Ueda T, Goka K et al (2009) Effects of Imidacroprid and Fipronil insecticide application on the larvae and adults of Sympetrum frequens (Libellulidae: Odonata). Trans Jpn Soc Irrig Drain Rural Eng 77:35–41
Jinguji H, Ueda T, Tsunoda M et al (2010) Effects of fipronil insecticide application on Sympetrum sp. Larvae and adults in experimental rice paddy field. Trans Jap Soc Irrig Drain Rural Eng 78:219–226
Jinguji H, Thuyet DQ, Uéda T et al (2013) Effect of imidacloprid and fipronil pesticide application on Sympetrum infuscatum (Libellulidae: Odonata) larvae and adults. Paddy Water Environ 11:277–284
Jinguji H, Ohtsu K, Ueda T et al (2018) Effects of short-term, sublethal fipronil and its metabolite on dragonfly feeding activity. PloS ONE 13:e0200299
Katayama N, Baba YG, Kusumoto Y, Tanaka K (2015) A review of post-war changes in rice farming and biodiversity in Japan. Agric Syst 132:73–84
Kato N, Yoshio M, Kobayashi R et al (2010) Differential responses of two anuran species breeding in rice fields to landscape composition and spatial scale. Wetlands 30:1171–1179
Kiritani K (2000) Integrated biodiversity management in paddy fields: shift of paradigm from IPM toward IBM. Integr Pest Manage Rev 5:175–183
Kleijn D, Sutherland WJ (2003) How effective are European agri-environment schemes in conserving and promoting biodiversity? J Appl Ecol 40:947–969
Kleijn D, Baquero RA, Clough Y et al (2006) Mixed biodiversity benefits of agri-environment schemes in five European countries. Ecol Lett 9:243–254
Lawler SP (2001) Rice fields as temporary wetlands: a review. Israel J Zool 47:513–528
Merckx T, Feber RE, Riordan P et al (2009) Optimizing the biodiversity gain from agri-environment schemes. Agric Ecosyst Environ 130:177–182
Miller RH, Masuoka P, Klein TA et al (2012) Ecological niche modeling to estimate the distribution of Japanese encephalitis virus in Asia. PLoS Negl Trop Dis 6:e1678
Ministry of the Environment Japan (2018) Red List 2018. http://www.env.go.jp/nature/kisho/hozen/redlist/index.html Accessed 20 July, 2018
Nakanishi K, Yokomizo H, Hayashi TI (2018) Were the sharp declines of dragonfly populations in the 1990s in Japan caused by fipronil and imidacloprid? an analysis of Hill’s causality for the case of Sympetrum frequens. Environ Sci Pollu Res 25:35352–35364
Oyama J, Kidokoro T (2003) Effects of insecticide application in rice cultivation on densities of spiders, larvae of Sympetrum and Chironomidae species in paddy field. Annu Rep Plant Prot N Jpn 54:123–125. (In Japanese)
Ozono A, Kawashima I, Futahashi R (2012) Dragonflies of Japan. Bunichi-Sogo Syuppan, Tokyo
Pither J, Taylor PD (1998) An experimental assessment of landscape connectivity. Oikos 83:166–174
R Development Core Team (2014) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, http://www.R-project.org.
Raebel EM, Merckx T, Riordan P et al (2010) The dragonfly delusion: why it is essential to sample exuviae to avoid biased surveys. J Insect Conserv 14:523–533
Raebel EM, Merckx T, Feber RE et al (2012a) Multi-scale effects of farmland management on dragonfly and damselfly assemblages of farmland ponds. Agric Ecosyst Environ 161:80–87
Raebel EM, Merckx T, Feber RE et al (2012b) Identifying high-quality pond habitats for Odonata in lowland England: implications for agri-environment schemes. Insect Conserv Div 5:422–432
Sugimura M, Ishida S, Kojima K, Ishida K, Aoki T (1999) Dragonflies of the Japanese Archipelago in color (in Japanese). Hokkaido University Press, Sapporo
Sugita N, Agemori H, Goka K (2018) Acute toxicity of neonicotinoids and some insecticides to first instar nymphs of a non-target damselfly, Ischnura senegalensis (Odonata: Coenagrionidae), in Japanese paddy fields. Appl Entomol Zool 53:519–524
Tada Y, Okabe N, Kimura M (2008) Travelers’ risk of malaria by destination country: a study from Japan. Travel Med Infect Dis 6:368–372
Takada MB, Takagi S, Iwabuchi S et al (2014) Comparison of generalist predators in winter-flooded and conventionally managed rice paddies and identification of their limiting factors. SpringerPlus 3:418
Tscharntke T, Klein AM, Kruess A (2005) Landscape perspectives on agricultural intensification and biodiversity–ecosystem service management. Ecol Lett 8:857–874
Tsutsui MH, Tanaka K, Baba YG et al (2016) Spatio-temporal dynamics of generalist predators (Tetragnatha spider) in environmentally friendly paddy fields. Appl Entomol Zool 51:631–640
Tuck SL, Winqvist C, Mota F et al (2014) Land-use intensity and the effects of organic farming on biodiversity: a hierarchical meta-analysis. J Appl Ecol 51:746–755
Ueda T (1998a) Dragonfly communities in paddy fields. In: Ezaki Y, Tanaka T (eds) Conservation of biological communities in rivers, ponds and paddy fields (in Japanese). Asakura Book Co., Tokyo, pp 93–110
Ueda T (1998b) Dragonfly communities in ponds. In: Ezaki Y, Tanaka T (eds) Conservation of biological communities in rivers, ponds and paddy fields (in Japanese). Asakura Book Co., Tokyo, pp 17–33
Ueda T, Jinguji H (2013) The ecological impact of the insecticides fipronil and imidacloprid on Sympetrum frequens in Japan. Tombo 55:1–12
Umenai T, Krzysko R, Bektimirov TA et al (1985) Japanese encephalitis: current worldwide status. Bull World Health Organ 63:625
Urabe K, Ikemoto T, Takei S (1990) Studies on Sympetrum frequens (Odonata: Libellulidae) nymphs as natural enemies of the mosquito larvae Anopheles sinensis in rice fields. 4 Prey–predator relationship in the rice field areas. Appl Entomol Zool 41:265–272
Uruma H, Kobayashi R, Nishijima Set al (2012) Effectiveness of conservation-oriented agricultural practices on amphibians inhabiting Sado Island, Japan, with a consideration of spatial structure. Jpn J Conserv Ecol 17:155–164
Usio N, Miyashita T (eds) (2015) Social-ecological restoration in paddy-dominated landscapes. Springer Japan
Washitani I (2007) Restoration of biologically-diverse floodplain wetlands including paddy fields. Glob Environ Res 11:135–140
Willer H, Lernoud J, Home R (2011) The world of organic agriculture 2011: Summary. The world of organic agriculture. Stat Emerg Trends 2011, 26–32
Acknowledgements
We thank Shu-ichi Sugiyama, Toshihiro Yoshida, Yoshiharu Yoshida, and Tamotsu Yoshida for allowing us to survey their paddy fields, and the late Katsuo Abe for helping with the field survey. This work was supported by grants from the Ministry of Agriculture, Forestry and Fisheries of Japan through the “Development of agricultural pest control system by the effective use of indigenous natural enemies” project.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflicts of interest.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Baba, Y.G., Kusumoto, Y. & Tanaka, K. Positive effect of environmentally friendly farming on paddy field odonate assemblages at a small landscape scale. J Insect Conserv 23, 467–474 (2019). https://doi.org/10.1007/s10841-019-00132-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10841-019-00132-2