
Abstract
We examined the effects of environmentally friendly (EF) farming and landscape factors on the abundances of major spider guilds and a rice pest, the small brown planthopper (SBPH), Laodelphax striatellus (Fallén), in a Japanese rice paddy ecosystem. The abundances of all spider guilds increased by EF farming, whereas different spider guilds showed contrasting responses to the size of the forest within 200 m of the fields. The abundances of ground spiders and horizontal web weavers increased with increasing forest area, unlike small ground spiders, whose abundance decreased. The abundances of SBPH nymphs and adults decreased by EF farming but responded differently to landscape: nymphs decreased with increasing forest area, but adults did not. Further analysis indicated a potential negative relationship between web-weaving spiders and SBPH nymphs. Our findings suggest that the size of the forest adjacent to fields is an important determinant of spider guild composition and pest abundance.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Pest insect control by generalist predators such as spiders and carabid beetles is an important ecosystem service in agro-ecosystems (Marc and Canard 1997; Riechert and Lockley 1984; Sunderland and Samu 2000). The importance of such pest control is increasing because environmentally friendly (EF) farming, in which the use of agrochemicals and chemical fertilizers is reduced, is being promoted around the world (Willer et al. 2010). Therefore, exploring factors affecting predator communities and their pest control services in the field is an important issue in conservation biological control (Tscharntke et al. 2007).
Both agricultural practices and environmental factors beyond the field scale are important determinants of predator communities and their pest control services (Bianchi et al. 2006; Tscharntke et al. 2007). In terms of agricultural practices, the functions of generalist predators can be enhanced by EF farming because predator communities within EF-farmed fields are generally more diverse and complex than those in conventionally farmed fields (Bengtsson et al. 2005; Tuck et al. 2014). Furthermore, the surrounding landscape influences the abundance and diversity of predators by providing alternative habitats for field organisms, including both prey and predators, when arable fields are unsuitable (Bianchi et al. 2006; Schmidt et al. 2008; Tscharntke et al. 2007). Because responses to landscapes likely differ among predator groups with different habitat uses, the guild and taxonomic compositions of predators potentially vary depending on the surrounding landscape, and this in turn has different consequences for pest control services. Many studies have explored how local and landscape factors determine the diversity and abundance of generalist predators in upland farmlands, such as in wheat fields and meadows (reviewed by Tscharntke et al. 2007), and the number of studies on the relationship between predator abundance or diversity and their pest control services is rapidly increasing. However, relatively few studies have integrated a series of effects from the landscape to pest control service via changes in the natural enemy community (but see Birkhofer et al. 2016; Picchi et al. 2016). Furthermore, these landscape-scale studies have been biased towards European upland fields; there have been very few studies of rice paddy ecosystems, which constitute the major type of farmland in Asia (Betz and Tscharntke 2017; Katayama et al. 2015; Tsutsui et al. 2016). The composition of natural enemies and pests and the surrounding landscape completely differ between European upland fields and Asian paddy fields, so studies on paddy ecosystems are expected to provide novel and useful knowledge for conservation biological control in Asian countries.
Here, we attempted to elucidate the insect pest control services provided by generalist predators and associated with local and landscape factors in Japanese rice paddy ecosystems. We focused on the small brown planthopper (SBPH), Laodelphax striatellus (Fallén 1826), one of the most important pest insects of rice plants in Asia (Cheng 2009). Although this species usually does not directly damage rice plants by feeding on them, it can transmit the rice stripe virus (RSV) to rice plants, which then develop rice stripe disease (Shinkai 1962). Recently, the density of SBPH has gradually been increasing (Shiba 2016), and the frequency of individuals carrying RSV has increased in the western (Matsumura and Otuka 2009) and eastern (Ogura et al. 2014; Okabe and Sugiyama 2016; Tsukahara 2016) regions of Japan. Additionally, the susceptibility of SBPH to certain kinds of insecticide (e.g., fipronil) has recently declined (Sanada-Morimura et al. 2011; Tsukahara 2016), so an effective pest control technique to replace insecticide application needs to be developed.
We focused on spiders, which are major generalist predators in rice paddy ecosystems (Tanaka 2016) and act as agents that regulate SBPH. Although many spider species occur in rice paddy fields, the following four spider guilds, which have different prey capture behaviors, are representative: horizontal web weavers (Tetragnatha), cursorial ground spiders (Lycosidae), small ground sheet-web weavers (Erigoninae), and small semi-wandering spiders (Pachygnatha). Although, to our knowledge, the suppression effects of spiders on the SBPH population have not been explored, spiders can be considered potential enemies, because some are major biocontrol agents for hemipteran pest insects (Kiritani et al. 1972; Kobayashi et al. 2011; Takada et al. 2012). Spider abundance and guild composition are spatially variable and are influenced by agricultural practices (Takada et al. 2014; Tanaka 2016) and surrounding landscapes (Amano et al. 2011; Baba and Tanaka 2016b; Tsutsui et al. 2016). These changes in spider abundance and composition influence pest populations. Therefore, to use the pest control function of spiders, we need to understand how both agricultural practices and landscape factors influence spider communities and their pest control functions.
Here, we evaluated local and landscape factors influencing the abundance of spiders and their pest control services in a rice paddy ecosystem. We first investigated the abundances of four major guilds of spiders and SBPH in conventional and EF-farmed paddy fields located across heterogeneous landscapes, and we analyzed the effects of agricultural practices and landscape factors on these abundances. Second, to detect the impacts of spiders on the pest population, we clarified the relationships between spiders and pest insect abundances, taking into account other confounding factors with generalized linear models. We hypothesized that, if the surrounding landscape were to change, the guild composition of spiders would also change to reflect the spiders’ divergent responses to landscape factors, and this would probably result in different effects on pest control services.
Materials and methods
Study area and sampling design
The field survey was conducted in 2012 and 2013 at Shioya-cho, Tochigi Prefecture, in the northern part of the Kanto region of Japan (Fig. 1). As study plots, ten EF-farmed paddy fields were surveyed in both years, and seven and ten conventional paddies were surveyed in 2012 and 2013, respectively. The size of paddy field is ranging from 1366 to 8586 m2. These paddy fields are located along a gradient from simple arable fields to forest-rich landscapes, thus allowing us to test the influences of fine-scale landscape differences on organisms. The major difference between EF and conventional farming practices was insecticide use (Table 1). In the case of the EF fields, no insecticides had been used since 2010, whereas neonicotinoid, phenylpyrazole, and/or synthetic pyrethroid insecticides had been applied to the rice-seedling nursery boxes and paddy fields in the conventional farming practices. In addition, the fungicides and herbicides applied to the EF fields had lower toxicity to organisms than those applied to the conventional fields.

Study area in Shioya-cho, Tochigi Prefecture, Japan. White and black circles indicate environmentally friendly farmed and conventionally farmed paddy fields, respectively. ✩ paddy fields were surveyed in 2013 only
Target organisms
Four major spider guilds with different prey-capture behaviors, namely Tetragnatha, lysosids, erigonines, and Pachygnatha, were investigated as predators. Tetragnatha spiders (Tetragnathidae) are among the predominant generalist predators in Asian rice paddy ecosystems (Okuma et al. 1978; Pei and Nakamura, 2012). These spiders prefer damp habitats and build horizontal orb-webs among rice plants. In our study fields, five Tetragnatha species, T. praedonia L. Koch 1878, T. maxillosa Thorell 1895, T. caudicula (Karsch 1879), T. extensa (Linnaeus 1758), and T. vermiformis Emerton 1884, have been observed (Baba and Tanaka 2016a). Lycosids are cursorial ground spiders and are effective biological control agents for the green rice leafhopper, Nephotettix cincticeps Uhler 1896 (Kiritani et al. 1972). The two dominant species, Pardosa pseudoannulata (Bösenberg and Strand 1906) and Pirata subpiraticus (Bösenberg and Strand 1906) (Baba and Tanaka 2016a), feed on prey insects above the water surface or on the bases of rice plants. Erigonine (Linyphiidae) communities are dominated by three species (Erigone prominens Bösenberg and Strand 1906, Gnathonarium exsiccatum (Bösenberg and Strand 1906) and Ummeliata insecticeps (Bösenberg and Strand 1906)) of small ground spiders. They construct small sheet webs in the lower parts of rice plants, but they often wander and capture prey directly without a web (Nakasuji et al. 1973). Pachygnatha spiders in our study system are two main species (P. tenera Karsch 1879 and P. quadrimaculata (Bösenberg and Strand 1906)) and are also categorized as small cursorial ground spiders despite belonging to the Tetragnathidae (most of which construct orb-webs), because they are likely to forsake web construction. However, their detailed life history remains unclear.
The SBPH as a pest insect is dimorphic, with fully winged (macropterous) and truncated winged (brachypterous) forms. The fully winged forms are potential migrants and are responsible for colonizing new fields. In the Kanto region, this species is thought to produce at least three generations during the rice growing season (Okabe and Sugiyama 2016). After immigration to rice fields from the surrounding grasslands and wheat fields in late May, they produce the next generation, in which most of the females develop as a brachypterous form and the males as a macropterous form. Adults usually mate on the day of emergence, and females start laying eggs the following day. Eggs hatch in about 6–9 days, and the nymphs undergo five instars to develop into adults. The second-generation adults occur from late June to mid-July (during the rice tillering to heading stages), and the third-generation adults occur starting in late July (the heading stage).
Sampling protocol
We sampled spiders and pest insects on 18 and 19 July and 8 and 9 August 2012, and 24 and 25 July and 7 and 8 August 2013, when spiders and SBPH were most abundant. To collect SBPH and spiders with different prey-capture behaviors, two sampling methods were adopted. Tetragnatha spiders and SBPH were collected by sweep-netting. The rice plants were swept with an insect net (36 cm internal diameter) 20 times while the researcher walked along an arbitrary transect within the paddy field. This procedure was conducted in two locations within each field. All arthropods collected were killed with a household insecticide and preserved in polypropylene vials filled with 75% ethanol. The target spiders and pest insects were then sorted and counted in the laboratory. In the second sampling procedure, the other spiders (lycosid, erigonine, and Pachygnatha spiders), which live near the paddy water or ground surface, were visually counted by pushing aside the rice plants. Five hills of rice plants were surveyed per plot, and four plots were investigated per rice paddy field (for a total of 20 rice hills per field).
Landscape metrics
We created polygon data on land uses in the study area (e.g., arable field, open water, forest, road, buildings, and abandoned fields) on the basis of a national basic map (1:10,000) and a high-resolution aerial chart by using the remote-sensing software ERDAS IMAGINE (Hexagon Geospatial, GA, USA) and ArcGIS 10.1 (Esri, Redlands, CA, USA). Landscape metrics were measured by using ArcView 3.10 GIS software (Esri, Redlands, CA, USA). The areas of land dedicated to each use were evaluated within 50, 100, and 200 m buffers from the periphery of the study fields. We used the sizes of the areas of land covered by forests or buildings in our analyses as potential factors influencing the abundances of the target organisms. The other land uses were not considered because the area of arable field showed a strong negative correlation with the forested area at two spatial scales (arable field vs. forest: 50 m, r = − 0.38, P = 0.10; 100 m, r = − 0.64, P = 0.002; 200 m, r = − 0.84, P < 0.001), and abandoned fields, open water, and road accounted for only small fractions of the total area of buffers.
Statistical analysis
To analyze the effects of local field management (environmentally friendly vs. conventional) and landscape factors (size of the areas of land covered by forests or buildings) on the abundances of the different spider guilds and pest insects, generalized linear models (GLMs) with a negative-binomial distribution and a log-link function were applied, including the number of surveys (2–4) as an offset term. The abundance data on spiders for different years and months were combined, as were those for pest insects. Possible two-way interactions of the explanatory variables were not included because of the relatively small sample sizes. If the opposite response of abundance to the explanatory variables was found between spiders and the pest insect, we added spider abundance to the original model explaining pest abundance to test the potential pest-suppression effect of spiders. The best model explaining the variability of the abundance was selected on the basis of the lowest akaike information criteria (AIC) value. We adopted the following two procedures to detect the best model. First, to specify the effective spatial scale of the landscape, we applied the statistical model to all possible combinations of explanatory variables for each buffer size and compared model performance by using the AIC values. The lowest AIC value of the model in each buffer size were compared, and the buffer size showing the lowest value was regarded as an effective spatial scale. Next, to identify explanatory variables influencing the abundance of organisms, we created a model for all explanatory variable combinations on the effective spatial scale and calculated the difference from the lowest AIC value (ΔAIC = AIC—lowest AIC). All models showing ΔAIC < 2 are considered as valid alternative models, so these models were shown in the results. We determined influential variables based on the z values of their coefficients (estimate/SE). Variables with z values > 1.96 were considered to be influential (Burnham and Anderson 2002). The statistical significance level of each coefficient was also assessed by the Wald test. These procedures were conducted by using the MASS packages of R 2.3.4 software (R Core Team 2014). In general, the geographical coordinates of each field should be taken into account to evaluate the effect of spatial autocorrelation, but it is difficult to distinguish it from landscape effect due to the relative small sample size and the spatial proximity. So we decide not to consider it here as well as in the previous study (Tsutsui et al. 2016).
Results
A total of 2466 Tetragnatha spider individuals were collected by net-sweeping, and 2257 other spiders were counted visually (Lycosidae, n = 1187; Erigoninae, n = 772; and Pachygnatha, n = 298). In total, 14,229 nymph and 5332 adult SBPH were collected.
Spiders
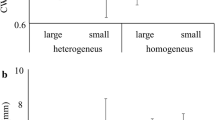
The model selection results showed that the abundances of the four spider guilds were influenced by local agricultural practices and landscape factors within 200 m of the periphery of the rice paddy fields. Environmentally friendly farming positively influenced the abundances of all of the spider guilds. However, landscape factors had contrasting effects (Table 2). Increasing forested area within 200 m had a positive effect on both Tetragnatha and lycosid spiders, but a negative effect on erigonine and Pachygnatha spiders (Table 2, Fig. 2).

Relationships between the size of the forested area within 200 m and the abundance of spiders (Tetragnatha, Lycosidae, Erigoninae, and Pachygnatha). Solid and dashed lines represent the fitting lines of environmentally friendly (EF) farmed and conventionally farmed paddy fields, respectively. Abundance indicates the number of spiders collected or counted in each field per survey
SBPH and its relationship to spiders
Abundances of SBPH nymphs and adults were influenced by different local and landscape factors. The best selected model showed that the nymphs were negatively influenced by EF farming and by increasing forest area within 200 m of the periphery of the rice field, whereas the adults were negatively influenced only by EF farming (Mean ± SE, EF farming 60.83 ± 7.73; conv. farming 90.18 ± 12.76) (Table 3). The results indicated a negative effect of Tetragnatha and lycosid abundance on SBPH nymphs. Therefore, we added spider abundances as explanatory variables to the original statistical models. In the model selection, a greater abundance of Tetragnatha spiders consistently had a negative effect on SBPH nymphs in the best and two competitive models, although nine competitive models (ΔAIC < 2) were found (Table 3, Fig. 3). The same analysis was also applied to the SBPH adults, but the best model did not include the effect of spider abundances, and there were seven competitive models (ΔAIC < 2), indicating no consistent effect of spiders.

Relationship between the abundance of SBPH nymphs and Tetragnatha spiders in environmentally friendly (EF) farmed and conventional farmed paddy fields proposed by the best explanatory model including only the effect of Tetragnatha spider abundance. Solid line represents the fitting line of GLM
Discussion
Our results showed that both agricultural practice and fine-scale landscape factors influenced the abundances of spiders with different foraging behaviors and the target pest insect. Different spider guilds exhibited different responses to increases in the amount of forested area within 200 m of the rice field, indicating that the guild composition of spiders potentially varied depending on the surrounding landscape. Furthermore, the abundance of Tetragnatha spiders had a negative influence on the abundance of SBPH nymphs, suggesting that the surrounding landscape indirectly influenced the pest control service mediated by spiders. Below we discuss the possible mechanisms of the landscape and EF farming effects on spiders and the pest insect.
Environmentally friendly farming (i.e., non-use of insecticides) had a positive effect on the abundance of all spider guilds, suggesting that the practice of applying insecticides twice a season decreased spider density. The mechanism by which insecticides decrease the abundance of spiders is thought to differ depending on the type of insecticide. Insecticides applied to rice fields in August—especially etofenprox (Tanaka et al. 2000)—may directly decrease spider abundance, whereas early insecticide application to rice-seedling nursery boxes may indirectly decrease it, because spiders are not directly exposed to the insecticide. Insecticides applied to nursery boxes, such as fipronil and imidacloprid, have negative effects on aquatic insect communities (Hayasaka et al. 2012). Consequently, the early-season (nursery-box) insecticides probably reduced spider abundances by decreasing the amount of available insect prey emerging from the paddy water (e.g., chironomid midges: Takada et al. 2014; Tsutsui et al. 2016).
Unlike the effect of agricultural practice, landscape effect varied among the spider guilds. An increase in the amount of forest area within the 200-m buffer had a positive effect on Tetragnatha and lycosid spiders but a negative one on erigonine and Pachygnatha spiders. Because the surrounding landscape serves as alternative habitat for spiders or a source habitat for their potential insect prey (Miyashita et al. 2012; Tsutsui et al. 2016), these opposite responses may reflect differences in habitat use or prey types among spider guilds. Because Tetragnatha and lycosid spiders do not typically inhabit forest (Baba, unpublished data), they may benefit from forest through its food supply. This possibility is strongly supported by a previous study conducted in the same area (Tsutsui et al. 2016), which revealed that the availability of chironomid midges—a major part of the diet of Tetragnatha—increased as the amount of surrounding forest area increased. On the other hand, there is an alternative possibility that spiders accumulate in paddy fields with a large surrounding forest area because there are no suitable habitats around them. Further investigation to confirm the ecological process is necessary.
In contrast, the negative relationship between the abundances of erigonine and Pachygnatha spiders and increasing forest area is likely related to the spiders’ habitat use. Some erigonine spiders are abundant in arable fields and not in the surrounding natural habitat (Schmidt and Tscharntke 2005), and they prefer disturbed habitats (Samu and Szinetár 2002). Thus, forest may be an unsuitable habitat for these small spiders, and an increase in forest area may fragment their preferred habitats at the landscape level. In addition, increased forest area may provide suitable habitat for their competitors or predators. For example, the abundance of lycosid spiders increases as the surrounding forest area increases and the abundance of small spiders decreases because the larger spiders prey on the smaller ones (e.g. Denno et al. 2004). To confirm the ecological processes related to the landscape effect, it is necessary to investigate the fundamental ecology of these spiders, such as habitat use, prey types, and inter-specific interactions.
Environmentally friendly farming negatively affected the abundance of SBPH nymphs and adults. An increase in the area of surrounding forest also negatively affected the nymphs, but landscape factors had no effect on the adults. The negative effect of this increase in forest area on the nymphs suggest two possibilities. One is the possibility that the potential habitat of pests (e.g. grassland) will decrease as surrounding forest increases. The other possibility is that the decrease in pests is attributable to the abundance of Tetragnatha spiders, which increased with increasing forest area. This possibility is supported by the result that Tetragnatha abundance had a negative effect on SBPH nymph abundance (Fig. 3). However, since there were many competitive models showing ΔAIC < 2 in the results of model selection, further verification is needed. On the other hand, the decrease in the number of adult SBPH in EF farming fields could not be explained by the increase in spider abundances, although adult SBPH are also likely to be captured by various spider guilds. This may be a result of a weakness in statistical power or a synergetic pest suppression effect by predators that is not simply equal to the sum of predator abundances (e.g. Losey and Denno 1998). Because the statistical results did not necessarily suggest a causal relationship of pest control by spiders and direct interaction between spiders and SBPH has never been investigated, there is a need to directly evaluate the prey-capture rate in spider webs (Birkhofer et al. 2016; Mader et al. 2016) or to perform a DNA-based diet-content analysis of spiders (Birkhofer et al. 2017; Kobayashi et al. 2011) in future studies.
Our results showed that landscape factors within narrow areas (within 200 m of the paddy field) determined spider and pest insect abundances. However, in this analysis, since only the landscape elements ranging from 50 to 200 m were analyzed, there is a possibility that spatial scale factor larger than 200 m may be important. Furthermore, since the landscape has a nested structure, smaller landscape elements included in the effective spatial scale may also have an important role. Therefore, to specify deterministic factor for spiders and pest abundances, it is also necessary to focus on a larger spatial scale and evaluate more detailed landscape structure. It is noteworthy that more pest insects occurred in conventionally farmed fields than in EF-farmed fields. This result may indicate a pest resurgence caused by a decline in the natural enemy population as a result of the use of insecticides (e.g. Hardin et al. 1995; Settle et al. 1996). Although the damage to rice caused by SBPH is currently limited, the number of individuals with RSV is increasing rapidly in the eastern region of Japan (Shiba 2016), and SBPH has become less susceptible to certain kinds of insecticide (Sanada-Morimura et al. 2011; Tsukahara 2016). In future it will be necessary to develop an effective technique to suppress SBPH without using insecticides, and EF farming practices seem to be an effective way to enhance the function of SBPH’s natural enemies.
Here, we were able to elucidate the responses of spider community and pest insect to landscape and agricultural practices in rice paddy ecosystems. Because the responses of spiders to landscape factors are diverse, the dominant spider species (or guilds) vary depending on landscape structure. For example, small Erigoninae and Pachygnatha spiders, which are negatively influenced by the presence of forest, are abundant in paddy-dominated landscapes and may play an important pest-suppression role there. As a future study, we need to test the pest control function of spider by considering a positive interaction among spiders in which different spider guilds cooperatively suppress pest insects (e.g. Takada et al. 2013). This synergetic pest suppression among multiple predators may be important, especially in rice paddy ecosystems, because predator diversity in rice paddy fields appears to be high (Kiritani 2000; Usio and Miyashita 2015). Therefore, to evaluate the comprehensive ecosystem function of spiders we need to conduct further investigations to clarify the individual and interactive pest-suppression functions of a broad range of guilds of spiders under various landscape contexts.
References
Amano T, Kusumoto Y, Okamura H, Baba YG, Hamasaki K, Tanaka K, Yamamoto S (2011) A macro-scale perspective on within-farm management: how climate and topography alter the effect of farming practices. Ecol Lett 14:1263–1272
Baba YG, Tanaka K (2016a) Environmentally friendly farming and multi-scale environmental factors influence generalist predator community in rice paddy ecosystems of Japan. In: Yagi K, Kuo CG (eds) The challenges of agro-environmental research in Monsson Asia, NIAES Series No.6. pp. 171–179
Baba YG, Tanaka K (2016b) Factors affecting abundance and species composition of generalist predators (Tetragnatha spiders) in agricultural ditches adjacent to rice paddy fields. Biol Control 103:147–153
Bengtsson J, Ahnström J, Weibull AC (2005) The effects of organic agriculture on biodiversity and abundance: a meta-analysis. J Appl Ecol 42:261–269
Betz L, Tscharntke T (2017) Enhancing spider families and spider webs in Indian rice fields for conservation biological control, considering local and landscape management. J Insect Conserv 21(3):495–508
Bianchi FJJA, Booij CJH, Tscharntke T (2006) Sustainable pest regulation in agricultural landscapes: a review on landscape composition, biodiversity and natural pest control. P Roy Soc B-Biol Sci 273:1715–1727
Birkhofer K, Arvidsson F, Ehlers D, Mader VL, Bengtsson J, Smith HG (2016) Organic farming affects the biological control of hemipteran pests and yields in spring barley independent of landscape complexity. Landscape Ecol 31:567–579
Birkhofer K, Bylund H, Dalin P, Ferlian O, Gagic V, Hambäck PA, Klapwijk M, Mestre L, Roubinet E, Schroeder M, Stenberg JA, Porcel M, Björkman C, Jonsson M (2017) Methods to identify the prey of invertebrate predators in terrestrial field studies. Ecol Evol 7:1942–1953
Burnham KP, Anderson DR (2002) Model selection and multimodel inference: a practical information-theoretic approach. Springer, New York
Cheng JA (2009) Rice planthopper problems and relevant causes in China. In: Heong KL, Hardy B (eds) Planthoppers: new threats to the sustainability of intensive rice production systems in Asia. International Rice Research Institute, Los Baños, pp. 157–178
Denno RF, Mitter MS, Langellotto GA, Gratton C, Finke DL (2004) Interactions between a hunting spider and a web-builder: consequences of intraguild predation and cannibalism for prey suppression. Ecol Entomol 29:566–577
Hardin MR, Benrey B, Coll M, Lamp WO, Roderick GK, Barbosa P (1995) Arthropod pest resurgence: an overview of potential mechanisms. Crop Protec 14:3–18
Hayasaka D, Korenaga T, Suzuki K, Saito F, Sánchez-Bayo F, Goka K (2012) Cumulative ecological impacts of two successive annual treatments of imidacloprid and fipronil on aquatic communities of paddy mesocosms. Ecotox Environ Safe 80:355–362
Katayama N, Baba YG, Kusumoto Y, Tanaka K (2015) A review of post-war changes in rice farming and biodiversity in Japan. Agric Syst 132:73–84
Kiritani K (2000) Integrated biodiversity management in paddy fields: shift of paradigm from IPM toward IBM. Integ Pest Manag Rev 5:175–183
Kiritani K, Kawahara S, Sasaba T, Nakasuji F (1972) Quantitative evaluation of predation by spiders on the green rice leafhopper, Nephotettix cincticeps Uhler, by a sight-count method. Res Popul Ecol 13:187–200
Kobayashi T, Takada M, Takagi S, Yoshioka A, Washitani I (2011) Spider predation on a mirid pest in Japanese rice fields. Basic Appl Ecol 12:532–539
Losey JE, Denno RF (1998) Positive predator–predator interactions: enhanced predation rates and synergistic suppression of aphid populations. Ecology 79:2143–2152
Mader V, Birkhofer K, Fiedler D, Thorn S, Wolters V, Diehl E (2016) Land use at different spatial scales alters the functional role of web-building spiders in arthropod food webs. Agric Ecosyst Environ 219:152–162
Marc P, Canard A (1997) Maintaining spider biodiversity in agroecosystems as a tool in pest control. Agric Ecosyst Environ 62:229–235
Matsumura M, Otuka A (2009) Recent occurrence of the small brown planthopper and the rice stripe virus disease in Japan. Plant Protect 63:293–296 (in Japanese)
Miyashita T, Chishiki Y, Takagi SR (2012) Landscape heterogeneity at multiple spatial scales enhances spider species richness in an agricultural landscape. Popul Ecol 54:573–581
Nakasuji F, Yamanaka H, Kiritani K (1973) The disturbing effect of micryphantid spiders on the larval aggregation of the tobacco cutworm, Spodoptera litura (Lepidoptera: Noctuidae). Kontyû 41:220–227
Ogura Y, Sakai H, Koike J (2014) Occurrence of rice stripe disease in Gunma Prefecture. Ann Rep Kanto-Tosan Plant Protec Soc 61:13–17 (in Japanese)
Okabe K, Sugiyama A (2016) Occurrence and control of rice stripe disease in Ibaraki Prefecture. Plant Protec 70:89–95 (in Japanese)
Okuma C, Lee MH, Hokyo N (1978) Fauna of spiders in a paddy field in Suwon, Korea I. Esakia 11:81–88
Pei H, Nakamura K (2012) Biodiversity and abundance of spider assemblages in restored terraced paddies. Acta Arachnol 61:31–39
Picchi MS, Bocci G, Petacchi R, Entling MH (2016) Effects of local and landscape factors on spiders and olive fruit flies. Agric Ecosyst Environ 222:138–147
R Core Team (2014) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.r-project.org/
Riechert SE, Lockley T (1984) Spiders as biological control agents. Annu Rev Entomol 29:299–320
Samu F, Szinetár C (2002) On the nature of agrobiont spiders. J Arachnol 30:389–402
Sanada-Morimura S, Sakumoto S, Ohtsu R, Otuka A, Huang SH, Van Thanh D, Matsumura M (2011) Current status of insecticide resistance in the small brown planthopper, Laodelphax striatellus, in Japan, Taiwan, and Vietnam. Appl Entomol Zool 46:65–73
Schmidt MH, Tscharntke T (2005) The role of perennial habitats for Central European farmland spiders. Agric Ecosyst Environ 105:235–242
Schmidt MH, Thies C, Nentwig W, Tscharntke T (2008) Contrasting responses of arable spiders to the landscape matrix at different spatial scales. J Biogeogr 35:157–166
Settle WH, Ariawan H, Astuti ET, Cahyana W, Hakim AL, Hindayana D, Lestari AS, Pajarningsih S (1996) Managing tropical rice pests through conservation of generalist natural enemies and alternative prey. Ecology 77:1975–1988
Shiba T (2016) Present state of rice stripe disease -Introduction-. Plant Protec 70:77–78 (in Japanese)
Shinkai A (1962) Studies on insect transmissions of rice virus diseases in Japan. Bull Nat Inst Agric Sci Ser C 14:1–112
Sunderland K, Samu F (2000) Effects of agricultural diversification on the abundance, distribution and pest control potential of spiders: a review. Entomol Exp Appl 95:1–13
Takada MB, Yoshioka A, Takagi S, Iwabuchi S, Washitani I (2012) Multiple spatial scale factors affecting mirid bug abundance and damage level in organic rice paddies. Biol Control 60:169–174
Takada MB, Kobayashi T, Yoshioka A, Takagi S, Washitani I (2013) Facilitation of ground-dwelling wolf spider predation on mirid bugs by horizontal webs built by Tetragnatha spiders in organic paddy fields. J Arachnol 41:31–35
Takada MB, Takagi S, Iwabuchi S, Mineta T, Washitani I (2014) Comparison of generalist predators in winter-flooded and conventionally managed rice paddies and identification of their limiting factors. SpringerPlus 3:418
Tanaka K (2016) Functional biodiversity indicators and their evaluation methods in Japanese farmlands. In: Yagi K, Kuo GC (eds) The challenges of agro-environmental research in Monsson Asia, NIAES Series No.6. pp. 157–168
Tanaka K, Endo S, Kazano H (2000) Toxicity of insecticides to predators of rice planthoppers: spiders, the mirid bug and the dryinid wasp. Appl Entomol Zool 35:177–187
Tscharntke T, Bommarco R, Clough Y, Crist TO, Kleijn D, Rand TA, Vidal S (2007) Conservation biological control and enemy diversity on a landscape scale. Biol Control 43:294–309
Tsukahara T (2016) Occurrence and control of rice stripe disease in Tochigi Prefecture. Plant Protec 70:96–99 (in Japanese)
Tsutsui MH, Tanaka K, Baba YG, Miyashita T (2016) Spatio-temporal dynamics of generalist predators (Tetragnatha spider) in environmentally friendly paddy fields. Appl Entomol Zool 51:631–640
Tuck SL, Winqvist C, Mota F, Ahnström J, Turnbull LA, Bengtsson J (2014) Land-use intensity and the effects of organic farming on biodiversity: a hierarchical meta-analysis. J Appl Ecol 51:746–755
Usio N, Miyashita T (2015) Social-ecological restoration in paddy-dominated landscapes. Springer, Tokyo
Willer H, Yussefi M, Sorensen N (2010) The world of organic agriculture: statistics and emerging trends 2008. Earthscan. https://doi.org/10.1017/S0014479709007807
Acknowledgements
We thank Shu-ichi Sugiyama, Toshihiro Yoshida, Yoshiharu Yoshida, and Tamotsu Yoshida for allowing us to survey their rice paddy fields. This work was supported by grants from the Ministry of Agriculture, Forestry, and Fisheries of Japan through a research project titled “Development of agricultural pest control system by the effective use of indigenous natural enemies” and by a Japan Society for the Promotion of Science Grant-in-Aid for Challenging Exploratory Research (17K20074).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no financial or other types of conflicts of interest.
Additional information
Handling Editor: Marta Montserrat
Rights and permissions
About this article

Cite this article
Baba, Y.G., Kusumoto, Y. & Tanaka, K. Effects of agricultural practices and fine-scale landscape factors on spiders and a pest insect in Japanese rice paddy ecosystems. BioControl 63, 265–275 (2018). https://doi.org/10.1007/s10526-018-9869-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10526-018-9869-5