
Abstract
This article gives an overview of high-cell-density cultures for polyhydroxyalkanoate (PHA) production and their modes of operation for increasing productivity. High cell densities are very important in PHA production mainly because this polymer is an intracellular product accumulated in various microorganisms, so a high cellular content is needed for the polymer production. This review describes relevant results from fed-batch, repeated batch, and continuous modes of operation without and with cell recycle for the production of these polymers by microorganisms. Finally, recombinant microorganisms for PHA production, as well future directions for PHA production, are discussed.
Similar content being viewed by others

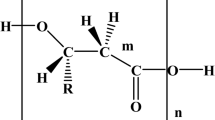
Avoid common mistakes on your manuscript.
Introduction
Polyhydroxyalkanoates (PHAs) are biodegradable biopolymers produced by microorganisms. Poly(3-hydroxybutyrate) (P(3HB)) and poly(3-hydroxybutyrate-co-3-hydroxyvalerate) (P(3HB-co-3HV)) are the best representatives of this polymer class that have properties similar to petrochemical polymers, and they can be produced from renewable sources [28]. These polymers have high production costs which limit their more widespread use [5]. Over the last decade, research has focused on the development of alternative cost-effective processes for PHA production, including the use of low-value substrates (agricultural or industrial waste or surplus feedstocks) [4, 12, 40, 61], mixed microbial cultures [13, 68], recombinant strains [1, 17, 36, 64], and the development of efficient culture strategies, among which are cultures for high-cell-density production in the shortest time [1, 31, 35].
The use of low-cost agroindustrial substrates can decrease the PHA production costs, because its represent 50 % of P(3HB) production costs [10]. These substrates have moderate carbon source concentrations, e.g., citrus molasses (80 g l−1 [4]), or low carbon source concentration, e.g., whey (40 g l−1 [1]). Therefore, strategies for feeding with these substrates are needed when high cell densities are desired, but few works about feedstock feed strategies for high-cell-density culture have been reported [1, 35, 74] owing to difficulties in operations for substrates containing moderate or low carbon concentrations.
One of the main goals for biotechnological process development is to maximize productivity, to obtain the highest density of products of interest in a reaction volume within the shortest time. High cell densities are a requirement for high productivities [65]. Historically, high-cell-density cultures (HCDC) were first established for yeast in the production of single-cell proteins, bioethanol, and biomass [65]. Later, dense cultures of other mesophiles (streptomyces, methylotrophs, etc.) producing other kinds of products (antibiotics, PHA, etc.) were developed [38, 72].
Continuous and fed-batch cultures are the main operation modes for HCDC of microbial cells. Usually, fed-batch cultures reach densities above 50 g l−1 of total dry cells [64, 77]. However, for the production of PHA, densities above 100 g l−1 of total dry cells and around 30–40 g l−1 of residual biomass (Xr) [active biomass without the polymer; Xr = total biomass − P(3HB)] are considered high cell densities [3, 67].
Once PHAs are accumulated inside the cell, and in most cases under limitation of an essential nutrient (nitrogen, phosphorus, oxygen, etc.), their productivity is directly proportional to the residual biomass production in the process. Hence, processes with high cell densities (higher Xr concentration) are desired because they favor PHA production, especially in terms of reduction of the culture volume, reduction of residual liquids, lower production costs, and lower investment in equipment compared to the low-cell-density processes [7, 27, 32, 67].
Currently, continuous, repeated batch, and fed-batch cultures are the main culture strategies used for high-cell-density PHA production. For P(3HB) production, fed-batch cultures have been widely used with the goal of reaching high cell density; such cultures increase the system productivity and require low investment and production costs [35, 67, 69, 75]. When agroindustrial wastes are used, a dilution of the cell occurs owing to the moderate or low carbon source concentration; therefore, in order to avoid this problem, cell recycle can be used to retain the cell inside the bioreactor and drain a partial volume of the fermentation broth [1].
Recombinant microorganisms have been investigated to produce PHA with some advantage in relation to wild-type strains to obtain HCDC for PHA bioprocess production. For example, the following recombinant strains have been used for PHA production: Escherichia coli [1, 11, 16, 73, 74], Cupriavidus necator [17, 22, 57, 63], Aeromonas hydrophila 4AK4 [42], Methylobacterium extorquens [23], Pseudomonas strains [25, 45], and yeasts strains [36, 55, 56, 78].
In this context, this review summarizes the trends in HCDC for PHA bioprocess production.
Main operation modes for production of polyhydroalkanoates in high-cell-density cultures
Fed-batch culture
The fed-batch technique was originally devised by yeast producers in the early 1900s to regulate the growth in batch culture of Saccharomyces cerevisiae [54]. Yeast producers observed that in the presence of high concentrations of malt, a by-product—ethanol—was produced, whereas in low concentrations of malt, the yeast growth was restricted. This problem was solved by a controlled feeding regime, so that yeast growth remained substrate limited [46]. The concept was then extended to the production of other products, such as some enzymes, antibiotics, growth hormones, microbial cells, vitamins [46], amino acids and other organic acids [48], and polymers such as PHA [29, 31, 65].
In fed-batch culture, cells are grown under a batch regime for some time, usually until close to the end of the exponential growth phase. At this point, the bioreactor is fed with a solution of substrates, without the removal of culture broth, until the desired volume in the bioreactor is achieved. This solution should have all necessary nutrients to keep a desired specific growth rate, and simultaneously reducing the production of by-products. These by-products may also affect the culture environment such that they might lead to early cell death even though sufficient nutrients are available or are still being provided.
Two cases can be considered in fed-batch cultures: the production of a growth-associated product and the production of a non-growth-associated product. For PHA production, a non-growth-associated product is verified when fed-batch with C. necator (formerly known as Wautersia eutropha, Ralstonia eutropha, Alcaligenes eutrophus, or Hydrogenomonas eutropha) is used. A culture with this microorganism should have two phases: a growth phase in which the cells are grown to the required concentration, and a production phase (in most cases under limitation of an essential nutrient, like nitrogen, phosphorus, oxygen, etc.) in which a carbon source and other requirements for production are fed to the bioreactor [46].
Table 1 summarizes the results of PHA production by HCDC fed-batch operations (without cell recycle) in terms of PHA content, productivity of PHAs, total biomass concentration, type of carbon source, and concentration of the carbon source solution.
Table 1 shows that efforts to attain high cell density by C. necator fed-batch cultures [7, 58, 67, 69, 75] presented high carbon source concentration in the feed solution. Ryu et al. [67] showed the importance of the glucose carbon source feeding during the fed-batch culture with the aim of reaching high cell density, high content, and high P(3HB) productivity. The glucose feeding (monitored on-line) was carried out to keep the carbon concentration in the bioreactor between 0 and 20 g l−1. The best results for the contents of P(3HB), residual biomass, total biomass, and productivity by C. necator were 81 %, 42 g l−1, 221 g l−1, and 3.75 g l−1 h−1, respectively. In a similar study, Shang et al. [69] established a strategy based on the on-line glucose feeding to obtain high cell density in a culture with C. necator in the production of P(3HB). The residual glucose concentrations of 2.5, 9, 16, and 40 g l−1 were maintained in four separate cultures. The best results were obtained for the culture which kept the concentration of residual glucose at 9 g l−1, reaching 208 g l−1 of total biomass, 69 g l−1 of residual biomass, 139 g l−1 of P(3HB), and 3.1 g l−1 h−1 of P(3HB) productivity. Kim et al. [32], after many trials with glucose feeding strategies, reported that keeping the glucose concentration between 10 and 25 g l−1 in the production phase is important for obtaining high productivity. Ienczak et al. [27] also have shown that the maintenance of the carbon source concentration in the media during culture is important. These authors showed that the carbon source pulse should not be performed upon carbon source exhaustion, but rather when the carbon source is close to its half-velocity coefficient for the substrate saturation concentration (Ks), values around 10 g l−1.
Cavalheiro et al. [7] established an HCDC for P(3HB-co-4HB) and P(3HB-4HB-3HV) production by C. necator using fed-batch mode (Table 1). The main carbon source was biodiesel waste glycerol (GRP) (1,147.5 g l−1) and the culture was supplemented with γ-butyrolactone (GBL) and propionic acid (PA). The best result showed a PHA content of 36.9 %, PHA concentration of 16.7 g l−1, 4HB content of 43.6 %, and 3HV content of 6.0 %. Total biomass concentration at the end of the culture was 45 g l−1. This is the only work presented in Table 1 that used waste to attain high cell density.
Burkholderia sacchari is a strain capable of producing PHA on both balanced and unbalanced culture media. Pradella et al. [58] performed an experiment in an airlift bioreactor for high-cell-density P(3HB) production from B. sacchari IPT 189 under limited nitrogen conditions. Total biomass, residual biomass, and P(3HB) production were 150, 87, and 63 g l−1, respectively.
Fed-batch cultures for obtaining high PHA productivity must have appropriate feeding strategies for the carbon source, because limitation of the carbon source in the production phase can lead to the degradation of the intracellular polymer owing to PHA depolymerase enzyme activity. Many research groups have therefore focused on the supply carbon source rather than that of the culture. For example, Rocha et al. [66] cultivated B. sacchari for the production of P(3HB-co-3HV) by means of feeding the best carbon source (C) to propionic acid (PA) ratio (C/PA) (Table 1). The authors concluded that concentrations of higher sucrose (250 g l−l) and PA in the C/PA ratio resulted in higher polymer production (45 %), total biomass concentration of 221 g l−1, and productivity of 1.04 g l−1 h−1 [66]. These results show that the carbon source concentration in the solution fed during a fed-batch culture is important for the polymer production.
Some wild-type strains are capable of growth-associated production of PHA. Yamane et al. [75] established such a fed-batch system using Alcaligenes latus (Table 1). The strategy used by these authors was based on feeding with sucrose (900 g l−1), inorganic element solutions, and ammonia solution by the pH-stat method (i.e., feeding the reactor in order to maintain a constant pH value), and the nutrient solutions were supplied in proportion to the ratio of their consumption rates, thereby maintaining the nutrient concentrations at nearly constant levels during fermentation. This approach afforded a residual biomass of 71.6 g l−1, total biomass of 143 g l−1, and polymer productivity of 3.97 g l−1 h−1. It is important to highlight that this work also mentions the importance of both the nitrogen residual concentration feed and the carbon source feed in obtaining high PHA productivity. Aragão et al. [3] reported the differences between P(3HV-co-3HB) production by cultures of C. necator, a microorganism that presented non-growth-associated PHA production, with and without nitrogen source feeding. Improvements in productivity were obtained in cultures fed with nitrogen and the proportion of 3HV incorporated was approximately twofold higher in these cultures (0.46 g l−1 h−1 without nitrogen supply, and 0.72 g l−1 h−1 with nitrogen supply).
Studies on PHA production have been conducted regarding glucose feeding [69] and the feeding of glucose and an organic acid [33] in order to reach high cell density and high productivity. Chen et al. [9] explored P(3HB-co-3HV) production by A. hydrophila 4AK4 from glucose and lauric acid, aiming at establishing an efficient large-scale production. A 20,000-l bioreactor was used for three fed-batch cultures, which differed in the time point of the pulses (residual glucose at 10 or 20 g l−1) and limiting nutrient (nitrogen or phosphorus). The results pointed to the importance of the residual carbon source concentration (20 g l−1) at the moment of the lauric acid pulses for obtaining 50 g l−1 of total biomass, 24.4 g l−1 of residual biomass, and 0.54 g l−1 h−1 polymer productivity.
Medium-chain-length PHAs (PHAMCL) can also be produced in HCDCs. These PHAs are made up of monomers with 6–14 carbon atoms and have different physical and mechanical characteristics compared to short-chain-length PHAs (PHASCL), which have 3–5 carbon atoms in the main chain. It is worth pointing out that P(3HB) belongs to the latter group. Lee et al. [39] established a strategy for obtaining high cell density with Pseudomonas putida from a culture in oleic acid for PHAMCL production by means of phosphorus limitation. The trials were carried out in a 20-l bioreactor with initial volume of 1.4 l and feeding of 15 g l−1 of oleic acid in pulses. The strategy was based on pH-stat for the growth phase and dissolved oxygen (DO)-stat (the same as pH-stat, i.e., feeding the medium in order to maintain a constant DO concentration) for the production phase. The best results were a residual biomass of 69.1 g l−1, total biomass of 141 g l−1, 51 % PHAs, and a productivity of 1.9 g l−1 h−1.
Sun et al. [70] recently studied the production of PHAMCL by P. putida KT2440 to attain high cell density. Different cultures were performed to compare a single-stage carbon-limited process with a two-stage process incorporating a nitrogen-limited stage. The results indicated that in the single-stage process, exponential feeding of nonanoic acid resulted in high PHA productivity and content but nitrogen limitation could perhaps further increase the synthesis of PHA. The results for productivity, PHA content, and total biomass concentration were 1.44 g l−1 h−1, 66.9 %, and 56 g l−1, respectively. It is important to highlight that hydroxynonanoate (C9) and hydroxyheptanoate (C7) PHA subunits were detected. Diniz et al. [14] studied three strategies for the production of PHAMCL by using HCDC of P. putida. The first was based on a fed-batch culture by pulses of the carbon source, the second was continuously fed with carbon source during linear growth of the residual biomass, and the third involved exponential feeding of the carbon source. These authors reported that, when a carbon source-limiting flow was used (i.e., the last two strategies), lower specific growth rates were observed. However all strategies yielded high cell density (about 30 g l−1 of Xr). Furthermore, in all strategies the carbon source concentration was lower than 1 g l−1 at the end of the growth phase and the biomass at the end of the culture had a low PHAMCL content (3–15 %).
In another study aiming at high-cell-density P(3HB) production by Bacillus megaterium, Kulpreecha et al. [35] (Table 1) used the feeding strategy of different C/N ratios (C = carbon source sucrose or sugar cane molasses and N = nitrogen source urea or ammonium sulfate). The feeding strategy for the fed-batch culture was based on DO- and pH-stat. A C/N ratio of 10 for sugar cane molasses and urea, with pH-stat set at 7.00 and DO-stat set at 60 %, afforded a residual biomass, total biomass, and productivity at the end of the culture of 42.1 g l−1, 72 g l−1, and 1.2 g l−1 h−1, respectively. It is important to highlight that is the only work presented in Table 1 that aimed to attain high cell density by using a renewable carbon source (sugar cane molasses).
Most of the studies [7, 58, 67, 69, 75] presented in Table 1 used higher carbon source concentrations to obtain high PHA concentrations, whereas when moderate concentrations were used (500 g l−1 [27], 400 g l−1 [35], and 250 g l−1 [66]) the lowest PHA concentrations were observed for fed-batch cultures.
Fed-batch culture with cell recycle
The cultures conducted through the fed-batch system can have inhibitory metabolite accumulation or limiting concentration of nutrients. The efficacy of several microbial cultures can, nevertheless, be increased by removing cell-growth-inhibiting metabolites [44]. One way of reducing the interference of these metabolites on microorganism growth is by using the internal or external cell recycle system. Internal recycle means a situation where a fraction of the cells is kept in the bioreactor through a simple sedimentation or by filtration inside the bioreactor, in order to retain the biomass, aiming at removing liquid from the bioreactor. In the case of external recycle, the effluent liquid circulates through a cell separator (e.g., filters, membranes, centrifuges, sedimentators, hydrocyclones) so a flow rich in cells returns to the bioreactor, while the other flow (filtered or permeated) leaves virtually free of cells [46]. Next, the bioreactor is re-fed with the fresh culture medium.
The traditional methods of separating cells from fermentation broths are conventional filtration, sedimentation, and centrifugation. Since cells are highly hydrated, have a low specific gravity, and tend to be rather glutinous in character, these methods of solid/liquid separation are generally problematic and inefficient. Cross-flow filtration on membranes has been proposed as an alternative method for the separation of cells and/or product from fermentation broths. The retentate is recirculated through the filtration module, and the filtrate (permeated) is continuously removed. Permeates are continually removed from the module, without accumulation within the filtration system with a substantial increase in concentration of solids in the retentate stream [20].
There are no works in the literature about PHA production with cell recycle by wild-type strain, only by recombinant E. coli [1], and this study will be presented in Sect. “Genetically modified organisms to produce polyhydroxyalkanoates”.
Repeated batch culture with and without cell recycle
Repeated or sequential batch culture without cell recycle is a system where the downtime for cleaning, medium preparation, and sterilization is eliminated, making the process more productive, besides not being restricted by the work volume. This system consists of removing a partial volume from the bioreactor’s work volume so that the same volume is immediately fed with the fresh culture medium. Repeated batch culture is a well-known method for enhancing the productivity of microbial cultures because it skips the turnaround time and the lag phase, thus increasing the process productivity [24, 59]. This method has been a very useful approach to increase the yields of several processes, such as the production of hyaluronic acid [24], biohydrogen [43], and lipase [76].
For P(3HB) production by C. necator, Khanna and Srivastava [31] investigated the repeated batch culture without cell recycle. Cultures in batch and repeated batch modes were carried out comparatively in a 7-l bioreactor with an initial volume of 4 l at 30 °C and pH 7.00. The system was characterized by the removal of 20 % (v/v) of the bioreactor medium when the residual glucose concentration was close to 10 g l−1. This procedure was carried out twice with the feeding of fresh medium (fructose solution stock with a concentration of 300 g l−1 and others nutrients). The results obtained for the repeated batch process, compared to the batch process, show an increase in productivity and total biomass concentration from 0.15 to 0.42 g l−1 h−1 and from 21 to 50 g l−1, respectively. It must be noted that the system used by these authors has the disadvantage of eliminating the cells from the bioreactor, which would cause a lower P(3HB) production in the production phase.
The cell recycle system, together with the repeated batch process, enables the achievement of a higher cell concentration and productivity in the process [52]. There are few studies about PHA production by repeated batch with cell recycle; on the other hand, for other products this process is well documented. Oh et al. [52] produced lactic acid through cell recycle repeated batch fermentation. Their results also indicated that only 26 % of the yeast extract dosage, compared with conventional batch fermentation, should be required to produce the same amount of lactic acid, which might result in a considerable reduction of production costs. The maximum cell concentration in their experiment was greater than 28 g l−1, which might contribute to the improvement of the productivity and reduction of nutrient supplementation. Others examples of the repeated batch recycle system include the cell immobilization with media recirculation for gibberellic acid [47] production and for fructooligosaccharide (FOS) fermentation [51].
Ibrahim and Steinbüchel [26] reported a repeated batch process with cell recycle to produce PHA. In this work different fermentation strategies for the cultivation of a new P(3HB)-accumulating thermophilic bacterium, Chelatococcus sp. strain MW10, were performed with the aim of achieving high-cell-density growth and high P(3HB) productivity. Enhanced cultivation was achieved by a repeated batch fermentation technique (42-l scale). The time for repeated batch was designed (50-h cultivation batches) and cultivation was started with 25 l (with 4 % [v/v] inoculum size), with a pulse fed, and a new batch was started. At the end of the culture (156 h), an HCDC of up to 115 g l−1, P(3HB) content of 11.8 %, and 13.6 g l−1 of P(3HB) were produced [26].
Continuous culture with and without cell recycle
A continuous production process is of great commercial interest owing to its high productivities, especially for strains with high maximum specific growth rates. The continuous system is characterized by continuously feeding the culture medium at a given constant flow, the work volume being kept constant by continuously removing the fermented broth. The microbial population in steady state continuous cultures is in a constant physiological state controlled by the variation in the residence time (relation between work volume and substrate feed flow to the system) in the bioreactor, amount of substrate fed into the bioreactor, and cell recycle level (when applicable) [62].
Table 2 summarizes the results of PHA production by HCDC-type continuous mode operations (without cell recycle) in terms of the PHA content, productivity of PHAs, total biomass concentration, type of carbon source, and concentration of the carbon source solution, using different microorganisms.
Ramsay et al. [60] investigated P(3HB) and P(3HB-co-3HV) production in single-stage cultures by C. necator and two-stage continuous cultures by A. latus, respectively (Table 2). In the single-stage continuous cultures, C. necator DSM 545 accumulated to 33 % of its dry mass in P(3HB) when fed with a medium containing glucose, mineral salts, and nitrogen limitation. P(3HB-co-3HV) was produced in similar experiments with A. latus (two-stage continuous cultures) when propionic or valeric acid was added to the feeding containing sucrose as the main carbon source. In the first culture stage in a chemostat, feeding PA at 5 g l−1 resulted in a copolymer with a molar fraction of 20 % of 3HV units. Replacing PA (three carbons) with valeric acid (five carbons) led to a higher content of 3HV units in the copolymer. When high PA concentrations were fed (8.5 g l−1), sucrose assimilation was inhibited. In this case, transferring the bioreactor effluent to a second stage led to the complete consumption of the sugar and afforded P(3HB-co-11-mol %3HV), representing 58 % of the total biomass.
Khanna and Srivastava [30] (Table 2) tested the continuous culture of C. necator at 30 °C, pH 7.00 using fructose as a carbon source. In this study, a two-stage continuous system was established, the first one being used for cell growth and the second for the P(3HB) production process. The bioreactor used for the first stage contained a high nitrogen concentration to avoid the limitation of this nutrient. The second bioreactor was fed with the effluent of the first one and with a concentrated fructose solution and nitrogen limitation occurred. The purpose of establishing a two-stage continuous process was justified by the authors because the maximum P(3HB) production takes place when nutrient (nitrogen) limitation occurs. The results obtained showed that only 20 g l−1 of the total biomass, 18.5 g l−1 of residual biomass, and 1.5 g l−1 of P(3HB) were accumulated with this system.
Du et al. [15] studied P(3HB) production by C. necator in a two-stage continuous culture (Table 2). The first stage was characterized by the microorganism growth phase, whereas the second stage occurred with nitrogen limitation for P(3HB) production. Among the values tested for the dilution rates (reciprocal of the residence time values), 0.075 h−1 showed the best results for P(3HB) percentage (73 %) and productivity (1.25 g l−1 h−1). The total biomass was around 50 g l−1 and the residual biomass was 13.5 g l−1.
A five-stage system for PHA production by C. necator was reported by Atlić et al. [2]. The first bioreactor was used for biomass growth and the other four bioreactors for PHA production. It is important to emphasize that each bioreactor presented a different dilution rate and residence times. The last bioreactor worked with a higher volume in comparison to the previous one in order to increase the substrate conversion. When the steady state was reached, the residual biomass, PHB content, and volumetric productivity were 18.6 g l−1, 77 %, and 1.85 g l−1 h−1, respectively.
It should be noted that P(3HB) is an intracellular product and, hence, the higher biomass density, the higher polymer density that could be accumulated. However in the literature there are no reports on continuous PHA production processes using cell recycle. The continuous culture with cell recycle allows for the operation at specific feed flows above the maximum specific growth rate of the microorganism, going beyond the limit imposed by ordinary continuous systems where the bioreactor maximum medium average output flow is determined by the maximum specific growth rate. Therefore, in systems with cell recycle, higher productivities could be expected [18].
In the case of PHAMCL, a Swiss group reported the continuous, growth-associated production of poly(3-hydroxyalkanoate-co-3-hydroxyalkenoates) in one-stage chemostat cultures of P. putida ATTC 29147 in a single continuous stirred tank reactor (CSTR). The applied substrates encompassed 5-phenylvalerate, octanoate, and 10-undecenoate. Multiple limited nutrient growth conditions were chosen at a dilution rate of D = 0.1 h−1. Different mixtures of the substrates in the feed resulted in the formation of copolyesters with varying compositions and different amounts of aromatic and unsaturated side chains that make the products accessible for further modification. The authors report that the steady state conditions in a continuous culture provide a strategy specially suited for the production of tailored PHA copolymers [21].
A few studies on a two-stage continuous system were conducted with other microorganisms than C. necator. In the case of poly(3HB-co-4HB) production, Delftia acidovorans P4a was cultivated in mixtures of acetic acid and GBL; here, poly(3HB-4HB) copolymers with a molar fraction of 2.7–19 % 4HB were obtained. The authors established that especially in the case of toxic substrates like acetic acid and GBL, the two-stage continuous production strategy is very convenient [49].
Tan et al. [71] developed an unsterile two-stage (bioreactor I and II) continuous process for P(3HB) production by Halomonas TD01, a halophilic strain. The start-up of the process was performed in bioreactor I for 2 weeks. Thereafter, the culture of bioreactor I was pumped to bioreactor II. Cell growth in bioreactor II was under nitrogen limitation conditions. Temperature, pH, and DO were kept at 37 °C, 9.00, and 50 %. The total biomass was 20 g l−1 (second stage) and PHA content was 52 % (second stage). The concentration of glucose fed into bioreactor I was 400 and 200 g l−1 for the first 2 days and days 3–14, respectively. For bioreactor II, a glucose solution (100 g l−1) was fed during all the operation time (14 days).
Genetically modified organisms to produce polyhydroxyalkanoates
After many years of screening for PHA producers, several useful wild-type strains were identified. However, few strains have presented potential for industrial scale use owing to low productivity and yield. In this sense, many efforts have been made to maximize productivity, kinetics parameters, and the capability to uptake renewable carbon source by these wild-type strains. In the past, the main focus of this line of research was on bioprocess engineering to increase productivity and the kinetic parameters. However, progress in metabolic engineering research and the combination of metabolic engineering and bioprocess engineering have been exploited. In this context, some research groups have focused on recombinant strain development for PHA production by the insertion of genes responsible for the PHA production in strains that lack this capability, or by the knock out of genes in wild-type strains focused on the increase of the yield and productivity, etc. At this moment few recombinant strains (E. coli and C. necator) have presented potential for industrial application on the basis of results observed on bench and pilot scale.
The best representative of this class of modified strains is E. coli. It holds promise as a source of economical PHA production because of its high productivity, no need for nutrient limitation, and the lack of a depolymerase system degrading the synthesized polymer [16, 19, 37]. Since all wild-type E. coli strains are unable to synthesize PHA, these organisms have to be equipped with at least the PHA-encoding gene (phbCAB operon) [32]. After the cloning of the C. necator PHA biosynthesis genes in E. coli [53], recombinant E. coli has been investigated for the production of P(3HB) because it has several advantages over other bacteria [16]. In particular, it has been shown that recombinant E. coli harboring the heterologous PHA biosynthesis genes has several advantages over wild-type PHA producers; these advantages include a wide range of utilizable carbon sources, accumulation of a large amount of P(3HB) with a high level of productivity [16, 19, 37].
Table 3 summarizes the results of P(3HB) production by recombinant E. coli under HCDC-type conditions in terms of productivity of PHA, total biomass concentration, type of carbon source, and concentration of the carbon source solution.
The production of P(3HB) by recombinant E. coli aiming at high cell density and high polymer productivity was studied in a fed-batch by Wang and Lee [73]. They carried out four cultures fed with different concentrations of thiamine (15, 30, 250, and 300 mg l−1) and 20 g l−1 of glucose. According to the authors, the use of recombinant E. coli for high-density P(3HB) production showed good results (residual biomass = 43.7 g l−1, total biomass = 156 g l−1, and productivity = 2.4 g l−1 h−1, with the use of 30 mg l−1 of thiamine), mainly because the biopolymer production is associated with growth and there is no PHA depolymerase enzyme in the cell.
Wong and Lee [74] used recombinant E. coli in an HCDC strategy for P(3HB) production using whey as the substrate. Two cultures were carried out fed with powder-milk whey [65 % (w/v)] and a whey solution (210 g l−1), respectively. The productivity was virtually the same for both cultures; however, the percentage of biopolymer and the conversion rates were higher for the second culture. The authors also point out that, owing to issues with the solubility of powder-milk whey, the downstream process could be hindered, because there would be no way of separating the substrate from the product during the cell separation step, which would result in low polymer purity.
In the literature, there is one study that reports the use of external cell recycle for P(3HB) production by recombinant E. coli GSC 4401 in a repeated fed-batch mode with pH-stat strategy for whey feeding [1]. Two fed-batch cultures with cell recycle were carried out, where run A was fed with a whey solution containing 210 g l−1 of the lactose carbon source, whereas run B was fed with a whey solution containing 280 g l−1 of the lactose carbon source. The external cell recycle system was carried out with a membrane filter. The results showed that run B had better total P(3HB) production results (168 g l−1) and P(3HB) productivity (4.6 g l−1 h−1) than run A. Run B also had total biomass concentration of 194 g l−1 and residual biomass of 25.2 g l−1. High productivity was obtained with this cell recycle system owing to the advantages of retaining the cells in the bioreactor, draining the fermented media (with low carbon source concentration), and feeding concentrated carbon source solution into the bioreactor. It is important to emphasize that PHA is an intracellular product, and by using the cell recycle system, it is possible to increase the PHA content owing to the larger carbon source consumption without volume variations in the bioreactor, and without dilution of the cell/PHA concentration.
Another representative of the PHA recombinant strain producers is C. necator. As notice before, this wild-type strain is known as the model organism for PHA production, mainly because it can store large amounts of PHA under nutrient limitation [27, 67]. However, this strain is able to produce only PHASCL. Focused on the replacement of petroleum-based plastics, Riedel et al. [63] modified C. necator for the production of poly(hydrobutyrate-co-hydroxyhexanoate) (P(HB-co-HHx), a PHAMCL product, by palm oil (PO) fermentations. Four different cultures were performed: batch, extended batch (with one pulse of palm oil), fed-batch (pH control with NH4OH and PO pulses), fed-batch (urea as a nitrogen source and PO pulses). The fed-batch with urea as a nitrogen source had the best results for PHA production (102 g l−1), PHA content (73.5 % with 19 mol % of HHx), total biomass concentration (138.8 g l−1), and yield (0.78 gPHA (gPO)−1). The authors pointed out that the plasmid maintenance was robust in HCDCs, and the results indicated that the plasmid-borne PHA production genes were still present at the end of the culture.
There are others reports about recombinant C. necator [17, 22]. Povolo et al. [57] constructed a C. necator recombinant strain capable of growing and producing PHA in lactose, hydrolyzed whey, and directly in whey permeate, with the aim of developing a useful strain able to use a cheap carbon source and with an improved PHA production by removing the polymer degradation mechanism. The recombinant strain produced 30 % of PHB and the capability to uptake lactose, glucose as the sole carbon source, and in a hydrolyzed whey permeate and non-hydrolyzed whey permeate solution. Another contribution of this work was the insertion of the lac operon within the phaZ gene that may reduce the amount of PHA depolymerized by the cell, thus improving the final polymer yield.
One way of making PHA synthesis cheaper could be the availability of bacterial PHB biosynthesis genes and the development of methods for transferring them into eukaryotic systems. In this way it will be possible to equip higher organisms like plants or yeasts with the ability to synthesize PHA. Yeasts are traditionally used in biotechnological procedures; yeast metabolism and nutrition physiology are sufficiently flexible to facilitate such production; wild types are not ecologically harmful [6].
It has been shown previously that poly(hydroxybutyrate) is synthesized in the cytosol of S. cerevisiae if the PHASCL polymerase from R. eutropha is expressed in this cell compartment [36]. This finding indicates that native S. cerevisiae is capable of synthesizing monomers of the correct enantiomeric configuration for the polymerase enzyme. It has been demonstrated recently that PHAMCL can be synthesized in the peroxisomes of S. cerevisiae and Pichia pastoris when the PHAMCL polymerase from Pseudomonas aeruginosa is expressed and targeted into this organelle [55, 56]. PHAMCL can be synthesized in the cytosol if the PHAMCL polymerase from Pseudomonas oleovorans is expressed in S. cerevisiae [78]. Nevertheless the productivity and yield of these strains is very low, and its scaling-up becomes difficult.
A wide range of strains have been modified for PHA production: A. hydrophila 4AK4 [42], M. extorquens [23], Pseudomonas strains [25, 45], and others. However the potential scale-up of these strains is not possible at this point because of the low productivity and yield.
Nowadays synthetic biology and engineering principles applied to bioprocess development have been studied by some research groups. This field of science involves the disassembly, redesign, and standardization of existing biological components (parts, devices, and genetic circuits) with the aim of creating novel genetic circuits, biosynthetic pathways, and living system from abiotic components [41, 50]. As describe by Zhu et al. [79] the introduction of a synthetic pathway for PHA biosynthesis into non-PHA-producing microorganisms can confer improved survival ability on host cells under adverse conditions such as starvation, desiccation, UV radiation, high osmotic pressure, and the presence of organic solvents [8]. The introduction of new biosynthetic pathways capable of producing stress-related molecules may confer redundancy for damage protection on host cells, thus increasing their stress resistance. However, the constitutive expression of enzymes involved in synthetic pathways could impose a physiological burden on the host, so an increase in the dynamic controllability of the synthetic pathway introduced, using the strategies illustrated above, seems necessary [79].
Conclusions and perspectives
Industrial PHA production only becomes economically viable when high cell densities and high polymer productivities are reached with low costs. In recent years, advanced techniques to control and monitor biotechnological processes have appeared, aiming at reaching high cell density in less costly processes. However, key factors in P(3HB) production must be taken into consideration for HCDCs: substrate and microorganism.
As a result of the high production costs of this polymer compared to those of the petrochemical polymers, there is a need for more productive strains and feeding strategies with low-cost substrates such as agroindustrial wastes and biomass. These substrates have low (20–40 g l−1) or moderate (80–200 g l−1) carbon source concentrations, and the hydrolyzed biomass has hexose and pentose sugars, which make the attainment of HCDCs difficult. For this reason, appropriate strategies in fed-batch, continuous, or repeated batch culture with cell recycle are necessary to attain HCDC. When hydrolyzed biomass is used for P(3HB) production, co-cultures can be used to uptake both substrates (hexose and pentose) present in a raw material biomass and convert them into polymer.
In general, recombinant microorganisms showed more competitive conditions for industrial processes in relation to wild-type strain owing to the absence of depolymerase enzyme, absence of the necessity of nutrient limitation, and the capacity to produce higher P(3HB) content in a short time. These factors are favorable for enhanced productivity and PHA production. On the other hand, recombinant microorganisms have shown instability for industrial processes (need of antibiotics and inductors for expressing the genes of interest). Therefore, creating strains with this genetic material inserted into DNA is needed for the stability of this process in an industrial plant.
Abbreviations
- (3HV):
-
3-Hydroxyvalerate
- AP:
-
Analytical purity
- (C/PA):
-
Carbon source/propionic acid ratio
- C/N:
-
Carbon source sucrose or sugar cane molasses/nitrogen source urea or ammonium sulfate
- DO:
-
Dissolved oxygen
- FOS:
-
Fructooligosaccharides
- GBL:
-
γ-Butyrolactone
- GRP:
-
Waste glycerol
- HCDC:
-
High-cell-density cultures
- MM:
-
Mineral medium
- P(3HB):
-
Poly(3-hydroxybutyrate)
- P(3HB-4HB):
-
Poly(3-hydroxybutyrate–4-hydroxybutyrate)
- P(3HB-4HB-3HV):
-
Poly(3-hydroxybutyrate–4-hydroxybutyrate–3-hydroxyvalerate)
- P(3HB-co-3HV):
-
Poly(3-hydroxybutyrate-co-3-hydroxyvalerate)
- P(3HB-co-4HB):
-
Poly(3-hydroxybutyrate-co-4-hydroxybutyrate)
- %PHAs:
-
Polyhydroxyalkanoate cell content
- PA:
-
Propionic acid
- PHA:
-
Polyhydroxyalkanoates
- PHASCL :
-
Short-chain-length polyhydroxyalkanoates
- PHASMCL :
-
Medium-chain-length polyhydroxyalkanoates
- r PHAS :
-
Productivity of polyhydroxyalkanoates
- STR:
-
Stirred tank reactors
- Xr:
-
Residual biomass
- Xt:
-
Total biomass concentration
References
Ahn WS, Park SJ, Lee SY (2001) Production of poly(3-hydroxybutyrate) from whey by cell recycle fed-batch culture of recombinant Escherichia coli. Biotechnol Lett 23:235–240. doi:10.1023/A:1005633418161
Atlić A, Koller M, Scherzer D, Kutschera C, Grillo-Fernandes E, Horvat P, Chiellini E, Braunegg G (2011) Continuous production of poly([R]-3-hydroxybutyrate) by Cupriavidus necator in a multistage bioreactor cascade. Appl Microbiol Biotechnol 91:295–304. doi:10.1007/s00253-011-3260-0
Aragão GMF, Lindley ND, Uribelarrea JL, Pareilleux A (1996) Maintaining a controlled residual growth capacity increases the production of polyhydroxyalkanoate copolymers by Alcaligenes eutrophus. Biotechnol Lett 18:937–942
Aragão GMF, Schmidell W, Ienczak JL, Schmidt FC, Dalcanton F, Fiorese ML, Rodrigues R, Deucher R, Marangoni C (2009) Preparation of polyhydroxyalkanoates from a citric residue. WO2009/149529 A1
Bengtssona S, Piscob AR, Johanssona P, Lemos PC, Reis MAM (2010) Molecular weight and thermal properties of polyhydroxyalkanoates produced from fermented sugar molasses by open mixed cultures. J Biotechnol 147(3):172–179
Breuer U, Terentiev Y, Kunze G, Babel W (2002) Yeasts as producers of polyhydroxyalkanoates: genetic engineering of Saccharomyces cerevisiae. Macromol Biosci 2:380–386. doi:10.1002/1616-5195(200211)
Cavalheiro JMBT, Raposo SR, Almeida MCMD, Sevrin MTCC, Grandfils C, Fonseca MMR (2012) Effect of cultivation parameters on the production of poly(3-hydroxybutyrate-co-4-hydroxybutyrate) and poly(3-hydroxybutyrate-4-hydroxybutyrate-3-hydroxyvalerate) by Cupriavidus necator using waste glycerol. Bioresourc Technol 111:391–397. doi:10.1016/j.biortech.2012.01.176
Chen GQ (2009) A microbial polyhydroxyalkanoates (PHA) based bioand materials industry. Chem Soc Rev 38:2434–2446
Chen GQ, Zhang G, Park SJ, Lee SY (2001) Industrial scale production of poly(3-hydroxybutyrate-co-3-hydroxyhexanoate). Appl Microbiol Biotechnol 57:50–55. doi:10.1007/s002530100755
Choi J, Lee SY (1997) Process analysis and economic evaluation for poly(3-hydroxybutyrate) production by fermentation. Bioprocess Eng 17:335–342. doi:10.1007/s004490050394
Choi J, Lee SY, Han K (1998) Cloning of the Alcaligenes latus polyhydroxyalkanoate biosynthesis genes and use of this genes for enhanced production of poly(3-hydroxybutyrate) in Escherichia coli. Appl Environ Microbiol 64:4879–4903
Dalcanton F, Ienczak JL, Fiorese ML, Aragão GMF (2010) Produção de poli(3-hidroxibutirato) por Cupriavidus necator em meio hidrolisado de amido de arroz com suplementação de óleo de soja em diferentes temperaturas. Quim Nova 33(3):552–556. doi:10.1590/S0100-40422010000300011
Dias JML, Lemos PC, Serafim LS, Oliveira C, Eiroa M, Albuquerque MGE, Ramos AM, Oliveira R, Reis MAM (2006) Recent advances in polyhydroxyalkanoate production by mixed aerobic cultures: from the substrate to the final product. Macromol Biosci 6:885–906. doi:10.1002/mabi.200600112
Diniz SC, Taciro MK, Gomez JGC, Pradella JGC (2004) High-cell-density cultivation of Pseudomonas putida IPT 046 and medium-chain-length polyhydroxyalkanoate production from sugarcane carbohydrates. Appl Biochem Biotechnol 119:51–69
Du G, Chen J, Yu J, Lun S (2001) Continuous production of poly-3-hydroxybutyrate by Ralstonia eutropha in a two-stage culture system. J Biotechnol 88:59–65. doi:10.1016/S0168-1656(01)00266-8
Fidler S, Dennis D (1992) Polyhydroxyalkanoate production in recombinant Escherichia coli. FEMS Microbiol Rev 103:231–236
Fukui T, Doi Y (1998) Efficient production of polyhydroxyalcanoates from plant oils by Alcaligenes eutrophus and its recombinant strain. Appl Microbiol Biotechnol 49:333–336
Grón S, Biedermann K, Emborg C (1995) Enzyme production in a cell recycle fermentation system evaluated by computer simulations. Bioprocess Eng 13:59–68
Hahn SK, Chang YK, Lee SY (1995) Recovery and characterization of poly(3-hydroxybutyric acid) synthesized in Alcaligenes eutrophus and recombinant Escherichia coli. Appl Environ Microbiol 61:34–39
Hanisch A (1986) Cell harvesting. In: McGregor WC (ed) Membrane separations in biotechnology. Marcel Dekker, New York
Hartmann R, Hany R, Geiger T, Egli T, Witholt B, Zinn M (2004) Tailored biosynthesis of olefinic medium-chain-length poly[(R)-3-hydroxyalkanoates] in Pseudomonas putida GPo1 with improved thermal properties. Macromolecules 37:6780–6785. doi:10.1021/ma040035+
Hoefel T, Wittmann E, Reinecke L, Weuster-Botz D (2010) Reaction engineering studies for the production of 2-hydroxyisobutyric acid with recombinant Cupriavidus necator H 16. Appl Microbiol Biotechnol 88:477–484. doi:10.1007/s00253-010-2739-4
Höffer P, Vermetteb P, Groleaua D (2011) Production and characterization of polyhydroxyalkanoates by recombinant Methylobacterium extorquens: combining desirable thermal properties with functionality. Biochem Eng J 54:26–33. doi:10.1016/j.bej.2011.01.003
Huang WC, Chen SJ, Chen TL (2008) Production of hyaluronic acid by repeated batch fermentation. Biochem Eng J 40:460–464. doi:10.1016/j.bej.2008.01.021
Huisman GW, Wonin E, Koning G, Preusting H, Witholt B (1992) Synthesis of poly(3-hydroxyalkanoates) by mutant and recombinant Pseudomonas strains. Appl Microbiol Biotechnol 38:1–5
Ibrahim MHA, Steinbüchel A (2010) High-cell-density cyclic fed-batch fermentation of a poly(3-hydroxybutyrate)-accumulating thermophile, Chelatococcus sp. strain MW10. Appl Environ Microbiol 76(23):7890–7895. doi:10.1128/AEM.01488-10
Ienczak JL, Quines LK, Melo AA, Brandellero M, Mendes CR, Schmidell W, Aragão GMF (2011) High cell density strategy for poly(3-hydroxybutyrate) production by Cupriavidus necator. Braz J Chem Eng 28(4):585–596. doi:10.1590/S0104-66322011000400004
Kahar P, Tsuge T, Taguchi K, Doi Y (2004) High yield production of polyhydroxyalkanoates from soybean oil by Ralstonia eutropha and its recombinant strain. Polym Degrad Stab 83:79–86. doi:10.1016/S0141-3910(03)00227-1
Khanna S, Srivastava AK (2006) Computer simulated fed-batch cultivation for over production of PHB: a comparison of simultaneous and alternate feeding of carbon and nitrogen. Biochem Eng J 27:197–203. doi:10.1016/j.bej.2005.08.006
Khanna S, Srivastava AK (2008) Continuous production of poly-β-hydroxybutyrate by high-cell-density cultivation of Wautersia eutropha. J Chem Technol Biotechnol 83:799–805. doi:10.1002/jctb.1868
Khanna S, Srivastava AK (2005) Repeated fed-batch cultivation of Ralstonia eutropha for poly(β-hydroxybutyrate) production. Biotechnol Lett 27:1401–1403. doi:10.1007/s10529-005-0688-2
Kim BS, Lee SC, Lee SY, Chang HN, Chang YK, Woo SI (1994) Production of poly(3-hydroxybutyric acid) by fed-batch culture of Alcaligenes eutrophus with glucose concentration control. Biotechnol Bioeng 43:892–898. doi:10.1002/bit.260430908
Klinke S, Ren Q, Witholt B, Kessler B (1999) Production of medium-chain-length by recombinant Escherichia coli poly(3-hydroxyalkanoates) from gluconate. Appl Environ Microbiol 65(2):540
Koyama N, Doi Y (1995) Continuous production poly(3-hydrobutyrate-co-3-hydroxyvalerate) by Alcaligenes eutrophus. Biotechnol Lett 17(3):281–284
Kulpreecha S, Boonruangthavorn A, Meksiriporn B, Thongchul N (2009) Inexpensive fed-batch cultivation for high poly(3-hydroxybutyrate) production by a new isolate of Bacillus megaterium. J Biosci Bioeng 107(3):240–245. doi:10.1016/j.jbiosc.2008.10.006
Leaf TA, Peterson MS, Stoup SK, Somers D, Srienc F (1996) Saccharomyces cerevisiae expressing bacterial polyhydroxybutyrate synthase produces poly-3-hydroxybutyrate. Microbiology 142:1169–1180
Lee SY (1996) Bacterial polyhydroxyalkanoates. Biotechnol Bioeng 49:1–4. doi:10.1002/(SICI)1097-0290(19960105)49:1<1:AID-BIT1>3.0.CO;2-P
Lee SY, Chang HN (1995) Production of poly(hydroxyalkanoic acid). Adv Biochem Eng Biotechnol 52:27–58
Lee SY, Wong HH, Choi J, Lee SH, Lee SC, Chang CS (2000) Production of medium-chain-length polyhydroxyalkanoates by high-cell-density cultivation of Pseudomonas putida under phosphorus limitation. Biotechnol Bioeng 68(4):466–470. doi:10.1002/(SICI)1097-0290(20000520)68:4<466:AID-BIT12>3.0.CO;2-T
Lee WH, Loo CY, Nomura CT, Sudesh K (2008) Biosynthesis of polyhydroxyalkanoate copolymers from mixtures of plant oils and 3-hydroxyvalerate precursors. Bioresourc Technol 99:6844–6851. doi:10.1016/j.biortech.2008.01.051
Leonard E, Nielsen D, Solomon K, Prather KJ (2008) Engineering microbes with synthetic biology frameworks. Trends Biotechnol 26:674–681
Liu F, Jian J, Shen X, Chung A, Chen J, Chen GQ (2012) Metabolic engineering of Aeromonas hydrophila 4AK4 for production of copolymers of 3-hydroxybutyrate and medium-chain-length 3-hydroxyalkanoate. Bioresourc Technol 102:8123–8129. doi:10.1016/j.biortech.2011.05.074
Liu DW, Zeng RJ, Angelidaki I (2008) Enrichment and adaptation of extreme-thermophilic (70 °C) hydrogen producing bacteria to organic household solid waste by repeated batch cultivation. Int J Hydrogen Energy 33:6492–6497. doi:10.1016/j.ijhydene.2008.08.014
Märkl H, Pörther R (1998) Dialysis culture. Appl Microbiol Biotechnol 50:403–414
Matsusaki H, Abe H, Doi Y (2000) Biosynthesis and properties of poly(3-hydroxybutyrate-co-3-hydroxyalkanoates) by recombinant strains of Pseudomonas sp. 61–3. Biomacromolecules 1:17–22. doi:10.1021/bm9900040
McNeil B, Harvey LM (1990) Fermentation, a practical approach. IRL, Tokyo
Meleigy SA, Khalaf MA (2009) Biosynthesis of gibberelic acid from milk permeate in repeated batch operation by a mutant Fusarium moliniforme cells immobilized on loofah sponge. Bioresour Technol 100:374–379. doi:10.1016/j.biortech.2008.06.024
Meszaros A, Bales V (1992) A contribution to optimal control of fed-batch biochemical processes. Bioprocess Eng 7:363–367. doi:10.1007/BF00369492
Mothes G, Ackermann JU (2005) Synthesis of poly(3-hydroxybutyrate-co-4-hydroxybutyrate) with a target mole fraction of 4-hydroxybutyric acid units by two-stage continuous cultivation of Delftia acidovorans P4a. Eng Life Sci 5:58–62. doi:10.1002/elsc.200420056
Mukherji S, van Oudenaarden A (2009) Synthetic biology: understanding biological design from synthetic circuits. Nat Rev Genet 10:859–871. doi:10.1038/nrg2697
Mussatto SI, Rodrigues LR, Teixeira JA (2009) β-Fructofuranosidase production by repeated batch fermentation with immobilized Aspergillus japonicas. J Ind Microbiol Biotechnol 36:923–928. doi:10.1007/s10295-009-0570-7
Oh H, Wee YJ, Yun JS, Ryu HW (2003) Lactic acid production through cell-recycle repeated-batch bioreactor. Appl Biochem Biotechnol 107:603–613
Peoples OP, Sinskey AJ (1989) Poly-beta-hydroxybutyrate (PHB) biosynthesis in Alcaligenes eutrophus H16—identification and characterization of the Phb polymerase gene (phbC). J Biol Chem 264:15298–15303
Pirt SJ (1990) The dynamics of microbial processes: a personal view. In: Poole RK, Bazin MJ, Keevil CM (eds) Microbial growth dynamics, vol 28. IRL, Tokyo, pp 1–16
Poirier Y, Erard N, Petetot JM (2001) Synthesis of polyhydroxyalkanoate in the peroxisome of Saccharomyces cerevisiae by using intermediates of fatty acid β-oxidation. Appl Environ Microbiol 67:5254–5260
Poirier Y, Erard N, MacDonald-Comber PJ (2002) Synthesis of polyhydroxyalkanoate in the peroxisome of Pichia pastoris. FEMS Microbiol Lett 207:97–102
Povolo S, Toffano P, Basaglia M, Casella S (2010) Polyhydroxyalkanoates production by engineered Cupriavidus necator from waste material containing lactose. Bioresourc Technol 101:7902–7907. doi:10.1016/j.biortech.2010.05.029
Pradella JGC, Taciro MK, Pataquiva AY (2010) High-cell-density poly (3-hydroxybutyrate) production from sucrose using Burkholderia sacchari culture in airlift bioreactor. Bioresourc Technol 101:8355–8360. doi:10.1016/j.biortech.2010.05.046
Radmann EM, Reinehr CO, Costa JAV (2007) Optimization of the repeated batch cultivation of microalga Spirulina platensis in open raceway ponds. Aquaculture 265:118–126. doi:10.1016/j.aquaculture.2007.02.001
Ramsay BA, Ramsay JA, Lomaliza K, Chavarie C, Bataille P (1990) Production of poly-(β-hydroxybutyric-co-β-hydroxyvaleric) acid copolymers. Appl Environ Microbiol 56:2093–2098
Reddy CSK, Ghai R, Rashmi T, Kahia VC (2003) Polyhydroxyalkanoates: an overview. Bioresourc Technol 87:137–146
Richardson KR (1988) Production of beta-hydroxybutyrate polymers. European Patent EP 0114086 B1
Riedel SL, Bader J, Brigham CJ, Budde CF, Yusof ZAM, Rha C, Sinskey AJ (2012) Production of poly(3-hydroxybutyrate-co-3-hydroxyhexanoate) by Ralstonia eutropha in high cell density palm oil fermentations. Biotechnol Bioeng 109:74–83. doi:10.1002/bit.23283
Riesenberg D (1991) High-cell-density cultivation of Escherichia coli. Curr Opin Biotechnol 2:380–384
Riesenberg D, Guthke R (1999) High-cell-density cultivation of microorganisms. Appl Microbiol Biotechnol 51:422–430. doi:10.1007/s002530051412
Rocha RCS, Silva LF, Taciro MK, Pradella JGC (2008) Production of poly(3-hydroxybutyrate-co-3-hydroxyvalerate) P(3HB-co-3HV) with a broad range of 3HV content at high yields by Burkholderia sacchari IPT 189. World J Microbiol Biotechnol 24:427–431
Ryu HW, Hahn SK, Chang YK, Chang HN (1997) Production of poly(3-hydroxybutyrate) by high cell density fed-batch culture of Alcaligenes eutrophus with phosphate limitation. Biotechnol Bioeng 55(1):25–32
Serafim LS, Lemos PC, Albuquerque MGE, Reis MAM (2008) Strategies for PHA production by mixed cultures and renewable waste materials. Appl Microbiol Biotechnol 81:615–628. doi:10.1007/s00253-008-1757-y
Shang L, Jiang M, Chang HN (2003) Poly(3-hydroxybutyrate) synthesis in fed-batch culture of Ralstonia eutropha with phosphate limitation under different glucose concentrations. Biotechnol Lett 25:1415–1419
Sun Z, Ramsay JA, Guay M, Ramsay BA (2007) Carbon-limited fed-batch production of medium-chain-length polyhydroxyalkanoates from nonanoic acid by Pseudomonas putida KT2440. Appl Microbiol Biotechnol 74:69–77. doi:10.1007/s00253-006-0655-4
Tan D, Xue YS, Aibaidula G, Chen GQ (2011) Unsterile and continuous production of polyhydroxybutyrate by Halomonas TD01. Bioresourc Technol 102:8130–8136. doi:10.1016/j.biortech.2011.05.068
Turner C, Gregory ME, Thornhill N (1994) Closed-loop control of fed-batch cultures of recombinant Escherichia coli using on-line HPLC. Biotechnol Bioeng 44:819–829. doi:10.1002/bit.260440707
Wang F, Lee SY (1998) High cell density culture of metabolically engineered Escherichia coli for the production of poly(3-hydroxybutyrate) in a defined medium. Biotechnol Bioeng 58:325–328. doi:10.1002/(SICI)1097-0290(19980420)58:2/3<325:AID-BIT33>3.0.CO;2-8
Wong HH, Lee SY (1998) Poly(3-hydroxybutyrate) production from whey by high cell density cultivation of recombinant Escherichia coli. Appl Microbiol Biotechnol 50:30–33
Yamane T, Fukunaga M, Lee YW (1996) Increased PHB production by high-cell-density fed-batch culture of Alcaligenes latus, a growth associated PHB producer. Biotechnol Bioeng 50:197–202. doi:10.1002/(SICI)1097-0290(19960420)50:2<197:AID-BIT8>3.0.CO;2-H
Yang X, Wang B, Cui F, Tan T (2005) Production of lipase by repeated batch fermentation immobilized Rhizopus arrhizus. Process Biochem 40:2095–2103. doi:10.1016/j.procbio.2004.07.015
Yee L, Blanch HW (1992) Recombinant protein expression in high cell density fed-batch cultures of Escherichia coli. Biotechnol Adv 10:1550–1556
Zhang B, Carlson R, Pederson EN, Srienc F (2005) Novel synthesis routes for polyhydroxyalkanoic acids with unique properties. Polym Biocatal Biomater 900:292–301
Zhu L, Zhu Y, Zhang Y, Li Y (2012) Engineering the robustness of industrial microbes through synthetic biology. Trends Microbiol 20:94–101. doi:10.1016/j.tim.2011.12.003
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Ienczak, J.L., Schmidell, W. & de Aragão, G.M.F. High-cell-density culture strategies for polyhydroxyalkanoate production: a review. J Ind Microbiol Biotechnol 40, 275–286 (2013). https://doi.org/10.1007/s10295-013-1236-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10295-013-1236-z