
Abstract
Seed production areas (SPAs) are critical infrastructure for ecological restoration, particularly in fragmented landscapes where wildland seed crops are unavailable or wildland harvest is unsustainable. SPAs are useful for a wide range of species that are amenable to cultivation. Despite increasing research on SPAs, their value for biodiversity conservation has yet to be comprehensively described. Here, we highlight the key benefits of SPAs to biodiversity conservation. First, SPAs allow restoration to be conducted on a much greater scale than could be accomplished with wildland-harvested seed, thus protecting key biodiversity assets from harvest pressures. Second, the native seed production industry adds to the base of stakeholders who are invested in restoring and enhancing biodiversity. Third, SPAs provide novel opportunities for research and public exposure to native biodiversity. We also describe how cultivation can alter plant fitness compared to wildland plants, which acts as a multiplier to their conservation implications. SPAs could lead to two interrelated negative consequences that generate risks for taxonomic and genetic diversity at multiple scales: (1) SPAs can cause and multiply negative genetic legacies as a result of cultivation practices, and (2) SPA progenies can numerically and genetically dominate wildland plant populations. Nevertheless, SPA cultivation offers an opportunity to genetically diversify SPA-derived populations for success in restoration and enlarge the pool of species available for restoration, thus mitigating or solving some of these risks. Targeted government policies toward SPAs, additional research, and sound SPA management are necessary to minimize genetic risks and taxonomic redundancy, and also to maximize the conservation benefits of SPAs.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Producing seed of appropriate quality and quantity is a major challenge for ecological restoration globally (Nevill et al. 2018). The need for seed to create de novo communities (Brudvig 2011), combined with larger restoration initiatives worldwide, requires massive, reliable, and cost-effective supplies of native seed (Broadhurst et al. 2008). In many contexts, using wildland-harvested seed is not a feasible strategy to meet these demands (Broadhurst et al. 2016; Cuneo et al. 2018), especially in degraded landscapes with substantial habitat destruction, where collection can compromise the long-term persistence of remnant populations (Nevill et al. 2016). Additionally, prospects for wildland seed harvests will likely worsen in many regions as climate change, habitat destruction, and degradation continue to erode or eliminate plant populations (Broadhurst et al. 2016; Nevill et al. 2018).
Seed production areas (SPAs) are important systems to overcome the limitations of wildland-harvested seed supply (Merritt and Dixon 2011; Tischew et al. 2011). Typical SPAs are intentional plantings in which managers use a combination of agricultural and horticultural practices to produce seed of native species for restoration (Delpratt and Gibson-Roy 2015). Other seed production systems are closely related to, and intergrade with, typical SPAs. For example, some wildland populations that are intensively managed for seed production might be considered SPAs (Pedrini et al. 2020), as might seed orchards (see Box 1 for definitions), which produce genetically improved tree seed (Hancock et al. 2020). All of these systems use management of wild plants for seed production for restoration, conservation or other purposes.
SPAs are found globally and are managed by a variety of stakeholders, including government programs, small local growers, and large commercial operations (Fig. 1). The need for seed production systems is expected to grow as natural environments increasingly require restoration due to human impacts (Ellis et al. 2010), including climate change (Harris et al. 2006) and vast deforestation (Bradshaw 2012).

SPAs are a global phenomenon and vary from small-scale demonstrative projects (such as that pictured in the Falkland Islands) to industrial commercial operations, such as Prairie Moon Nursery in Minnesota (USA). Despite the different regions and contexts, SPA practices frequently resemble traditional agricultural or horticultural plantings of high intraspecific density. Many SPAs are concentrated in regions where natural areas have been highly impacted by habitat destruction and degradation, where established restoration industries lead to substantial demands for native seed. Pictured from upper left, clockwise: Texas (USA), Minnesota (USA), Saxony (Germany), a seed orchard in Chiang Mai Province (Thailand), New South Wales (Australia), the Falkland Islands (UK), and Florida (USA)
Studies that investigate the ecological and genetic consequences of SPAs have become more frequent (Broadhurst et al. 2008; Pedrini et al. 2020). Many of these studies are framed in the context of ecological restoration genetics or ecotypes (see Kettle et al. 2008; Dolan et al. 2008; Broadhurst et al. 2017). Others are more practical in nature, such as focusing on SPA infrastructure (e.g., Tischew et al. 2011; Kirmer et al. 2015; Gibson-Roy 2018; Jones 2019) or describing the establishment and role of SPAs in particular restoration projects (Cuneo et al. 2018). Because of the increasing interest in using SPAs to help meet the global demand for seed for restoration, considerably more research is required about the management and ecological effects of SPAs (Merritt and Dixon 2011; Nevill et al. 2016).
In this review, we synthesize existing literature to explore the conservation relevance of SPAs. We first discuss the benefits of SPAs to conservation and describe how these are contingent upon and influenced by government policies, programs, and restoration markets. We then explain how the targeted manipulation of field-based SPA populations through agronomic-like cultivation can lead to negative consequences, as well as altered or enhanced fitness potential in SPA stocks and their progenies relative to wildland populations. We discuss how the enhanced fitness of SPA stock can amplify effects on biodiversity and address methods to mitigate or counter potential negative effects of SPAs through sound management actions. We conclude by identifying long-term strategies to maximize the value of SPAs for ecological restoration and biodiversity conservation.
Practical benefits to conservation outcomes
SPAs have several attributes that enhance their value for biodiversity conservation. First, SPAs can upscale seed production required to meet the global demand for native seed. Globally, the reliable supply and affordability of seed are significant constraints for restoration practitioners and conservationists (Peppin et al. 2010; Nevill et al. 2016; Jalonen et al. 2018). SPA-cultivated seed can relieve supply chain and financial pressures due to their scalability and production efficiency (Pedrini et al. 2020; León-Lobos et al. 2020), as well as provide a greater diversity of species for planting (Jones 2019). These factors would make ambitious, landscape-scale restoration and rehabilitation goals more logistically and financially feasible than sourcing unreliable, limited, or expensive wildland seed, despite their high upfront costs (Merritt and Dixon 2011; Broadhurst et al. 2016). Upscaled restoration could lead to improved conservation outcomes by connecting remnants, restoring habitat for pollinators, providing food for higher trophic levels, or generating valuable ecosystem services throughout a landscape (Perring et al. 2015).
SPAs have several advantages over wildland seed harvesting. Wildland seed harvesting will often be insufficient to provision native plant materials for restoration efforts, though it may be the best option when SPA production is not viable (Jones 2019; Pedrini et al. 2020) or when wildland sources are readily available (e.g., Jalonen et al. 2018). Wildland harvesting can be unreliable due to fluctuating weather, seed predation pressure, or other disturbances (Mortlock 2000; Gibson-Roy et al. 2010). Furthermore, wildland harvesting is potentially damaging to natural populations, especially for short-lived species that rely on seed recruitment for persistence (Meissen et al. 2015). Wildland seed collectors also face various logistical and bureaucratic challenges to meet the demand for native seed (Mortlock 2000). Remnants suitable for wildland harvests can be scattered, and target species may be sparsely distributed both within a population and across a landscape, which may encourage seed collectors to sample easily accessible seed sources that are few in number (Nyoka et al. 2015; Dedefo et al. 2017). Responsible wildland harvesters must receive proper government permits or property-holder permission (Pedrini et al. 2020), and afterward they must conservatively harvest seed to avoid overstressing the seed resource (Delpratt and Gibson-Roy 2015; Meissen et al. 2015). A recent native seed sourcing guideline recommended that only 20% of a particular individual’s seed crop (and only 10% for annual species) should be harvested per season in wildlands (Pedrini and Dixon 2020). Together, these factors can cause wildland-harvested seed to be low in volume, expensive, and only sporadically available to practitioners (Pedrini et al. 2020). Furthermore, in many instances wildland seed collectors can only speculate on the genetic quality of their target populations (Neff et al. 2019). For example, collectors could unknowingly harvest poor-quality wildland seed if remnant seed sources are genetically depauperate (Broadhurst 2013; Broadhurst et al. 2017).
In contrast, SPA harvesting is more efficient due to a clustered, convenient location for SPA plants, the harvest of all seed for the life of the stand, and a reduced potential for seed contamination by non-target species (Broadhurst et al. 2016; Pedrini et al. 2020). SPAs therefore protect remnant communities by relieving them from harvest pressure, as well as by bringing more of their taxonomic and genetic diversity into this type of ex situ conservation planting (Box 2). SPAs provide a unique opportunity to access seed materials for species which are currently absent or underutilized in restoration (Hancock et al. 2020), and they can even be integrated into conservation strategies to propagate threatened and endangered species (Gibson-Roy 2010; Han et al. 2020). Despite the genetic challenges with SPAs (discussed in the Risks to conservation outcomes section), SPA management confers a greater degree of control and knowledge of a stock’s population genetics than wildland harvesting (Neff et al. 2019). Such management could lead to improved genetic diversity or post-dispersal performance than could be expected from some wildland sources (e.g., Broadhurst et al. 2017).
SPAs lead to additional conservation benefits by increasing stakeholder engagement. The management of SPAs creates a skilled conservation workforce and involves communities in restoration and conservation. Specialized growers and managers, including many in small and entrepreneurial private enterprises (De Vitis et al. 2017; Jones 2019), have developed in response to increasing native seed demand. SPAs can support economic activity and employment, particularly for rural or indigenous communities (Nevill et al. 2016; Gibson-Roy 2018; León-Lobos et al. 2020). For example, cultivating native species can diversify household incomes (Schmidt et al. 2019) and provide farmers an alternative source of income if traditional crops go through unfavorable market changes (Gibson-Roy 2018). In turn, these stakeholders can generate nonfinancial rewards. Private SPA managers often have under-recognized, specialized expertise and can advance biological or propagation knowledge of their cultivated species (Haase and Davis 2017; Jones 2019). Community-based wildland seed collectors are well-described sources of traditional and local ecological knowledge (de Urzedo et al. 2020). SPA managers provide information and feedback to ecologists and practitioners, particularly regarding the state of the restoration industry (e.g., Dunne and Dunne 2003); they also commonly establish collaborations with researchers (De Vitis et al. 2017).
SPAs have also been excellent initial locations for ecological studies of plant species (Gibson-Roy et al. 2010). Examples include pollinator interactions, phenology, seed germination, growth and fitness responses to environmental variation, and the effects of particular interspecific interactions in polycultures. For instance, Jiménez-Alfaro et al. (2020) studied the phenology and functional traits of 35 Mediterranean herbaceous species in seed-production systems. Studies of pollinators in SPAs have also been conducted (Tangmitcharoen et al. 2006; White et al. 2017; Schmidt et al. 2020). SPAs may be particularly valuable as restoration research study sites because they provide information about how site conditions determine target species’ performance, which can therefore be integrated into restoration planning and management (e.g., Cuneo et al. 2018). Despite these examples, restoration research at SPAs is uncommon, and there might be great capacity to increase the use of SPAs as research hubs.
Furthermore, SPA managers can introduce biodiversity into nontraditional conservation settings, because they can provide seed for agroforestry systems (Jiménez-Alfaro et al. 2020) and public roadsides (Gibson-Roy 2018), as well as gardens and landscaped areas (Hancock et al. 2020). SPAs may also present educational opportunities for schools, which can be observed in well-developed nurseries (Haase and Davis 2017). Such efforts expose and normalize the broader public to native flora and restoration efforts (Maunder et al. 2004). SPAs therefore provide benefits for multiple stakeholders (e.g., practitioners, communities, and educators).
Linking policies, programs, and economic markets
SPAs are often the result of emerging or established native seed markets, many of which are privately run enterprises. Native seed markets, and the restoration and conservation benefits of SPAs, are contingent on direct (e.g., mandates) and indirect (e.g., programs) government support for restoration. Government policies and programs can either create native seed markets or stimulate existing ones. In some cases, government agencies are the main purchasers of native seed (Jones 2019; Hancock et al. 2020). For example, the native seed industry in the western United States developed as a response to restoring large tracts of land for post-fire rehabilitation on federal lands (Peppin et al. 2010; Camhi et al. 2019), while the midwestern United States traces much of its SPA infrastructure to conservation programs in rural agricultural areas (Gibson-Roy 2018). In the European Union, habitat directives have stimulated restoration markets and SPA demand (Tischew et al. 2011; De Vitis et al. 2017). Government-funded Land Care programs, along with mandated reforestation and mine rehabilitation projects, have stimulated a native seed industry in Australia (Hancock et al. 2020), though this seed sector is described as a smaller “cottage” industry compared to other regions (Gibson-Roy 2018). Demands in Asia, Latin America, and parts of Africa are driven by reforestation initiatives, as well as agroforestry (Nyoka et al. 2015). Informal native seed markets can be found in some of these regions where native seed markets are emerging (e.g., eastern Africa, Dedefo et al. 2017). Similarly, in Amazonia, native seed demand is driven by mandatory landscape-scale restoration initiatives, which has led to the formation of decentralized, community-based seed collection networks (Schmidt et al. 2019; de Urzedo et al. 2019, 2020). Wildland-harvested seed, rather than SPA-produced seed, likely comprises most or all of seed sales in some of these regional examples (Jalonen et al. 2018). Nonetheless, we suggest SPAs could become more valuable contributors to native seed supplies in such developing native seed markets and could supplement or replace wildland harvesting.
Given the importance of government support for conservation and restoration, SPA managers are sensitive to changes in policies and market demands. Industry development can be limited by inadequate incentives for vegetation conservation or restoration (Hancock et al. 2020). Fluctuating government funding of conservation or restoration projects can destabilize native seed markets, both in producer finances and their seed supply (Dunne and Dunne 2003). Native seed prices can be highly variable due to inconsistent demand (Peppin et al. 2010; Hancock et al. 2020), which can be exacerbated by unpredictable yields from native species (Dunne and Dunne 2003). SPAs can have high up-front costs (e.g., land, infrastructure, equipment, or crop installation), laborious management requirements, and steep learning curves for managers who often lack information for cultivating native species (Jalonen et al. 2018; Camhi et al. 2019; Jones 2019). Greater and more consistent investment in restoration and conservation programs may enhance or financially stabilize SPA operations.
In unfavorable cases, policies may even stifle private stakeholders in the seed supply chain. Small-scale, private seed stakeholders in some emerging native seed markets can be undercut if government- and NGO-run nurseries flood the seed market with low-cost or free seed (Nyoka et al. 2015; Hancock et al. 2020). Abbandonato et al. (2018) and de Urzedo et al. (2019) have described how mandates, initially intended to manage commercially important plant species, were incongruously applied to native seed markets, thus restricting and burdening producers. Government regulation of seed provenancing and certification is also risk-laden for growers. Strict policies to use local seed and source certification could limit growers to small sales areas and a select few financial opportunities (see Mainz and Wieden 2019). On the other hand, a lack of effective certification or sourcing legislation can allow distant nurseries selling undesirable seed material to dominate the market and smother local businesses (Mainz and Wieden 2019). These examples highlight the need to advance native seed market policies to balance the needs of seed producers and end users.

The case for enhanced fitness potential
We posit that plants in SPAs (i.e., stock) can have enhanced fitness potential relative to unmanaged wildland individuals. Here, we use enhanced fitness potential as a descriptor of SPA plants to characterize significant advantages in propagule production, seed dispersal, and establishment relative to unmanaged wildland individuals and populations. Taken together, we speculate that SPA plants can be some of the most reproductively successful individuals throughout a landscape. SPA enhanced fitness can be viewed as beneficial in some contexts; large-scale restorations can be achieved that use large volumes of genetically diverse and appropriate seeds. However, enhanced fitness might also amplify the negative effects of SPAs on multiple levels of biodiversity (see “Risks to conservation outcomes” section).
Under natural conditions, plants face many barriers to successful reproduction, such as pollen and pollinator limitations, mate availability, stress and/or competitive interactions that reduce fitness, and seed predation (Fenner 2000; Wilcock and Neiland 2002). SPA management seeks to minimize these multiple constraints to boost yields. Mass plantings in SPAs may make them more attractive to pollinators. For example, Tangmitcharoen et al. (2006) found a greater number of potential insect pollinators in teak seed orchards compared to nearby wildland trees. High plant densities of target species can also promote gene flow by facilitating cross-pollination (McCallum et al. 2018, 2019). Managers intentionally boost growth through irrigation, fertilization, density manipulation, and pesticide application (Dunne and Dunne 2003; Gibson-Roy 2018; Nagel et al. 2019). SPAs are also potential safe sites from interspecific competition (Delpratt and Gibson-Roy 2015) and stresses due to extreme weather or other disturbance events (Gibson-Roy et al. 2010). The cultivation and harvesting practices of SPAs can reduce seed predation, which can be intense in the wild. Henderson (2017) described an anecdotal case where SPA cultivation of Baptisia bracteata precluded seed loss from the Baptisia seed specialist weevil Trichapion rostrum, which was comparatively intense in remnant natural areas. On the other hand, there is some evidence that agricultural-like production techniques might stimulate disease (Dyer et al. 2016) and pest infestation, reduce interspecific complementarity (Saxhaug et al. 2020), and/or reduce pollinator mutualisms (Tangmitcharoen et al. 2006), thus decreasing the fitness of SPA stocks. Taken together, however, SPA stocks generally should have greater survival and growth compared to unmanaged wildland plants, leading to greater and more consistent seed production (Fig. 3a; see Gibson-Roy et al. 2010).

A hypothetical comparison of production and dispersal distance of wildland and SPA progenies. The cultivation practices of SPAs generally create more volume and consistency of seed output (a). Wildland plant seed dispersal is leptokurtic, with most offspring dispersing close to the parent (b). Given constraints of wildland seed production and collection, relative to unmanaged wildland plant progenies, SPA plants have a permanent and consistent ability to disperse their progenies through human-vectored transport
Dispersal and establishment barriers can be significant population constraints for wildland plants. In contrast, SPA progenies are frequently and strategically moved throughout the landscape to establish persisting populations (Knapp and Dyer 1998). In other words, recruitment should be greater for seed destined for restorations versus seeds that naturally disperse in wildland populations (Broadhurst et al. 2008). SPA seed can also be moved substantially farther than the natural dispersal range of wildland plant seed through human-vectored dispersal (Fig. 3b; see Bullock et al. 2018), especially when limited markets for local seed create financial incentives for unrestricted sales (Jones 2019). In the Australian native seed sector, seed transports beyond local provenance ranges are common across restoration activities, most of which are components of biodiversity protection and Land Care projects (Hancock et al. 2020). Additional examples include the trans-ecoregional movement of C4 grasses in prairie restoration (Gustafson et al. 2001), post-fire rehabilitation seed in the western United States (Peppin et al. 2010), roadside wildflower beautification plantings in the southern United States (Gibson-Roy 2018; Turner et al. 2018), and the wildflower industry moving herbaceous species across Europe (Laikre et al. 2010; Mainz and Wieden 2019).
There are some exceptions to the dispersal advantages of SPA progenies. First, wildland seed can receive similar human-vectored dispersal advantages if it is collected for use in restoration. The average advantages conferred to harvested wildland plants, however, will likely be lower compared to SPA stocks (see challenges of wildland harvesting in “Practical benefits to conservation outcomes” section), especially when considered across those plants’ lifespans. Second, there are cases where SPAs have been specifically created to supply on-site restoration projects. For example, prairie SPAs were established on-site in the Kankakee Sands (Indiana, USA) to provide seed for unrestored acreage (Dolan et al. 2008). Cuneo et al. (2018) describe similar smaller-scale SPA operations to provide seed quantities and species diversity for a specific geographic region in Australian restoration projects. Despite this approach of establishing SPAs close to their primary utilization destination, there are nonetheless many more examples of SPA seed dispersing over long distances.
Another exception concerns the prohibition of nonlocal seed provenances, which has the potential to restrict long-distance movement. This especially applies to areas where seed transfer zones and seed certification schemes are developed and legally enforced (De Vitis et al. 2017; Durka et al. 2017; León-Lobos et al. 2020). Seed transfer zones are geographically defined areas within which materials are considered local to ensure local provenancing; plant adaptive traits and environmental factors (e.g., climate, soils) are considered to minimize maladaptation in restorations (Gibson et al. 2019; Cevallos et al. 2020). Seed certification encompasses a variety of quality assurance standards and documentation, e.g., seed identity and origin (Mainz and Wieden 2019). Frameworks to promote local provenancing may ineffectively regulate seed sales from distant or unspecified origins due to a lack of regulatory cohesion, as well as nontransparent market practices (Tischew et al. 2011; Mainz and Wieden 2019). Additionally, evidence is equivocal for the “local is best” strategy that drives these restrictions, and the approach is not universally accepted by practitioners or scientists (Broadhurst et al. 2008; Jones 2013; Jones et al. 2015). Using nonlocal seed may become a respected and more common practice in future restorations for multiple reasons (e.g., Hancock et al. 2020). First, practitioners may buy seed from broader geographic ranges for pragmatic reasons. For example, distant sources may have favorable cost-to-benefit ratios or more reliable availability than local sources (Broadhurst et al. 2016, 2018). Second, there is growing scientific interest in the strategic use of nonlocal sources. Some seeding strategies now call for using genotypes more suited for future climates (so-called “predictive provenancing;” Prober et al. 2015), mixing regional genotypes to enhance genetic diversity (“admixture provenancing;” Breed et al. 2013), or forming novel ecosystems when re-creations of historic ecosystems are impractical (Jones 2013). For these reasons, we suspect that there are, and will continue to be, frequent human-vectored long-distance dispersal of SPA progenies.
Seed destined for restoration projects, especially that from SPAs, may have unique properties beyond just considerations of distance. In well-developed native seed markets, SPA seed is often mixed and aggregated in storage facilities until sold (Haase and Davis 2017; Gibson-Roy 2018; Box 2). The storage and subsequent movement of this seed creates a spatial and temporal “bet hedging” of destination sites for a single seed set, similar to the temporal bet hedging of germination in seed banks known in wild annual species (see Evans and Dennehy 2005). For example, a hypothetical SPA plant’s seed set could be primarily dispersed westward to an adjacent ecoregion one year, while the next year, most sales could occur inside of the ecoregion. This can confer a highly stochastic, varied pool of vectored dispersal sites throughout the lifespan of SPA stocks.
Additionally, favorable maternal effects may occur for SPA seed. Maternal effects characterize how the maternal environment can affect the phenotype of offspring via environmental, genetic, and epigenetic factors (Roach and Wulff 1987; Bossdorf et al. 2008). SPA cultivation practices, such as irrigation and fertilization, are known to promote maternal effects in offspring that differ from those in wildland seed grown under more natural, and often more stressful, conditions. Examples of desirable maternal effects in SPAs include increased seed size, germination capability, or offspring growth rate (see Espeland and Hammond 2013). Although some have argued that SPAs should mimic the environmental conditions of wildland areas to promote adaptive maternal effects (Espeland et al. 2017), agronomic cultivation could also enhance germination or post-germination survival of SPA seed compared to wildland seed (see Drenovsky et al. 2016). However, the trade-offs associated with different maternal environments in SPAs are not well understood.
Risks to conservation outcomes
Detrimental genetic legacies
There is a heightened potential of detrimental genetic impacts resulting from the purposeful manipulation of SPA stock. Practically every decision made by SPA managers can affect the genetic composition of stock and their progenies, including the source from which the establishing seed materials are acquired and the specific management techniques used to produce and harvest seed (Vander Mijnsbrugge et al. 2010; Basey et al. 2015; Nevill et al. 2016; Espeland et al. 2017). Such actions can lead to a variety of genetic outcomes (e.g., erosion, drift, or selection) that compromise adequate or desirable genetic diversity and evolutionary potential (Broadhurst et al. 2008), conferring detrimental “genetic legacies” on SPA progenies.
Several genetic challenges exist for maintaining genetic integrity and preventing genetic erosion in SPA systems. Some commonly discussed issues include genetic erosion and inbreeding depression, inadvertent selection, and outbreeding depression (Vander Mijnsbrugge et al. 2010). If not addressed through best SPA practices, these challenges could result in lower progeny fitness and long-term persistence potential, as well as reduce the desirability of the seed to practitioners (Broadhurst et al. 2008; Abbandonato et al. 2018).
The risk of genetic erosion and inbreeding is a relatively well-characterized genetic legacy in SPA case studies. Using molecular markers, multiple studies have found evidence of genetic bottlenecks in cultivated plant populations relative to populations in remnants (Dolan et al. 2008; Kettle et al. 2008). For example, commercial sources of Lychnis flos-cuculi were less heterozygous and had higher inbreeding coefficients compared to natural populations (Aavik et al. 2012), though these results did not lead to detectable fitness declines (Aavik et al. 2014). The creation of SPA stocks using inbred wildland populations can compromise their genetic viability. Broadhurst et al. (2017) found that inbreeding depression was prevalent in SPA seed stocks due to the low genetic diversity of the remnant vegetation used to establish them. Similarly, botanic garden studies document how repeated generations of ex situ cultivation can lead to genetic drift and erosion (Ensslin et al. 2011).
Inadvertent or intentional selection is another genetic legacy that can influence the eventual fitness and biotic integrity of SPA seed. Examples of selection events potentially relevant to SPAs include inadvertent selection (1) against seed-shattering genotypes when harvesting seed en masse (Espeland et al. 2017) and (2) under cultivation, promoting faster and more consistent germination, which has been found in botanical gardens (Ensslin et al. 2018). Dyer et al. (2016) demonstrated unintended selection in two perennial grasses caused by SPA cultivation, including a phenological shift, as well as greater reproductive success from some source populations due to differential disease tolerance. Similarly, Nagel et al. (2019) describe changes in flowering and biomass phenotypes of Medicago lupulina, possibly due to the harvest time at a production farm. However, Nagel et al. (2019) did not find consistent phenotypic changes among some other study species. Similarly, maternal effects due to SPA cultivation practices could also be considered a form of accidental selection, and these are of special concern if they are maladaptive at the seeds’ destinations (Espeland et al. 2017). Advertent selection in SPAs has also been used to develop plant materials with enhanced progeny performance (Jones and Robins 2011; Chivers et al. 2016).
In some situations, using SPA seed could undermine local adaptation and seed sourcing guidelines, which may harm the eventual performance and biological integrity of the native populations. As suggested by many long-distance movements, genotypes being propagated in SPAs may be those that are currently available rather than those genetically suited to a target area, resulting in environmental and genetic mismatch. Reproduction between divergent populations could promote outbreeding depression and reduce favorable local adaptation (Hufford and Mazer 2003). The assumption of local superiority in plants (Broadhurst et al. 2008; Weeks et al. 2011; Jones 2013) is buttressed by studies documenting evidence for local adaptation (Vander Mijnsbrugge et al. 2010; Baughman et al. 2019). For example, studies of seven German grassland species in common gardens have found genetic, phenological, and fitness proxy differences among some of the taxa (Bucharova et al. 2017; Durka et al. 2017). These researchers interpreted their findings as support for the 22 seed transfer zones in Germany. However, many translocation and common garden experiments suggest local adaptation is more context-dependent and less common than often assumed (Leimu and Fischer 2008). Local adaptation can also be rapidly developed by natural selection, especially when there is high genetic variance (see Oduor et al. 2016). Consequently, while negative adaptive genetic legacies might result from SPAs at the population level, they could be smaller than otherwise believed, and perhaps less problematic than other challenges (e.g., loss of populations due to large-scale habitat destruction).
Additionally, SPAs could compromise genetic integrity at higher genetic levels through alternative types of outbreeding depression. There is a possibility of unfavorable hybridization events occurring in situ in SPAs among closely related species (Basey et al. 2015). The spread and establishment of SPA progenies ex situ could also result in interspecific hybrid zones (e.g., Winkler and Massatti 2020). There is heightened risk of outbreeding depression in SPAs if a stock has intraspecific chromosomal variations (namely, ploidy variation; Kramer et al. 2018). These examples highlight the potential genetic effects of SPAs across a hierarchy of genetic levels.
Distinct population and community composition
Due to the unequal production and commercial viability of the species cultivated in SPAs, as well as genetic legacies of SPA-produced seed, restoration that relies on SPAs can result in distinct populations and communities that differ compositionally and genetically from remnant populations and communities (Broadhurst et al. 2016). There can be an overemphasis on the use of workhorses, i.e., species that produce high volumes of seed due to their agronomic suitability (sensu Leger and Baughman 2014), that establish consistently in restoration projects, and are generally available at a low price in the marketplace (Dunne and Dunne 2003). Workhorse species are used in restoration projects throughout the world, including eucalypt and acacia overstory trees in Australian woodland restoration (Broadhurst et al. 2018), historical fodder crops in European grassland restorations (Ladouceur et al. 2018), and a minority of agronomically suitable legumes in the southern United States (Muir et al. 2018). In Veracruz, Mexico, Ramírez-Soto et al. (2018) documented an extreme case of workhorse overemphasis they term “pinarization.” Despite the native tree diversity in Veracruz (> 1000 species), five Pinus species comprised over two-thirds of reforestation trees grown in nurseries. The neglect of underutilized species, despite their presence in and ecological importance to original communities (Broadhurst et al. 2016), limits practitioners’ abilities to fully restore complex and functional plant communities. Underutilized species may be absent from SPAs and restoration projects due to poor agronomic suitability; a lack of interest or awareness from seed producers or consumers; permitting restrictions to obtain stock seed from the wild; or a combination of these factors (Broadhurst et al. 2016; Muir et al. 2018; Hancock et al. 2020). In many cases, entire functional groups are underutilized in restorations, such as herbaceous understory flora in Australian restorations (Gibson-Roy and McDonald 2014), lianas and epiphytes in Brazilian Atlantic forests (Vidal et al. 2020), or hemi-parasitic plants in North American prairies (Barak et al. 2017).
The prolific production of seed in SPAs could promote numeric dominance of many species. “Numeric dominance” may occur when SPA-derived seed constitutes a large proportion of the individuals and coverage of a species across a landscape due to restoration efforts. We hypothesize that numeric dominance is likely in the future for many species, particularly workhorses. This can be partially attributed to the enhanced fitness we described for SPAs and the need for reasonably priced seed in large-scale restoration markets (Jackson 1992). For example, large-scale conservation easement programs in the United States (such as the Conservation Reserve Program) might drive numeric dominance of SPA-derived and workhorse C4 grasses, especially when improved cultivars of these species are commonly used (see Chang et al. 2018). In the Intermountain West (USA), perennial C3 grass seed produced in SPAs is in high demand from government agencies for the restoration of weed-infested and wildfire-damaged public lands (Jones et al. 2015). Numeric dominance is likely in highly impacted areas with scarce remnant habitats, such as the prairies of midwestern North America or temperate grasslands in southeastern Australia. In Brazilian Atlantic forests, Moreira da Silva et al. (2017) described how large-scale production of a small subset of a hyper-diverse pool of forest species, produced in a few large nurseries, could encourage a compositional mismatch between restorations and remnants. The loss and degradation of remnant habitats will continue in many impacted landscapes (Millennium Ecosystem Assessment 2005), which could set a low threshold for SPA numeric dominance for some species.
Closely related to numeric dominance, “genetic dominance” of SPA offspring could also occur through numeric superiority and human-vectored dispersal. This idea is similar to genetic swamping, but here we use genetic dominance more broadly since genetic swamping implies an unfavorable loss of localized alleles driven by asymmetric gene flow (Laikre et al. 2010). It is also possible that continued habitat loss and fragmentation will limit gene flow between remnants and restored populations, leading to a mosaic of remnant and SPA-derived populations. This could promote genetic differentiation among these populations; in other cases, local populations could also become extirpated before gene flow with SPA progenies can occur. Multiple studies have found cultivars of widely used C4 grasses in prairie restorations can genetically differ from remnant sources (e.g., Mutegi et al. 2014; Chang et al. 2018), including having different ploidy levels. Similarly, in a study of three forbs in German grassland restorations, Kaulfuß and Reisch (2019) found that commercial sources of Knautia arvensis were genetically differentiated from nearby wild populations due to the commercial sources differing in ploidy. These results were similar to those of Aavik et al. (2012), who found that commercial sources of Lychnis flos-cuculi were genetically differentiated from wildland sources. Winkler and Massatti (2020) found evidence that a widely used cultivar in dryland plantings in the southwestern United States, Hilaria jamesii ‘Viva’, may have resulted from hybridization between H. jamesii and a closely related species, H. mutica. They also found that the cultivars may have established into the core range of H. jamesii, with unknown implications for that species’ genetics. In another potential case of genetic dominance, Turner et al. (2018) found a lack of population structure (i.e., panmixia) between wild and commercially seeded Texas populations of Lupinus texensis, a popular wildflower planted for aesthetics. The reason for this finding was unclear, but the data could be explained by homogenization of gene flow directed by human-vectored dispersal (Turner et al. 2018).
Large numbers of SPA-derived plants can influence reproduction dynamics across landscapes as plants colonize new habitats, integrate into the landscape, and exchange genes with wildland populations (Jackson 1992; Selbo and Snow 2005; Winkler and Massatti 2020). SPA progenies could therefore parallel the genetic effects of large-scale releases of economically valuable animal and tree species (Laikre et al. 2010). For example, Mejnartowicz (1996) found a large genetic distance between an isolated, declining Abies alba remnant population and nearby, more numerous anthropogenic populations in Polish forests, though these populations were not necessarily reproducing. In contrast, salmon hatchery progenies have been known to spread to wild habitats and genetically homogenize wild populations (e.g., Vasemägi et al. 2005). The dominance potentials of SPA stocks and their progenies, facilitated by their enhanced fitness advantages, could potentially homogenize populations genetically and communities taxonomically, with cascading impacts on other functional groups. For example, phenological differences among SPA plants could affect pollinators in restorations (Bucharova et al. 2017), and morphological differences of workhorse species can influence insect herbivores (e.g., Gustafson et al. 2001). Yet, for many SPA species, these possibilities are emerging concerns with as yet limited evidence or discussion.
Mitigating risks
Although SPAs may create or amplify negative biodiversity consequences, these challenges can be addressed. There is a tendency in the SPA literature to focus primarily on genetic risks, particularly at the population level, a justified concern because cultivation and large-scale restorations cause inevitable genetic changes (Nevill et al. 2016; Espeland et al. 2017). Nevertheless, there is no need for SPAs to exacerbate genetic risks; rather, SPAs are ideal places for managers to genetically manipulate SPA propagules to produce favorable genetic legacies. Favorable SPA management strategies can utilize their enhanced fitness potential, thereby leading to diverse, resilient, and functional populations and communities.
Genetic risks can be minimized or addressed through multiple approaches. Basey et al. (2015), Nevill et al. (2016), and Espeland et al. (2017) list general strategies of creating stocks for seed production that limit genetic erosion and accidental selection. Examples of these recommendations include collecting seed from multiple target sites with large population sizes, varying harvesting times and germination conditions of source seed, and limiting generation times for cultivated plants (Gibson-Roy et al. 2010). Genetic screening activities can assist the delineation of taxonomic or population genetic boundaries to prevent likely scenarios of outbreeding depression (e.g., ensuring same ploidy levels within SPAs, Nevill et al. 2016). Furthermore, continuous introduction of wildland-harvested plant materials can also contribute favorable spatial or temporal genetic variation to SPAs.
An additional approach that may be favorable to the population genetics of many SPAs is to “prime the pump” for natural selection by incorporating greater and/or more adaptive genetic diversity on SPA sites (Broadhurst et al. 2016; Chivers et al. 2016). Pump-priming enhances the evolutionary and persistence potential of SPA-produced seed when these are released throughout the landscape (Broadhurst et al. 2008), and it may also promote stock survival within SPAs (e.g., Suraj et al. 2019). However, there is no “one size fits all” policy for the genetic priming of SPA stocks because there can be varying goals of SPA seed destinations. For example, priming an orchard from which seed is desired for large-scale carbon sequestration or timber production in Southeast Asia will differ from an on-site SPA meant to buffer or connect prairie remnants. The control SPA managers have over stock creation and cultivation practices can accommodate these diverse goals. For example, SPA managers could focus on conserving local ecotypes at risk of extinction by genetically rescuing inbred seed sources (Gibson-Roy 2010; Broadhurst et al. 2017). Alternatively, admixture and predictive seed provenancing strategies (Prober et al. 2015; Bucharova et al. 2019) can be accomplished in SPAs by selectively mixing multiple distant source populations, creating productive and sustainable seed resources on a regional basis. A strategic genetic manipulation within SPAs should encourage capturing a large amount of targeted genetic variation (see Weeks et al. 2011) and consider both SPA manager and seed-user goals. Managers and end users may therefore benefit from standardized record-keeping regarding SPA stocks as quality assurance for genetic integrity (Pedrini and Dixon 2020). However, some of these suggestions could logistically complicate creation and management of SPAs for managers and may need to be better incentivized.
As a whole, SPAs must generate taxonomically and genetically diverse seed to promote biodiversity. With continuing habitat loss, a species’ representation in SPAs will become important to its long-term conservation—a form of domesticated nature (Kareiva et al. 2007). Studying SPA management techniques to cultivate underutilized species and to move beyond, or diversify, workhorse species will thus be critical (Broadhurst et al. 2016; Hancock et al. 2020). SPA practices should not lead to workhorse overuse as long as there are incentives and efforts to truly restore biodiverse, functional plant communities (Cuneo et al. 2018). In some cases, market incentives for growers to diversify the restoration species pool (Ladouceur et al. 2018) exist, encouraging some growers to diversify their holdings and cultivate more agronomically difficult species to establish a market niche (Jones 2019). Conservation easement and biodiversity offsetting programs could also increase market demand and interest for diversified SPA holdings (Gibson-Roy 2018), especially since many metrics used to assess easements and offsets prioritize greater species and functional diversity. When there are government-sponsored restoration efforts, SPA managers may cultivate a significant portion of the total regional species pool (Ladouceur et al. 2018; White et al. 2018; Vidal et al. 2020).
Moving forward: how can we more strategically utilize SPAs?
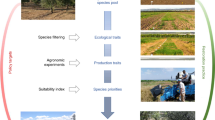
SPAs are not just a means to an end for ecological restoration. SPAs are strategic management tools with substantial value to multiple stakeholders, and have influence across multiple levels of biodiversity (Fig. 4). The strategic importance of SPAs to restoration and biodiversity conservation programs deserves greater recognition (Nevill et al. 2016). SPA practices should be viewed as opportunities for securing seed supplies and enhancing global conservation for practitioners and scientists. Although SPAs can generate challenges, they have the potential to augment biodiversity protection by delivering large volumes of high-quality and diverse plant materials (Jones 2019). Therefore, we believe that SPAs are the most comprehensive solution for responsibly and expeditiously satisfying the increasing demands for native seed (see also Merritt and Dixon 2011; Nevill et al. 2016, 2018). The importance and positive potential of SPAs drives the question: what should be done to maximize and enhance the use of SPAs? In response, we make three recommendations: (1) promote robust native seed industries and collaborations; (2) acquire knowledge pertaining to SPAs; and (3) enhance their genetic and taxonomic diversity.

A conceptual summary of the strategic importance of SPAs. SPAs are associated with interrelated benefits to biodiversity conservation (left). SPA management and use can also lead to certain risks relating to genetic legacies and distinct ecological communities (right). Enhanced fitness potential (center) characterizes the conceptual advantages of managed SPA plants compared to wildland settings in seed production, as well as their progenies’ dispersal and establishment abilities. Enhanced fitness could intensify opposing outcomes to biodiversity conservation, depending on SPA management practices
First, we recommend promoting the financial stability of SPA operations and stronger collaborations between multiple seed-industry stakeholders. Large SPA infrastructures and the skillsets of their managers did not develop spontaneously. SPA managers, especially those in the private sector, are often reliant on institutional or government policies promoting restoration or conservation initiatives. Additional and consistent governmental investment in vegetation-based restoration and conservation projects could nurture a robust network of stakeholders (Camhi et al. 2019; Hancock et al. 2020). However, flawed and poorly executed policies can work to the detriment of the native seed industry. A balance must be struck, therefore, between market stimulation, quality assurance of seed products, and bureaucratization.
Globally, SPA managers are crucial stakeholders who are often under-recognized and susceptible to unfavorable policies and fluctuating markets. These managers arguably possess local ecological knowledge and may be excellent project or research collaborators. We suggest SPAs can become valuable and sustainable sources of income to people living in areas with developing native seed markets, including economically empowering indigenous peoples and involving them in biodiversity protection (Nevill et al. 2016; Gibson-Roy 2018; León-Lobos et al. 2020). Further collaborations between researchers and these stakeholders could advance knowledge about SPAs and improve the economic viability and management of SPAs. One mutually beneficial policy we recommend is the development of organized seed-exchange networks that include private SPA managers (Schmidt et al. 2019). Seed-exchange networks can provide new species for restorations and valuable genetic materials to SPA managers, in turn improving the biodiversity benefits of produced seed (Jalonen et al. 2018). Seed-exchange networks could be formed among key stakeholders (Nevill et al. 2016); key members could include government or nongovernmental organizations producing seed or actively managing wild areas, botanic gardens, and seed banks; volunteer wildland seed collectors; and private SPA managers and nurseries.
Second, SPAs merit further scientific investigation. A larger knowledge base is critical to understand the conservation and restoration implications of SPAs. Research is lacking regarding SPA stakeholder perceptions, market forces, economics of seed production and seed procurement, and harvest and seed-conditioning methodologies. The yields, quality characteristics, ultimate destinations, and performance of SPA products remain largely uninvestigated, suggesting substantial knowledge gaps in restoration ecology. While we hypothesized that SPA practices will typically enhance stock fitness, research that explicitly compares wildland and SPA ecology is uncommon. Practical studies of SPAs are also imperative for long-term biodiversity protection (Delpratt and Gibson-Roy 2015). Examples include studying methods to boost yields of native seed crops and cultivate underutilized species, recording interannual variations in yields, or determining trade-offs associated with SPA monocultures versus polycultures (Saxhaug et al. 2020). Additional research could characterize which species form the restoration species pool (Ladouceur et al. 2018), as well as which factors lead to an underutilized versus workhorse status. Moreover, SPAs may be unique ecological or social study systems in their own right, but these topics are virtually unexplored.
Similarly, there are genetic questions about SPAs, as well as restorations broadly, that warrant further study. Studies could compare population genetic differences between remnants, SPA stocks, and then the eventual populations propagated for ecological restorations. Future research could investigate how the genetic composition in SPAs affects higher trophic levels and restoration success. In addition, the flaws and merits of improved plant materials in restoration and conservation projects should be further characterized (Chivers et al. 2016).
Lastly, SPA managers must emphasize diversifying SPAs both at the species and population genetic diversity level. In highly impacted landscapes, SPAs could become uniquely valuable repositories, sources, and propagators of both taxonomic and genetic diversity. To avoid overemphasis on workhorses, we suggest governments could require or financially reward taxonomically and functionally diverse restorations, develop regulatory or permitting systems to enable seed growers to source underutilized species, and spearhead research efforts for cultivating underutilized species and functional groups. Managers should capture large amounts of genetic diversity on SPAs to facilitate establishment success and evolutionary potential. If SPA genetic risks are addressed, detrimental “genetic legacies” and numeric and genetic dominance by SPA progenies could become less problematic. To address the genetic concerns, there are already several published guidelines about appropriate stock creation and seed collection practices that should be followed. Managers should emphasize maintaining large effective population sizes, minimizing time and generations in cultivation, and capturing a large amount of target genetic diversity and variation from remnants or improved seed sources (Nevill et al. 2016; Espeland et al. 2017). For example, managers can capture greater genetic diversity by sampling large populations over multiple sampling periods, cultivating stocks from multiple and distant populations in SPAs, and avoiding exclusive collections from populations with a high risk of being genetically depauperate. Genetic and taxonomic diversity of SPAs might be improved by continuous addition of wildland-harvested materials. This practice could also be logistically facilitated by the seed-exchange networks we recommended above.
The global necessity to maintain and restore native plant diversity in human-impacted landscapes is both a substantial challenge and opportunity. Native seed supplies must be upscaled and in high quality to achieve ambitious restoration and conservation goals. SPAs have a substantial influence on biodiversity and are an emerging pathway for the success of these endeavors.
References
Aavik T, Edwards PJ, Holderegger R, Graf R, Billeter R (2012) Genetic consequences of using seed mixtures in restoration: a case study of a wetland plant Lychnis flos-cuculi. Biol Conserv 145:195–204. https://doi.org/10.1016/j.biocon.2011.11.004
Aavik T, Bosshard D, Edwards PJ, Holderegger R, Billeter R (2014) Fitness in naturally occurring and restored populations of a grassland plant Lychnis flos-cuculi in a Swiss agricultural landscape: fitness in natural and sown plant populations. Restor Ecol 22:98–106. https://doi.org/10.1111/rec.12020
Abbandonato H, Pedrini S, Pritchard HW, De Vitis M, Bonomi C (2018) Native seed trade of herbaceous species for restoration: a European policy perspective with global implications. Restor Ecol 26:820–826. https://doi.org/10.1111/rec.12641
Barak RS, Williams EW, Hipp AL, Bowles ML, Carr GM, Sherman R, Larkin DL (2017) Restored tallgrass prairies have reduced phylogenetic diversity compared with remnants. J Appl Ecol 54:1080–1090. https://doi.org/10.1111/1365-2664.12881
Basey AC, Fant JB, Kramer AT (2015) Producing native plant materials for restoration: 10 rules to collect and maintain genetic diversity. Native Plants J 16:37–53. https://doi.org/10.3368/npj.16.1.37
Baughman OW, Agneray AC, Forister ML, Kilkenny FF, Espeland EK, Fiegener R, Horning ME, Johnson RC, Kaye TN, Ott J, St Clair JB, Leger EA (2019) Strong patterns of intraspecific variation and local adaptation in Great Basin plants revealed through a review of 75 years of experiments. Ecol Evol 9:6259–6275. https://doi.org/10.1002/ece3.5200
Bossdorf O, Richards CL, Pigliucci M (2008) Epigenetics for ecologists. Ecol Lett 11:106–115. https://doi.org/10.1111/j.1461-0248.2007.01130.x
Bradshaw CJA (2012) Little left to lose: deforestation and forest degradation in Australia since European colonization. J Plant Ecol 5:109–120. https://doi.org/10.1093/jpe/rtr038
Breed MF, Stead MG, Ottewell KM, Gardner MG, Lowe AJ (2013) Which provenance and where? Seed sourcing strategies for revegetation in a changing environment. Conserv Genet 14:1–10. https://doi.org/10.1007/s10592-012-0425-z
Broadhurst LM (2013) A genetic analysis of scattered yellow box trees (Eucalyptus melliodora A. Cunn. ex Schauer, Myrtaceae) and their restored cohorts. Biol Conserv 161:48–57. https://doi.org/10.1016/j.biocon.2013.02.016
Broadhurst LM, Lowe A, Coates DJ, Cunningham SA, McDonald M, Vesk PA, Yates C (2008) Seed supply for broadscale restoration: maximizing evolutionary potential. Evol Appl 1:587–597. https://doi.org/10.1111/j.1752-4571.2008.00045.x
Broadhurst LM, Jones TA, Smith FS, North T, Guja L (2016) Maximizing seed resources for restoration in an uncertain future. Bioscience 66:73–79. https://doi.org/10.1093/biosci/biv155
Broadhurst LM, Hopley T, Li L, Begley J (2017) A genetic assessment for seed production areas (SPAs) for restoration. Conserv Genet 18:1257–1266. https://doi.org/10.1007/s10592-017-0977-z
Broadhurst LM, Waters C, Coates DJ (2018) Native seed for restoration: a discussion of key issues using examples from the flora of southern Australia. Rangel J 39:487–498. https://doi.org/10.1071/RJ17055
Brudvig LA (2011) The restoration of biodiversity: where has research been and where does it need to go? Am J Bot 98:549–558. https://doi.org/10.3732/ajb.1000285
Bucharova A, Michalski S, Hermann JM, Heveling K, Durka W, Hölzel N, Kollmann J, Bossdorf O (2017) Genetic differentiation and regional adaptation among seed origins used for grassland restoration: lessons from a multispecies transplant experiment. J Appl Ecol 54:127–136. https://doi.org/10.1111/1365-2664.12645
Bucharova A, Bossdorf O, Hölzel N, Kollmann J, Prasse R, Durka W (2019) Mix and match: regional admixture provenancing strikes a balance among different seed-sourcing strategies for ecological restoration. Conserv Genet 20:7–17. https://doi.org/10.1007/s10592-018-1067-6
Bullock JM, Bonte D, Pufal G, da Silva CC, Chapman DS, García C, García D, Matthysen E, Delgado MM (2018) Human-mediated dispersal and the rewiring of spatial networks. Trends Ecol Evol 33:958–970. https://doi.org/10.1016/j.tree.2018.09.008
Camhi AL, Perrings C, Butterfield B, Wood T (2019) Market-based opportunities for expanding native seed resources for restoration: a case study on the Colorado Plateau. J Environ Manag 252:109644. https://doi.org/10.1016/j.jenvman.2019.109644
Cevallos D, Bede-Fazekas A, Tanács E, Szitár K, Halassy M, Kövendi-Jakó A, Török K (2020) Seed transfer zones based on environmental variables better reflect variability in vegetation than administrative units: evidence from Hungary. Restor Ecol 28:911–918. https://doi.org/10.1111/rec.13150
Chang H, Alexander HM, Mutegi E, Snow AA (2018) Habitat restoration and native grass conservation: a case study of switchgrass (Panicum virgatum). Restor Ecol 26:506–515. https://doi.org/10.1111/rec.12599
Chivers IH, Jones TA, Broadhurst LM, Mott IW, Larson SR (2016) The merits of artificial selection for the development of restoration-ready plant materials of native perennial grasses. Restor Ecol 24:174–183. https://doi.org/10.1111/rec.12323
Cuneo P, Gibson-Roy P, Fifield G, Broadhurst LM, Berryman T, Crawford A, Freudenberger D (2018) Restoring grassy woodland diversity through direct seeding: insights from six ‘best-practice’ case studies in southern Australia. Ecol Manag Restor 19:124–135. https://doi.org/10.1111/emr.12315
Dedefo K, Derero A, Tesfaye Y, Muriuki J (2017) Tree nursery and seed procurement characteristics influence on seedling quality in Oromia, Ethiopia. For Trees Livelihoods 26:96–110. https://doi.org/10.1080/14728028.2016.1221365
Delpratt J, Gibson-Roy P (2015) Sourcing seed for grassland restoration. In: Williams N, Marshall A, Morgan J (eds) Land of sweeping plains: managing and restoring the grasslands of south-eastern Australia. CSIRO Publishing, Clayton, pp 285–330
De Vitis M, Abbandonato H, Dixon KW, Laverack G, Bonomi C, Pedrini S (2017) The European native seed industry: characterization and perspectives in grassland restoration. Sustainability 9:1682. https://doi.org/10.3390/su9101682
Dolan RW, Marr DL, Schnabel A (2008) Capturing genetic variation during ecological restorations: an example from Kankakee Sands in Indiana. Restor Ecol 16:386–396. https://doi.org/10.1111/j.1526-100X.2007.00318.x
Drenovsky RE, Thornhill ML, Knestrick MA, Dlugos DM, Svejcar TJ, James JJ (2016) Seed production and seedling fitness are uncoupled from maternal plant productivity in three aridland bunchgrasses. Rangel Ecol Manag 69:161–168. https://doi.org/10.1016/j.rama.2016.01.003
Dunne RA, Dunne CG (2003) Trends in the western native plant seed industry since 1990. Native Plants J 4:88–94. https://doi.org/10.3368/npj.4.2.88
Durka W, Michalski SG, Berendzen KW, Bossdorf O, Bucharova A, Hermann JM, Hölzel N, Kollmann J (2017) Genetic differentiation within multiple common grassland plants supports seed transfer zones for ecological restoration. J Appl Ecol 54:116–126. https://doi.org/10.1111/1365-2664.12636
Dyer AR, Knapp EE, Rice KJ (2016) Unintentional selection and genetic changes in native perennial grass populations during commercial seed production. Ecol Restor 34:39–48. https://doi.org/10.3368/er.34.1.39
Ellis EC, Goldewijk KK, Siebert S, Lightman D, Ramankutty N (2010) Anthropogenic transformation of the biomes, 1700 to 2000. Glob Ecol Biogeogr 19:589–606. https://doi.org/10.1111/j.1466-8238.2010.00540.x
Ensslin A, Sandner TM, Matthies D (2011) Consequences of ex situ cultivation of plants: genetic diversity, fitness and adaptation of the monocarpic Cynoglossum officinale L. in botanic gardens. Biol Conserv 144:272–278. https://doi.org/10.1016/j.biocon.2010.09.001
Ensslin A, van de Vyver A, Vanderborght T, Godefroid S (2018) Ex situ cultivation entails high risk of seed dormancy loss on short-lived wild plant species. J Appl Ecol 55:1145–1154. https://doi.org/10.1111/1365-2664.13057
Espeland EK, Hammond D (2013) Maternal effects on growth and competitive ability in a commonly used restoration species. Native Plants J 14:231–242. https://doi.org/10.3368/npj.14.3.231
Espeland EK, Emery NC, Mercer KL, Woolbright SA, Kettenring KM, Gepts P, Etterson JR (2017) Evolution of plant materials for ecological restoration: insights from the applied and basic literature. J Appl Ecol 54:102–115. https://doi.org/10.1111/1365-2664.12739
Evans MEK, Dennehy JJ (2005) Germ banking: bet-hedging and variable release from egg and seed dormancy. Q Rev Biol 80:431–451. https://doi.org/10.1086/498282
Fenner M (2000) Seeds: the ecology of regeneration in plant communities, 2nd edn. CABI Publishing, Wallingford
Gibson A, Nelson CR, Rinehart S, Archer V, Eramian A (2019) Importance of considering soils in seed transfer zone development: evidence from a study of the native Bromus marginatus. Ecol Appl 29:e01835. https://doi.org/10.1002/eap.1835
Gibson-Roy P (2010) Reconstructing new populations of the endangered Rutidosis leptorrhynchoides (button wrinklewort) at two sites in south-western Victoria. Australas Plant Conserv 19:13–14
Gibson-Roy P (2018) Restoring grassy ecosystems: feasible or fiction? An inquisitive Australian’s experience in the USA. Ecol Manag Restor 19:11–25. https://doi.org/10.1111/emr.12327
Gibson-Roy P, McDonald T (2014) Reconstructing grassy understories in south-eastern Australia: interview with Paul Gibson-Roy. Ecol Manag Restor 15:111–122. https://doi.org/10.1111/emr.12116
Gibson-Roy P, Moore G, Delpratt J, Gardner J (2010) Expanding horizons for herbaceous ecosystem restoration: the Grassy Groundcover Restoration Project. Ecol Manag Restor 11:176–186. https://doi.org/10.1111/j.1442-8903.2010.00547.x
Gustafson DJ, Gibson DJ, Nickrent DL (2001) Characterizing three restored Andropogon gerardii Vitman (big bluestem) populations established with Illinois and non-Illinois seed: established plants and their offspring. In: Bernstein NP, Ostrander LJ (eds) Proceedings of the seventeenth North American Prairie Conference. Northern Iowa Area Community College, Mason City, IA, pp 118–124
Haase DL, Davis AS (2017) Developing and supporting quality nursery facilities and staff are necessary to meet global forest and landscape restoration needs. Reforesta 4:69–93. https://doi.org/10.21750/REFOR.4.06.45
Han L, Love K, Peace B, Broadhurst LM, England N, Li L, Bush D (2020) Origin of planted Eucalyptus benthamii trees in Camden NSW: checking the effectiveness of circa situm conservation measures using molecular markers. Biodivers Conserv 29:1301–1322. https://doi.org/10.1007/s10531-020-01936-4
Hancock N, Gibson-Roy P, Driver M, Broadhurst LM (2020) The Australian native seed survey report. Australian Network for Plant Conservation, Canberra
Harris JA, Hobbs RJ, Higgs E, Aronson J (2006) Ecological restoration and global climate change. Restor Ecol 14:170–176. https://doi.org/10.1111/j.1526-100X.2006.00136.x
Henderson R (2017) Of checks, balances, and seed production. Prairie Promoter 30:6–8
Hufford KM, Mazer SJ (2003) Plant ecotypes: genetic differentiation in the age of ecological restoration. Trends Ecol Evol 18:147–155. https://doi.org/10.1016/S0169-5347(03)00002-8
Jackson LL (1992) The role of ecological restoration in conservation biology. In: Hall CA (ed) Conservation biology: the theory and practice of nature conservation, preservation, and management. Springer, New York, pp 433–451
Jalonen R, Valette M, Boshier D, Duminil J, Thomas E (2018) Forest and landscape restoration severely constrained by a lack of attention to the quantity and quality of tree seed: insights from a global survey. Conserv Lett 11:e12424. https://doi.org/10.1111/conl.12424
Jiménez-Alfaro B, Frischie S, Stolz J, Gálvez-Ramírez C (2020) Native plants for greening Mediterranean agroecosystems. Nat Plants 6:209–214. https://doi.org/10.1038/s41477-020-0617-3
Jones TA (2013) When local isn’t best. Evol Appl 6:1109–1118. https://doi.org/10.1111/eva.12090
Jones TA (2019) Native seeds in the marketplace: meeting restoration needs in the Intermountain West, United States. Rangel Ecol Manag 72:1017–1029. https://doi.org/10.1016/j.rama.2019.07.009
Jones TA, Robins JG (2011) Appropriate use of genetic manipulation for the development of restoration plant materials. In: Lüttge UE, Beyschlag W, Büdel B, Francis D (eds) Progress in botany, vol 72. Springer, Berlin, pp 249–264
Jones TA, Monaco TA, Rigby CW (2015) The potential of novel native plants materials for the restoration of novel ecosystems. Elementa 3:e000047. https://doi.org/10.12952/journal.elementa.000047
Kaulfuß F, Reisch C (2019) Restoration of grasslands using commercially produced seed mixtures: genetic variation within and among natural and restored populations of three common grassland species. Conserv Genet 20:373–384. https://doi.org/10.1007/s10592-018-01138-0
Kareiva P, Watts S, McDonald R, Boucher T (2007) Domesticated nature: shaping landscapes and ecosystems for human welfare. Science 316:1866–1869. https://doi.org/10.1126/science.1140170
Kettle CJ, Ennos RA, Jaffré T, Gardner M, Hollingsworth PM (2008) Cryptic genetic bottlenecks during restoration of an endangered tropical conifer. Biol Conserv 141:1953–1961. https://doi.org/10.1016/j.biocon.2008.05.008
Kirmer A, Valkó O, Török P, Tóthmérész B, Tischew S, Kapocsi I, Radócz S, Miglécz T, Tóth K, Klemen A, Deák B (2015) Large-scale grassland restoration: high diversity seeding and knowledge transfer of regional seed propagation to Hungary. Debrecen University Press, Debrecen
Knapp EE, Dyer AR (1998) When do genetic considerations require special approaches to ecological restoration? In: Fieldler PL, Kareiva PM (eds) Conservation biology for the coming decade. Chapman and Hall, New York, pp 345–363
Kramer AT, Wood TE, Frischie S, Havens K (2018) Considering ploidy when producing and using mixed-source native plant materials for restoration. Restor Ecol 26:13–19. https://doi.org/10.1111/rec.12636
Ladouceur E, Jiménez-Alfaro B, Marin M, De Vitis M, Abbandonato H, Iannetta PPM, Bonomi C, Pritchard HW (2018) Native seed supply and the restoration species pool. Conserv Lett 11:e12381. https://doi.org/10.1111/conl.12381
Laikre L, Schwartz MK, Waples RS, Ryman N, GeM Working Group (2010) Compromising genetic diversity in the wild: unmonitored large-scale release of plants and animals. Trends Ecol Evol 25:520–529. https://doi.org/10.1016/j.tree.2010.06.013
Leger EA, Baughman OW (2014) What can natural selection tell us about restoration? Finding the best seed sources for use in disturbed systems. In: Kirmer A, Kiehl K (eds) Guidelines for native seed production and grassland restoration. Cambridge Scholars, Newcastle, pp 14–36
Leimu R, Fischer M (2008) A meta-analysis of local adaptation in plants. PLoS ONE 3:e4010. https://doi.org/10.1371/journal.pone.0004010
León-Lobos P, Bustamante-Sánchez MA, Nelson CR, Alarcón D, Hasbún R, Way M, Pritchard HW, Armesto JJ (2020) Lack of adequate seed supply is a major bottleneck for effective ecosystem restoration in Chile: friendly amendment to Bannister et al. (2018). Restor Ecol 28:277–281. https://doi.org/10.1111/rec.13113
Mainz AK, Wieden M (2019) Ten years of native seed certification in Germany: a summary. Plant Biol 21:383–388. https://doi.org/10.1111/plb.12866
Maunder M, Havens K, Guerrant EO Jr, Falk DA (2004) Ex situ methods: a vital but underused set of conservation resources. In: Guerrant EO, Havens K, Maunder M (eds) Ex situ plant conservation. Island Press, Washington, DC, pp 389–408
McCallum KP, Lowe AJ, Breed MF, Paton DC (2018) Spatially designed revegetation: why the spatial arrangement of plants should be as important to revegetation as they are to natural systems. Restor Ecol 26:446–455. https://doi.org/10.1111/rec.12690
McCallum KP, Breed MF, Paton DC, Lowe AJ (2019) Clumped planting arrangements improve seed production in a revegetated eucalypt woodland. Restor Ecol 27:638–646. https://doi.org/10.1111/rec.12905
Meissen JC, Galatowitsch SM, Cornett MW (2015) Risks of overharvesting seed from native tallgrass prairies. Restor Ecol 23:882–891. https://doi.org/10.1111/rec.12295
Mejnartowicz L (1996) Cisovka: the relic population of Abies alba and its relationship to man-made silver-fir stands in Białowieża primeval forest. Acta Soc Bot Pol 65:319–328. https://doi.org/10.5586/asbp.1996.049
Merritt DJ, Dixon KW (2011) Restoration seed banks: a matter of scale. Science 332:424–425. https://doi.org/10.1126/science.1203083
Millennium Ecosystem Assessment (2005) Ecosystems and human well-being: current state and trends. Findings of the condition and trends working group. In: Hassan R, Scholes R, Ash N (eds) Millennium ecosystem assessment series. Island Press, Washington, DC
Moreira da Silva AP, Schweizer D, Rodrigues-Marques H, Cordeiro-Teixeira AM, Nascente dos Santos TVM, Sambuichi RHR, Badari CG, Gaudare U, Brancalion PHS (2017) Can current native tree seedling production and infrastructure meet an increasing forest restoration demand in Brazil? Restor Ecol 25:509–515. https://doi.org/10.1111/rec.12470
Mortlock W (2000) Local seed for revegetation. Where will all that seed come from? Ecol Manag Restor 1:93–101. https://doi.org/10.1046/j.1442-8903.2000.00029.x
Muir JP, Pitman WD, Smith FS, Lloyd-Reilley J, Shadow RA (2018) Challenges to developing native legume seed supplies: the Texas experience as a case study. Native Plants J 19:224–238. https://doi.org/10.3368/npj.19.3.224
Mutegi E, Stottlemyer AL, Snow AA, Sweeney PM (2014) Genetic structure of remnant populations and cultivars of switchgrass (Panicum virgatum) in the context of prairie conservation and restoration. Restor Ecol 22:223–231. https://doi.org/10.1111/rec.12070
Nagel R, Durka W, Bossdorf O, Bucharova A (2019) Rapid evolution in native plants cultivated for ecological restoration: not a general pattern. Plant Biol 21:551–558. https://doi.org/10.1111/plb.12901
Neff D, Kettle C, Gotor E (2019) Costs of tree seed and seedling supply systems. The cost of integrating genetic diversity into forest landscape restoration. Biodiversity International, Rome, p 42
Nevill PG, Tomlinson S, Elliott CP, Espeland EK, Dixon KW, Merritt DJ (2016) Seed production areas for the global restoration challenge. Ecol Evol 6:7490–7497. https://doi.org/10.1002/ece3.2455
Nevill PG, Cross AT, Dixon KW (2018) Ethical seed sourcing is a key issue in meeting global restoration targets. Curr Biol 28:R1378–R1379. https://doi.org/10.1016/j.cub.2018.11.015
Nyoka BI, Roshetko J, Jamnadass R, Muriuki J, Kalinganire A, Lillesø JPB, Beedy T, Cornelius J (2015) Tree seed and seedling supply systems: a review of the Asia, Africa and Latin America models. Small-Scale For 14:171–191. https://doi.org/10.1007/s11842-014-9280-8
Oduor AMO, Leimu R, van Kleunen M (2016) Invasive plant species are locally adapted just as frequently and at least as strongly as native plant species. J Ecol 104:957–968. https://doi.org/10.1111/1365-2745.12578
Pedrini S, Dixon KW (2020) International principles and standards for native seeds in ecological restoration. Restor Ecol 28:S286–S303. https://doi.org/10.1111/rec.13155
Pedrini S, Gibson-Roy P, Trivedi C, Gálvez-Ramírez C, Hardwick K, Shaw N, Frischie S, Laverack G, Dixon KW (2020) Collection and production of native seeds for ecological restoration. Restor Ecol 28:S228-238. https://doi.org/10.1111/rec.13190
Peppin DL, Fulé PZ, Lynn JC, Mottek-Lucas AL, Hull Sieg C (2010) Market perceptions and opportunities for native plant production on the Southern Colorado Plateau. Restor Ecol 18:113–124. https://doi.org/10.1111/j.1526-100X.2010.00656.x
Perring MP, Standish RJ, Price JN, Craig MD, Erickson TE, Ruthrof KX, Whiteley AS, Valentine LE, Hobbs RJ (2015) Advances in restoration ecology: rising to the challenges of the coming decades. Ecosphere 6:1–25. https://doi.org/10.1890/ES15-00121.1
Prober SM, Byrne M, McLean EH, Steane DA, Potts BM, Vaillancourt RE, Stock WD (2015) Climate-adjusted provenancing: a strategy for climate-resilient ecological restoration. Front Ecol Evol 3:65. https://doi.org/10.3389/fevo.2015.00065
Ramírez-Soto A, Villa-Bonilla B, Lucio-Palacio CR, Libreros LL, Sánchez-Velásquez LR, Inzunza ER (2018) Mexico’s official reforestation programs are shrinking and narrowing its vision at a higher financial expense. For Policy Econ 94:32–34. https://doi.org/10.1016/j.forpol.2018.06.006
Roach DA, Wulff RD (1987) Maternal effects in plants. Ann Rev Ecol Syst 18:209–235. https://doi.org/10.1146/annurev.es.18.110187.001233
Saxhaug KF, Jungers JM, Hegeman AD, Wyse DL, Sheaffer CC (2020) Cultivation of native plants for seed and biomass yield. Agron J 112:1815–1827. https://doi.org/10.1002/agj2.20195
Schmidt IB, de Urzedo DI, Piña-Rodrigues FCM, Vieira DLM, de Rezende GM, Sampaio AB, Junqueira RGP (2019) Community-based native seed production for restoration in Brazil–the role of science and policy. Plant Biol 21:389–397. https://doi.org/10.1111/plb.12842
Schmidt LA, Gilpin AM, Cook JM, Rymer P, Gibson-Roy P, Craigie S, Power SA (2020) Restoration of native wildflower patches in agronomic settings for diverse and healthy pollinator populations. Australas Plant Conserv 28:17–19
Selbo SM, Snow AA (2005) Flowering phenology and genetic similarity among local and recently introduced populations of Andropogon gerardii in Ohio. Restor Ecol 13:441–447. https://doi.org/10.1111/j.1526-100X.2005.00055.x
Suraj PG, Nagabhushana K, Kamalakannan R, Varghese M (2019) Impact of fertility variation on genetic diversity and phenotypic traits in second generation seed production areas and clonal seed orchards of Eucalyptus camaldulensis. Silvae Genet 68:29–40. https://doi.org/10.2478/sg-2019-0006
Tangmitcharoen S, Takaso T, Siripatanadilox S, Tasen W, Owens JN (2006) Behavior of major insect pollinators of teak (Tectona grandis L. f.): a comparison of clonal seed orchard versus wild trees. For Ecol Manag 222:67–74. https://doi.org/10.1016/j.foreco.2005.10.041
Tischew S, Youtie B, Kirmer A, Shaw N (2011) Farming for restoration: building bridges for native seeds. Ecol Restor 29:219–222. https://doi.org/10.3368/er.29.3.219
Turner KG, Huang DI, Cronk QCB, Rieseberg LH (2018) Homogenization of populations in the wildflower, Texas bluebonnet (Lupinus texensis). J Hered 109:152–161. https://doi.org/10.1093/jhered/esx094
de Urzedo DI, Fisher R, Piña-Rodrigues FCM, Freire JM, Junqueira RGP (2019) How policies constrain native seed supply for restoration in Brazil. Restor Ecol 27:768–774. https://doi.org/10.1111/rec.12936
de Urzedo DI, Piña-Rodrigues FCM, Feltran-Barbieri R, Junqueira RGP, Fisher R (2020) Seed networks for upscaling forest landscape restoration: is it possible to expand native plant sources in Brazil? Forests 11:259. https://doi.org/10.3390/f11030259
Vander Mijnsbrugge K, Bischoff A, Smith B (2010) A question of origin: where and how to collect seed for ecological restoration. Basic Appl Ecol 11:300–311. https://doi.org/10.1016/j.baae.2009.09.002
Vasemägi A, Gross R, Paaver T, Koljonen ML, Nilsson J (2005) Extensive immigration from compensatory hatchery releases into wild Atlantic salmon population in the Baltic Sea: spatio-temporal analysis over 18 years. Heredity 95:76–83. https://doi.org/10.1038/sj.hdy.6800693
Vidal CY, Naves RP, Viani RAG, Rodrigues RR (2020) Assessment of the nursery species pool for restoring landscapes in southeastern Brazil. Restor Ecol 28:427–434. https://doi.org/10.1111/rec.13096
Weeks AR, Sgrò CM, Young AG, Frankham R, Mitchell NJ, Miller KA, Byrne M, Coates DJ, Eldridge MDB, Sunnucks P, Breed MF, James EA, Hoffmann AA (2011) Assessing the benefits and risks of translocations in changing environments: a genetic perspective. Evol Appl 4:709–725. https://doi.org/10.1111/j.1752-4571.2011.00192.x
White N, Gibson-Roy P, Cook JM (2017) Plant–pollinator interactions within a seed production area for native grassland restoration. In: Johnson SN (ed) Invertebrate ecology of Australasian grasslands. Proceedings of the ninth ACGIE. Western Sydney University, Hawkesbury, pp 22–25
White A, Fant JB, Havens K, Skinner M, Kramer AT (2018) Restoring species diversity: assessing capacity in the U.S. native plant industry. Restor Ecol 26:605–611. https://doi.org/10.1111/rec.12705
Wilcock C, Neiland R (2002) Pollination failure in plants: why it happens and when it matters. Trends Plant Sci 7:270–277. https://doi.org/10.1016/S1360-1385(02)02258-6
Winkler DE, Massatti R (2020) Unexpected hybridization reveals the utility of genetics in native plant restoration. Restor Ecol 28:1047–1052. https://doi.org/10.1111/rec.13189
Acknowledgements
Comments from two anonymous reviewers greatly improved the previous version of the manuscript. We thank the following individuals and organizations for providing images used in Fig. 1: F. Ross, S. and A. Felton (Falklands Conservation), Native American Seed (Junction, TX), Prairie Moon Nursery (Winona, MN), A. Kareen (Verband deutscher Wildsamen und Wildpflanzenproduzenten), T. Zinn, S. Tangmitcharoen, and Ernst Seed (Meadville, PA). Special thanks to C. Carpenter and S. Lindholm for assistance with Fig. 3, and S. Flamand for editing.
Funding
No funding was received to assist with the preparation of this manuscript.
Author information
Authors and Affiliations
Contributions
All authors contributed to the conception of the article contents. The drafts of the article were primarily written by JZ. LB, PG-R, and TJ contributed to the writing of the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest to disclose.
Additional information
Communicated by Eckehard G. Brockerhoff.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Zinnen, J., Broadhurst, L.M., Gibson-Roy, P. et al. Seed production areas are crucial to conservation outcomes: benefits and risks of an emerging restoration tool. Biodivers Conserv 30, 1233–1256 (2021). https://doi.org/10.1007/s10531-021-02149-z
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10531-021-02149-z