
Abstract
Marine macroalgae (seaweeds) consist of a diverse range of species with diverse morphology and bioactive properties. The potential for genetic improvement to increase the value of the production of such molecules and other economically important traits is likely to be high as procedures for cultivation of many species are well established, genetic diversity is often high, sexual propagation is often possible and seaweeds generally have a short generation time that allows rapid transition from one selected generation to the next. The need for genetic improvement has been voiced by industry for many years. Despite this, there is little published research describing seaweed genetic improvement methodology, results or impact. This review draws on knowledge from other groups of species and describes options, benefits and knowledge needed for the genetic improvement of macroalgae. In conclusion, the review highlights how a well-designed genetic improvement program, which targets one or a few select economically valuable traits whilst limiting inbreeding, could provide potentially large benefits for industries reliant on the culture of seaweeds. Progress will, to a large extent, rely on the magnitude of genetic variation for the trait in question.
Similar content being viewed by others



Avoid common mistakes on your manuscript.
Introduction
Macroalgae consist of a diverse range of some 25,000 species with differing morphologies and bioactive properties (see Raja et al. 2008; Holdt and Kraan 2011 for reviews). As the basic requirements for production of marine macroalgae are sunlight, nutrients and salt water, seaweeds are very efficiently grown in aquaculture and contribute approximately 16 million tons valued at US$7.4 billion to the global trade of aquaculture production (Food and Agriculture Organisation of the United Nations 2010). Considering that this scale of production employs many thousands of people worldwide and is recognised as important to the future of food security and sustainable blue carbon concepts (Food and Agriculture Organisation of the United Nations 2010), reliable cultivation strategies are needed to mitigate the risks and remove barriers to the growth and sustainability of this industry. In comparison to terrestrial crops and some other aquaculture industries, it is evident that seaweed cultivation, although of considerable scale, is still in its infancy and, thus, vulnerable. Strategic management of cultivated genetic seaweed resources has been lacking.
Like any cultured organism, performance in aquaculture depends, in part, on the environment in which a particular species or strain is grown and also on genetic variability. All traits in living organisms are controlled, to some extent, by genetics (inherited effects), environment and the interaction of genes and environment (G × E). Understanding the genetic and environmental drivers of performance has provided strong advances in crop and livestock production. Increased use of selectively bred stocks leads to more efficient utilisation of nutrient, labour and water resources and will, therefore, be important for meeting increasing future demands for food production (Gjedrem et al. 2012). There are several reasons why we would expect the potential to make genetic improvement in seaweed species to be high:
-
Procedures for the cultivation of some species are established,
-
A range of diverse phenotypes exist for macroalgal species along the world’s coastlines and recent ecological and evolutionary studies have demonstrated that high genetic diversity exists between and within some species (e.g., Hays 2007; Monro and Poore 2005),
-
Sexual propagation under cultivation is possible for a number of commercially important species (e.g., Gutierrez et al. 2006),
-
Seaweed species generally have a short generation time, allowing a rapid transition from one selected generation to the next, and
-
The need for genetic improvement has been voiced by industry for many years.
Despite the above, little published work exists describing seaweed genetic improvement methodology, research results or impact. In a literature search on selective breeding (Google Scholar and Web of Knowledge with search terms “selective breeding” and “plant” or “microalgae” or “macroalgae” or “animal” or “aquaculture”), only 3 % of references referred to algae, and of these, only 10 % (or 0.3 % of the total) referred to macroalgae. In contrast to the development and progress of functional genomics in terrestrial plants, Chan et al. (2006) found that seaweeds had received little attention worldwide and were not included in efforts to elucidate gene functions.
From the literature, it appears that the creation of homogenous strains or production of clones with useful traits from wild populations has been the aim of “genetic improvement” initiatives so far. Van Der Meer (1990) recognised that traditional selective breeding had only been applied to a limited extent for the improvement of commercial red algal cultivars and wrote that, “Despite the advances of molecular genetics, it is likely that such traditional selective breeding, rather than genetic engineering will be the most important commercial aspect of red algae genetics in the foreseeable future”. Here, an updated review of genetic improvement initiatives for macroalgae is provided to support our contention that selective breeding has not progressed significantly since Van Der Meer’s prediction over 20 years ago. Building on this, an overview of three key strategies for genetic improvement of macroalgae based on knowledge from other industries is provided. Finally, we consider the benefits that a selective breeding approach might provide and the knowledge that would be needed to initiate a program for macroalgae genetic improvement.
Status of genetic improvement in macroalgal crops
Selection of Porphyra spp. has been practised, to some extent, since knowledge of the biology and control of reproduction became known in 1949. At the time of Van Der Meer’s review (1990), strain selection over 10–15 years in Japan had resulted in 30 recognised cultivars as well as hundreds of informally recognised strains. Improvement in the size of fronds had been achieved by selection for delayed fertility (Miura 1975), colour variants had been cultivated on a small scale and breeding objectives were being directed toward increased handling efficiency and quality (e.g., strains that produce monospores, that are disease resistant or adapted to local conditions). The methods practiced for intensive breeding of the most commonly grown strain in Japan, Porphyra yezoensis f. narawaensis, resulted in increased uniformity of the stock used for nori cultivation (Niwa et al. 2009). However, the breeding of P. yezoensis in Japan, China and South Korea has involved numerous exchanges of different cultivars with diverse genetic backgrounds, and records of the maintenance and breeding of stock are lacking (Hu et al. 2010). Mostly, however, the approach for such genetic improvement has been based on selection of a few strains from the wild with limited representation or knowledge of existing genetic diversity, limited knowledge of other characteristics of importance, and mutation/hybridisation of these strains to deliver novel or improved attributes (Zhang et al. 2011a; Niwa et al. 2004). Such approaches do not address the long-term opportunity for improvement through selective breeding but deliver shorter term and narrowly focused improvements. There is constant demand amongst cultivators of nori to make improvement to production traits, and different market segments (consumers and manufacturers of seaweed products) also have specific traits they would like to see improved (e.g., Chan et al. 2006). Therefore, even in the most well-established and high value cultivated genus of macroalgae, Porphyra, there is much to be gained from more strategic approaches to genetic improvement. Recent evidence that shows that most cultivars in Asia are probably a species of the genus Pyropia rather than Porphyra, highlights the lack of genetic knowledge for this important food crop (Sutherland et al. 2011).
Other red seaweeds are cultivated primarily for their hydrocolloid production and provide important income for many developing nations; however, in the near future, a lack of strategy in maintaining diverse genetic resources may negatively affect this important economic activity. In Chile, the strategy for cultivating Gracilaria chilensis has been to bypass sexual reproduction and maintain a pure gene pool (Guillemin et al. 2008). This has resulted in the creation of homogeneous strains or clones that cannot be genetically improved without going back to the wild stock or hybridising with another cultivated strain. In the Philippines, “selection” of Eucheuma has resulted in the production of a vigorous non-sporulating clone of Kappaphycus alvarezii known as Tambalang (Doty and Alvarez 1975). Ask and Azanza (2002), in their study of this clone, conclude that, “no true advances have taken place in commercial eucheumatoid farming in over a decade” as no net increase in production per unit time, effort, area and cost has occurred in this period. They suggest that research should focus on replacing the vegetative system for reproduction with sexual propagation of the crop and that strains should be developed through “continuous selection of wild varieties, breeding programs and genetic manipulation”.
Clones or homogenous strains are also likely to be less able to cope with the outbreak of new diseases. In Chile, the cultivation bottleneck with establishment of farmed populations of G. chilensis, coupled with subsequent vegetative (clonal) propagation, resulted in low genetic diversity compared to wild source populations (Guillemin et al. 2008). A sharp increase in the number of farms during the 1980s led to a large increase in production of this species; however, problems have arisen such as production declines and outbreaks of pests and pathogens (Buschmann et al. 2001). Reduced genetic diversity in naturally outbreeding species is known to affect the ability of such populations to resist disease outbreaks and to limit the scope for future genetic improvement, amongst other negative consequences, and could be contributing to the problems that have arisen in Chile. Similarly, the consequences of limited genetic variation in the seaweed farms of the Philippines is emerging, with increasing disease and epiphytism that will have consequences for local livelihoods and the vertically integrated processing industries that rely on this production (Bixler and Porse 2011; Loureiro et al. 2012). The narrow genetic cultivation base used in Asia has also been used to establish seaweed cultivation in Eastern Africa with the transfer of Kappaphycus and Eucheuma cultivars (Halling et al. 2012). Little attention is being paid to the choice of genetic material, establishment or maintenance of genetic diversity or potential benefits from genetic improvement by nations starting seaweed cultivation.
The other major seaweed crop for food, iodine and alginate products are species of brown algae, such as Laminaria/Saccharina (a split of the genus known as Laminaria into Saccharina and Laminaria clades has been proposed by Lane et al. 2006), most of which are cultivated in China. Here, some recent efforts have been made in establishing improvements to growth and iodine content through short-term line breeding trials (Li and Wu 2012). Hybridisation of different selected inbred lines produces Laminaria of higher yield due to heterosis (26–76 % higher than control parental lines) and has been used to introduce new traits selected in the parent lines, such as tolerance to strong irradiance, high seawater temperatures and tissue rot (Li et al. 2007, 2008a). The degree of heterosis for blade dry weight, fresh weight and length has been found to be significantly and positively correlated with genetic distance as measured using microsatellite markers (Li et al. 2008b). The growth rate, blade and hapteron morphology of self-crosses and hybrids of several natural geographic populations of Alaria esculenta have also been compared with regard to cultivation potential (Kraan et al. 2000). Further, genetic manipulation of Laminaria (Saccharina) japonica has been trialled successfully and used to incorporate a vaccine gene into the kelp genome, and methods of genetic transformation have been established for Undaria sp. and several red algae genera such as Porphyra, Gracilaria and Ceramium (reviewed by Qin et al. 2004). Although the introduction of new functional genes in this way may add value to the crop, there has been no consideration of the overall longevity of the industry and what traits of interest should be considered in the crop as a whole.
In summary, algal producers have long recognised that there would be large economic benefits from genetic improvement. The seaweed biomass itself is the primary cost for the production of hydrocolloid-based products (Bixler and Porse 2011) and any genetic improvements that reduce those costs will have large benefits. Despite this, quantitative genetic information about algal species is lacking. The focus of genetic improvement initiatives for macroalgae has been largely short-term and restricted to the production of homogeneous strains or clones and their hybrids. It seems that little has been learnt or adopted from the research efforts for terrestrial plant, livestock or aquaculture species.
Benefiting from the knowledge developed for other species
The establishment of well-designed genetic improvement programs has proven key to the successful development and ongoing viability of major livestock, plant and aquaculture industries worldwide. The same genetic principles apply to genetic improvement across the different kingdoms of plants and animals, and although some genetic improvement techniques are specific to particular (e.g., animal) lifecycles, most techniques should be broadly applicable across different kingdoms. Genetic improvement programs have been established for most important outcrossing crops. Notably, the rice industry has achieved major productivity and value gains at a scale of billions of dollars due to selective breeding strategies and this approach is anticipated to continue to deliver gains in the traditional terrestrial crops into the future (Conway and Toenniessen 1999). In addition, forest plants such as the eucalypts and pines have benefited significantly from selective breeding approaches (Eldridge et al. 1993; Kirst et al. 2004).
More recently, but still over a number of decades, aquaculture, excluding macroalgae, has embraced the selective breeding approach. Like algae, fish and shellfish are aquatic species that are often highly fecund and release gametes into the water column for external fertilisation; therefore, much can be learnt from modern fish selective breeding programs and adapted to breeding programs for algae.
Major world aquaculture species (e.g., Atlantic salmon, rainbow trout, carp, Mediterranean sea bass, white shrimp, catfish, tilapia and oysters) are typically serviced by two or three “centralised” selective breeding programs (“centralised”, meaning a selective breeding program that acts in a coordinated way as one program with a pool or nucleus of breeding individuals, often operating as a separate business entity). Evaluation of the impact of these selective breeding programs has been very positive. Thodesen et al. (1999) compared the performance of unselected animals to selectively bred Atlantic salmon grown under the same conditions after five generations of selection on growth rate. The growth rate of the selectively bred fish was more than doubled (+113 %), feed consumption increased by less than 50 %, protein retention was 9 % improved, energy retention was 14 % improved and the feed conversion ratio (feed to gain) was 20 % improved. After seven generations, this improvement in feed conversion ratio alone was estimated to have resulted in a saving of around US$300 million in feed costs each year to the US$1–2 billion export industry in Norway. There are now several examples of successful fish selective breeding programs providing large benefits of this type to the industries they serve (e.g., Khaw et al. 2008; Ponzoni et al. 2008). Typically, fish breeding programs have been shown to achieve improvements of 10–15 % per generation for quantitative traits like growth rate and can yield benefit–cost ratios of around 15:1 per generation of selection (Gjedrem 2000, 2005b).
Options for genetic improvement
Here, we consider three general options for the genetic improvement of macroalgae, learning from the experiences of other cultivated species:
-
1.
Selective breeding (option utilised for some plants and normally utilised for fish and livestock—Fig. 1),
Fig. 1 
Selective breeding approach for genetic improvement. Field tests, when the performance of specimens from each family is evaluated under different conditions outside of the breeding nucleus (e.g., with growth in different culture systems). Chall. tests, when specimens from each family are evaluated with regard to performance under extreme conditions (e.g., extreme temperatures, nutrients, disease, herbivores, etc.)
-
2.
Line breeding (option sometimes used for plants and most common option used for algae species in the past—Fig. 2), and
Fig. 2 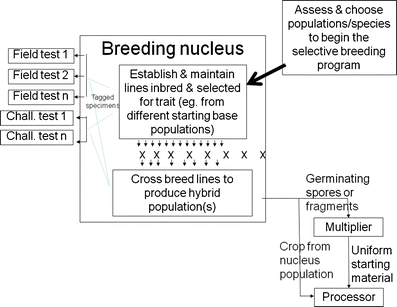
Line breeding approach for genetic improvement
-
3.
Genetic modification (use of molecular biology technologies to alter the regulation of gene expression, addition of new genes or changed ploidy—Fig. 3).
Fig. 3 
Genetic modification approach for genetic improvement



In later sections, we will explain how these options can be evaluated in a way that accounts for the scale of the breeding program, the biology of the species, traits of key interest, potential cost–benefits and the structure/degree of development of the industry. General advantages and disadvantages of the three options for genetic improvement (and some combinations of options) are shown in Table 1.
Selective breeding
Most livestock and fish and some plant genetic improvement programs utilise continuous selective breeding programs (Option 1) that aims to make stepwise genetic improvement with every generation of selection. By extending the process of natural selection in a controlled way, each generation of improvement builds on the improvements made in previous generations (Table 1 and Fig. 1). The science of quantitative genetics for selective breeding is well advanced, particularly for livestock and aquaculture (Gjedrem 2005b; Falconer and Mackay 1996), and years of research have led to the incorporation of new procedures that allow effective mate choice to limit inbreeding (e.g., Meuwissen and Sonesson 2004) and make effective use of modern technologies such as genomics (e.g., Meuwissen et al. 2001).
Selective breeding programs need to be continuously maintained and funded into the future if improvement is to continue. In essence, such breeding programs are for the greater good and viability of whole industries and work as an effective collaboration with industry where offspring are challenged to field cultivation conditions. The response in the field and feedback from farmers are important to this approach. A strong commitment by the industry is needed to continue breeding through good and bad times in the production sector, and the breeding entity itself (business or cooperative) needs to be profitable. The main biological prerequisite for genetic improvement by selective breeding is that additive genetic variation plays a role in controlling traits that are included in the breeding goal, meaning that there are a number of genes affecting the trait of interest, and that the effect of each allele for each of these genes is additive.
Line breeding
Some plant and the few existing seaweed genetic improvement programs utilise line breeding where inbreeding with trait selection is used to create lines, and the lines are crossed to produce superior crossbred or F1 individuals (Option 2, Fig. 2). Crossbred or F1 individuals will be superior to the original population if the selection intensity is high when creating the inbred lines and if heterosis occurs for non-additive genetic variants affecting the trait of interest with the crossing of the lines.
In comparing the current system of algal line breeding used in Japan to an option for continuous genetic improvement with selective breeding, we need to consider the magnitude of additive compared to non-additive genetic variation for traits of interest to the industry. If the non-additive genetic variance is considerable, a long-term cross-breeding program could be applied, for example, using reciprocal recurrent selection (RRS), which aims to improve both the general and specific combining abilities, thus potentially taking advantage of selection and cross-breeding (Gjedrem 1985; Falconer and Mackay 1996). If both additive and non-additive genetic variations are considerable, an additional potential consideration of importance is to what extent and for how many years will the benefits from the selection used when creating the inbred lines and hybrid vigour outweigh the accumulated genetic improvement expected with continuous selective breeding. There have been no demonstrations of trait heterosis or hybrid vigour amongst red algae when strains are crossed (Patwary and Van Der Meer 1983; Van Der Meer 1990; Zhang and Van Der Meer 2012), and little has been published about the presence or absence of heterosis in other macroalgae (heterosis was detected when crossing Laminaria gametophyte clones, Li et al. 2007, 2008a, b). Finally, the F1 from line breeding, where genotypes are likely to be replicated in most individuals of the crosses, would be expected to be more uniform and consistent than those produced from selective breeding. If a good cross is found using line breeding, it is possible to continue performing the same cross as long as both lines continue to exist.
There have been no evaluations of the success of current and past line breeding genetic improvement programs for macroalgae, and information about the original source of cultivated seaweed strains is lacking and there is no record of systematic comparison testing of possible hybrids and pure strains. Eleven of the 28 species of Porphyra (nori) that are found in Japan are thought to be under cultivation, the most commonly used strain being P. yezoensis f. narawaensis A. Miura. Molecular analysis of the nuclear internal transcribed spacer sequence (ITS) and plastid RuBisCo spacer regions revealed much higher levels of genetic variation in natural populations of P. yezoensis and P. tenera than existed in Porphyra strains that are commonly used for cultivation, and Niwa et al. (2009) concluded that, in fact, four separate, wild species had been sampled for cultivation. Although F1 from interspecific hybridisation between two of the strains developed normally and released numerous conchospores, almost all conchospore germlings failed to survive far beyond the four-cell stage (Niwa et al. 2010). Allodiploidy and polyploidy resulting from interspecific hybridisation is a natural mechanism maintaining species differences amongst some Porphyra spp. (Niwa et al. 2010).
Genetic transformation/modification
Genetic improvement can also be made by genetic modification (Option 3) such as changes to ploidy, mutagenesis (e.g., mutagenesis and phenotypic selection of microalgae for biofuel production, Bonente et al. 2011) or genetic manipulation to add genes or control their expression, (e.g., as reviewed by Kathiresan and Sarada (2009) for Haemotococcus pluvialis microalgae). Whilst large one-off gains can be achieved with genetic manipulation, a biosecure facility may be required to evaluate the organism, and there is a risk that public acceptance of genetically modified food organisms will be low and there may be other traits of importance that will be negatively affected by the modification. Also, in the case of targeted gene insertion or expression control, large and expensive research programs would be needed to determine which gene(s) to modify and how they should be modified (e.g., transformation technique).
If a particular gene is known to have a major effect on a desirable trait or if the desirable traits from two species or lines is to be rapidly combined in the one plant, some form of genetic modification or cell manipulation could be used to produce a new plant that contains the desired genes or that expresses genes at the desired level. A lot of attention has been given to the genetic transformation and propagation of algae in the literature (Qin et al. 2004; Reddy et al. 2008a, b; Kathiresan and Sarada 2009; Wang et al. 2010). This is because it is possible to artificially introduce new products or processes into the algal genome using genetic transformation (Raja et al. 2008). Model genetic transformation systems have been developed for Dunaliella salina (Feng et al. 2009) and for Saccharina japonica as described above (Qin et al. 2004). Genetic transformation methods and site-directed mutagenesis are being applied to improve and regulate ketocarotenoid and astaxanthin production in Hematococcus algae (Steinbrenner and Sandmann 2006; Kathiresan and Sarada 2009). However, transformation is normally only used to manipulate or introduce single genes. When quantitative traits (traits controlled by many genes of small and large effect) or multiple traits are considered, selective breeding is likely to be a more effective and efficient option.
Manipulation of ploidy is another means of genetic manipulation that can sometimes enhance performance (e.g., for oysters, Nell et al. 1994; Guo et al. 1996). This has been tried as a way of generating one-off genetic improvement in red algae, but the polyploids created so far have had reduced performance (Patwary and van der Meer 1984). Ploidy is known to affect the production of polysaccharides in some algae (Chen et al. 1975). In addition, somatic cell hybridisation has been undertaken with macroalgae and studies on interspecific and intergeneric protoplast fusion clearly demonstrated the possibilities of developing new, and in certain cases, improved strains of seaweeds (Reddy et al. 2008a).
Finally, sequential mutagenesis and selection could be used as a process for enhancing the selective breeding or line breeding strategies outlined above by introducing new genetic variants each generation. For example, two successive rounds of chemical mutagenesis and selection resulted in isolates of Chlamydomonas reinhardtii with greatly enhanced O2 tolerance (Flynn et al. 2002).
Production of uniform clones
Regardless of the option chosen above, one way to produce a uniform starting material for downstream processing might be to rely on asexual reproduction of algae (e.g., the distribution of clones or spores/gametophytes from one or a few elite specimens for production). The testing of clones could also be useful for providing more accurate estimates of the genotypic value for particular traits (rather than a breeding value).
Why might a genetic improvement program be beneficial for macroalgae?
Reduced reliance on wild harvest
Harvest from the wild can be dependent on climatic effects, season, etc. and could have local environmental impacts (Beattie et al. 2011). Wild harvest practices are facing management challenges and impacts on ecosystems are being detected (Maine Seaweed Council 2002; Vasquez 2011). Cultivation already contributes to the bulk of seaweeds marketed for human consumption and addressing the growing seaweed food industry through wild harvest will not be a scalable option. By culturing algae for genetic improvement in land- or sea-based systems, it should be possible to ensure year-round supply and eliminate harvest impacts. Once established, a genetic improvement program would not depend on the input of harvested wild stock. The supply of material to processors or on-growers for the purification of bioactive molecules would be derived from the latest generation of genetically improved stock sourced from the breeding program.
Quality and consistency
When wild seaweed is harvested and used as the basis for production of bioactives, compounds with inconsistent nutritional and health values are produced (Rioux et al. 2009). Traceability and quality assurance is of importance when producing high value products, especially functional foods and bioactive components (Hafting et al. 2012). The consistency of stock can be controlled and ensured in various ways by the genetic improvement program. In order to give the highest order of consistency, genetic diversity could be maintained within the breeding population, whilst the breeding program supplies clones from an “optimal” specimen to growers. These specimens could be chosen because of their suitability to specific environmental conditions or specific desired end markets.
Substantiation and labelling of health benefits is receiving increasing attention by the public and food regulators (Tapsell 2008). Being able to substantiate the health benefits of a product gives companies competitive advantages over rivals and establishes quality assurance. One key problem is that the final manufactured product may contain inconsistent quantities and types of compounds from batch to batch. By producing a more consistent starting material with higher levels of healthy compounds, a selective breeding program could allow industry to better substantiate and maintain consistency over the health benefits of their product.
Improved production resulting in reduced risks and greater value to the farmer
The main need of most aquaculture producers is to increase production and value and to reduce costs per tank/pond/cage (e.g., Robinson et al. 2010a, b). In the case of seaweed production for bioactives, potential benefits could be derived through improvement of:
-
Efficiency (production per unit area, sun hours or gram of nutrient): typically for a quantitative trait that can be easily and directly measured on the breeding candidates like growth rate, it is possible to achieve genetic improvement of around 10–14 % per generation with selective breeding (Gjedrem and Thodesen 2005). In the case of rapid lifecycle species like Ulva, assuming a generation interval of 3 months, growth rate could be twice that of wild Ulva within 1.5 years if growth rate was the only trait selected (authors’ own calculations, assumptions and estimates). Since maintenance costs (e.g., feed) are much lower than for animals, this would result in 10–14 % benefit per generation with few additional costs. Annual taxa like Laminaria/Saccharina spp. will require longer time frames for improvement.
-
Specific target compounds or compound combinations for food: in order to realise the value of seaweed, we need to be able to increase and control the content and quality of valuable compounds or metabolites. Natural variability in many of these compounds exists; however, little is known about the extent that genetic variability affects the production and quality of such compounds.
The presence, content and function of certain polysaccharides, such as fucoidans and ulvans, is species specific (Craigie 2010; Eluvakkal et al. 2010, Winberg pers comm.), and molecular or other identification is needed to choose the best species for optimising production (Brown et al. 2004). Once the most valuable genetic sources of the compound are identified, this source can be used as the basis for a selective breeding program. Other potentially valuable target compounds are known to be largely under environmental control, for example, pigment concentrations (Stengel et al. 2011), and genetic improvement may, therefore, be less of an issue. Further traits, e.g., fatty acid profiles, may have attributes characterised, in part, by the environment and, in part, by genetics (McCauley et al. in press) that then can be optimised through both breeding and according to environmental conditions. Design of the genetic breeding program, therefore, needs to be guided by knowledge regarding the control of valuable traits and the needs of the industry.
-
Avoidance of maturity during production: a challenge for the reproductive control of macroalgal species is that the life cycle is often taxon specific with differences in life history cycles and stages across the three main macroalgal groups, as well as unique environmental triggers and chemical signals amongst closely related taxa (e.g., Wichard and Oertel 2010). For Porphyra and Ulva production, sporing or reproductive maturity has a negative effect on condition and production, for example, by reducing biomass (Miura 1975; Han et al. 2003; Lüning et al. 2008). Increased tolerance to stress and increased control over the reproductive cycle would be traits of importance in these species.
-
Resistance to grazing or disease: many macroalgae species are highly susceptible to grazing pressure and low levels of secondary metabolites associated with resistance to herbivory have been detected (e.g., Erickson et al. 2006; Molis et al. 2008). Increased resistance to herbivory could be deliberately selected based on values for such chemical traits. Even if little deliberate emphasis is put on resistance to herbivory directly, good condition (high condition factor score) could provide a means for indirect selection.
-
Tolerance to high nutrients: increasing tolerance to specific high nutrient conditions could also reduce grazing pressure and epiphytism. For example, Ulva spp. thrive and dominate under conditions with high nitrogen in the reduced form of ammonia (e.g., Kennison et al. 2011). By increasing tolerance to nitrogen further, the purity of Ulva culture would increase, as other species would be less likely to grow under these conditions. Increasing the tolerance to high nitrogen or other compounds also opens up further possibilities for the use of macroalgae for bioremediation purposes (e.g., to extract nutrients from sewage, aquaculture outflows or in mine tailings).
-
Cooking or eating attributes: these are difficult traits to select because they typically require the use of trained taste panels (e.g., specialised anori assessors who exist in Japan) and because tastes and preparation methods vary depending on the market and value. One way such traits can be valued is through market surveys where buyers are given a range of choices and prices, and some money to spend (e.g., colour or welfare choices for buyers of salmon, Alfnes et al. 2006; Olesen et al. 2010). Taste stimulators such as natural glutamates (“umami peptides”) or other chemicals could be associated with the perception of flavour (Temussi 2012), and tests of such compounds might provide objective measurements for selective breeding. They might also affect the palatability of algal-based fish feeds and other products (e.g., the taste of abalone is affected by consumption of dimethylsulfoniopropionate in algal-based diets, Smit et al. 2007). Finally, an important consideration is whether there will be an economic benefit to growers, producers or processors if cooking or eating attributes are improved.
Avoidance of inbreeding and loss of genetic variability (with positive flow on effects on all of the above benefits)
A selective breeding program could suffer from high rates of inbreeding and loss of allelic variation if relationships between breeding candidates are not considered when making selection decisions. By maintaining a genetically healthy population (by minimising inbreeding and loss of genetic variability), there are long- and short-term benefits for the industry. In other species, inbreeding and loss of variability have been found to be associated with reduced production performance, greater susceptibility to disease and reduced capacity for genetic improvement (e.g., review for fish and shellfish species, Fjalestad 2005), and evidence for similar effects in macroalgal industries is emerging as reviewed above.
Knowledge needed to initiate a genetic improvement program for macroalgae?
Control of reproduction
Two important prerequisites for making genetic improvement are that (1) the life cycle can be closed in the breeding nucleus and (2) that it is possible to synchronise the production of gametes so that desirable mate combinations can be produced and compared. Both prerequisites need to be met to ensure that genetic improvement is possible to achieve and can occur at a rapid rate. It is well established that life history cycles and stages differ not only across the three main groups of macroalgae, Chlorophyta, Phaeophyta and Rhodophyta, but also within these (Araújo et al. 2011). In addition, different reproductive responses to environmental cues are known to occur across species (Han et al. 2003; Wichard and Oertel 2010) as well as life stages (Roleda et al. 2007). Seasonal variation in the mode of reproduction is documented for some populations (Lüning et al. 2008; Alstrom-Rapaport et al. 2010); however, knowledge of species-specific life histories is considered to be a key aspect of knowledge required as a precursor to successful macroalgae industries with or without selective breeding programs (Baweja et al. 2009).
There are additional considerations needed for the design of selective breeding programs for macroalgae because of the life cycle typically observed in this kingdom. Fertilisation will produce sporophyte plants (2n) and for breeding, there is a need to produce gametophytes (1n). Therefore, there is a need to be able to induce meiosis in sporophytes and mitosis in gametophytes, determine which gametophytes produce +ve gametes and which produce −ve gametes, collect gametes from the plants and control fertilisation so that desirable mate combinations are produced. Sometimes, the identification of gametophyte and sporophyte stages can be a challenge for macroalgae.
One feature of macroalgae life cycles that opens possibilities for selective breeding is the possibility for producing and testing the performance of clones of individuals in different test environments or under different challenges (e.g., Figs. 1, 2 and 3). This is an interesting possibility that could be explored for some traits. Testing cloned replicates, particularly gametophyte (1n) clones, could allow the genetic value of individuals to be measured with little error. Of note is that although the lifecycle knowledge across many species is limited, there has been a lot of effort to develop seaweed tissue culture techniques (as reviewed by Reddy et al. 2008b; Baweja et al. 2009) that will assist in this way.
Evaluation of starting material for breeding and formation of base population for breeding
During the establishment phase, an important consideration is the choice of “seed stock” used to found the base population, as this affects the species’ production and profitability for subsequent generations. Two considerations are important for base stocks: (1) will adequate genetic variability be captured by the base population to allow for the selection of a range of traits into the future? (2) Have the best-suited variants (e.g., those with the highest production of a desirable compound in the aquaculture environment) been used? To answer both of these questions we need to have knowledge about what variation exists and how this variation can be crossed and utilised to yield “the best” variants. Knowledge of traits of importance and the influence of genetics and environmental effects is important (Stengel et al. 2011).
Recent work in China is moving towards establishing rapid methods to assess differences in photosynthetic performance and pigment composition across Porphyra species (Zhang et al. 2011b). Similar methods can be applied to other macroalgae, using crosses grown in a common environment, to compare the performance of different variants.
Heritability and correlated traits
Genetic improvement relies on the estimation of the genetic value of individuals and/or families. Heritability is a measure of the relative importance of genetic variance in determining a trait (expressed as a proportion of the total phenotypic variance). The expected rate of genetic improvement can be predicted once we have estimated the extent of genetic variability, heritability of the trait and the intensity of selection that can be placed on the trait. If there is more than one trait of interest, the intensity of selection for any trait will be affected by the relative weighting placed on that trait in a selection index. Also, some traits can be negatively or positively correlated with other traits. These correlations will affect the strategy that is used for making selection decisions in the breeding program and will also affect the rate of genetic improvement of all the traits in the index.
The direct measurement of the type and quantity of high value bioactives (oils, minerals, vitamins and proteins) typically involves complex and expensive chemistry (e.g., for sulphated polysaccharide ulvan in Ulva, see Robic et al. 2009; Alves et al. 2010). There would be large benefits for the breeding program if it were possible to measure such traits in a simple and reliable way. For instance, if the polysaccharide content (e.g., ulvan) is directly correlated with growth rate (i.e., kilogram of dry weight after a set period of growth), then the growth rate can be measured and used as a simple way of indirectly selecting for polysaccharide production (quantity).
The only published studies of heritability in marine algae relate to the genetics of anti-grazing compounds (e.g., furanones in red alga Delisea pulchra, Wright et al. 2004). The concentration of all four furanone metabolites were positively correlated, indicating that selection for one (e.g., compound 3) or total furanone concentration resulted in increased concentrations of all four (Wright et al. 2004). As consumption was negatively correlated with furanone concentration, the measurement of furanone levels could be used as a marker in a selective breeding program to improve the resistance of this alga to grazing.
Similarly, relatively high broad sense heritability (0.63) exists for phlorotannin content in Fucus vesiculosus in two different environments (Jormalainen and Honkanen 2004). Phlorotannin content is also believed to affect resistance to herbivory. When calculated from field data collected from different environments, genotype alone was found to explain around 30 %, whilst genotype together with genotype–environment interactions explained 46 % of the phenotypic variation in phlorotannin content. The broad-sense heritability of growth (weight and length) was also significant in this species (25–30 %).
Niwa et al. (2004) found that genetic variation for blade shape exists by comparing the development of blade shape between two completely homozygous conchocelis from two pure lines of Porphyra (HG-1 and HG-4). However, the heritability of blade shape, which directly relates to and is essential basic knowledge needed for improving crop production, is still yet to be evaluated for Porphyra spp.
Genetic environment (G × E) interactions
G × E are important interactions to consider if the seaweed to be genetically improved is to be grown out in different environments. If there is a strong genetic by environment interaction, then plants with genotypes that perform well in one environment might perform relatively poorly in other environments. If the breeding nucleus also grows and directly supplies a company with algae for the production of bioactives, G × E (if it exists for traits of interest to the company) will not be a problem; however, if other facilities are needed to supply the company, some specimens from each family should be sent for testing in those environments to check for and correct for G × E. The only reported research describing G × E in seaweeds has been on the genetics of compounds affecting herbivory (in relation to the genetics of natural ecosystems). “Fitness costs” with the production of phlorotannins have been found to vary in different environments (i.e., possible G × E effect on fitness, Jormalainen and Honkanen 2004).
Trait prioritisation, economic weighting and selection index
Trait prioritisation by industry, in consultation with geneticists, is the first step for developing breeding objectives for a genetic improvement program (Gjedrem 2005a). Traits need to be ranked on the basis of likely economic impact, potential for genetic improvement, practicality of measurement and selection and amount of research needed for implementing selection. The emphasis that is put on particular traits is important and usually a multi-trait “economic index” (which weights or emphasises specific traits, depending on their likely economic impact) is used as the basis of selection. The more traits that are included in the breeding objective for the breeding program, the less emphasis we can put on any single trait and the less genetic improvement we can achieve for any single trait. Therefore, a balance is needed where the most economically important traits are emphasised whilst looking after other features that might be more or less important in the future.
Ploidy determination
The morphology of the different gametophyte and sporophyte phases of numerous macroalgal taxa is identical, and there is a need to be able to determine ploidy so that gametophytes can be selected for breeding. Ploidy can have large effects on some traits (e.g., polysaccharide production, Chen et al. 1975). There are a number of techniques that have been applied to determine ploidy, e.g., flagella counts (Hiraoka et al. 2003), iridescence, fluorescence and light reflectance techniques (Brown et al. 2004; Zitta et al. 2011); however, some of these techniques are very time consuming. A promising technique that might be used to distinguish between haploid and diploid life phases is laser scanning cytometry (Kagami et al. 2005). Polymerase chain reaction (PCR)-based assays have also been devised for assessing ploidy in Chlamydomonas (Zamora et al. 2004).
If ploidy is found to affect the trait of interest, a number of questions will arise, as was the case for the cultivation of Chondrus crispus (Chen et al. 1975). Should sporophytes or gametophytes be grown for production? Are the best genes affecting traits in the sporophyte also the best affecting the trait in the gametophyte? Should selection be based on the performance of the gametophyte, sporophyte or both?
Cost–benefits, commercialisation and risks
As public funding is often scarce or unreliable, the establishment and continuation of a breeding program will normally depend on the existence of a commercially viable entity (breeding company or cooperative) to oversee the program, a viable business plan, access to suitably scaled facilities and the willingness of producers and/or multipliers to pay for the genetically improved stock. Risks need to be identified by performing strength, weakness, opportunity and threat (SWOT) and risk analyses. What economic issues might affect the program? What environmental risks, including biosecurity issues, could impact the program? The SWOT analysis by Bolton et al. (2009) on growing Ulva in integrated systems, as a commercial crop for abalone feed, has already highlighted many issues that will be relevant to establishing breeding programs for Ulva and can be used as a starting point.
Conclusion
In conclusion, although more than 24 % of total world aquaculture production consists of seaweed (FAO 2010), there are few published studies focussed on the genetic improvement of macroalgae. There is already evidence for poor genetics reducing the viability and value of some algal industries. This has occurred because of a lack of coordinated genetic management. Much can be learnt from the success of genetic improvement programs for plant, fish and livestock. A well-designed plan for genetic improvement, which targets selection on a few select economically valuable traits whilst limiting inbreeding, could provide potentially large benefits to existing and developing seaweed industries. Considering the existing investment, as well as the socio-economic value and future potential to contribute to food and nutrition limitations (e.g., Msuya 2011), large benefits will come from improving the long-term viability, growth and value of the global seaweed cultivation industry by making genetic improvement.
References
Alfnes F, Guttormsen AG, Steine G, Kolstad K (2006) Consumers’ willingness to pay for the color of salmon: a choice experiment with real economic incentives. Am J Agric Econ 88:1050–1061
Alstrom-Rapaport C, Leskinen E, Parnilo P (2010) Seasonal variation in the mode of reproduction of Ulva intestinalis in a brackish water environment. Aquat Bot 93:244–249
Alves A, Caridade SG, Mano JF, Sousa RA, Reis RL (2010) Extraction and physico-chemical characterization of a versatile biodegradable polysaccharide obtained from green algae. Carbohydr Res 345:2194–2200
Araújo R, Serrão EA, Sousa-Pinto I, Åberg P (2011) Phenotypic differentiation at southern limit borders: the case study of two fucoid macroalgal species with different life-history traits. J Phycol 47:451–462
Ask EI, Azanza RV (2002) Advances in cultivation technology of commercial eucheumatoid species: a review with suggestions for future research. Aquaculture 206:257–277
Baweja P, Sahoo D, García-Jiménez P, Robaina RR (2009) Review: seaweed tissue culture as applied to biotechnology: problems, achievements and prospects. Phycol Res 57:45–58
Beattie AJ, Hay M, Magnusson B, de Nys R, Smeathers J, Vincent JFV (2011) Ecology and bioprospecting. Austral Ecol 36:341–356
Bixler HJ, Porse H (2011) A decade of change in the seaweed hydrocolloids industry. J Appl Phycol 23:321–335
Bolton JJ, Robertson-Andersson DV, Shuuluka D, Kandjengo L (2009) Growing Ulva (Chlorophyta) in integrated systems as a commercial crop for abalone feed in South Africa: a SWOT analysis. J Appl Phycol 21:575–583
Bonente G, Formighieri C, Mantelli M, Catalanotti C, Giuliano G, Morosinotto T, Bassi R (2011) Mutagenesis and phenotypic selection as a strategy toward domestication of Chlamydomonas reinhardtii strains for improved performance in photobioreactors. Photosynth Res 108:107–120
Brown MT, Neish A, Harwood D (2004) Comparison of three techniques for identifying isomorphic phases of Chondrus crispus (Gigartinaceae). J Appl Phycol 16:447–450
Buschmann AH, Correa JA, Westermeier R, Hernandez-Gonzalez MD, Norambuena R (2001) Red algal farming in Chile: a review. Aquaculture 194:203–220
Chan CX, Ho CL, Phang SM (2006) Trends in seaweed research. Trends Plant Sci 11:165–166
Chen LCM, Craigie JS, McCandless EL, McLachlan JL, Neish AC, Shacklock PF, Walter JA (1975) Algal polysaccharide production. U.S. Patent 3879890
Conway G, Toenniessen G (1999) Feeding the world in the twenty-first century. Nature 402:C55–C58
Craigie JS (2010) Seaweed extract stimuli in plant science and agriculture. J Appl Phycol 23:371–393
Doty MS, Alvarez VB (1975) Status, problems, advances and economics of Eucheuma farms. Mar Technol Soc 9:30–35
Eldridge K, Davidson J, Harwood C, van Wyk G (1993) Eucalypt domestication and breeding. Clarendon, Oxford
Eluvakkal T, Sivakumar SR, Arunkumar K (2010) Fucoidan in some Indian brown seaweeds found along the Coast Gulf of Mannar. Int J Bot 6:176–181
Erickson AA, Paul VJ, Van Alstyne KL, Kwiatkowski LM (2006) Palatability of macroalgae that use different types of chemical defenses. J Chem Ecol 32:1883–1895
Falconer DS, Mackay TFC (1996) An introduction to quantitative genetics, 4th edn. Addison Wesley Longman Limited, Edinburgh Gate
Feng SY, Xue LX, Liu HT, Lu PJ (2009) Improvement of efficiency of genetic transformation for Dunaliella salina by glass beads method. Mol Biol Rep 36:1433–1439
Fjalestad K (2005) Breeding strategies. In: Gjedrem T (ed) Selection and breeding programs in aquaculture. Springer, Dordrecht, pp 145–158
Flynn T, Ghirardi ML, Seibert M (2002) Accumulation of O2-tolerant phenotypes in H2-producing strains of Chlamydomonas reinhardtii by sequential applications of chemical mutagenesis and selection. Int J Hydrogen Energy 27:1421–1430
Food and Agriculture Organisation of the United Nations (2010) The state of world fisheries and aquaculture. FAO, Rome
Gjedrem T (1985) Improvement of productivity through breeding schemes. GeoJournal 10:233–241
Gjedrem T (2000) Genetic improvement of cold-water fish species. Aquac Res 31:25–33
Gjedrem T (2005a) Breeding plans. In: Gjedrem T (ed) Selection and breeding programs in aquaculture. Springer, Dordrecht, pp 251–277
Gjedrem T (2005b) Selection and breeding programs in aquaculture. Springer, Dordrecht
Gjedrem T, Robinson N, Rye M (2012) The importance of selective breeding in aquaculture to meet future demands for animal protein: a review. Aquaculture 350–353:117–129
Gjedrem T, Thodesen J (2005) Selection. In: Gjedrem T (ed) Selection and breeding programs in aquaculture. Springer, Dordrecht, pp 89–112, c2005
Guillemin ML, Faugeron S, Destombe C, Viard F, Correa JA, Valero M (2008) Genetic variation in wild and cultivated populations of the haploid diploid red alga Gracilaria chilensis: how farming practices favour asexual reproduction and heterozygosity. Evolution 62:1500–1519
Guo XM, DeBrosse GA, Allen SK (1996) All-triploid Pacific oysters (Crassostrea gigas Thunberg) produced by mating tetraploids and diploids. Aquaculture 142:149–161
Gutierrez A, Correa T, Munoz V, Santibanez A, Marcos R, Caceres C, Buschman AH (2006) Farming of the giant kelp Macrocystis pyrifera in southern Chile for development of novel food products. J Appl Phycol 18:259–267
Hafting JT, Critchley AT, Cornish ML, Hubley SA, Archibald AF (2012) On-land cultivation of functional seaweed products for human usage. J Appl Phycol 24:385–392
Halling C, Wikström S, Lilliesköld-Sjöö G, Mörk M, Lundsør E, Zuccarello GC (2012) Introduction of Asian strains and low genetic variation in farmed seaweeds: indications for new management practices. J Appl Phycol. doi:10.1007/s10811-012-9842-0
Han TJ, Han YS, Kim KY, Kim JH, Shin HW, Kain JM, Callow JA, Callow ME (2003) Influences of light and UV-B on growth and sporulation of the green alga Ulva pertusa Kjellman. J Exp Mar Biol Ecol 290:115–131
Hays CG (2007) Adaptive phenotypic differentiation across the intertidal gradient in the alga Silvetia compressa. Ecol Lett 88:149–157
Hiraoka M, Shimada S, Ohno M, Serisawa Y (2003) Asexual life history by quadriflagellate swarmers of Ulva spinulosa (Ulvales, Ulvophyceae). Phycol Res 51:29–34
Holdt SL, Kraan S (2011) Bioactive compounds in seaweed: functional food applications and legislation. J Appl Phycol 23:543–597
Hu ZM, Liu FL, Shao ZR, Yao JT, Duan DL (2010) NrDNA internal transcribed spacer revealed molecular diversity in strains of red seaweed Porphyra yezoensis and genetic insights for commercial breeding. Genet Resour Crop Evol 57:791–799
Jormalainen V, Honkanen T (2004) Variation in natural selection for growth and phlorotannins in the brown alga Fucus vesiculosus. J Evol Biol 17:807–820
Kagami Y, Fujishita M, Matsuyama-Serisawa K, Yamamoto M, Kuwano K, Saga N, Kawano S (2005) DNA content of Ulva compressa (Ulvales, Chlorophyta) nuclei determined with laser scanning cytometry. Phycol Res 53:77–83
Kathiresan S, Sarada R (2009) Towards genetic improvement of commercially important microalga Haematococcus pluvialis for biotech applications. J Appl Phycol 21:553–558
Kennison RL, Kamer K, Fong P (2011) Rapid nitrate uptake rates and large short-term storage capacities may explain why opportunistic green macroalgae dominate shallow eutrophic estuaries. J Phycol 47:483–494
Khaw HL, Ponzoni RW, Danting JC (2008) Estimation of genetic change in the GIFT strain of Nile tilapia (Oreochromis niloticus) by comparing contemporary progeny produced by males born in 1991 or in 2003. Aquaculture 275:64–69
Kirst M, Myburg AA, De Leon JP, Kirst ME, Scott J, Sederoff R (2004) Coordinated genetic regulation of growth and lignin revealed by quantitative trait locus analysis of cDNA microarray data in an interspecific backcross of eucalyptus. Plant Physiol 135:2368–2378
Kraan S, Verges Tramullas A, Guiry MD (2000) The edible brown seaweed Alaria esculenta (Phaeophyceae, Laminariales): hybridization, growth and genetic comparisons of six Irish populations. J Appl Phycol 12:577–583
Lane CE, Mayes C, Druehl LD, Saunders GW (2006) A multi-gene molecular investigation of the kelp (Laminariales Phaeophyceae) supports substantial taxonomic re-organization. J Phycol 42:493–512
Li D, Wu C (2012) Genetic analysis of Laminaria japonica; new strains bred by crossing of different gametophyte clone lines. In: Borowitzka MA (ed) Asia Pacific Conference on Algal Biotechnology. Adelaide
Li XJ, Cong YZ, Yang GP, Shi YY, Qu SC, Li ZL, Wang GW, Zhang ZZ, Luo SJ, Dai HL, Xie JZ, Jiang GL, Liu JL, Wang TY (2007) Trait evaluation and trial cultivation of Dongfang No. 2, the hybrid of a male gametophyte clone of Laminaria longissima (Laminariales, Phaeophyta) and a female one of L-japonica. J Appl Phycol 19:139–151
Li XJ, Liu JL, Cong YZ, Qu SC, Zhang ZZ, Dai HL, Luo SJ, Han XB, Huang SS, Wang QY, Liang GJ, Sun J, Jin Y, Wang DQ, Yang GP (2008a) Breeding and trial cultivation of Dongfang No. 3, a hybrid of Laminaria gametophyte clones with a more than intraspecific but less than interspecific relationship. Aquaculture 280:76–80
Li XJ, Yang GP, Shi YY, Cong YZ, Che S, Qu SC, Li ZL (2008b) Prediction of the heterosis of Laminaria hybrids with the genetic distance between their parental gametophyte clones. J Appl Phycol 20:1097–1102
Loureiro RR, Reis RP, Berrogain FD, Critchley AT (2012) Extract powder from the brown alga Ascophyllum nodosum (Linnaeus) Le Jolis (AMPEP): a “vaccine-like” effect on Kappaphycus alvarezii (Doty) Doty ex P.C. Silva. J Appl Phycol 24:427–432
Lüning K, Kadel P, Pang SJ (2008) Control of reproduction rhythmicity by environmental and endogenous signals in Ulva pseudocurvata (Chlorophyta). J Phycol 44:866–873
Maine Seaweed Council (2002) Harvest guidelines for Maine seaweeds. M. S. Council, Brunswick
McCauley J, Winberg PC, Skropeta D (in press) Genetic and environmental influences on the lipid profiles of three Ulva spp. (Chlorophyta). J Appl Phycol
Meuwissen TH, Hayes BJ, Goddard ME (2001) Prediction of total genetic value using genome-wide dense marker maps. Genetics 157:1819–1829
Meuwissen THE, Sonesson AK (2004) Genotype-assisted optimum contribution selection to maximize selection response over a specified time period. Genet Res 84:109–116
Miura A (1975) Studies on the breeding of cultivated Porphyra (Rhodophyceae). Third International Ocean Development Conference (Aug. 5–8). Mar Res 3:81–93
Molis M, Korner J, Ko YW, Kim JH (2008) Specificity of inducible seaweed anti-herbivory defences depends on identity of macroalgae and herbivores. Mar Ecol Prog Ser 354:97–105
Monro K, Poore AGB (2005) Light quantity and quality induce shade-avoiding plasticity in a marine macroalga. J Evol Biol 18:426–435
Msuya FE (2011) The impact of seaweed farming on the socio-economic status of coastal communities in Zanzibar, Tanzania. World Aquacult 42:45–48
Nell JA, Cox E, Smith IR, Maguire GB (1994) Studies on triploid oysters in Australia.1. The farming potential of triploid Sydney rock oysters Saccostrea commercialis (Iredale and Roughley). Aquaculture 126:243–255
Niwa K, Iida S, Kato A, Kawai H, Kikuchi N, Kobiyama A, Aruga Y (2009) Genetic diversity and introgression in two cultivated species (Porphyra yezoensis and Porphyra tenera) and closely related wild species of Porphyra (Bangiales, Rhodophyta). J Phycol 45:493–502
Niwa K, Kikuchi N, Iwabuchi M, Aruga Y (2004) Morphological and AFLP variation of Porphyra yezoensis Ueda form. narawaensis Miura (Bangiales, Rhodophyta). Phycol Res 52:180–190
Niwa K, Kobiyama A, Sakamoto T (2010) Interspecific hybridization in the haploid blade-forming marine crop Porphyra (Bangiales Rhodophyta): occurrence of alloploidy in surviving F-1 gametophytic blades. J Phycol 46:693–702
Olesen I, Alfnes F, Rora MB, Kolstad K (2010) Eliciting consumers’ willingness to pay for organic and welfare-labelled salmon in a non-hypothetical choice experiment. Livest Sci 127:218–226
Patwary MU, Van Der Meer JP (1983) An apparent absence of heterosis in hybrids of Gracilaria tikvahiae (Rhodophyceae). Proc N S Inst Sci 33:95–99
Patwary MU, van der Meer JP (1984) Growth experiments on autopolyploids of Gracilaria tikvahiae (Rhodophyceae). Phycologia 23:21–27
Ponzoni RW, Nguyen N, Khaw HL, Ninh NH (2008) Accounting for genotype by environment interaction in economic appraisal of genetic improvement programs in common carp Cyprinus carpio. Aquaculture 285:47–55
Qin S, Jiang P, Tseng CK (2004) Molecular biotechnology of marine algae in China. Hydrobiologia 512:21–26
Raja R, Hemaiswarya S, Kumar NA, Sridhar S, Rengasamy R (2008) A perspective on the biotechnological potential of microalgae. Crit Rev Microbiol 34:77–88
Reddy C, Gupta M, Mantri V, Jha B (2008a) Seaweed protoplasts: status, biotechnological perspectives and needs. J Appl Phycol 20:619–632
Reddy C, Jha B, Fujita Y, Ohno M (2008b) Seaweed micropropagation techniques and their potentials: an overview. J Appl Phycol 20:609–617
Rioux LE, Turgeon SL, Beaulieu M (2009) Effect of season on the composition of bioactive polysaccharides from the brown seaweed Saccharina longicruris. Phytochemistry 70:1069–1075
Robic A, Bertrand D, Sassi JF, Lerat Y, Lahaye M (2009) Determination of the chemical composition of ulvan, a cell wall polysaccharide from Ulva spp. (Ulvales, Chlorophyta) by FT-IR and chemometrics. J Appl Phycol 21:451–456
Robinson N, Li X, Hayes B (2010a) Testing options for commercialisation of abalone selective breeding using bioeconomic simulation modelling. Aquacult Res 41:e268–e288
Robinson NA, Schipp G, Bosmans J, Jerry DR (2010b) Modelling selective breeding in protandrous, batch-reared Asian sea bass (Lates calcarifer, Bloch) using walkback selection. Aquacult Res 41:e643–e655
Roleda MY, Wiencke C, Hanelt D, Bischof K (2007) Sensitivity of the early life stages of macroalgae from the Northern Hemisphere to ultraviolet radiation. Photochem Photobiol 83:851–862
Smit AJ, Robertson-Andersson DV, Peall S, Bolton JJ (2007) Dimethylsulfoniopropionate (DMSP) accumulation in abalone Haliotis midae (Mollusca: Prosobranchia) after consumption of various diets, and consequences for aquaculture. Aquaculture 269:377–389
Steinbrenner J, Sandmann G (2006) Transformation of the green alga Haematococcus pluvialis with a phytoene desaturase for accelerated astaxanthin biosynthesis. Appl Environ Microbiol 72:7477–7484
Stengel DB, Connan S, Popper ZA (2011) Algal chemodiversity and bioactivity: sources of natural variability and implications for commercial application. Biotechnol Adv 29:483–501
Sutherland JE, Lindstrom SC, Nelson WA, Brodie J, Lynch MDJ, Hwang MS, Choi H-G, Miyata M, Kikuchi N, Oliviera MC, Farr TJ, Neefus CD, Mols-Mortensen A, Milstein D, Muller KM (2011) A new look at an ancient order: generic revision of the Bangiales (Rhodophyta). J Phycol 47:1131–1151
Tapsell LC (2008) Evidence for health claims: a perspective from the Australia–New Zealand region. J Nutr 138:1206S–1209S
Temussi PA (2012) The good taste of peptides. J Pept Sci 18:73–82
Thodesen J, Grisdale-Helland B, Helland SJ, Gjerde B (1999) Feed intake, growth and feed utilization of offspring from wild and selected Atlantic salmon (Salmo salar). Aquaculture 180:237–246
Van Der Meer JP (1990) Genetics. In: Cole KM, Sheath RG (eds) Biology of the red algae. Cambridge University Press, Cambridge, pp 103–120
Vasquez JA (2011) Production, use and fate of Chilean brown seaweeds: re-sources for a sustainable fishery. J Appl Phycol 20:7–17
Wang JF, Jiang P, Cui YL, Deng XY, Li FC, Liu JG, Qin S (2010) Genetic transformation in Kappaphycus alvarezii using micro-particle bombardment: a potential strategy for germplasm improvement. Aquacult Int 18:1027–1034
Wichard T, Oertel W (2010) Gametogenesis and gamete release of Ulva mutabilis and Ulva lactuca (Chlorophyta): regulatory effects and chemical characterisation of the “swarming inhibitor”. J Phycol 46:248–259
Wright JT, de Nys R, Poore AGB, Steinberg PD (2004) Chemical defense in a marine alga: heritability and the potential for selection by herbivores. Ecology 85:2946–2959
Zamora I, Feldman JL, Marshall WF (2004) PCR-based assay for mating type and diploidy in Chlamydomonas. Biotechniques 37:534–536
Zhang BL, Yan XH, Huang LB (2011a) Evaluation of an improved strain of Porphyra yezoensis Ueda (Bangiales, Rhodophyta) with high-temperature tolerance. J Appl Phycol 23:841–847
Zhang T, Shen Z, Xu P, Zhu J, Lu Q, Shen Y, Wang Y, Yao C, Li J, Wang Y, Jiang H (2011b) Analysis of photosynthetic pigments and chlorophyll fluorescence characteristics of different strains of Porphyra yezoensis. J Appl Phycol 24:881–886
Zhang XC, Van Der Meer JP (2012) A study of heterosis in diploid gametopytes of the marine red alga Gracilaria tikvahiae. Bot Mar 30:309–314
Zitta C, Oliveira E, Bouzon Z, Hayashi L (2011) Ploidy determination of three Kappaphycus alvarezii strains (Rhodophyta, Gigartinales) by confocal fluorescence microscopy. J Appl Phycol 24:495–499
Acknowledgments
Thanks to Trygve Gjedrem for his helpful suggestions about the manuscript. Thanks also to the Rural Industries Research and Development Corporation in Australia for funding the project and to our project collaborators at Marinova.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Robinson, N., Winberg, P. & Kirkendale, L. Genetic improvement of macroalgae: status to date and needs for the future. J Appl Phycol 25, 703–716 (2013). https://doi.org/10.1007/s10811-012-9950-x
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10811-012-9950-x