
Abstract
Oxidative stress can occur in different parts of plant cells. We employed two oxidants that induce reactive oxygen species (ROS) in different intracellular compartments: methyl viologen (MV, in chloroplasts) and menadione (MD, in mitochondria). The responses of pea (Pisum sativum) leaf discs to MV or MD after 4-h incubation in dark or moderate (300 μE m−2 s−1) or high light (1200 μE m−2 s−1) were examined. Marked increase in ROS levels was observed, irrespective of compartment targeted. The levels of proline, a compatible solute, increased markedly much more than that of ascorbate or glutathione during oxidative/photo-oxidative stress, emphasizing the importance of proline. Further, the activities and transcripts of enzymes involved in biosynthesis or oxidation of proline were studied. An upregulation of biosynthesis and downregulation of oxidation was the basis of proline accumulation. Pyrroline-5-carboxylate synthetase (P5CS, involved in biosynthesis) and proline dehydrogenase (PDH, involved in oxidation) were the key enzymes regulated under oxidative stress. Since these two enzymes—P5CS and PDH—are located in chloroplasts and mitochondria, respectively, we suggest that proline metabolism can help to mediate inter-organelle interactions and achieve redox homeostasis under photo-oxidative stress.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Exposure to oxidative stress typically raises the reactive oxygen species (ROS) levels in plant tissues. Disturbance in redox can affect the functioning of chloroplasts or mitochondria or both (Gill and Tuteja 2010). The ROS levels and antioxidants provide an effective network of adaptation (Foyer and Noctor 2003). One of the benefits from the cross-talk between chloroplasts and mitochondrial interactions is redox stabilization so as to maintain optimal levels of ROS (Raghavendra and Padmasree 2003; Sunil et al. 2013). However, it is not clear if the different organelles respond similarly under variable stress conditions.
Among the well-known antioxidant compounds, ascorbate (ASC) can mediate the cross-talk between mitochondria and chloroplasts (Talla et al. 2011; Szarka et al. 2013). Glutathione (GSH) is mostly synthesized in chloroplasts, but is required for redox balancing in cytoplasm and mitochondria (Noctor et al. 2012; Szalai et al. 2009). Besides these two classical antioxidants ASC and GSH, other compatible solutes such as proline and GABA may also have a role in redox responses of plant cells (Liang et al. 2013; Li et al. 2016).
Apart from antioxidants, accumulation of osmolytes is one of the common responses to abiotic stresses in many plants. Among the osmolytes, proline accumulates in large quantities, when plants are exposed to stress. A strong correlation has been established between proline accumulation, abiotic stress tolerance, and adaptation to metabolic disturbance (Hebbelmann et al. 2012; Sperdouli and Moustakas 2012; Kishor and Sreenivasulu 2014). Proline seems to have multifunctional roles, besides being an osmolyte. For e.g., proline can play a role in scavenging ROS, stabilizing proteins, supplying energy in the form of reducing equivalents, modulating cellular redox homeostasis and interacting with other metabolic pathways under stress (Kishor et al. 2005; Verbruggen and Hermans 2008; Szabados and Savoure 2010; Sharma et al. 2011; Wang et al. 2015).
There is extensive literature, on the biochemical basis of proline accumulation under varying abiotic stresses like drought/dehydration (Kiyosue et al. 1996; Parida et al. 2008) or salt (Huang et al. 2013; Wang et al. 2015). The levels of proline depends on the balance between the rates of biosynthesis and degradation (Szabados and Savoure 2010; Hayat et al. 2012). Enzymes mediating proline biosynthesis are localized in chloroplasts and cytosol, while those of proline oxidation are located in mitochondria (Szabados and Savoure 2010). Under abiotic stress, plant metabolism will be disturbed and the metabolic network must be reconfigured for acclimation to prevailing stress conditions.
We have attempted to examine the consequences of triggering oxidative and photo-oxidative stress in chloroplasts and mitochondria, with a focus on proline. Oxidative and photo-oxidative stress was induced by either methyl viologen (MV) or menadione (MD), which acts primarily on chloroplasts or mitochondria, respectively. MV, a redox-active molecule, induces ROS production and causes light dependent oxidative damage in leaves (Moustaka et al. 2015). MV acts primarily on the chloroplasts in the light through the PS-I mediated generation of superoxide (Donahue et al. 1997; Li et al. 2013; Hawkes et al. Hawkes 2014). In contrast MD induces oxidative stress mainly in mitochondria, by competing with the ubiquinone pool for electrons from complexes I and II (Sweetlove et al. 2002; Obata et al. 2011). A redox-sensitive roGFP was used by Lehmann et al. (2009) to demonstrate that exposure to MD led to a rapid disturbance in the redox state of mitochondria and later in other cellular compartments. When exposed to paraquat/MV, a rise in the ascorbate and glutathione content (Hemavathi et al. 2011), increased antioxidant enzymatic activity of APX and SOD (Moustaka et al. 2015), and accumulation of proline were observed (Rejeb et al. 2015).
The present article describes our efforts to evaluate if proline can be a factor to mediate the interaction between chloroplasts and mitochondria, when exposed to oxidative stress. The differential regulation of proline biosynthesis/oxidation under photo-oxidative stress conditions created specifically in chloroplasts or mitochondria has not been much studied. We emphasize that the disturbance in redox of either chloroplasts or mitochondria would promote accumulation of proline, by promoting biosynthesis in chloroplasts and limiting oxidation in mitochondria. We emphasize that proline could be another important metabolite during interactions of mitochondria and chloroplasts, particularly under oxidative stress.
Materials and methods
Plant growth
Seeds of pea (Pisum sativum L., cv. Arkel) were procured from Vipul seeds, Indore. The seeds were soaked overnight in water and sterilized with sodium hypochlorite. They were covered with filter paper and allowed to germinate. The germinated seeds were sown in plastic trays filled with farmyard manure and soil. The plants were raised in temperature controlled green houses, with average temperature of 30 °C day/20 °C night.
Preparation and treatment of leaf discs
Discs (ca. 0.25 cm2) were prepared from leaves with a sharp paper punch, from either side of the midrib, under water. These discs were incubated in 5-cm-diameter Petri dishes containing the medium (2 mM KH2PO4, 1 mM CaCl2, 1 mM KCl, pH 6.5) and respective test compound (10 μM MV or 10 μM MD) for 4 h in either dark or moderate (300 μE m−2 s−1, ML) or high light (1200 μE m−2 s−1, HL). The oxidants MV and MD are known to trigger stress in chloroplasts and mitochondria, respectively (Li et al. 2013; Obata et al. 2011).
Levels of H2O2, superoxide (O2 •−) and lipid peroxidation
The detection of H2O2 and superoxide was by 3, 3′-diaminobenzidine (DAB) and nitro blue tetrazolium chloride (NBT), respectively. Leaf discs were vacuum infiltrated (at 25 psi) with 1 mg ml−1 DAB (pH 3.8) or 1 mg ml−1NBT for 5 min and then with the oxidants for 3 min. Later leaf discs were placed in respective treatment conditions: dark, ML (300 μE m−2 s−1) or HL (1200 μE m−2 s−1). H2O2 was visualized as a reddish brown color, whereas superoxide radicals were detected as blue color formazan. Quantification of both H2O2 and superoxide was done according to Kwon et al. (2013).
The H2O2 levels in leaves with or without treatments were measured by also using Amplex red reagent (Molecular Probes) according to Lee et al. (2012). One hundred milligrams of leaf sample was powdered in liquid nitrogen and extracted in sodium phosphate buffer (pH 6.5). Mixture was centrifuged at 9500×g for 10 min at 4 °C and the supernatant was examined for absorbance at 560 nm. H2O2 content was determined by using a standard curve. Lipid peroxidation assay was performed according to Hodges et al. (1999).
Proline, ascorbate, and glutathione levels
Proline was estimated according to Bates et al. (1973). Leaf samples were ground in liquid nitrogen with 3% (w/v) sulfosalicylic acid and the extract was centrifuged at 15200×g for 10 min. An aliquot of the cleared extract was mixed with sulfosalicylic acid, glacial acetic acid, and acid-ninhydrin and incubated for 1 h at 96 °C. The reaction was stopped by transferring the tubes to ice. To this mixture, 2 ml of toluene was added, stirred for 20 s and left for 5 min to allow phase separation. The absorbance of toluene (upper layer) was measured at 520 nm with toluene as reference. Proline content was determined using a standard curve.
Approximately 100 mg of pea leaf discs were harvested and immediately frozen in liquid nitrogen. Total ASC content was measured according to Gillespie and Ainsworth (2007). Ascorbic acid was calculated using ASC as the standard. The levels of GSH were determined spectrophotometrically as described by Griffith (1980) by measuring the absorbance at 412 nm, using a standard curve generated with GSH.
P5CS and PDH: activity and protein levels
The leaf discs (approx. 100-mg fresh weight) after treatment were collected and ground to powder in liquid nitrogen. The assay of P5CS was done according to Garcia-Rios et al. (1997). Activity was measured as the consumption of NADPH, monitored as a decrease in absorbance at 340 nm. PDH activity was measured according to Sanchez et al. (Sánchez et al. 2001). Reduction of NAD+ at 340 nm was measured to determine enzyme activity.
Western blotting
The leaf discs (100 mg) after respective treatment were ground to powder in liquid nitrogen. The powdered samples of leaf discs were homogenized in 125 mM Tris–HCl (pH 6.8) containing 5% (w/v) sodium dodecyl sulfate (SDS) and 1 mM PMSF followed by a brief centrifugation step at 1000×g for 1 min and supernatant was collected. Protein was estimated by the Bradford method (Bradford 1976).
SDS-PAGE was performed as per the principles of Laemmli (1970) using mini gels. The proteins were then transferred electrophoretically from the gel onto polyvinylidene difluoride (PVDF) membranes (Towbin et al. 1979), using power supply of 30 V, for 4 h at 4 °C. The PVDF membrane was blocked and was treated for 1 h with primary antibody (1: 2000) followed by 1: 2000 dilution of secondary antibody (goat anti-rabbit IgG, Agrisera AB, Sweden) conjugated with alkaline phosphatase. We used antibodies raised against Arabidopsis (Cecchini et al. 2011). We could not check the specificity of the PDH antibody against pea PDH enzyme, as we do not have the pure protein. Therefore, we checked the similarity of PDH proteins of Arabidopsis thaliana and pea, by the analysis of relevant databases.
The gene sequences of PDH1 of Arabidopsis thaliana and PDH of pea were retrieved from the NCBI (gene) database. When these two gene sequences were translated to amino acid sequences using ExPASy Translate tool, the amino acid sequences of Arabidopsis PDH 1 and pea PDH showed up a similarity of > 82%, when checked by LALIGN tool. The small deviation from the ideal 100% similarity might be the reason that higher quantities of antibody were required to detect PDH in pea leaf extract. We had to increase the level of antibody (1:2000 dilution) to detect PDH in pea, compared to the dilution used for Arabidopsis (1:5000). The washed blot was developed with 1 ml of pre-mixed BCIP/NBT solution (Sigma-Aldrich, St. Louis, USA).
RNA extraction and cDNA synthesis
The leaf discs (approx. 100-mg fresh weight) were ground to powder in liquid nitrogen. The extraction of RNA was done, using guanidine thiocyanate acid phenol-based method (Chomczynski and Sacchi 1987) using TRI-Reagent® (Sigma-Aldrich, USA). cDNA was synthesized following manufacturer’s instructions (RevertAid cDNA synthesis kit, Fermentas) and was used as template for RT-PCR analysis.
RT-PCR analysis
The complete genome of Pisum sativum is not yet available (Kulaeva et al. 2017). Semi-quantitative reverse transcription-PCR (RT-PCR) was performed using primers designed on the basis of the published sequences (http://www.ncbi.nlm.nih.gov) and using ESTs available in cool season food legume database (https://www.coolseasonfoodlegume.org/) using Primer3 software (Rozen and Skaletsky 2000). The sequences of the primers used for various enzymes and the house keeping gene are given in Supplemental Table 1. The PCR conditions were optimized by testing annealing temperatures. Actin 2 was used as an internal (constitutive) control, and the amplified products were resolved by electrophoresis. Band intensities after electrophoresis were quantified using Image J software and normalized with reference to band intensities of Actin 2.
Replications
The data presented are the average of results (mean ± SE) from three to four experiments conducted on different days. Significance of difference was tested by one-way ANOVA and student’s t test using SIGMA PLOT 11.
Results
Production of ROS: peroxides and superoxide
The oxidants directed at either chloroplast (MV) or mitochondria (MD) triggered an increase in ROS generation, as indicated by staining with DAB and NBT, for H2O2 and superoxide, respectively. The extent of ROS accumulation was higher at 4 h of incubation than that at 2 h. Representative images of DAB and NBT stained leaf discs with oxidants at 2 h and 4 h were given in Supplemental Fig. 1 and the quantified values were given in Supplemental Table 2. In an earlier report, Wang et al. (2015) reported that proline content increased by 37% in leaves of Kosteletzkya viriginica after 2 h of salt stress, whereas proline levels further increased up to twofold at the end of 6 h. In another report, threefold increases in proline was observed in leaves of grapevine after treatment with H2O2 for 6 h (Ozden et al. 2009). We could see an increase of approximately threefold in proline, even by 4 h of exposure to oxidants particularly under HL. In view of the significant increase in proline, we limited all our subsequent experiments to 4 h. We also felt that exposure to periods exceeding 4 h, particularly under HL, could be harmful.
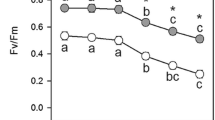
The levels of H2O2 measured by DAB staining (Fig. 1a), were much higher than the levels determined by Amplex red reagent (Fig. 1b). Highest H2O2 levels were in light compared to those in dark, in both MV and MD treated samples (Fig. 1a, b). The levels of H2O2 were threefold higher in MV treated samples under HL compared to control, while the levels of superoxide were about twofold higher in HL (Fig. 1c). The extent of lipid peroxidation, as indicated by MDA levels, was not much altered in dark conditions. In contrast, the level of lipid peroxidation was twofold higher in HL treated samples on treatment with MV (Fig. 1d).

The levels of ROS and lipid peroxidation were monitored in leaf discs after treatment with oxidants for 4 h in either dark, 300 or 1200 μE m−2 s−1 light intensities. The amount of DAB in stained discs was quantified to assess the levels of H2O2 (a). The levels of H2O2 were measured also by using Amplex red (b). Superoxide levels were quantified as formazan formed after incubating with NBT (c). Lipid peroxidation in leaf discs was determined as MDA equivalents (d). The images for DAB and NBT stained leaf discs in a typical experiment are in Supplemental Fig. 1. Each data point is the average of three replicates, and error bars represent ± SE. Asterisks indicate significance of the effects of MV or MD, compared to respective control. *P < 0.05; **P < 0.01; and ***P < 0.001
Levels of proline, ascorbate, and glutathione
There was no significant change in levels of proline when treated with MV or MD in dark. In contrast, there was marked increase in the level of proline (> threefold) in either MD or MV treated samples, under HL (Fig. 2a). Ascorbate and glutathione contents increased on treatment with MV or MD, but, the increase (about 50 to 60%) was much less than that of proline (Fig. 2b, c). Enhanced antioxidant enzymatic activities and upregulation of transcripts were observed upon exposure to oxidants for 4 h (Supplemental Figs. 2 and 3).

Proline (a) or ascorbate (b) or glutathione (c) content in leaf discs after exposure to oxidants for 4 h in dark, 300 or 1200 μE m−2 s−1 light intensities. Proline or antioxidant levels were expressed as μmol g−1 FW. Each data point is the average of three replicates, and error bars represent ± SE. Asterisks indicate significance of MV or MD-effect, compared to respective control. *P < 0.05; and ***P < 0.001
Enzymes of proline biosynthesis and oxidation: activities, protein levels, and transcripts
Since proline accumulation was more pronounced upon oxidant (MV or MD) treatment, we measured the activities of enzymes involved in proline metabolism. The activity of P5CS increased markedly while the activity of PDH decreased. These changes were pronounced in HL compared to that in ML or dark. For e.g., the activity of P5CS increased more than threefold when treated with MV under HL (Fig. 3a). In contrast, the activity of PDH decreased by as much as 50% on treatment with MV under HL (Fig. 3b).

Activities of P5CS (a) and PDH (b) (enzymes involved in proline metabolism) were measured in extracts after treatment for 4 h with oxidants in dark, 300 or 1200 μE m−2 s−1 light intensities. Activity was expressed as μmol mg−1 protein min−1. Each data point is the average of three replicates, and error bars represent ± SE. Asterisks indicate significant difference determined by t test, compared to respective control. *P < 0.05; and ***P < 0.001
When the levels of PDH protein were determined by using an antibody, there was no marked change in PDH protein content when treated with MV or MD in dark. However, the amount of PDH protein decreased on treatment with oxidants, MV or MD, particularly under HL (Fig. 4a). The decrease in PDH on exposure to MV or MD was significant, particularly under HL (Fig. 4b). Protein levels of P5CS could not be determined due to non-availability of antibody.

Western blot analysis of proline dehydrogenase (PDH) enzyme showing protein levels in leaves treated with MV or MD for 4 h in dark, 300 or 1200 μE m−2 s−1 light intensities. Ponceau-stained gels showing Rubisco large subunits are in lower panel to indicate the protein loading in each gel (a). The ratios of PDH to Rubisco were calculated based on the ImageJ analysis and plotted (B). Each data point is an average of three replicates, and error bars represent ± SE. Asterisks indicate significant difference determined by t test, compared to respective control. *P < 0.05; and **P < 0.01
The transcripts of genes involved in proline metabolism were studied and quantified (Fig. 5a). Both P5CS and P5CR genes were upregulated in light-treated conditions and no predominant changes were observed in dark (Fig. 5b, c). Under light conditions, 1.8-fold increase of P5CS transcripts was observed in MV or MD treated samples and the stimulation was significant (Fig. 5b). In contrast, the upregulation of P5CR was significant only by MV but not with MD (Fig. 5c). Though downregulation of P5CDH and PDH genes was noticed, a significant change of P5CDH was not observed under given conditions (Fig. 5d). Significant decrease in transcripts of PDH was seen in both ML and HL conditions (Fig. 5e). Maximum downregulation was seen in case of ML-treated MD samples, with a decrease from 1 to 0.24, whereas the levels were reduced to 0.54 only in case of MV-treated ML samples (Fig. 5d, e).

Expression pattern of genes involved in proline metabolism. a The transcripts from semi-quantitative RT-PCR of leaf extracts, after treatment with oxidants for 4 h in dark, 300 or 1200 μE m−2 s−1 light intensities. Actin 2 was used as an internal control. b Extent of changes in expression of genes after normalizing with actin. Each data point is the average of three replicates, and error bars represent ± SE. Asterisks indicate significance of effects of MV or MD, compared to respective control. *P < 0.05; **P < 0.01; and ***P < 0.001
Discussion
There was a marked rise in the levels of ROS (H2O2 or superoxide or MDA equivalents) when treated with an oxidant, that acts primarily on either chloroplasts (e.g., MV) or mitochondria (MD). The following three points emerged from the results.
-
i.
Photo-oxidative stress (particularly HL) elicited stronger response than the oxidative stress in dark. Thus, the disturbance in chloroplasts was more stressful than that in mitochondria.
-
ii.
Proline accumulation was more pronounced than that of ascorbate and glutathione.
-
iii.
P5CS and PDH, key enzymes of proline biosynthesis and oxidation respectively, were strongly modulated and hence more important than the other two enzymes P5CR and P5CDH during oxidative/photo-oxidative stress.
ROS accumulation was maximal under photo-oxidative stress
An increase in H2O2 content on exposure to MV was observed in leaf discs of tobacco (Kwon et al. 2013). Similarly in another study, treatment with MD increased H2O2 in Arabidopsis suspension cultures (Sweetlove et al. 2002). Our study with both MV and MD in the same experimental system is the first such attempt. A marked increase in H2O2 and superoxide occurred when leaf discs were treated with either MV or MD, particularly under HL (Fig. 1). Though the ROS accumulation occurred irrespective of compartment targeted, high accumulation was noticeable when chloroplasts were stressed (for e.g., by MV). Since, accumulation of both H2O2 and superoxide was always much higher in light than that in dark, we felt that photo-oxidative conditions could create more stress than oxidative conditions.
Marked increase in proline
The levels of ASC and GSH (important antioxidants in plant cells) usually increase on exposure to oxidative stress (Foyer 2001). In the present study too, the total ASC and GSH content increased on oxidant treatment under both dark and light conditions (Fig. 2b, c). It is obvious that oxidative stress triggered in chloroplasts (by MV) or mitochondria (by MD) lead to an increase in levels of antioxidants. Increase of both ASC and GSH upon exposure to MV has been observed in potato plants (Hemavathi et al. 2011).
Accumulation of compatible solutes such as proline is one of the common responses to abiotic stresses in plants. We observed that the increase in proline levels (Fig. 2a) was more pronounced than that of ASC/GSH (Fig. 2b, c). This suggests that proline can be as crucial as the other well-known antioxidants, when there is a redox disturbance in either chloroplasts or mitochondria. Though proline was not much accumulated in dark, increased proline levels were quite evident in light particularly in MV-treated samples where chloroplasts are the major targets. We suggest that light may be playing a crucial role in accumulating proline under photo-oxidative stress.
Basis of proline accumulation: increased synthesis and decreased oxidation
An important point from our results is that the modulation of biosynthesis/oxidation leading to an elevation of proline levels is mediated by oxidative stress in either chloroplasts or mitochondria. The aggravation of stress in HL was reflected again in higher proline levels (Fig. 2a). Proline biosynthesis occurs from glutamate in chloroplasts, while proline oxidation occurs in mitochondria. The levels of proline appear to be tightly regulated by biosynthesis and oxidation under oxidative/photo-oxidative stress. An alternative pathway of proline biosynthesis from ornithine was found during seedling development (Armengaud et al. 2004).
Based on the enzymatic activities and gene expression patterns, we suggest that the modulation of P5CS and PDH could be the major reason for proline accumulation under oxidative or photo-oxidative stress by MV or MD (Figs. 3 and 5). Changes in expression of P5CDH and P5CR were much less than those of P5CS and PDH. Similar pattern of proline accumulation was observed under various abiotic stresses (Parida et al. 2008; Huang et al. 2013; Kishor and Sreenivasulu 2014; Wang et al. 2015).
Relevance to chloroplast-mitochondrial interactions
Since two key enzymes of biosynthesis and oxidation (namely, P5CS in chloroplasts and PDH in mitochondria) were modulated by oxidants targeted either to chloroplasts (MV) or mitochondria (MD), we feel that proline could be another key metabolite during chloroplast-mitochondrial interactions. The ability of proline to directly scavenge the ROS is debated (Signorelli 2016), but the importance of proline in maintenance of redox balance in chloroplasts and mitochondria has been suggested earlier (Szabados and Savoure 2010; Shinde et al. 2016). In our experiments, the responses to stress in chloroplasts were always higher than in mitochondria, indicating that stress or redox disturbance within chloroplasts was stronger. Proline synthesis could be an important sink for reducing equivalents in chloroplasts and can help to minimize the excess reduction.
Concluding remarks
Disturbance in redox state of either chloroplasts (by MV) or mitochondria (by MD) increased the ROS levels in leaf tissues, followed by a marked increase in levels of proline, maximum being under HL. Upregulation of P5CS and downregulation of PDH were the reasons for enhanced proline levels. Since these two enzymes of P5CS and PDH are located in chloroplasts and mitochondria,, proline metabolism could mediate the utilization of reductants in chloroplasts and regeneration in mitochondria, thus maintaining an optimal redox status. Additionally, proline could team up with other metabolic pathways such as lipids to ensure an efficient redox (Shinde et al. 2016). We propose that proline, like ASC, could be an important metabolite to facilitate the cross-talk between mitochondria and chloroplasts, during adaptive responses to photo-oxidative response. Proline transporters are known to operate in mitochondria (Szabados and Savoure 2010; Shinde et al. 2016), but the mechanism of proline transport across chloroplasts is not clear.
Abbreviations
- ASC:
-
Ascorbate
- DAB:
-
3,3′-Diaminobenzidine
- GSH:
-
Glutathione
- HL:
-
High light
- MD:
-
Menadione
- MDA:
-
Malondialdehyde
- ML:
-
Moderate light
- MV:
-
Methyl viologen
- NBT:
-
Nitro blue tetrazolium chloride
- P5CDH:
-
Pyrroline-5-carboxylate dehydrogenase
- P5CR:
-
Pyrroline-5-carboxylate reductase
- P5CS:
-
Pyrroline-5-carboxylate synthetase
- PDH:
-
Proline dehydrogenase
- PVDF:
-
Polyvinylidene difluoride
- ROS:
-
Reactive oxygen species
References
Armengaud P, Thiery L, Buhot N, Grenier-DeMarch G, Savouré A (2004) Transcriptional regulation of proline biosynthesis in Medicago truncatula reveals developmental and environmental specific features. Physiol Plant 120:442–450
Bates LS, Waldren RP, Teare ID (1973) Rapid determination of free proline for water-stress studies. Plant Soil 39:205–207
Bradford MM (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72:248–254. https://doi.org/10.1016/0003-2697(76)90527-3
Cecchini NM, Monteoliva MI, Alvarez ME (2011) Proline dehydrogenase contributes to pathogen defense in Arabidopsis. Plant Physiol 155:1947–1959
Chomczynski P, Sacchi N (1987) Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal Biochem 162:156–159
Donahue JL, Okpodu CM, Cramer CL, Grabau EA, Alscher RG (1997) Responses of antioxidants to paraquat in pea leaves. Plant Physiol 113:249–257
Foyer CH (2001) Prospects for enhancement of the soluble antioxidants, ascorbate and glutathione. BioFactors 15:75–78
Foyer CH, Noctor G (2003) Redox sensing and signalling associated with reactive oxygen in chloroplasts, peroxisomes and mitochondria. Physiol Plant 119:355–364
Garcia-Rios M, Fujita T, LaRosa PC, Locy RD, Clithero JM, Bressan RA, Csonka LN (1997) Cloning of a polycistronic cDNA from tomato encoding γ -glutamyl kinase and γ- glutamyl phosphate reductase. Proc Natl Acad Sci U S A 94:8249–8254
Gill SS, Tuteja N (2010) Reactive oxygen species and anti-oxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol 148:909–930
Gillespie KM, Ainsworth EA (2007) Measurement of reduced, oxidizied and total ascorbate content in plants. Nat Protoc 2:871–874
Griffith OW (1980) Determination of glutathione and glutathione disulfide using glutathione reductase and 2-vinyl-pyridine. Anal Biochem 106:207–212
Hawkes TR (2014) Mechanisms of resistance to paraquat in plants. Pest Manag Sci 70:1316–1323
Hayat S, Hayat Q, Alyemeni MN, Wani AS, Pichtel J, Ahmad A (2012) Role of proline under changing environments. Plant Signal Behav 7:1456–1466
Hebbelmann I, Selinski J, Wehmeyer C, Goss T, Voss I, Mulo P, Kangasjarvi S, Aro EM, Oelze ML, Dietz KJ, Nunes-Nesi A, Do PT, Fernie AR, Talla SK, Raghavendra AS, Linke V, Scheibe R (2012) Multiple strategies to prevent oxidative stress in Arabidopsis plants lacking the malate valve enzyme NADP-malate dehydrogenase. J Exp Bot 63:1445–1459
Hemavathi UCP, Akula N, Kim HS, Jeon JH, Ho OM, Chun SC, Kim DH, Park SW (2011) Biochemical analysis of enhanced tolerance in transgenic potato plants overexpressing D-galacturonic acid reductase gene in response to various abiotic stresses. Mol Breed 28:105–115
Hodges DM, DeLong JM, Forney CF, Prange RK (1999) Improving the thiobarbituric acid-reactive-substances assay for estimating lipid peroxidation in plant tissues containing anthocyanin and other interfering compounds. Planta 207:604–611
Huang Z, Zhao L, Chen D, Liang M, Liu Z (2013) Salt stress encourages proline accumulation by regulating proline biosynthesis and degradation in Jerusalem artichoke plantlets. PLoS One 8:e62085
Kishor PBK, Sreenivasulu N (2014) Is proline accumulation per se correlated with stress tolerance or is proline homeostasis a more critical issue? Plant Cell Environ 37:300–311
Kishor PBK, Sangam S, Amrutha RN, SriLaxmi P, Naidu KR, Rao KRSS (2005) Regulation of proline biosynthesis, degradation, uptake and transport in higher plants: its implications in plant growth and abiotic stress tolerance. Curr Sci 88:424–438
Kiyosue T, Yoshiba Y, Yamaguchi-Shinozaki K, Shinozaki K (1996) A nuclear gene encoding mitochondrial proline dehydrogenase, an enzyme involved in proline metabolism, is upregulated by proline but downregulated by dehydration in Arabidopsis. Plant Cell 8:1323–1335
Kulaeva OA, Zhernakov AI, Afonin AM, Boikov SS, Sulima AS, Tikhonovich IA, Zhukov VA (2017) Pea marker database (PMD) - a new online database combining known pea (Pisum sativum L.) gene-based markers. PLoS One 12:e018713
Kwon KC, Verma D, Jin S, Singh ND, Daniell H (2013) Release of proteins from intact chloroplasts induced by reactive oxygen species during biotic and abiotic stress. PLoS One 8:e67106
Laemmli UK (1970) Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227:680–685
Lee S, Seo PJ, Lee HJ, Park C (2012) A NAC transcription factor NTL4 promotes reactive oxygen species production during drought-induced leaf senescence in Arabidopsis. Plant J 70:831–844
Lehmann M, Schwarzlander M, Obata T, Sirikantaramas S, Burow M, Olsen CE, Tohge T, Fricker MD, Møller BL, Fernie AR, Sweetlove LJ, Laxa M (2009) The metabolic response of arabidopsis roots to oxidative stress is distinct from that of heterotrophic cells in culture and highlights a complex relationship between the levels of transcripts, metabolites, and flux. Mol Plant 2:390–406
Li J, Mu J, Bai J, Fu F, Zou T, An F, Zhang J, Jing H, Wang Q, Li Z, Yang S, Zuo J (2013) Paraquat resistant 1, a golgi-localized putative transporter protein, is involved in intracellular transport of paraquat. Plant Physiol 162:470–483
Li Z, Yu J, Peng Y, Huang B (2016) Metabolic pathways regulated by γ-aminobutyric acid (GABA) contributing to heat tolerance in creeping bentgrass (Agrostis stolonifera). Sci Rep 6:30338
Liang X, Zhang L, Natarajan SK, Becker DF (2013) Proline mechanisms of stress survival. Antioxid Redox Signal 19:998–1011
Moustaka J, Tanou G, Adamakis ID, Elefteriou EP, Moustakas M (2015) Leaf age-dependent photoprotective and antioxidative response mechanisms to paraquat-induced oxidative stress in Arabidopsis thaliana. Int J Mol Sci 16:13989–14006
Noctor G, Mhamdi A, Chaouch S, Han Y, Neukermans J, Marquez-Garcia B, Queval G, Foyer CH (2012) Glutathione in plants: an integrated overview. Plant Cell Environ 35:454–484
Obata T, Matthes A, Koszior S, Lehmann M, Araujo WL, Bock R, Sweetlove LJ, Fernie AR (2011) Alteration of mitochondrial protein complexes in relation to metabolic regulation under short-term oxidative stress in Arabidopsis seedlings. Phytochemistry 72:1081–1091
Ozden M, Demirel U, Kahraman A (2009) Effects of proline on antioxidant system in leaves of grapevine (Vitis vinifera L.) exposed to oxidative stress by H2O2. Sci Hort 119:163–168
Parida AK, Dagaonkar VS, Phalak MS, Aurangabadkar LP (2008) Differential responses of the enzymes involved in proline biosynthesis and degradation in drought tolerant and sensitive cotton genotypes during drought stress and recovery. Acta Physiol Plant 30:619–627
Raghavendra AS, Padmasree K (2003) Beneficial interactions of mitochondrial metabolism with photosynthetic carbon assimilation. Trends Plant Sci 8:546–553
Rejeb KB, Vos LD, Le Disquet I, Leprince AS, Bordenave M, Maldiney R, Asma J, Abdelly C, Savour A (2015) Hydrogen peroxide produced by NADPH oxidases increases proline accumulation during salt or mannitol stress in Arabidopsis thaliana. New Phytol 208:1138–1148
Rozen S, Skaletsky H (2000) Primer3 on the WWW for general users and for biologist programmers. Methods Mol Biol 132:365–386
Sánchez E, López-Lefebre LR, García PC, Rivero RM, Ruiz JM, Romero L (2001) Proline metabolism in response to highest nitrogen dosages in green bean plants (Phaseolus vulgaris L. cv. Strike). J Plant Physiol 158:593–598
Sharma S, Villamor JG, Verslues PE (2011) Essential role of tissue- specific proline synthesis and catabolism in growth and redox balance at low water potential. Plant Physiol 157:292–304
Shinde S, Villamor JG, Lin W, Sharma S, Verslues PE (2016) Proline coordination with fatty acid synthesis and redox metabolism of chloroplast and mitochondria. Plant Physiol 172:1074–1088
Signorelli S (2016) The fermentation analogy: a point of view for understanding the intriguing role of proline accumulation in stressed plants. Front Plant Sci 7:1339
Sperdouli I, Moustakas M (2012) Interaction of proline, sugars, and anthocyanins during photosynthetic acclimation of Arabidopsis thaliana to drought stress. J Plant Physiol 169:577–585
Sunil B, Talla SK, Aswani V, Raghavendra AS (2013) Optimization of photosynthesis by multiple metabolic pathways involving interorganelle interactions: resource sharing and ROS maintainance as the bases. Photosynth Res 117:61–71
Sweetlove LJ, Heazlewood JL, Herald V, Holtzapffel R, Day DA, Leaver CJ, Millar AH (2002) The impact of oxidative stress on Arabidopsis mitochondria. Plant J 32:891–904
Szabados L, Savoure A (2010) Proline: a multifunctional amino acid. Trends Plant Sci 15:89–97
Szalai G, Kellos T, Galiba G, Kocsy G (2009) Glutathione as an antioxidant and regulatory molecule in plants under abiotic stress conditions. J Plant Growth Regul 28:66–80
Szarka A, Banhegyi G, Asard H (2013) The inter-relationship of ascorbate transport, metabolism and mitochondrial, plastidic respiration. Antioxid Redox Signal 19:1036–1044
Talla SK, Riazunnisa K, Padmavathi L, Sunil B, Rajsheel P, Raghavendra AS (2011) Ascorbic acid is a key participant during the interactions between chloroplasts and mitochondria to optimize photosyhthesis and protect against photoinhibition. J Biosci 36:163–173
Towbin H, Staehlin T, Gordon J (1979) Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets. Procedure and some applications. Proc Natl Acad Sci U S A 76:4350–4354
Verbruggen N, Hermans C (2008) Proline accumulation in plants: a review. Amino Acids 35:753–759
Wang H, Tang X, Wang H, Shao HB (2015) Proline accumulation and metabolism-related genes expression profiles in Kosteletzkya virginica seedlings under salt stress. Front Plant Sci 6:792
Acknowledgments
Authors thank Prof. Maria Elena Alvarez, Faculty of Chemistry, National University of Cordoba, Argentina for kindly providing us with proline dehydrogenase antibodies for our experiments.
Funding
This work was supported by grants to ASR from Council of Scientific and Industrial Research (No. 38(1404)/15/EMR-II), JC Bose National Fellowship (No. SR/S2/JCB-06/2006). VA, PR, and RBB were all supported by Research Fellowships from University Grants Commission, New Delhi, India. We also thank grants from DST-FIST, UGC-SAP-CAS, and DBT-CREBB, from New Delhi, India for support of infrastructure in Department/School.
Author information
Authors and Affiliations
Contributions
ASR designed the study. VA performed most of the experiments. PR, RBB, and BS performed some experiments. VA and ASR analyzed results and wrote the manuscript. VA, PR, RBB, BS, and ASR revised and finalized the manuscript. All the authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Rights and permissions
About this article

Cite this article
Aswani, V., Rajsheel, P., Bapatla, R.B. et al. Oxidative stress induced in chloroplasts or mitochondria promotes proline accumulation in leaves of pea (Pisum sativum): another example of chloroplast-mitochondria interactions. Protoplasma 256, 449–457 (2019). https://doi.org/10.1007/s00709-018-1306-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00709-018-1306-1