
Abstract
A new species of caligid copepod, Caligus mulli n. sp., is described based on specimens collected from surmullet Mullus surmuletus Linnaeus from Atlantic waters off the Azores, Portugal, and from red mullet Mullus barbatus barbatus (Linnaeus) caught in the Mediterranean Sea, off the south-eastern coast of Turkey. Sixteen specimens (both sexes) were collected from the gill cavity of M. surmuletus, and one female was collected from the gill cavity of M. barbatus barbatus. The new species is described from both fish hosts. Detailed comparisons are presented with two closely related species, C. ocyurus Cressey, 1991 and C. biaculeatus Brian, 1914: the former can be distinguished by differences in shape of the dorsal cephalothoracic shield and postantennal process, while the latter differs in relative size of the distal spines on leg 4 and in the shape of the sternal furca. The new species can easily be distinguished from its congeners by a combination of the following additional characteristics: (1) the corpus of the female maxilliped bears a prominent projection proximally and the myxal area is ornamented with a hyaline flange and two sensillae, and (2) the male maxilliped has a conical proximal projection on the corpus and a bifid myxal process opposing the claw.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The genus Caligus O.F. Müller, 1785 was first established to accommodate Caligus curtus Müller, 1785, the type species. Since then, it has grown to become the most speciose genus in the family Caligidae, currently comprising 266 valid species (Boxshall 2018) which are, with rare exceptions, parasitic on marine and brackish water fishes. During the twentieth century, the number of known species increased significantly, with the recognition of 177 additional species, an average of 1.77 new species per year (Fig. 1). Since the beginning of the twenty-first century, the rate of discovery of new species has increased, with the description of another 40 new species, at a rate of 2.22 per year. This includes the recent addition of a further nine new species of Caligus plus the transfer of four other species into Caligus from other caligid genera (Boxshall 2018).

The accumulation of valid species of the genus Caligus from 1785 to the present
The parasite fauna of Mediterranean fishes has been intensively studied and is better known than other regional faunas. Raibaut et al. (1998) reviewed historic records and reported the presence of 26 species of Caligus within the Mediterranean, although this number included at least six nomina nuda listed by Richiardi (1880) but never described. Since Raibaut et al.’s excellent review, another nine species have been reported and these represent a mix of new species (e.g., C. solea Demirkale, Özak, Yanar & Boxshall 2014), invasives from the Red Sea (e.g. C. lagocephali Pillai, 1961 (as C. fugu Yamaguti & Yamasu, 1959) and first reports of Atlantic species hitherto unreported from the Mediterranean (e.g. C. temnodontis Brian, 1924 and C. macrurus Heller, 1865) (Demirkale et al. 2014; Özak et al. 2010, 2012, 2017). Currently, there are 29 valid species of the genus Caligus reported from Mediterranean fish species (Özak et al. 2017).
Surmullet, Mullus surmuletus (Linnaeus) and Red mullet, Mullus barbatus barbatus (Linnaeus), are common and commercially important fishes of the family Mullidae along the eastern Atlantic coast and in the Mediterranean basin. Their parasites have been repeatedly surveyed and, to date, a total of only eight species of parasitic copepods have been reported from these two mullid species (Table 1).
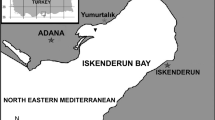
Considering the intensity of sampling of these hosts, it was surprising to discover a species of Caligus that was previously unknown from the Atlantic and the Mediterranean. The new species described below was first collected from M. b. barbatus captured during a parasitological survey of the fishes of Iskenderun Bay, Turkey in 2014. However, only a single female was collected from the gill cavity of its host after the examination of a large sample of 3000 specimens of this commercially important fish. Two years later, 16 specimens were collected from M. surmuletus captured during a research project on “Developing methods for capture, husbandry and live transportation of aquarium fishes” conducted in oceanic waters off Faial Island, in the Azores archipelago. Five (three females and two males) of the 16 specimens collected were sent to the first author for identification. Comparative morphological examination of the Caligus specimens collected from the two different mullid hosts revealed that the single female Caligus collected from M. b. barbatus from the Mediterranean was identical with the Atlantic specimens collected from M. surmuletus. The discovery of the new species of Caligus on these two hosts raises the number of parasitic copepods reported from M. barbatus barbatus from five to six and from M. surmuletus from seven to eight. In addition, it raises the number of species of Caligus known from the Mediterranean to 30.
Materials and methods
Sixteen specimens were collected from Mullus surmuletus (n = 14; total body length range 8–10 cm) caught in oceanic waters off Faial Island, in the Azores archipelago, and one female was sampled from the gill cavity of Mullus barbatus barbatus (n = 3000; total body length range 15–22 cm) caught in Iskenderun Bay, Turkey. The Atlantic specimens were collected from the host by applying the freshwater treatment method of Wildgoose (2001) as follows: the fish was initially placed in a 5-L tank containing 36 ppt seawater; the salinity was then decreased to 9 ppt within 35 min; the fish was kept at 9 ppt for 30 min, after which time the salinity was increased slowly from 9 to 36 ppt. After the osmotic shock, a total of 16 specimens of a parasitic copepod were collected from the bottom of the tank by filtering the water through a plankton net (mesh size 100 μ). The Mediterranean specimen was collected during the macroscopic examination of the fishes. The parasitic copepods were immediately preserved in 70% ethanol. Atlantic and Mediterranean specimens were cleared in lactic acid for 2 h prior to examination using a Nikon SMZ 800N dissecting microscope and an Olympus BX51 compound microscope. Subsequently, the specimens were mounted as temporary preparations in a drop of lactic acid on a cavity slide. Measurements were made using an ocular micrometre and drawings were made with the aid of a drawing tube. All measurements are in millimetres unless otherwise stated. The scientific and common names of fishes follow Froese and Pauly (2018) and the morphological terminology for the copepods follows Boxshall (1990) and Huys and Boxshall (1991). The holotype female (CUMAP-COP/2017-29) and holotype male (CUMAP-COP/2017-30) are stored in the collections of the Aquatic Parasitology Museum at the Faculty of Fisheries, University of Çukurova, Adana-Turkey.
Family Caligidae Burmeister, 1835
Genus Caligus Müller, 1785
Caligus mulli n. sp.
Atlantic specimens: (n = 16; 14 females, 2 males)
Type host: Mullus surmuletus Linnaeus, 1758 (Mullidae)
Type locality: Atlantic waters off Faial Island, Azores, Portugal; collected by Alfredo M.V. Rodrigues on 19 Oct. 2016.
Site on host: Dorsal body surface.
Prevalence: 85.7% (12 fish infected of a total of 14 examined)
Mediterranean specimen: (n = 1, 1 female)
Host: Mullus barbatus barbatus (Linnaeus, 1758) (Mullidae)
Locality: Northeastern Mediterranean waters off Karatas¸ Iskenderun Bay, Turkey; collected by A. A.Özak (15 Feb. 2014).
Depth range: 30–50 m; mean surface water temperature 13.5 °C; salinity 35 ppt.
Site on host: Gill cavity
Prevalence: 0.0003% (1 fish infected of a total of 3000 examined)
Type material: holotype female [CUMAP-COP/2017–29]; holotype male [CUMAP-COP/2017–30] stored in the collection of the Aquatic Parasitology Museum of the Faculty of Fisheries, University of Çukurova (CUMAP), Adana, Turkey.
Etymology: The species name refers to the host genus.
Adult female
Body caligiform, comprising cephalothorax incorporating first to third pedigerous somites, free fourth pedigerous somite, genital complex and 1-segmented abdomen (Fig. 2a). Body length 2.72 (2.68–2.77) (n = 4) excluding caudal setae. Dorsal cephalothoracic shield subtriangular, longer than wide, 1.44 (1.34–1.49) × 1.25 (1.20–1.31); thoracic zone of shield 0.75 (0.7–0.79) × 0.97 (0.9–1.1), comprising slightly more than half total length (52%) of cephalothorax; posterior margin of thoracic zone extending beyond posterior margins of lateral zones. Fourth pedigerous somite distinctly separate from genital complex, wider than long, 0.13 (0.09–0.15) × 0.30 (0.24–0.36). Genital complex (Fig. 2b) 1.36 times longer than wide, 0.94 (0.91–0.97) × 0.69 (0.63–0.74); with slightly rounded anterior angles, parallel sides and with rounded posterolateral corners; about 4.95 times longer than 1-segmented abdomen. Abdomen slightly longer than wide, 0.19 (0.15–0.22) × 0.18 (0.14–0.2). Combined length of genital complex and entire abdomen (excluding caudal rami) approximately 78% of length of cephalothorax. Caudal rami longer than wide, 0.11 (0.09–0.13) × 0.08 (0.05–0.1), armed with 6 pinnate setae.

Caligus mulli n. sp. holotype female. a Habitus, dorsal view; b genital complex, ventral view; c antennule; d antenna; e postantennal process; f maxillule. Scale bars: a and b, 0.5 mm; c–f, 100 μm
Antennule (Fig. 2c) 2-segmented, proximal segment distinctly wider than distal, armed with 25 plumose setae on anterior and antero-ventral surfaces plus 2 unarmed setae located dorsally; distal segment armed with 1 subterminal seta on posterior margin and 11 setae plus 2 aesthetascs around distal margin. Antenna (Fig. 2d) uniramous, 3-segmented; proximal segment produced posteriorly into blunt spinous process; middle segment with small adhesion pad on dorsal surface; distal segment forming recurved claw with small setae proximally and distally. Postantennal process (Fig. 2e) weakly curved, carrying 2 bi-sensillate papillae and similar sensillate papilla located on body surface adjacent to process. Maxillule (Fig. 2f) comprising dentiform posterior process with a minute denticle on medioventral surface and rounded tip, anterior papilla bearing 3 setae, 1 long and 2 small. Mandible (Fig. 3a) curved inward distally and with 12 teeth on one side near apex. Sternal furca (Fig. 3b) with small box and bluntly pointed, divergent tines curved inward distally, each with large flange along outer margin. Maxilla (Fig. 3c) brachiform and 2-segmented; proximal segment (lacertus) unarmed, distal segment (brachium) slender bearing subterminal hyaline membrane on outer margin, plus short canna and long calamus distally (Fig. 3d). Maxilliped (Fig. 3e) comprising large proximal segment (corpus) and distal subchela representing fused endopodal segments plus claw; outer margin of corpus with small subtriangular process proximally; subchela armed with small seta at base of claw; claw extending about to mid-length of corpus.

Caligus mulli n. sp. holotype female. a Mandible; b sternal furca; c maxilla; d calamus and canna of maxilla; e maxilliped; f leg 1; g, distal exopodal segment of leg 1. Scale bars: a–c, f, g 100 μm; d, 50 μm
Swimming leg 1 (Fig. 3f) biramous, with 2-segmented exopod and unsegmented vestigial endopod. Sympod armed with lateral plumose seta and inner seta. First exopodal segment ornamented with row of setules along free posterior margin and bearing small spine at outer distal corner. Distal exopodal segment (Fig. 3g) with 3 plumose setae posteriorly plus 4 distal margin elements; spine 1 (outermost) simple, spines 2 and 3 about equal in length and each bearing single accessory process, seta 4 (innermost) longer than spines but shorter than segment.
Leg 2 (Fig. 4a) biramous with 3-segmented rami. Coxa small with large pinnate seta on posterior margin. First and second exopodal segments (Fig. 4b) each with pinnate seta on inner margin and long spine at outer distal corner reflexed obliquely back across surface of segment; both spines finely bilaterally serrated. Third exopodal segment (Fig. 4b) with five inner plumose setae plus three outer spines; proximal outer spine (smallest) naked, middle spine rounded, extending over base of distal spine; distal spine ornamented with hyaline membrane along outer margin and row of setules along inner margin. First endopodal segment (Fig. 4c) armed with inner plumose seta and ornamented with row of setules along outer margin; second endopodal segment with two inner plumose setae plus dense ornamentation of setules along outer margin; third segment with six plumose setae (Fig. 4c).

Caligus mulli n. sp. holotype female. a Leg 2; b exopodal segments of leg 2; c endopodal segments of leg 2. Scale bars: a, 200 μm; b and c, 100 μm
Leg 3 (Fig. 5a) forming flattened plate closing posterior part of cephalothoracic sucker as typical for genus. Protopodal part flattened and joined by plate-like intercoxal sclerite, forming apron ornamented with marginal membrane posteriorly and along lateral margin anterior to exopod: bearing inner plumose seta at junction with intercoxal plate and outer plumose seta dorsal to base of exopod; sensillae located adjacent to inner coxal seta and adjacent to origin of endopod. Exopod (Fig. 5a) 3-segmented; first segment armed with straight outer claw directed over ventral surface of ramus, slightly shorter than segment; second segment with slender outer spine and inner plumose seta; third with three outer spines and four inner plumose setae; middle spine shorter than proximal or distal; outer margins of segments 2 and 3 ornamented with row of slender setules. Endopod 2-segmented (Fig. 5a); first segment forming flap-like velum ornamented with row of fine setules along free margin, closing space between rami and armed with inner plumose seta; compound distal segment expanded laterally, armed with six setae increasing in length from outermost to innermost.

Caligus mulli n. sp. holotype female. a Leg 3; b leg 4; c leg 5. Scale bars: a 200 μm; b and c 100 μm
Leg 4 (Fig. 5b) uniramous, comprising long protopodal segment armed with outer pinnate seta plus 2-segmented exopod; first exopodal segment with one short distal spine, not quite reaching middle of margin of second exopodal segment; compound second segment with one lateral spine plus three apical spines along oblique distal margin, longest spine about twice as long as shortest; each spine with pecten at base.
Spine (Roman numerals) and seta (Arabic numerals) formula of legs 1–4 as follows:
Exopod | Endopod | |
Leg 1 | I-0; III,1,3 | Vestigial |
Leg 2 | I-1; I-1; II,I,5 | 0-1; 0-2; 6 |
Leg 3 | I-0; I-1; III,4 | 0-1; 6 |
Leg 4 | I-0; I,III | Absent |
Leg 5 (Fig. 5c) located at posterolateral corner of genital complex, represented by two papillae; outer (protopodal) papilla bearing single pinnate seta; inner (exopodal) papilla bearing two plumose setae.
Adult male. Body (Fig. 6a) 1.37–1.39 mm (n = 2) long, excluding caudal setae. Cephalothoracic shield subtriangular, slightly longer than wide, 0.8–0.9 × 0.72–0.77, excluding marginal hyaline membranes. Free thoracic zone of shield wider than long, 0.43–0.48 × 0.56–0.59. Fourth pedigerous somite 0.09–0.12 × 0.17–0.19, indistinctly divided from genital complex. Genital complex (Fig. 6a) 0.17–0.19 × 0.17–0.19, with convex sides and subtriangular posterolateral corners. Abdomen (Fig. 6f) comprising two somites; first free abdominal somite 0.07–0.09 × 0.1–0.1, shorter than anal somite, 0.18–0.2 × 0.16–0.18; ventral surface of anal somite ornamented with sensillae. Caudal rami (Fig. 6f) longer than wide, 0.1–0.09 × 0.07–0.06, bearing six pinnate setae.

Caligus mulli n. sp. holotype male. a Habitus, dorsal view; b antenna; c postantennal process; d maxillule; e maxilliped; f legs 5 and 6. Scale bars: a, 0.5 mm; b–e, 100 μm; f, 200 μm
Antennule as in female. Antenna (Fig. 6b) 3-segmented; proximal segment long, narrow, with corrugated adhesion pad on outer surface proximally; middle segment largest, with corrugated pads proximally and on outer and medial distal surfaces; distal segment of antenna forming recurved claw consisting of three overlapping triangular plates and armed with two slender basal setae. Postantennal process (Fig. 6c) more strongly curved than that of female, inner margin with small protuberance on mid-part, carrying two bi-sensillate papillae; similar bi-sensillate papilla located on body surface near postantennal process. Maxillule (Fig. 6d) with small dentiform knob located medially on posterior spinous process plus an anterior papilla carrying two medium-length and one long setae; deniform process corrugated at base. Mandible and maxilla as in female. Maxilliped (Fig. 6e) with massive corpus carrying conspicuous bifid process on myxal margin, opposing claw; corpus with prominent conical process on outer ventral surface; subchela armed with small seta at base of claw. Sternal furca and legs 1–4 as in female. Leg 5 (Fig. 6f) represented by two papillae located on posterolateral margins of genital complex, outer papilla with 1 and inner (exopodal) papilla with two plumose setae. Leg 6 (Fig. 6f) represented by single papilla bearing two unequal setae on posteroventral side of genital complex.
Remarks
The new species belongs to a large group of species within Caligus that is united by the possession of a 3-segmented leg 4 in which the distal exopodal segment is compound (representing the fused second and third exopodal segments) and carries four spines. More than 90 valid species of Caligus share this same configuration of leg 4. Specimens of the new species also retain the three plumose setae on the posterior margin of the distal exopodal segment of leg 1. This eliminates all members of the Caligus productus-group which are characterised by the loss or major reduction of these posterior margin setae (Boxshall and El-Rashidy 2009; Boxshall 2018). A total of nearly 50 species share these two character states. However, only ten of these also have a genital complex in the female that is both markedly longer than wide and more than three times longer than the free abdomen. Within this group of ten species, only two, Caligus biaculeatus Brian, 1914 and C. ocyurus Cressey, 1991, share the possession of a genital complex in the female that is about five times longer than the 1-segmented free abdomen (the anal somite).
Caligus ocyurus was described on the basis of material of both sexes collected from the yellow snapper, Ocyurus chrysurus Bloch, caught at Carrie Bow Cay in Belize (Cressey 1991). Estimating from the measurements provided by Cressey (1991), the genital complex of the adult female is 1.4 times longer than wide and 5.2 times longer than the 1-segmented abdomen. This species also has bisensillate papillae on the postantennal process, outer margin spines on exopodal segments 1 and 2 of leg 2 that are reflexed obliquely across the surface of the ramus and a weakly curved outer spine on the first exopodal segment of leg 3 that does not reach the distal margin of the second segment. Like most Caligus species, C. ocyurus has three outer margin spines on the third exopodal segment of leg 3 but, unusually, the middle spine is shorter than both the proximal and the distal spines. All of these features are shared with the new species. However, the new species differs in having a straight postantennal process, compared to a recurved and flanged process in C. ocyurus, and the dorsal cephalothoracic shield is subtriangular whereas that of C. ocyurus is rather elongate with nearly parallel lateral margins (in dorsal view). In addition, the sternal furca of the new species has inwardly curved, blunt tipped tines (vs. straight and sharply pointed tines), the proximal segment of the male antenna is subrectangular (vs. irregular in C. ocyurus) and the recurved terminal segment of male antenna comprises three overlapping subtriangular elements (vs. two subtriangular recurved elements in C. ocyurus).
Caligus biaculeatus was originally described on the basis of material recovered free within a deep-water trawl that contained specimens of Macrourus Bloch and Bathygadus Günther taken off the Madeira Islands (Brian 1914). The assumption was, presumably, that it may have utilised such deep-sea fishes as hosts even though we now know that caligids in general are rarely found in deep water (Boxshall 1998). Cressey (1991) challenged Brian’s assumption: he considered that the hosts were more likely fishes of the shallow waters surrounding the Madeira Islands. He re-examined the holotype female of C. biaculeatus stored in the Musée Océanographique, Monaco and compared it with 19 lots of new material collected from eight different hosts caught off the coast of Belize (Cressey 1991). The most common hosts of C. biaculeatus in Belize were Aulostomus maculatus Valenciennes, 1841, Halichoeres bivittatus (Bloch, 1791) and Sparisoma viride (Bonnaterre, 1788)—three very different fish from three different families, Aulostomidae, Labridae and Scaridae, respectively. Cressey (1991) redescribed female C. biaculeatus and concluded that this parasite was “not very host specific”.
Calculating from measurements given by Cressey (1991), the genital complex of female C. biaculeatus is about 2.3 times longer than wide and about 4.9 times longer than the 1-segmented abdomen. This species also has bisensillate papillae on the weakly curved postantennal process, spine 1 on the distal margin of the exopod of leg 1 is distinctly shorter than spines 2 and 3, the outer margin spines on exopodal segments 1 and 2 of leg 2 that are reflexed obliquely across the surface of the ramus, and the outer spine on the first exopodal segment of leg 3 is weakly curved and does not reach the distal margin of the second segment. It also shares with C. ocyurus, the unusual configuration of the three outer margin spines on the third exopodal segment of leg 3 in which the middle spine is shorter than both the proximal and the distal spines. The new species also shares this configuration of spines on the distal exopodal segment of leg 3, has bisensillate papillae on the postantennal process, a short spine 1 on the distal margin of the exopod of leg 1 and reflexed outer margin spines on the first and second exopodal segments of leg 2. It is clearly closely related to C. biaculeatus.
The females redescribed by Cressey (1991) carried four spines on the compound distal exopodal segment of leg 4, and the outermost spine on the oblique distal margin was tiny. It appeared easy to overlook, so Cressey highlighted it in his illustrations (Cressey 1991: arrowed in his Fig. 64). In the original description, Brian (1914) figured the female leg 4 as having two large distal spines on the oblique distal margin, plus a tiny spine that was largely hidden by its associated pecten (Brian 1914: Fig. 5). In the new species, this outermost spine on the oblique distal margin is well developed and about equal in size to the spine on the lateral margin of the same segment. Given the diagnostic importance of leg 4 in the systematics of Caligus, this is a significant difference between the new species and both Brian’s eastern Atlantic (Madeira) females and Cressey’s Gulf of Mexico (Belize) females of C. biaculeatus, which share the possession of the tiny outer distal spine on leg 4. The lengths of the armature elements on leg 4 are reliable characters in Caligus species, showing little if any geographic variation, unlike the shape of the female genital complex which varies markedly with the reproductive state of the individual female (Parker et al. 1968). An additional minor difference is that the tines of the sternal furca are relatively longer in C. biaculeatus than in the new species.
The new species also resembles Caligus suffuscus Wilson, 1913. However, the new species differs from C. suffuscus in the possession of a blunt-tipped posterior dentiform process on the maxiilule (vs. slender process tapering towards the tip); the sternal furca has divergent and inwardly curved tines (vs. divergent but straight tines); the outer spine on the distal exopodal segment of leg 1 is slightly shorter than the adjacent elements (vs. distinctly shorter than other three distal elements); the middle endopodal segment of leg 2 has fine setules along the outer margin (vs. with a row of spinules) and the outer spine on the distal exopodal segment of leg 4 is more than half as long as the middle spine (vs. distinctly shorter).
Although closely related to C. biaculeatus, we consider that the new species described herein exhibits sufficient differences to justify the establishment of a new species to accommodate it. It can be most readily distinguished from C. biaculeatus by the size of the outermost distal spine on leg 4, which is equal in length to the lateral spine on the same segment.
References
Boxshall GA (1990) The skeletomusculature of siphonostomatoid copepods, with an analysis of adaptive radiation in structure of the oral cone. Philos Trans R Soc Lond B 328:167–212. https://doi.org/10.1098/rstb.1990.0113
Boxshall GA (1998) Host specificity in copepod parasites of deep-sea fishes. J Mar Syst 15:215–223. https://doi.org/10.1016/S0924-7963(97)00058-4
Boxshall GA (2018) The sea lice (Copepoda: Caligidae) of Moreton Bay (Queensland, Australia), with descriptions of thirteen new species. Zootaxa 4398(1):172
Boxshall GA, El-Rashidy HH (2009) A review of the Caligus productus species group, with the description of a new species, new synonymies and supplementary descriptions. Zootaxa 2271:1–26
Brian A (1914) Copépodes parasites provenant des récentes campagnes scientifiques de S.A.S. le Prince Albert Ier de Monaco ou déposés dans les collections du Musée océanographie. Bull. Inst. Oceanogr Monaco 286:1–14
Cordero del Campillo, M. (1978) Indice-catálogo de zooparásitos ibéricos. 5. Acantocefalos. 6. Anelidos. 7. Crustaceos. 8. Malofagos. Universidad de Oviedo, León, Spain 1–65, map. (Crustaceos: 29–36). (Addition)
Cressey RF (1991) Parasitic copepods from the Gulf of Mexico and Caribbean Sea, III: Caligus. Contrib Zool (497):1–53 (including appendix), figs 1–220
Demirkale I, Özak AA, Yanar A, Boxshall GA (2014) Caligus solea n. sp. (Copepoda: Caligidae) parasitic on common sole, Solea solea (Linnaeus) from the North-Eastern Mediterranean off the Turkish coast. Syst Parasitol 89:23–32. https://doi.org/10.1007/s11230-014-9505-4
Essafi KP, Raibaut A, Boudaoud-Krissat K (1983) Colobomatus steenstrupi (Richiardi, 1876) and Colobomatus mulli n. sp. (Copepoda: Philichthyidae), parasitic on fish of the genus Mullus (Mullidae) in the western Mediterranean. Syst Parasitol 5:135–142. https://doi.org/10.1007/BF00049240
Froese R., Pauly D. (Eds) (2018) FishBase. World Wide Web electronic publication. http://www.fish base.org. Accessed 10 May 2018
Huys R, Boxshall GA (1991) Copepod evolution. The Ray Society, London, p 468
Kabata Z. (2003) Copepods parasitic on fishes. Synopses of the British Fauna No. 47, 2nd revised edn. Backhuys No. 4: 1-274, 288 figs
Nuñes-Ruivo L (1954) Parasites de poissons de mer ouest-africains récoltés par M. XJ. Cadenat. III. Copépodes (2e note). Genres Prohatschekia n. gen. et Hatschekia Poche. Bull Inst Fr Afr Noire Sér A 16:479–505
Özak AA, Demirkale I, Yanar A (2012) First record of two species of parasitic copepods on immigrant pufferfishes (Tetraodontiformes: Tetraodontidae) caught in the eastern Mediterranean Sea. Turk J Fish Aquat Sci 12:675–681. https://doi.org/10.4194/1303-2712-v12_3_16
Özak AA, El-Rashidy HH, Demirkale I, Boxshall GA (2010) The discovery of Caligus temnodontis Brian, 1924 (Copepoda: Caligidae) from the bluefish Pomatomus saltatrix (Linnaeus) in the eastern Mediterranean Sea. Syst Parasitol 76(3):223–230. https://doi.org/10.1007/s11230-010-9253-z
Özak AA, Yanar A, Boxshall GA (2017) The discovery of Caligus macrurus Heller, 1865 (Copepoda: Caligidae) in the Mediterranean Sea, and the recognition of Sciaenophilus van Beneden, 1852 as a junior synonym of Caligus Müller, 1785. Syst Parasitol 94:97–109. https://doi.org/10.1007/s11230-016-9682-4
Papoutsoglou SE (1976) Metazoan parasites of fishes from Sacronicos gulf Athens-Greece. Thalassografica 1:62–102
Parker RR, Kabata Z, Margolis L, Dean MD (1968) A review and description of Caligus curtus Müller, 1785 (Caligidae: Copepoda), type species of its genus. J Fish Res Board Can 25:1923–1969
Radujkovic BM, Raibaut A (1987) Copépodes parasites des poissons des côtes du Monténégro (Adriatique sud). Première série. Acta Adriat 28(1–2):121–142
Raibaut A, Combes C, Benoit F (1998) Analysis of the parasitic copepod species richness among Mediterranean fish. J Mar Syst 15:185–206. https://doi.org/10.1016/S0924-7963(97)00079-1
Ramdane Z, Trilles JP, Ho J-S (2010) Caligus uranoscopi Vaissière, 1955 (Copepoda, Caligidae), a rare parasitic copepod from Algeria. Acta Parasitol 55(4):379–385. https://doi.org/10.2478/s11686-010-0053-z
Richiardi S. (1880) Contribuzione alla fauna d’Italia. I. Catalogo systematico di crostacei chez vivono sul corpo di animali aquatici. Catalogo degli Espozitioni e della cosa Esposte, Espozitione internationale di Pesca in Berlino. p. 147–152
Wildgoose WH (2001) BSAVA manual of ornamental fish. Wiley, Gloucester
Acknowledgements
We would like to thank local fishermen working in Karataş in Adana, Turkey, for their help during our fish sampling and alsoFlying Sharks team for support and materials provided during the collection and sampling process.
Funding
This research was funded by the Çukurova University academic research projects unit (Project No. FBA-2017-7490).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that there is no conflict of interest.
Ethical approval
All procedures performed in studies involving animals were in accordance with the ethical standards of the University of Cukurova in Adana, Turkey and Flying Sharks, in Horta, Portugal or practice at which the studies were conducted. All applicable international, national and/or institutional guidelines for the use and care of animals were followed. Fish samples studied in Turkey and Portugal were maintained according to the guidelines of the animal facility at Faculty of Fisheries of the Cukurova University in Turkey and the Flying sharks in Portugal, respectively.
Additional information
Section Editor: Sascha L Hallett
Rights and permissions
About this article

Cite this article
Rodrigues, A.M.V., Özak, A.A., Silva, L.M.H. et al. Caligus mulli n. sp. (Copepoda: Caligidae) parasitic on two Mullid fishes from the eastern Mediterranean and adjacent Atlantic waters. Parasitol Res 117, 3843–3850 (2018). https://doi.org/10.1007/s00436-018-6090-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00436-018-6090-5