
Abstract
Key message
The daily stem radial growth of P. orientalis during the main growing period in semi-arid areas of North China is mainly limited by moisture availability.
Abstract
Finer resolution studies of tree stem radial growth on short temporal scales can be useful for exploring the main environmental variables that trigger radial growth. The objective of this study was to determine the most important environmental factors that impact the intra-annual radial growth of Platycladus orientalis (Linn.) Franco in a semi-arid area of North China. The stem radial growth of P. orientalis was studied daily during the growing season (May to October 2013) using high-resolution automatic point dendrometers. Our results showed that the daily stem radial increment (SRI) of P. orientalis during the main growing period was primarily impacted by relative air humidity, precipitation, vapor pressure deficit and daily maximum air temperature. Those environmental factors mainly influenced the moisture availability to the trees, thereby affecting their daily stem radial growth. The rapid release of stem radial growth was observed when heavy precipitation events occurred after a drought period. The maximum stem growth rate of P. orientalis was observed in early August and was regulated by soil water content in the summer; this was regarded as an adaptation to the local climate conditions. Our results have further implications for sensitivity analyses of tree growth to climate changes in semi-arid areas.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Tree growth in semi-arid areas is sensitive to climate change, which has potentially great impacts on vegetation changes, regional climate feedback and the carbon cycle (Rotenberg and Yakir 2010; Williams et al. 2010; Liu et al. 2013). Previous dendroclimatological studies have reported that tree growth in semi-arid areas is generally limited by moisture availability (Liang et al. 2001, 2008; Cherubini et al. 2003; Gebrekirstos et al. 2008; Shi et al. 2008; Dulamsuren et al. 2009). Lebourgeois et al. (2012) reported that spring drought (May–June) is the primary factor limiting conifer growth in the Mediterranean, with narrower tree rings observed in years with a drier spring. A broader study by Liu et al. (2013) showed that tree-ring width at semi-arid forest sites throughout Inner Asia is positively correlated with precipitation from March to June of the growth year, and negatively correlated with vapor pressure deficit (VPD) from April to July of the growth year.
Dendroclimatology aims to establish long-term relationships between radial growth and climatic variables by examining correlations with monthly, seasonal and annual resolution (Fritts and Shatz 1975). The relative low resolution of dendroclimatology limits its interpretation power to the intra-annual short-term radial growth, such as daily stem radial increment and radial growth rate, response to environmental factors (Gutiérrez et al. 2011; Köcher et al. 2012). Finer resolution studies, which address radial growth responses on a daily time scale during the growing season, can be useful to explore the main environmental variables that influence daily stem radial growth (Pérez et al. 2009).
Dendrometer measurements provide a continuous time series of intra-annual stem radial variation with high temporal resolution (minutes to hours) without invasive sampling (Drew and Downes 2009 ); this technique has been widely used for different tree species and environments to describe the relationship between intra-annual radial growth and climate in the last decade (Downes et al. 1999; Mäkinen et al. 2008; Bräuning et al. 2009; Volland-Voigt et al. 2011; Köcher et al. 2012). For example, Gutiérrez et al. (2011) reported that the seasonal radial increment of Quercus ilex L., a dominant evergreen oak species in the Western Mediterranean Basin, is bimodal and characterized by a greater increase in May and a lesser, more variable increase peak in September. Xiao et al. (2014) showed that groundwater depth and the related soil moisture content are key factors potentially limiting the radial growth of Populus euphratica, a dominant tree species in desert riparian forests in arid regions of northwest China (Westermann et al. 2008).
Platycladus orientalis is an evergreen tree species with a well-developed root system and a long lifespan that originated in China that is also naturalized in Korea, Japan and Iran (Lei et al. 2010). Due to its resistance to cold, drought and nutrient deficiency, P. orientalis has become one of the dominant evergreen tree species in semi-arid areas of North China. Recently, it has been frequently used for afforestation in the semi-arid Loess Plateau of China (Guo et al. 2007). In recent years, a few dendroclimatological studies have examined the stem radial growth of P. orientalis. Huang et al. (2006) found that the ring width of P. orientalis has a significant positive response to precipitation and a significant negative response to temperature, both from June to August, the current growing season. However, basic studies on the intra-annual daily stem radial growth of P. orientalis are still lacking.
In this study, we examined the intra-annual weather response of the stem radial growth of P. orientalis in a semi-arid area of North China using high-resolution automatic point dendrometers. We hypothesized that moisture availability throughout the growing season determines its daily stem radial growth. To verify this hypothesis, we aim to (a) test the environmental factors air temperature, relative air humidity, vapor pressure deficit, precipitation, soil temperature and soil water content in their effect on daily stem radial increment during the main growing period, and (b) identify the most important climatic factors related to the maximum growth rate of P. orientalis during the main growing period.
Materials and methods
Study area and local climate
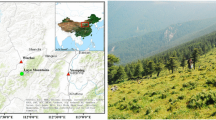
This study was conducted in a pure P. orientalis forest with an area of approximately 0.5 ha in Huailai (40°15′37.9″N, 115°36′39.6″E; 495 m a.s.l.), Hebei province, North China (Fig. 1). P. orientalis is the dominant forest tree species at the study sites, which were commonly on the tops and slopes of hills. Climatically this area belongs to the temperate semi-arid zone, and it has a temperate, semi-arid, continental monsoonal climate, which is cold and dry in winter and hot and rainy in summer (Tang and Zhang 2003). The mean annual precipitation is 376 mm, most of which falls from June to August (65 % of total annual precipitation). The mean annual temperature is 9.6 °C with a warmer period (>24 °C) from late June to early August (Fig. 2). The soil type is sandy loam, according to the United States Department of Agriculture (U.S.D.A.) standards.

Study area in North China (a), and the landscape of study area (b)

Comparison of the mean climatic conditions between May and October from 1963 to 2012 and during 2013 (lines 10-day mean air temperature, bars 10-day sum of precipitation). P precipitation, AT average temperature
Trees and dendrometer measurements
Four sample trees of P. orientalis, with stem diameters at breast height of 7.5 ± 0.5 cm and a stem height of 4.0 ± 0.4 m with well-developed crowns and no obvious wounds on the trunk, were selected for this study. From 7 May to 31 October 2013, the stem diameter variation at breast height was continuously monitored using a high-resolution automatic point dendrometer (Type DR, Ecomatic, Munich, Germany) with a resolution of 1 μm and a temperature coefficient <0.1 μm/K. The operating principle of the dendrometer is based on the use of a linearly variable differential transformer. As the stem expands and contracts, the sensing rod moves out and in and transmits a signal to the potentiometer (Deslauriers et al. 2003). Before installing the dendrometers, the outer dead bark was peeled off to reduce the influence of hygroscopic swelling and shrinkage of the bark. The radial variation values were recorded every 30 min using a data logger (DL10, Ecomatik, Munich, Germany).
Meteorological measurements
At an altitude of 2 m, air temperature (AT, °C), relative humidity (Rh, %) and precipitation (P, mm) were measured by an automatic weather station (HOBO, Onset, Pocasset, USA), which was located in a relatively flat area near the research site. The volumetric soil water content (SWC, %; Em50, Decagon, WA, USA) and soil temperature (ST, °C; Em50, Decagon, Washington, USA) were measured at 20-cm depth in close proximity to the trees studied. The vapor pressure deficit (VPD, kPa) was calculated from the values of ST and Rh. The environmental factors were recorded every 30 min, which was synchronous to the dendrometer measurements.
Data analysis
Based on the stem circadian cycle approach described by Downes et al. (1999), the typical diurnal cycle can be divided into three distinct phases as follows: (a) contraction phase, period between the morning maximum and daily minimum; (b) recovery phase, from the daily minimum to the position observed at that morning’s maximum; (c) stem radial increment (SRI) phase, from when the stem radius exceeds the morning maximum until the subsequent maximum. According to these definitions, some days did not experience any stem radial increment (SRI) phase (e.g., 23 July in Fig. 3), and those data were treated as missing values in the subsequent analyses (Downes et al. 1999). The daily stem mean value (R) was the average of 48 values measured in 1 day by the dendrometers, and the cumulative radial variation (CRV) was calculated as:

A stem’s circadian cycle divided into three distinct phases, recovery, contraction and stem radial increment (SRI)
where CRV n and R n are the cumulative radial variation (CRV) and daily stem mean value (R) on the day n, respectively, and R 1 is the daily stem mean value (R) on the first day. The RCRV was defined as the running mean of CRV for a time windows of 21 days, and the running means were calculated at the mid-point (days 11) of the 21-day window position. The RCRV was calculated as:
where RCRV n is the running mean of cumulative radial variation (CRV) for a time windows of 21 days on the day n. To describe the radial growth rate (RGR) of P. orientalis during the main growing season (Fig. 4) with negligible bias from putative stem shrinking and swelling effects, the RGR was determined by calculating the differences between the RCRV values for two consecutive days, according to Köcher’s study (2012), and a negative growth rate never occurred during the main growing period by this calculation. The RGR was calculated as:
where RGR n is the radial growth rate on the day n.
Pearson’s correlation coefficients were calculated using SPSS 17.0 software (SPSS Inc., Chicago, USA) to quantify the association between the daily stem radial increments (SRI) and the daily environmental variables during the main growing period (Deslauriers et al. 2007).

Daily time series (from 7 May to 31 October 2013) of cumulative radial variation (CRV) of P. orientalis (a), daily stem radial increment (SRI) during the main growing period (from 10 June to 20 September) (b), soil water content (c), precipitation (d), daily mean (ATmean), minimum (ATmin) and maximum air temperature (ATmax) (e), soil temperature (f), relative air humidity (g), and vapor pressure deficit (h). Rectangular boxes indicate the main growing period for P. orientalis, and bars represent the standard error
Results
Environmental variables during the growing season
From May to October 2013, the growing season, it was warmer by 0.7 °C than the long-term mean values (LTM 1963–2012), and the daily mean, maximum and minimum air temperatures during the growing season ranged from 4.6 to 28.5 °C, −2.5 to 24.1 °C and 11.4 to 39.0 °C, respectively (Fig. 4e). The variation pattern of soil temperature and of mean air temperature was similar with small fluctuations. The precipitation during the study period was higher (67.8 mm) than the LTM, and the highest amount of 10-day precipitation (70.7 mm) was recorded in early August (Fig. 4d). A serious drought period was recorded in May, when the total monthly precipitation was only 3 mm (LTM 28.5 mm), and the mean monthly air temperature was 20.9 °C (LTM 18.6 °C) (Fig. 2). The soil water content (SWC) showed a large fluctuant time course related to the occurrence of precipitation, and the highest SWC was observed in early August (Fig. 4c). The vapor pressure deficit (VPD) reached its highest value in May (Fig. 4h), while the relative air humidity (Rh) exhibited a different course from VPD, varying during the growing season from 20 to 100 % (Fig. 4g).
Dynamics of cumulative stem variation and daily stem radial increment
Serious drought period lasting for about 30 days was observed during the early growing season (in May). Under these drought conditions, the cumulative radial variation (CRV) of P. orientalis showed a persistent contraction until the occurrence of heavy precipitation in early June. Then, due to the precipitation, the CRV sharply increased within 4 days and resulted in an increase in soil water content (Fig. 4a). From mid-June to mid-September, the CRV was characterized by rapid growth as shown by an S-shaped growth curve, and this period was regarded as the main growing period for P. orientalis. During the late growing season (from late September to October), the CRV plateaued with small fluctuations, suggesting that the radial growth tended to stop at that time. During the main growing period (from mid-June to mid-September), the daily stem radial increment (SRI) was highly variable (Fig. 4b), ranging from 0.9 to 105.7 μm. The daily stem radial increment of P. orientalis during the main growing period was 27.6 ± 1.6 μm (mean ± SE) on average.
Stem radial growth rates
The stem radial growth rate of P. orientalis during the main growing period, calculated as the difference between the RCRV values (Fig. 5a) for two consecutive days, decreased from 20 June to 10 July and was followed by a unimodal curve from 10 July to 10 September (Fig. 5b). The radial growth rate reached its highest value (22.9 μm/day) in early August, the time with the best hydrothermal conditions, and its lowest value (0.3 μm/day) in mid-July, which was a short dry period.

Running mean of the cumulative radial variations (RCRV) for time windows of 21 days during the main growing period; the running means were calculated at the mid-point (days 11) of the 21-day window position (a), and daily time series of stem radial growth rate and soil water content (b)
Daily stem radial increment in relation to environmental factors
The daily stem radial increment (SRI) of P. orientalis was analyzed for its relation to environmental factors during the main growing period (Fig. 6). SRI showed the highest direct correlation with relative air humidity (Rh) (r = 0.675 p < 0.001) and precipitation (r = 0.572, p < 0.001). A significant indirect relationship was observed between SRI and vapor pressure deficit (VPD) (r = −0.653, p < 0.001). Non-significant correlations were found between SRI and soil temperature and soil water content. As for air temperature, ATmean and ATmax had significantly negative impacts on SRI.

Pearson’s correlations between the daily stem radial increments (SRI) of P. orientalis and climate parameters (daily mean air temperature, minimum air temperature, maximum air temperature, relative air humidity, vapor pressure deficit, precipitation, soil temperature and soil water content) during the main growing period
Discussion
Response to drought
In our study, a serious drought period was observed in May, and at the same time the stem of P. orientalis showed persistent shrinking (Fig. 4a). Zweifel et al. (2006) reported that for a certain period of drought, stem cambial cell division is not inhibited, and only when drought conditions last beyond that period, cell division is stopped. The inhibition of cambial cell expansion with the loss of cell turgor during drought is a well-known phenomenon (Abe and Nakai 1999). When drought ends after certain rainfall events and soil water is replaced, continuous rapid stem growth for days can be measured immediately (Drew et al. 2009). In our study, P. orientalis experienced a sharp stem radial increment within 4 days of a precipitation event in early June, and there was an increase in soil water content. To some extent, this rapid growth could be explained by the release of the low-pressure conditions in the cambium by a sudden enlargement of the already existing cells after division to their mature sizes (Zweifel et al. 2006). Although some of this growth was likely to be reversible swelling of phloem and cambial tissue (Zweifel et al. 2001), the stem of P. orientalis did not show an obvious contraction when the soil water content sharply decreased after 10 June (Fig. 4a, c), suggesting that irreversible cell growth resulted in a real net increment, with an increase of approximately 210 μm from early May to early June.
Weather dependence of growth
During the main growth period, the daily stem radial increment (SRI) of P. orientalis has been found to be significantly and positively related to relative air humidity (Rh), precipitation and soil water content (SWC) (Fig. 6). Physiological studies have shown that cellular enlargement is basically water dependent, because turgor pressure drives irreversible cell expansion (Proseus and Boyer 2005), and cell enlargement can be considered the major driving force of the net daily radial increase (Deslauriers et al. 2003). In our study, the strong positive effect of Rh on SRI could be the result of high Rh lowering transpiration, which reduces the negative pressure in the conducting system and increases the turgor in the stem cambial cells, thereby favoring cell division and expansion (Köcher et al. 2012). Similarly, Oberhuber and Gruber (2010), using dendrometers and a microsampling method, and Liang et al. (2006), using tree-ring analysis, found that the stem radial growth of Pinus sylvestris, Sabina przewalskii and Picea crassifolia were positively correlated with Rh under dry conditions. With respect to precipitation, on the one hand, large amounts of precipitation (>10 mm) caused an evident soil water content increase (Fig. 4c, d), which increases stem water status and cambial cell turgor, which then influences radial growth (Ryan et al. 1994). With precipitation, reversible variations due to the swelling of extensible tissues outside the cambium inevitably occurs (Deslauriers et al. 2007). On the other hand, small amounts of precipitation (<10 mm) exert a positive effect on cambial activity, not by increasing soil water content (Fig. 4c, d), but through wetting the crown; this leads to a partial release of the low leaf water potential, resulting in an increase in the cambial turgor and, thus, to an enlargement of the cambial cells already produced (Zweifel et al. 2006). This would also explain why there was a decoupling between radial growth and soil water content. Breshears et al. (2008) observed that foliar absorption of intercepted rainfall led to a substantial improvement in the water status of Juniperus monosperma during drought. In agreement with our results, a dendroclimatological study by Liang et al. (2008) found that monthly precipitation and relative air humidity in the growing season were major limiting factors for the radial growth of Pinus tabulaeformis in semi-arid areas of northern central China. These three environmental factors [relative air humidity (Rh), precipitation and soil water content (SWC)] can influence the moisture availability for the tree and, thereby, the stem radial growth, which also confirms that generally moisture availability is the dominant limiting factor for tree radial growth in semi-arid areas of North China based on dendroclimatological studies (Liang et al. 2001, 2008; Shi et al. 2008).
The significant negative response of SRI to vapor pressure deficit (VPD) and, especially, to daily maximum temperature (ATmax) and daily mean air temperature (ATmean) (Fig. 6) also indicates moisture stress. Under the conjoined influences of high VPD and ATmax, due to the close correlation between them (r = 0.76, p < 0.0001), the rate of transpiration of P. orientalis is increased. First, the negative pressure in the conducting system is increased, which decreases the turgor in the cambial cells and thereby decreases cell expansion (Köcher et al. 2012); this contrasts with the impact of Rh. Second, the tree closes its stomata to avoid hydraulic failure (Guehl and Aussenac 1987; Ripullone et al. 2007). By closing the stomata, carbon uptake and photosynthesis are affected, followed by a decrease of carbohydrate availability in the stem and the subsequent inhibition of radial growth (Oribe et al. 2003; Michelot et al. 2012). These results are supported by observations by Tian et al. (2005) who reported that P. orientalis showed a mid-day depression of net photosynthesis in a semi-arid region of North China related to high mid-day temperatures, namely the daily maximum temperature (ATmax). Moreover, high temperatures lead to higher respiration losses (Kajimoto 1990; Ryan et al. 1995), resulting in a faster consumption of carbohydrates and, hence, in a decreased radial growth; this also decreases the carbohydrate availability for earlywood formation in the next spring (Lebourgeois et al. 2004; Friedrichs et al. 2009). By dendroclimatological study, Huang et al. (2006) found that the ring width of P. orientalis has a significant positive response to precipitation and a significant negative response to temperature during the current growing season, which was consistent with our results. Soil temperature and daily minimum temperature did not have significant impacts on the SRI (Fig. 6). In contrast, the stem radial growth of conifers growing in cold conditions, such as at the alpine tree line, may benefit from high soil and air temperatures during the growing season (Rossi et al. 2007; Gruber et al. 2009; Dong et al. 2011), because tree growth is strongly regulated by low temperatures (Körner and Paulsen 2004).
Timing of maximum growth rate
Several authors (Rossi et al. 2006; Seo et al. 2008; Gruber et al. 2009; Duchesne et al. 2012) reported that the maximum daily growth rate of confiners in cold environments, i.e., at the alpine tree line and boreal forests, peaked around the summer solstice, and not during the warmest period of the year. Rossi et al. (2006) hypothesized that the connection of the growth constraint with the maximum photoperiod could be an adaptation of conifers to cold conditions, allowing them enough time to complete cell wall formation and lignification before winter. However, in our study, the maximum growth rate of P. orientalis was observed in early August with its favorite growing conditions, i.e., the highest soil water content during the main growing season (Fig. 4c), and a rather steady daily mean air temperature of approximately 24 °C in summer (mid-June to mid-August) (Figs. 2, 4e). There was a significant correlation between the stem radial growth rate and daily soil water content (r = 0.39, p < 0.001). Zhang et al. (2012) reported that the net photosynthesis rate of P. orientalis was closely and positively correlated with soil water content between 3 and 16 %, which also explains our results with respect to the physiological process. Our results support the conclusion by Henttonen et al. (2009) that the timing of the highest growth rate does not seem to be related to maximum day length as strictly as was suggested by Rossi et al. (2006), and in milder climatic conditions, other environmental variables, such as temperature and water availability, may have an effect on the timing of the maximum growth rate. We suggest that the timing of the maximum growth rate reflects the ability of the species to exploit favorable environmental conditions and could be considered as an adaptation to local climatic conditions. In our study, the maximum radial growth rate of P. orientalis in a semi-arid area of North China was regulated by soil water content in summer. This seems to be an adaptation to the local, temperate continental, monsoon climate conditions, with high precipitation and high temperature at the same time in early August simultaneously (Fig. 2). In accordance with our results, Popescu et al. (1968) reported that the maximum increment rate for Fagus sylvatica in Romania was reached in late July under conditions of optimal temperature and water availability.
Conclusions
From our analysis, we conclude that the daily stem radial increment (SRI) of P. orientalis during the main growing period in a semi-arid area of North China was primarily influenced by relative air humidity, precipitation, vapor pressure deficit and daily air maximum temperature, which mainly influenced the moisture availability for the tree and, thereby, the daily stem radial growth. Daily mean air temperature and soil water content played a secondary role. The stem radial growth was slowed down under drought conditions, but a rapid growth release occurred when water became available. The stem radial growth rate of P. orientalis reached its highest value in early August, which was related to soil water content in summer, and this effect appears to be an adaptation to the local, temperate continental, monsoon climate.
References
Abe H, Nakai T (1999) Effect of the water status within a tree on tracheid morphogenesis in Cryptomeria japonica D-Don. Trees 14:124–129
Bräuning A, Volland-Voigt F, Burchardt I, Ganzhi O, Nauss T, Peters T (2009) Climatic control of radial growth of Cedrela montana in a humid mountain rainforest in Southern Ecuador. Erdkunde 63:337–345
Breshears DD, McDowell NG, Goddard KL, Dayem KE, Martens SN, Meyer CW, Brown KM (2008) Foliar absorption of intercepted rainfall improves woody plant water status most during drought. Ecology 89:41–47
Cherubini P, Gartner BL, Tognetti R, Braker OU, Schoch W, Innes JL (2003) Identification, measurement and interpretation of tree rings in woody species from mediterranean climates. Biol Rev 78:119–148
Deslauriers A, Morin H, Urbinati C, Carrer M (2003) Daily weather response of balsam fir (Abies balsamea (L.) Mill.) stem radius increment from dendrometer analysis in the boreal forests of Quebec (Canada). Trees 17:477–484
Deslauriers A, Rossi S, Anfodillo T (2007) Dendrometer and intra-annual tree growth: what kind of information can be inferred? Dendrochronologia 25:113–124
Dong MY, Jiang Y, Zhang WT, Yang YG, Yang HC (2011) Effect of alpine treeline conditions on the response of the stem radial variation of Picea meyeri Rebd. et Wils. to environmental factors. Pol J Ecol 59:729–739
Downes G, Beadle C, Worledge D (1999) Daily stem growth patterns in irrigated Eucalyptus globulus and E. nitens in relation to climate. Trees 14:102–111
Drew DM, Downes GM (2009) The use of precision dendrometers in research on daily stem size and wood property variation: a review. Dendrochronologia 27:159–172
Drew DM, Downes GM, Grzeskowiak V, Naidoo T (2009) Differences in daily stem size variation and growth in two hybrid eucalypt clones. Trees 23:585–595
Duchesne L, Houle D, D’Orangeville L (2012) Influence of climate on seasonal patterns of stem increment of balsam fir in a boreal forest of Quebec, Canada. Agric For Meteorol 162:108–114
Dulamsuren C, Hauck M, Bader M, Osokhjargal D, Oyungerel S, Nyambayar S, Runge M, Leuschner C (2009) Water relations and photosynthetic performance in Larix sibirica growing in the forest-steppe ecotone of northern Mongolia. Tree Physiol 29:99–110
Friedrichs DA, Trouet V, Buntgen U, Frank DC, Esper J, Neuwirth B, Loffler J (2009) Species-specific climate sensitivity of tree growth in Central-West Germany. Trees 23:729–739
Fritts HC, Shatz DJ (1975) Selecting and characterizing tree-ring chronologies for dendroclimatic analysis. Tree Ring Bull 35:31–46
Gebrekirstos A, Mitlohner R, Teketay D, Worbes M (2008) Climate-growth relationships of the dominant tree species from semi-arid savanna woodland in Ethiopia. Trees 22:631–641
Gruber A, Zimmermann J, Wieser G, Oberhuber W (2009) Effects of climate variables on intra-annual stem radial increment in Pinus cembra (L.) along the alpine tree line ecotone. Ann For Sci 66:503. doi:10.1051/forest/2009038
Guehl JM, Aussenac G (1987) Photosynthesis decrease and stomatal control of gas-exchange in Abies Alba Mill in response to vapor-pressure difference. Plant Physiol 82:316–322
Guo ZJ, Song XD, Zhao HG (2007) Biomass and soil characteristics of Hippophae rhamnoides and Platycladus oriental in mixed forest and its root system distribution. Bull Soil Water Conservat 27:18–23 (in Chinese)
Gutiérrez E, Campelo F, Julio Camarero J, Ribas M, Muntan E, Nabais C, Freitas H (2011) Climate controls act at different scales on the seasonal pattern of Quercus ilex L stem radial increments in NE Spain. Trees 25:637–646
Henttonen HM, Makinen H, Nöjd P (2009) Seasonal dynamics of the radial increment of Scots pine and Norway spruce in the southern and middle boreal zones in Finland. Can J Forest Res 39:606–618
Huang RF, Zhao YK, Lv JX, Bao FC (2006) Response of ring width and ring density of Platycladus orientalis to climate change in Beijing. Scientia Silvae Sinicae 42:78–82 (in Chinese)
Kajimoto T (1990) Photosynthesis and respiration of Pinus pumila needles in relation to needle age and season. Ecol Res 5:333–340
Köcher P, Horna V, Leuschner C (2012) Environmental control of daily stem growth patterns in five temperate broad-leaved tree species. Tree Physiol 32:1021–1032
Körner C, Paulsen J (2004) A world-wide study of high altitude treeline temperatures. J Biogeogr 31:713–732
Lebourgeois F, Cousseau G, Ducos Y (2004) Climate-tree-growth relationships of Quercus petraea Mill stand in the Forest of Berce (“Futaie des Clos”, Sarthe, France). Ann For Sci 61:361–372
Lebourgeois F, Merian P, Courdier F, Ladier J, Dreyfus P (2012) Instability of climate signal in tree-ring width in Mediterranean mountains: a multi-species analysis. Trees 26:715–729
Lei HP, Wang YG, Liang FY, Su WW, Feng YF, Guo XL, Wang N (2010) Composition and variability of essential oils of Platycladus orientalis growing in China. Biochem Syst Ecol 38:1000–1006
Liang EY, Shao XM, Hu YX, Lin JX (2001) Dendroclimatic evaluation of climate-growth relationships of Meyer spruce (Picea meyeri) on a sandy substrate in semi-arid grassland, north China. Trees 15:230–235
Liang EY, Shao XM, Eckstein D, Huang L, Liu XH (2006) Topography- and species-dependent growth responses of Sabina przewalskii and Picea crassifolia to climate on the northeast Tibetan Plateau. For Ecol Manag 236:268–277
Liang EY, Eckstein D, Liu HY (2008) Climate-growth relationships of relict Pinus tabulaeformis at the northern limit of its natural distribution in northern China. J Veg Sci 19:393–406
Liu HY, Park Williams A, Allen CD, Guo DL, Wu XC, Anenkhonov OA, Liang EY, Sandanov DV, Yin Y, Qi ZH, Badmaeva NK (2013) Rapid warming accelerates tree growth decline in semi-arid forests of Inner Asia. Glob Chang Biol 19:2500–2510
Mäkinen H, Seo J-W, Nöjd P, Schmitt U, Jalkanen R (2008) Seasonal dynamics of wood formation: a comparison between pinning, microcoring and dendrometer measurements. Eur J Forest Res 127(3):235–245
Michelot A, Simard S, Rathgeber C, Dufrene E, Damesin C (2012) Comparing the intra-annual wood formation of three European species (Fagus sylvatica, Quercus petraea and Pinus sylvestris) as related to leaf phenology and non-structural carbohydrate dynamics. Tree Physiol 32:1033–1045
Oberhuber W, Gruber A (2010) Climatic influences on intra-annual stem radial increment of Pinus sylvestris (L.) exposed to drought. Trees 24:887–898
Oribe Y, Funada R, Kubo T (2003) Relationships between cambial activity, cell differentiation and the localization of starch in storage tissues around the cambium in locally heated stems of Abies sachalinensis (Schmidt) Masters. Trees 17:185–192
Pérez CA, Carmona MR, Aravena JC, Fariña JM, Armesto JJ (2009) Environmental controls and patterns of cumulative radial increment of evergreen tree species in montane, temperate rainforests of Chiloe Island, southern Chile. Austral Ecol 34(3):259–271
Popescu O, Parascan D, Marcu M (1968) On the annual course of radial growth of beech. Allg Forst Jagdztg 139:190–195 (in German)
Proseus TE, Boyer JS (2005) Turgor pressure moves polysaccharides into growing cell walls of Chara corallina. Ann Bot 95:967–979
Ripullone F, Guerrieri MR, Nole A, Magnani F, Borghetti M (2007) Stomatal conductance and leaf water potential responses to hydraulic conductance variation in Pinus pinaster seedlings. Trees 21:371–378
Rossi S, Deslauriers A, Anfodillo T, Morin H, Saracino A, Motta R, Borghetti M (2006) Conifers in cold environments synchronize maximum growth rate of tree-ring formation with day length. New Phytol 170:301–310
Rossi S, Deslauriers A, Anfodillo T, Carraro V (2007) Evidence of threshold temperatures for xylogenesis in conifers at high altitudes. Oecologia 152:1–12
Rotenberg E, Yakir D (2010) Contribution of semi-arid forests to the climate system. Science 327:451–454
Ryan DAJ, Allen OB, Mclaughlin DL, Gordon AM (1994) Interpretation of sugar maple (Acer Saccharum) ring chronologies from central and southern Ontario using a mixed linear-model. Can J Forest Res 24:568–575
Ryan MG, Gower ST, Hubbard RM, Waring RH, Gholz HL, Cropper WP, Running SW (1995) Woody tissue maintenance respiration of 4 conifers in contrasting climates. Oecologia 101:133–140
Seo JW, Eckstein D, Jalkanen R, Rickebusch S, Schmitt U (2008) Estimating the onset of cambial activity in Scots pine in northern Finland by means of the heat-sum approach. Tree Physiol 28(1):105–112
Shi JF, Liu Y, Vaganov EA, Li JB, Cai QF (2008) Statistical and process-based modeling analyses of tree growth response to climate in semi-arid area of north central China: a case study of Pinus tabulaeformis. J Geophys Res 1113:G1026. doi:10.1029/2007JG000547
Tang HP, Zhang XS (2003) Establishment of optimized eco-productive paradigm in the farming-pastoral zone of northern China. Acta Bot Sin 45:1166–1173
Tian JH, He KN, Wang BT, Guo H, Zhang WQ, Yin J (2005) Diurnal course of gas exchange and water use efficiency of Platycladus orientalis in a semi-arid region of the Loess Plateau. J Beijing For Uni 27:42–46 (in Chinese)
Volland-Voigt F, Bräuning A, Ganzhi O, Peters T, Maza H (2011) Radial stem variations of Tabebuia chrysantha (Bignoniaceae) in different tropical forest ecosystems of southern Ecuador. Trees 25:39–48
Westermann J, Zerbe S, Eckstein D (2008) Age structure and growth of degraded Populus euphratica floodplain forests in North-West China and perspectives for their recovery. J Integr Plant Biol 50(5):536–546
Williams AP, Allen CD, Millar CI, Swetnam TW, Michaelsen J, Still CJ, Leavitt SW (2010) Forest responses to increasing aridity and warmth in the southwestern United States. Proc Natl Acad Sci USA 107:21289–21294
Xiao SC, Xiao HL, Peng XM, Tian QY (2014) Daily and seasonal stem radial activity of Populus euphratica and its association with hydroclimatic factors in the lower reaches of China’s Heihe River basin. Environ Earth Sci 72:609–621
Zhang GC, Xia JB, Shao HB, Zhang SY (2012) Grading woodland soil water productivity and soil bioavailability in the semi-arid Loess Plateau of China. Clean Soil Air Water 40:148–153
Zweifel R, Item H, Hasler R (2001) Link between diurnal stem radius changes and tree water relations. Tree Physiol 21:869–877
Zweifel R, Zimmermann L, Zeugin F, Newbery DM (2006) Intra-annual radial growth and water relations of trees: implications towards a growth mechanism. J Exp Bot 57:1445–1459
Author contribution statement
Study conception and design: J.Y. and D.M.Y. Acquisition of data: W.B.Q., W.M.C., H.Y.M. and W.B. Analysis and interpretation of data: W.B.Q. and D.M.Y. Drafting of manuscript: J.Y., W.B.Q. and D.M.Y.
Acknowledgments
This work was supported by grants from the National Natural Science Foundation of China (No. 41401061, 41171067, 41271059), the Fundamental Research Funds for the Central Universities (No. 2013NT63), and the Special Basic Research Program of Science and Technology, Ministry of Science and Technology of China (No. 2011FY110300). The authors would like to thank the reviewers for their invaluable suggestions.
Conflict of interest
The authors declare that they have no conflict of interest.
Author information
Authors and Affiliations
Corresponding authors
Additional information
Communicated by U. Luettge.
Rights and permissions
About this article

Cite this article
Jiang, Y., Wang, BQ., Dong, MY. et al. Response of daily stem radial growth of Platycladus orientalis to environmental factors in a semi-arid area of North China. Trees 29, 87–96 (2015). https://doi.org/10.1007/s00468-014-1089-8
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00468-014-1089-8