
Abstract
There are several conflicting hypothesis that deal with the influence of flooding in the natural river–floodplain systems. According to the Flood Pulse Concept, the flood pulses are not considered to be a disturbance, while some recent studies have proven that floods can be a disturbance factor of phytoplankton development. In order to test whether flooding acts as a disturbance factor in the shallow Danubian floodplain lake (Lake Sakadaš), phytoplankton dynamics was investigated during two different hydrological years—extremely dry (2003) without flooding and usually flooded (2004). A total of 18 phytoplankton functional groups were established. The sequence of phytoplankton seasonality can be summarized P/D → E (W1, W2) → C/P (only in potamophase) → S2/H1/SN/S1 → W1/W2 → P/D. The canonical correspondence analysis (CCA) demonstrated that the water level was a significant environmental variable in 2004. Due to the higher total biomass of Bacillariophyceae established under potamophase conditions, floodings in the early spring seem to be a stimulating factor for phytoplankton development. On the other hand, the flood pulses in May and June had dilution effects on nutrients, so that a significantly lower phytoplankton biomass was established indicating that flooding pulses can be regarded as a disturbance event. Such conditions supported diatom development (D, P, C species) and prolonged its dominance in the total phytoplankton biomass. A long-lasting Cyanoprokaryota bloom (various filamentous species—S1, S2, SN and H1 representatives) with very high biomass characterized the limnophase (dry conditions) in summer and autumn of both years. In-lake variables (lake morphology, internal loadings of nutrients from sediments, light conditions) seem to be important for the appearance of Cyanoprokaryota bloom. The equilibrium phase was found during the Cyanoprokaryota bloom only in the extremely dry year. This study showed that depending on the time scale occurrence, flood pulses can be a stimulating or a disturbance factor for phytoplankton development in Lake Sakadaš.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Knowledge of floodplains has increased considerably since the flood pulse concept (FPC) was published (Junk et al., 1989). This concept promulgated the view that rivers and their fringing floodplains are integrated components of a single dynamical system, linked by strong interactions between hydrological and ecological processes. The major driving force is the pulsing of river discharge, which determines the connectivity and fluctuation processes of matter and organisms across river-floodplain gradients. However, according to a review of the present state and future trends of riverine floodplains (Tockner & Standford, 2002), there are many remaining questions about floodplain ecology that need to be investigated. Long-term trends in floodplain populations and the complexity and interdependence of processes that occur when river or lake water inundates a floodplain area are not yet fully understood. Currently, questions arise about the importance of landscape connectivity and the dynamics of flooding on biodiversity (Junk & Wantzen, 2003).
Floodplain lakes are the most common lentic water bodies in the tropics, in contrast to temperate latitudes where lakes are predominately of glacial origin (Lewis, 2000). Many recent studies have been undertaken to determine the responses of phytoplankton to water fluctuation in floodplain lakes of tropical rivers: Amazon (Ibañez, 1998), Paraguay (de Oliveira & Calheiros, 2000), Araguaia (Nabout et al., 2006), Mary (Townsend, 2006), Murray (Butler et al., 2007), and Parana (Zalocar de Domitrovic, 2003; Izaguirre et al., 2001). Results indicate that phytoplankton dynamics are hydrology driven, and that flood pulse influences the composition and population densities of the phytoplankton communities.
The timing of flooding and the total inundated area of the floodplain at high and low water varies in both temperate and tropical floodplains. In the South American wetlands, the flood pulse characterizes distinct low water (limnophase) and high water (potamophase) periods (Zalocar de Domitrovic, 2003), while the wetlands of South Asia are strongly influenced by seasonal monsoons and the water cycle can be divided into four phases (Muzaffar & Ahmed, 2007). Wetlands in arid and semi-arid regions, such as Mediterranean Europe, are faced with frequent droughts and low rainfall, unevenly distributed in space and time (Angeler et al., 2000). Many of the remaining European floodplains in temperate regions are far from being pristine. Floodplain areas have been separated by levees and have therefore become functionally extinct, since regular flooding and morphological dynamics are missing. Floodplains along the Oder River, the Danube River, or along Eastern European river corridors are impacted systems that retain some semblance of natural functions (Tockner & Standford, 2002).
European riverine wetlands have largely remained outside the focus of scientific investigations. There have been some recent studies of phytoplankton in floodplain areas along the Danube River, e.g., side-arms near Wiena (Riedler et al., 2006), floodplains along the Bulgarian section of the Danube (Stoyneva, 1998, 2003), Danube Delta (Török, 2006). However, a very important and fundamental question remains—to explain the occurrence of species and their communities in river-floodplain systems (Henle et al., 2006). The body of knowledge about structural changes of phytoplankton in deep and shallow lakes may apply less to wetlands (Padisák et al., 2003) where a different, weaker pattern has been established (Rojo et al., 2000).
The aim of this investigation is to contribute to an understanding of how changes in hydrological connectivity and habitat fragmentation affect phytoplankton assemblages, particularly the appearance of phytoplankton equilibrium in the shallow Lake Sakadaš, a part of the Danubian floodplain. For this reason, the study compares seasonal changes of phytoplankton assemblages in two hydrologically different years—usually a flood year and an extremely dry year.
Materials and methods
Study area
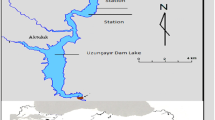
Nature Park Kopački Rit, a Ramsar site (No: 3HR002), is a fluvial floodplain along the Danube River (1410–1383 r. km), located in Northeast Croatia (Fig. 1). It is one of the largest riverine wetlands in Europe, covering more then 23 km2. The dominant wetland types in the floodplain are permanent freshwater marshes/pools, ponds (their surface areas <0.08 km2), marshes, and swamps on organic soils, with emergent vegetation water-logged for most of the growing season. The shallow lakes and river side arms are very significant sub-systems of the floodplain complex, determined by their interaction with Danube flooding. During periods of high water levels, usually in spring and early summer, there is an inflow of Danube water into the floodplain area. During low water conditions, floodplain water subsystems are isolated from one another. Thus, the highly variable hydrology results in fluctuating hydrological connectivity patterns and hydroperiods. Lake Sakadaš is located in the western part of the floodplain and through a system of natural channels is in direct hydrological connection with the Danube River (Fig. 1). During normal water conditions, the lake has an average depth of about 5 meters (at the deepest depression in the floodplain) and an area of about 0.15 km2. The lakeshore is infested with Phragmites, adjoined by floodplain forests of white willows.

Study area: Lake Sakadaš-Nature Park Kopački Rit (M 1:200 000)
Limnological methods
Water samples were collected from the central part of Lake Sakadaš at monthly intervals during the ice-free season (March through November) of 2003 and 2004. Water samples were taken just beneath the surface of the lake and close to the sediments, using the Ruttner bottle. Chemical variables were measured according to APHA (1985). Chlorophyll concentration was estimated according to Komárková (1989). Integrated phytoplankton samples were collected from the whole water column and fixed with Lugol`s solution with acetic acid. Phytoplankton species were identified by light microscopic observations (Carl Zeiss Jena). Taxonomic monographs of cyanophytes were used to supplement the standard literature for species determination. Quantitative assessment of phytoplankton was done by counting the cells settled in Utermöhl chambers (Utermöhl, 1958). The abundance of each species is presented as the number of individuals per liter (ind. l−1). To estimate biovolumes of the algae and cyanoprokaryotes, individual cells were measured and their volumes calculated according to geometrical solids (Rott, 1981). Biovolume was converted to biomass (Javornický & Komárková, 1973; Sournia, 1978) and expressed as mg l−1 fresh mass (FM). Functional groups of phytoplankton were defined according to Reynolds et al. (2002). Dominant species was estimated from percentage contribution of individual species to total biomass. Only those species which had a minimum of 5% contribution to total biomass were considered to be the dominants (Padisák et al., 2003).
Relationships between biological assemblages of species and their environment were analyzed with Canonical correspondence analysis (CCA) using the program CANOCO 4.5 (ter Braak & Šmilauer, 2002). The significance with which environmental variables explain the variance of species data was tested using Monte Carlo simulations with 99 unrestricted permutations. Variables were considered to be significant when P < 0.05. A total of 13 limnological variables were taken into consideration: nitrates (NO3), nitrites (NO2), total nitrogen (TN), total phosphorous (TP), ortho-phosphates (PO4), dissolved oxygen—surface (DO-surface), dissolved oxygen—bottom (DO-bottom), pH, water temperature (WT), Secchi depth (SD), water depth (WD), water level (WL), and FM.
Results
Environmental characteristics
Annual courses of the Danube water level are shown in Fig. 2. Flooding of the floodplain area of Kopački Rit began when the Danube water level rose above 3 m (Mihaljević et al., 1999). It is evident that extreme dry conditions (long-lasting limnophase) characterized the hydrological conditions during the whole of 2003. There were only three brief floods lasting no more than a few days in January, March, and October. In spite of that, the usual fluctuation of dry (limnophase) and flood periods (potamophase) characterized the hydrological conditions in 2004—flooding in January, February, April, June, and the beginning of July were followed by dry periods until the end of the year. Values of the main physical and chemical parameters of Lake Sakadaš during the study are shown in Table 1. Due to the frequent incoming of Danube water during the flooding in 2004, the mean value of water depth, as well as water temperature, was higher in comparison with 2003. The mean value of Secchi depth was lower during the long-term dry periods in 2003 than in 2004. Lower DO and anoxic conditions were registered during the characteristic regular fluctuation of dry and flood periods (2004), and not in conditions of hydrological isolation of the lake. High concentrations of TP (mean value 265.8 μg l−1 in 2003 and 129.3 μg l−1 in 2004) and TN (mean value 856.3 μg l−1 in 2003 and 1190.0 μg l−1 in 2004) were found. Low values of TN/TP (0.8–7.9) were established during the whole investigated period in 2003, while the TN/TP values were in the range 3.7–20.7 during 2004. The system supported mean Chla concentrations of 111.9 μg l−1, closely associated with Secchi depth. Total phytoplankton biomass was significantly correlated with Chla (r = 0.65, P < 0.05 in 2003; r = 0.72, P < 0.05 in 2004).

Annual courses of the Danube water level at r. km 1401.4
Phytoplankton characterization
A total of 70 phytoplankton taxa were detected during counting, among which 29 taxa were registered as having achieved a biomass higher than 5% (Table 2). The mean phytoplankton abundance was high (89.3 × 106 ind. l−1 in 2003 and 57.9 × 106 ind. l−1 in 2004) with cyanoprokaryotes and diatoms numerically dominant. The annual changes in phytoplankton biomass were in the range 45.9–274.9 mg l−1 in 2003 and 12.6–255.8 mg l−1 in 2004 (Fig. 3). Remarkably large summer maxima occurred earlier (July) during the dry period in 2003 rather than after the spring and early summer flood period (until September) in 2004 (Fig. 3). The total phytoplankton biomass was significantly correlated with phytoplankton abundance (r = 0.86, P < 0.05 in 2003; r = 0.77, P < 0.05 in 2004).

The annual changes in phytoplankton abundance and biomass in Lake Sakadaš
Canonical correspondence analysis (CCA) was initially performed on the environmental and all species datasets, for each year separately. Figure 4 shows the distribution of the taxa with minimum fit = 5%. The eigenvalues for CCA axis 1 (0.60) and axis 2 (0.46) explained 54.9% of the variance in the species in 2003. In 2004, eigenvalues for CCA axis 1 (0.63) and axis 2 (0.50) explained 63.9% of the variance in the species. Species-environment correlations for CCA axis 1 (0.94 in 2003; 0.95 in 2004) and axis 2 (0.92 in 2003; 0.92 in 2004) were high in both years, indicating a significant relationship between the 12 environmental variables and the 70 phytoplankton taxa selected.

In unusual hydrological conditions, e.g., in the extremely dry year of 2003, most of the variance contained in the first CCA axis is described by three environmental parameters—water temperature (r1CCA = −0.77, P < 0.05), TP (r 1CCA = −0.73, P < 0.05), and Secchi depth (r 1CCA = 0.69, P < 0.05). In the second CCA axis most of the variance is explained by the water level (r 2CCA = 0.88, P < 0.05), TN (r 2CCA = 0.80, P < 0.05), and PO4 (r 2CCA = −0.62, P < 0.05). Early spring phytoplankton development in 2003 (March) started with the dominance of Bacillariophyceae (64.8% of total FM). The well-developed populations in March were representatives from functional groups P (Aulacoseira granulata, Fragilaria capucina) and D (Stephanodiscus hantzschii). In April, Bacillariophyceae fell to 26.5% of total FM, while representatives from W1 (euglenoids), W2 (Peridinium aciculiferum), and E (Dinobryon divergens) were abundant. Then, from May until the end of the ice-free season (November), phytoplankton was dominated by cyanoprokaryotes. Their contributions in total FM reached up to more than 80% in the period from June to September (Fig. 5). Cyanoprokaryota bloom achieved a peak biomass of 246.5 mg l−1 in July (Fig. 3). Different filamentous species (Fig. 5) from several functional groups were replaced during the Cyanoprokaryota bloom: Planktolyngbya limnetica (S2) in June; dinitrogen-fixing nostocaleans (H1) with dominance of Aphanizomenon flos-aquae in July; Cylindrospermopsis raciborskii plankton (SN) with complement of Planktothrix agardhii (S1) in August and September; Pseudanabaena limnetica and Planktothrix agardhii (S1) in October and November. Bacillariophyceae (Stephanodiscus hantzschii—D group), Chlorococcales (Scenedesmus sp. div.—J group) and Euglenophyceae (W1 group) were additions to the autumnal composition of phytoplankton.

Share of Cyanoprokaryota species biomass on the total phytoplankton biomass in the Lake Sakadaš (see Table 2 for species code)
In usual hydrological conditions in 2004, most of the variance contained in the first CCA axis is described by the following environmental parameters—Secchi depth (r 1CCA = 0.88, P < 0.05), water level (r 1CCA = 0.61, P < 0.05), and PO4 (r 1CCA = −0.72, P < 0.05). In the second CCA axis most of the variance is explained by the water temperature (r2CCA = −0.64, P < 0.05) and NO2 (r2CCA = 0.62, P < 0.05).
At the beginning of spring, Bacillariophyceae also characterized the phytoplankton composition (Fig. 3). In March and April, they contributed 93.8% of total FM. Stephanodiscus hantzschii (D group) was the most abundant, while species from P functional group (Aulacoseira granulata and Fragilaria crotonensis) were abundant. At the end of May, Dinobryon divergens (E) was dominant. Through survival of potamophase, Bacillariophyceae, representatives from the functional groups C (Asterionella formosa) and P (Fragilaria crotonensis), were recurrent in June and were characterized by high water level (Fig. 4), e.g., lake flooding. In limnophase conditions, from July through October 2004, Cyanoprokaryota bloom developed and achieved a peak biomass of 145.8 mg l−1 in September (Fig. 3). “Clear assemblage” bloom composed of Limnothrix redekei, Planktothrix agardhii, and Pseudanabaena limnetica, representatives of S1 group, occurred during the cyanoprokaryotes bloom with the supplement of Planktolyngbya limnetica (S2) in July and Cylindrospermopsis plankton (SN) in August (Fig. 5). The characterization of cyanoprokaryotes by phosphate is evident from CCA (Fig. 4). In autumnal conditions (September and October) the contribution of cyanoprokaryotes decreased by ca. 50% of total FM, while the representatives of functional group W2 (Peridinium aciculiferum) with supplement of W1 (Euglenophyceae), P (Aulacoseira granulata), and D (Stephanodiscus hantzschii) were abundant.
Discussion
Due to the landscape position and geomorphology of Lake Sakadaš, flooding is an important water source for structuring its habitat (Mihaljević et al., 1999). The obtained results indicated that floodwater had altered the physical and chemical environment of Lake Sakadaš (Table 1). The high levels of nutrients (especially TP) established during the limnophase indicated that the river water did not serve as a major source of nutrients as is characteristic for tropical floodplains (Nabout et al., 2006; Weilhoefer and Pan, 2007). Thus, the nutrient enrichment during potamophase can be considered as the result of the in-lake processes due to the natural input and accumulation of organic matter from terrestrial vegetation in the river-floodplain system, generally recognized as “zones of storage and turnover of organic matter” (Keckeis et al., 2003; Pithart et al., 2007). The appearance of dissolved oxygen demand during the usually flooded year confirmed strong organic matter degradation and interface nitrogen transformation (McCarthy et al., 2008) consequently expressed as high NO2 concentrations.
In the annual courses of phytoplankton development there was a dominance of Bacillariophyceae in the early spring, which is a mutual characteristic regardless of hydrological conditions (March) and can be considered a natural phase of early spring phytoplankton development (PEG-model, Sommer, 1989). Higher total biomass of Bacillariophyceae under potamophase conditions contributed to the fact that floodings in the early spring seem to be a stimulating factor rather than a disturbance (Paidere et al., 2007) for phytoplankton development.
Under conditions of prolonged spring floods (in May and June, 2004), the dilution effects were expressed (higher Secchi depth and lower amounts of nutrients) and as a result, there was a significantly lower (ca. 400%) mean phytoplankton biomass compared to the limnophase in the previous year (Table 1, Fig. 4). Similarly, a decrease in phytoplankton biomass was established in the Danube floodplain near Vienna under conditions of high connectivity with the Danube (Keckeis et al., 2003). Thus, flooding in the late spring and at the beginning of summer seems to be a disturbance factor (Reynolds et al., 1993) in the seasonal development of phytoplankton, which is in line with many other studies where floods are considered as a disturbance (Tockner et al., 1999; Hein et al., 1999; Roozen et al., 2003). Due to the turbulence caused by the inflow of diatom-rich Danube River waters, (Riedler et al., 2006), this period was characterized by the dominance of Bacillariophyceae, representatives from D, P, and C functional groups. Although the Stephanodiscus hantzschii and Fragilaria (D representatives) are considered typical, constant Danube phytoplankton species (Schmidt, 1992; Török, 2006), the given results indicated that the limnological conditions in Lake Sakadaš were favorable for the development of this group, tolerant of the overall water conditions (Reynolds et al., 2002).
The long-term dominance of Cyanoprokaryota, with the main protagonists being various filamentous species (representatives of S1, H1, SN, and S2 groups), was a general characteristic of the limnophase in both years of investigation. The high trophic level (eutrophic-hypertrophic) of Lake Sakadaš, established according to its assessment by traditional methods (due to the concentrations of Chla, TP, and Secchi depth), seems to be one of a multiplicity of factors (Dokulil & Teubner, 2000) favoring the bloom of cyanoprokaryotes. Also, Cyanoprokaryota bloom was established under hypertrophic conditions of the wetlands along the Bulgarian section of the Danube River (Stoyneva, 2003). Seasonal sequences of functional groups appeared as H1 → SN → S1 during the Cyanoprokaryota bloom in 2003. This is recognizable as a common succession due to the fact that species belonging to these groups are frequently in competition (Padisák et al., 2003). The S1 “clear assemblage,” which frequently appears in shallow, eutrophic lakes (Nixdorf et al., 2003), characterized the bloom in 2004. Representatives of the S1 group have high nitrogen affinity (Mischke & Nixdorf, 2003) so that nitrites were significant in its characterization (Fig. 4).
Hypertrophic conditions of Lake Sakadaš are connected with low-light conditions, in which Cyanoprokaryotes of the SN, S1 and S2 groups have proven to be successful (O’Farrell et al., 2007; de Tezanos Pinto et al., 2007). Thus, the additional decrease in underwater light availability in Lake Sakadaš caused by the shadowing of well-developed floating vegetation (Lemna sp. div., Spirodella polyrrhiza) in 2004, can be responsible (Cattaneo et al., 1998) for the absence of dinitrogen-fixing Nostocaleans (H1), sensitive to light-deficient conditions (Reynolds et al., 2002).
Following the definition offered by Sommer et al. (1993), an equilibrium phase (coexistence of only three species contributing more than 80% of the total biomass with no significant changes in biomass) was established in the period June–September 2003 (Fig. 5), under conditions of an extremely long-lasting potamophase. The established dominant species, Aphanizomenon flos-aquae and Cylindrospermopsis raciborskii, are known as the dominant species of a relatively stationary state in shallow eutrophic lakes (Naselli-Flores et al., 2003), as well as floodplain lakes (Dokulil & Taubner, 2000; Stoyneva, 2003; Borics et al., 2000; Rojo & Álvarez-Cobelas, 2003). However, in the potamophase, which followed the long-lasting limnophase in 2004, the equilibrium phase was not established because of the expected “time lag” between the floods and the time at which the effects of homogenization become apparent (Thomaz et al., 2007). Furthermore, flooding has a destabilizing effect on plankton food web dynamics in wetlands (Margalef, 1997; Keckeis et al., 2003).
The decreasing ambient temperature in the late autumn conditions, as well as the high organic matter content that resulted due to the collapse of Cyanoprokaryota bloom, supported the development of representatives of functional groups W2 (Trachelomonas, Peridinium aciculiferum) and W1 (Euglenophyceae). Temperature and organic matter content were recognized by Grigorszky et al. (2006) as the most important factors controlling the distribution of Dinophyta species. Previous investigations (Mihaljević & Novoselić, 2000; Horvatić et al., 2003; Stević et al., 2005) report a frequent appearance of the bloom of Peridinium in Lake Sakadaš during the late autumn. Euglenoids, known as tolerant on low values of DO (de Oliveira & Calheiros, 2000), usually coincide with hypoxic conditions in Lake Sakadaš. Furthermore, it is interesting to note that there was no development of the Synura bloom which usually appears in late autumn and winter periods in Lake Sakadaš.
The obtained results of the investigation of phytoplankton in Lake Sakadaš during two hydrologically different years, supports a substantial influence of flood dynamics on the limnological characteristics of the lake, and consequently on phytoplankton dynamics. The long-lasting potamophase supported diatom development (D, P, C species) and prolonged its dominance in the total phytoplankton biomass until the beginning of the limnophase (dry conditions). The stable environment during the limnophase led to high biomass phytoplankton assemblages, dominated by cyanoprokaryotes (H1, SN, and S1 species), and forced phytoplankton communities toward equilibrium. A long-term limnological investigation of phytoplankton is necessary to completely elucidate the interactions between the Danube River and the fringe floodplain of Kopački Rit.
References
Angeler, D. G., M. Alvarez-Cobelas, C. Rojo & S. Sánchez-Carrillo, 2000. The significance of water inputs to plankton biomass and trophic relationships in a semiarid freshwater wetland (central Spain). Journal of Plankton Research 22: 2075–2093.
APHA, 1985. American Public Health Association Standard Methods for the Examination of Water and Wastewaters. Washington, DC, USA.
Borics, G., I. Grigorszky, S. Szabó & J. Padisák, 2000. Phytoplankton associations in a small hypertrophic fishpond in East Hungary during a change from bottom–up to top–down control. Hydrobiologia 424: 79–90.
Butler, J., R. Croome & G. N. Rees, 2007. The composition and importance of the phytoneuston in two floodplain lakes in south-eastern Australia. Hydrobiologia 579: 135–145.
Cattaneo, A., G. Galanti, S. Gentinetta & S. Romo, 1998. Epiphytic algae and macroinvertebrates on submerged and floating-leaved macrophytes in an Italian lake. Freshwater Biology 39: 725–740.
de Oliveira, M. D. & D. F. Calheiros, 2000. Flood pulse influence on phytoplankton communities of the south Pantanal floodplain, Brazil. Hydrobiologia 427: 101–112.
de Tezanos Pinto, P., L. Allende & I. O’Farrell, 2007. Influence of free-floating plants on the structure of a natural phytoplankton assemblage: an experimental approach. Journal of Plankton Research 29: 47–56.
Dokulil, M. T. & K. Teubner, 2000. Cyanobacterial dominance in lakes. Hydrobiologia 438: 1–12.
Grigorszky, I., K. T. Kiss, V. Béres, I. Bácsi, M. M-Hamvas, C. Máthé, G. Vasas, J. Padisák, G. Borics, M. Gligora & G. Borbély, 2006. The effects of temperature, nitrogen, and phosphorus on the encystment of Peridinium cinctum, Stein (Dinophyta). Hydrobiologia 563: 527–535.
Hein, T., C. Baranyi, G. Heiler, C. Holarek, P. Riedler & F. Schiemer, 1999. Hydrology as a major factor determining plankton development in two floodplain segments and the River Danube, Austria. Archiv für Hydrobiologie Supplement 115: 439–452.
Henle, K., M. Scholz, F. Dziock, S. Stab & F. Foeckler, 2006. Bioindication and functional response in floodplain systems: Where to from here? International Review of Hydrobiology 91: 380–387.
Horvatić, J., M. Mihaljević & F. Stević, 2003. Algal growth potential of Chlorella kessleri FOTT et NOV. in comparison with in situ microphytoplankton dynamics in the water of Lake Sakadaš marshes. Periodicum Biologorum 105: 307–312.
Ibañez, M. S. R., 1998. Phytoplankton composition and abundance of a central Amazonian floodplain lake. Hydrobiologia 362: 79–83.
Izaguirre, I., I. O’Farrell & G. Tell, 2001. Variation in phytoplankton composition and limnological features in a water-water ecotone of the Lower Paraná Basin (Argentina). Freshwater Biology 46: 63–74.
Javornický, P. & J. Komárková, 1973. The changes in several parameters of plankton primary productivity in Slapy Reservoir 1960–1967, their mutual correlations and correlations with the main ecological factors. In Hrbáček, J. & M. Straškraba (eds), Hydrobiological Studies 2. Academia, Prague: 155–211.
Junk, W. J., P. B. Bayley & R. E. Sparks, 1989. The flood pulse concept in river floodplain system. Canadian Special Publication of Fisheries Aquatic Sciences 106: 110–127.
Junk, W. J. & K. M. Wantzen, 2003. The flood pulse concept: New aspects, approaches, and applications—an update. In Welcomme R. L. & T. Petr (eds), Proceedings of the Second International Symposium on the Management of Large Rivers for Fisheries, Vol. 2. Food and Agriculture Organization & Mekong River Commission. FAO Regional Office for Asia and the Pacific, Bangkok. RAP Publication 2004/17.
Keckeis, S., C. Baranyi, T. Hein, C. Holarek, P. Riedler & F. Schiemer, 2003. The significance of zooplankton grazing in a floodplain system of the River Danube. Journal of Plankton Research 25: 243–253.
Komárková, J., 1989. Primárni produkce ř as ve slatkovodních ekosysteméch. In Dykyová, D. (ed.), Metody studia ecosystémů. Academia Praha, Praha: 330–347.
Lewis, W. M., 2000. Basis for the protection and management of tropical lakes. Lakes & Reservoirs Research and Management 5: 34–48.
Margalef, R., 1997. Our Biosphere. Excellence in Ecology 10. Ecology Institute, Oldenburg/Luhe, Germany.
McCarthy, M. J., K. S. McNeal, J. W. Morse & W. S. Gardner, 2008. Bottom-water hypoxia effects on sediment–water interface nitrogen transformations in a seasonally hypoxic, shallow bay (Corpus Christi Bay, TX, USA). Estuaries and Coasts 31: 521–531.
Mihaljević, M., D. Getz, Z. Tadić, B. Živanović, D. Gucunski, J. Topić, I. Kalinović & J. Mikuska, 1999. Kopački Rit—Research Survey and Bibliography. Croatian Academy of Arts and Sciences, Zagreb.
Mihaljević, M. & D. Novoselić, 2000. Trophic state of Lake Sakadaš (Nature Reserve Kopački rit) in the late autumnal and winter period of 1997/98. Periodicum Biologorum 102: 253–257.
Mischke, U. & B. Nixdorf, 2003. Equilibrium phase conditions in shallow German lakes: how Cyanoprokaryota species establish a steady state phase in late summer. Hydrobiologia 502: 123–132.
Muzaffar, S. B. & F. A. Ahmed, 2007. The effects of the flood cycle on the diversity and composition of the phytoplankton community of a seasonally flooded Ramsar wetland in Bangladesh. Wetlands Ecology and Management 15: 81–93.
Nabout, J. C., I. S. Noguera & L. G. Oliviera, 2006. Phytoplankton community of floodplain lakes of the Araguaia River, Brazil, in the rainy and dry seasons. Journal of Plankton Research 28: 181–193.
Naselli-Flores, L., J. Padisák, M. T. Dokulil & I. Chorus, 2003. Equilibrium/steady-state concept in phytoplankton ecology. Hydrobiologia 502: 395–403.
Nixdorf, B., U. Mischke & J. Rücker, 2003. Phytoplankton assemblages and steady state in deep and shallow eutrophic lakes–an approach to differentiate the habitat properties of Oscillatoriales. Hydrobiologia 502: 111–121.
O’Farrell, I., P. d. T. Pinto & I. Izaguirre, 2007. Phytoplankton morphological response to the underwater light conditions in a vegetated wetland. Hydrobiologia 578: 65–77.
Padisák, J., G. Borics, G. Fehér, I. Grigorszky, I. Oldal, A. Schmidt & Z. Zámbóné-Doma, 2003. Dominant species, functional assemblages and frequency of equilibrium phases in late summer phytoplankton assemblages in Hungarian small shallow lakes. Hydrobiologia 502: 157–168.
Paidere, J., D. Gruberts, A. Škute & I. Druvietis, 2007. Impact of two different flood pulses on planktonic communities of the largest floodplain lakes of the Daugava River (Latvia). Hydrobiologia 592: 303–314.
Pithart, D., R. Pichlová, M. Bílý, J. Hrbáček, K. Novotná & L. Pechar, 2007. Spatial and temporal diversity of small shallow watersin river Lužnice floodplain. Hydrobiologia 584: 256–275.
Reynolds, C. S., V. Huszar, C. Kruk, L. Naselli-Flores & S. Melo, 2002. Towards a functional classification of the freshwater phytoplankton. Journal of Plankton Research 24: 417–428.
Reynolds, C. S., J. Padisák & U. Sommer, 1993. Intermediate disturbance in the ecology of phytoplankton and the maintenance of species diversity: a synthesis. Hydrobiologia 249: 183–188.
Riedler, P., C. Baranyi, T. Hein, S. Keckeis & M. Schagerl, 2006. Abiotic and biotic control of phytoplankton development in dynamic side-arms of the River Danube. Archiv für Hydrobiologie Supplement 158/4. Large Rivers 16: 577–594.
Rojo, C. & M. Álvarez-Cobelas, 2003. Are there steady-state phytoplankton assemblages in the field? Hydrobiologia 502: 3–12.
Rojo, C., E. Ortega-Mayagoitia & M. Álvarez-Cobelas, 2000. Lack of pattern among phytoplankton assemblages. Or, what does the exception to the rule mean? Hydrobiologia 424: 133–139.
Roozen, F. C. J. M., G. J. Geest, B. W. Ibelings, R. Roijackers, M. Scheffer & A. D. Buijse, 2003. Lake age and water level affect the turbidity of floodplain lakes along the lower Rhine. Freshwater Biology 48: 519–531.
Rott, E., 1981. Some results from phytoplankton counting intercalibrations. Swiss Journal of Hydrology 43: 34–62.
Schmidt, A., 1992. Das Phytoplankton, das Phytobenthos und die Makrophyten der Donau. Wien, Limnologische Berichte Donau 77–101.
Sommer, U., 1989. Toward a Darwinian ecology of plankton. In Sommer, U. (ed.), Plankton Ecology. Springer-Verlag, Berlin, Heildelberg, New York, London, Paris, Tokyo: 1–8.
Sommer, U., J. Padisák, C. S. Reynolds & P. Juhász-Nagy, 1993. Hutchinson’s heritage: the diversity-disturbance relationship in phytoplankton. Hydrobiologia 249: 1–7.
Sournia, A. (ed.), 1978. Phytoplankton Manual. UNESCO, Paris: 337.
Stević, F., M. Mihaljević & J. Horvatić, 2005. Interactions between microphytoplankton of the Danube, its sidearms and wetlands (1426–1388 r. km, Croatia). Periodicum biologorum 107: 299–304.
Stoyneva, M. P., 1998. Development of the phytoplankton of the shallow Srebarna Lake (North-Eastern Bulgaria) across a trophic gradient. Hydrobiologia 369(370): 259–267.
Stoyneva, M. P., 2003. Steady-state phytoplankton assemblages in shallow Bulgarian wetlands. Hydrobiologia 502: 169–176.
ter Braak, C. J. F. & P. Šmilauer, 2002. CANOCO reference manual and CanoDraw for Windows user’s guide: Software for canonical community ordination (version 4.5). Microcomputer Power (Ithaca, NY, USA).
Thomaz, S. M., L. M. Bini & R. L. Bozelli, 2007. Floods increase similarity among aquatic habitats in river-floodplain systems. Hydrobiologia 579: 1–13.
Tockner, K., N. P. D. Reiner, F. Schiemer & J. Vard, 1999. Hydrological connectivity and the exchange of organic matter and nutrients in a dynamic river floodplain system (Danube, Austria). Freshwater Biology 41: 521–535.
Tockner, K. & J. A. Standford, 2002. Riverine flood plains: present state and future trends. Environmental Conservation 29: 308–330.
Török, L., 2006. Contribution to the knowledge on quantitative and qualitative status of diatoms population from plankton of some lakes of the Danube Delta at the end of second millenium (1995–2000). Doctoral thesis. Danube Delta National Institute, Tulcea, Romania, p. 87. (http://www.indd.tim.ro).
Townsend, S. A., 2006. Hydraulic phases, persistant stratification, and phytoplankton in a tropical floodplain lake (Mary River, northern Australia). Hydrobiologia 556: 163–179.
Utermöhl, H., 1958. Zur Vervollkommnung der quantitativen Phytoplankton-Methodik. Mitteilungen der internationale Vereinigung für theoretische und angewandte Limnologie 9: 1–38.
Weilhoefer, C. L. & Y. Pan, 2007. Relationships between diatoms and environmental variables in wetlands in the Willamette valley, Oregon, USA. Wetlands 27: 668–682.
Zalocar de Domitrovic, Y., 2003. Effect of fluctuations in water level on phytoplankton development in three lakes of the Parana river floodplain (Argentina). Hydrobiologia 510: 175–193.
Acknowledgments
This project was supported by the Croatian Ministry of Science, Education, and Sports. We thank the anonymous reviewers for their constructive comments and helpful suggestions which substantially improved this manuscript.
Author information
Authors and Affiliations
Corresponding author
Additional information
Handling editor: J. Padisak
Rights and permissions
About this article
Cite this article
Mihaljević, M., Stević, F., Horvatić, J. et al. Dual impact of the flood pulses on the phytoplankton assemblages in a Danubian floodplain lake (Kopački Rit Nature Park, Croatia). Hydrobiologia 618, 77–88 (2009). https://doi.org/10.1007/s10750-008-9550-6
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10750-008-9550-6