
Abstract
We examined whether inspiratory muscle training (IMT) improved cycling time-trial performance and changed the relationship between limit work (W lim) and limit time (T lim), which is described by the parameters critical power (CP) and anaerobic work capacity (AWC). Eighteen male cyclists were assigned to either a pressure-threshold IMT or sham hypoxic-training placebo (PLC) group. Prior to and following a 6 week intervention subjects completed a 25-km cycling time-trial and three constant-power tests to establish the W lim–T lim relationship. Constant-power tests were prescribed to elicit exercise intolerance within 3–10 (Ex1), 10–20 (Ex2), and 20–30 (Ex3) min. Maximal inspiratory mouth pressure increased by (mean ± SD) 17.1 ± 12.2% following IMT (P < 0.01) and was accompanied by a 2.66 ± 2.51% improvement in 25-km time-trial performance (P < 0.05); there were no changes following PLC. Constant-power cycling endurance was unchanged following PLC, as was CP (pre vs. post: 249 ± 32 vs. 250 ± 32 W) and AWC (30.7 ± 12.7 vs. 30.1 ± 12.5 kJ). Following IMT Ex1 and Ex3 cycling endurance improved by 18.3 ± 15.1 and 15.3 ± 19.1% (P < 0.05), respectively, CP was unchanged (264 ± 62 vs. 263 ± 61 W), but AWC increased from 24.8 ± 5.6 to 29.0 ± 8.4 kJ (P < 0.05). In conclusion, these data provide novel evidence that improvements in constant-power and cycling time-trial performance following IMT in cyclists may be explained, in part, by an increase in AWC.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
There is now little doubt that the structure and function of the respiratory muscles can be modified through specific training. Respiratory muscle training (RMT) can be performed using voluntary isocapnic hyperpnoea (VIH), flow-resistive loading, and pressure-threshold loading, otherwise commonly referred to as inspiratory muscle training (IMT). All three techniques increase breathing endurance whilst IMT increases diaphragm thickness (Downey et al. 2007), and maximal inspiratory muscle strength, endurance, shortening velocity and power output (for a full review see McConnell and Romer 2004).
Whether RMT has an ergogenic effect remains somewhat more controversial (for reviews see McConnell and Romer 2004; Sheel 2002). Comparisons of the literature are complicated by the potentially variable outcomes with each RMT technique (or concurrent IMT-VIH training (Sonetti et al. 2001)) and by inter-study differences in RMT protocols and durations. In addition, the mode, intensity, duration and type (time-trial or constant-power) of exercise performance evaluation test has differed considerably between studies, which also hinders comparison and interpretation. A further consideration is that some of the literature has been characterised by weak experimental design (McConnell and Romer 2004). A brief synopsis is that VIH training improved 4-mile running time-trial performance (Leddy et al. 2007) and constant-power cycling endurance at moderate (70–85% maximum power or maximal oxygen uptake, \( \dot W_{\max } \) and \(\ifmmode\expandafter\dot\else\expandafter\.\fi{V}{\text{O}}_{{2\max }},\) respectively) (Boutellier et al. 1992; Markov et al. 2001; McMahon et al. 2002), but not high (90–95% \( {\text{ }}{{\dot W_{\max } } \mathord{\left/ {\vphantom {{\dot W_{\max } } {\dot V{\text{O}}_{2\max } }}} \right. \kern-\nulldelimiterspace} {\dot V{\text{O}}_{2\max } }} \)) (Fairbarn et al. 1991; Morgan et al. 1987), exercise intensities. Accordingly, it is suggested that the efficacy of RMT is more apparent when the performance evaluation test is submaximal and prolonged, with performance gains becoming less discernable, although not eradicated, at higher exercise intensities (Leddy et al. 2007; McConnell and Romer 2004). IMT improved constant-power cycling endurance at both high \( \left( {100\% {\text{ }}\dot V{\text{O}}_{2\max } } \right) \) (Edwards and Cooke 2004) and moderate (75–80% \(\ifmmode\expandafter\dot\else\expandafter\.\fi{W}_{{\max }} /\ifmmode\expandafter\dot\else\expandafter\.\fi{V}{\text{O}}_{{2\max }} \))) exercise intensities (Gething et al. 2004; Guenette et al. 2006; Sonetti et al. 2001), although others failed to observe improved cycling and running endurance at 85% \( \dot V{\text{O}}_2 \max \) (Downey et al. 2007; Williams et al. 2002). Improvements of 1.0–4.6% have been observed for 6 min rowing and 20- and 40-km cycling time-trials following IMT in trained athletes (Griffiths and McConnell 2007; Romer et al. 2002a; Volianitis et al. 2001), whereas a 1.8% improvement in 8-km cycling time-trial performance following concurrent IMT and VIH training in trained cyclists failed to surpass that observed in a sham training placebo group (Sonetti et al. 2001). The lack of consensus is thus apparent but may be explained by the principle of training specificity: RMT ergogenicity may be dependent upon the nature of the performance evaluation test and the underlying mechanism(s) of action (McConnell and Romer 2004).
The mechanism(s) by which RMT may improve exercise tolerance has yet to be fully revealed although it has been argued that constant-power and time-trial exercise performances are predominantly governed by the parameters critical power (CP) and anaerobic work capacity (AWC) (Brandon 1995; Bulbulian et al. 1986; Fernández-García et al. 2000; Fukuba and Whipp 1999; Smith et al. 1999). Therefore, if RMT elicits a genuine ergogenic effect one might expect one or both of these parameters to be affected. CP and AWC can be derived from a series of exhaustive constant-power exercise tests in which exercise duration (T lim) is measured (Hill 1993). The power-T lim relationship is hyperbolic but can be linearised by plotting total work performed (W lim) against T lim, with the resulting gradient and y-intercept representing CP and AWC, respectively (Hill 1993; Monod and Scherrer 1965). Theoretically, CP is an inherent component of the aerobic energy supply system that characterises the highest exercise intensity at which a steady-state can be maintained in \( \dot V{\text{O}}_2 , \) blood lactate concentration, and blood acid–base balance (Hill et al. 2002; Poole et al. 1988). Conversely, although the physiological mechanisms of AWC are less clear, it may represent a constant, but finite, energy store that can be utilised when exercise intensity exceeds CP (Morton 2006). The magnitude of AWC may also be determined by fatigue-inducing metabolite accumulation (Fukuba et al. 2003), and/or the capacity to resist the adverse consequences of metabolic perturbations in heavy exercise (Jenkins and Quigley 1993).
The aim of this study was to examine whether IMT improves cycling time-trial performance and results in a corresponding increase in CP and/or AWC. Therefore, we examined the effects of 6 weeks IMT upon 25-km cycling time-trial performance and the W lim–T lim relationship in competitive cyclists.
Methods
Subjects
Following approval from Nottingham Trent University’s ethics committee, 18 non-smoking, competitive male cyclists provided written informed consent to participate in the study. Throughout the study subjects were instructed to adhere to their usual training regimen and not to partake in strenuous exercise the day before test days, during which subjects abstained from ingesting caffeine and arrived at the laboratory at least 2 h post-prandial. For each participant, tests were performed at a similar time of day. Descriptive characteristics of the subjects are presented in Table 1.
Experimental design
Subjects were initially familiarised with the test procedures and subsequently performed pulmonary and inspiratory muscle function tests. Subjects were then randomly, and equally, assigned to either a pressure-threshold IMT group or a sham hypoxic training placebo group. Prior to the intervention subjects completed a simulated 25-km cycling time-trial test and three constant-power cycling tests to establish the W lim–T lim relationship. Both groups then completed the prescribed training regimen for 6 weeks, after which the battery of exercise tests was repeated in random order, starting at least 48 h after the final training session. All tests were completed on separate days and separated by at least 48 h.
Pulmonary function and maximal inspiratory pressure
Pulmonary function was assessed using a pneumotachograph spirometer (Compact II, Vitalograph, Buckinghamshire, UK) previously calibrated using a 1 l syringe. Each test was repeated three times and the highest recorded value was used for subsequent analysis (Cotes 1993). A hand-held mouth pressure meter (P.K. Morgan, Kent, UK) measured maximal inspiratory mouth pressure (MIP) as an index of global inspiratory muscle strength. The mouthpiece assembly incorporated a 1 mm orifice to prevent glottic closure during inspiratory efforts. Manoeuvres were performed in an upright standing posture, were initiated from residual volume, and were sustained for at least 1 s. Repeat measurements separated by 30–60 s were taken until consistent values within 5 cm H2O of each other were produced (Volianitis et al. 2001). The highest recorded value was then used for subsequent analysis.
25-km time-trial performance
Subjects performed a 25-km cycling time-trial on their own racing bicycle, which was mounted on an air-braked ergometry system (Kingcycle, High Wycombe, Buckinghamshire, UK). Use of this system has been described previously (Palmer et al. 1996). Subjects performed a 2 min warm-up at a self-selected intensity and began the test from a rolling start. Subjects were instructed to complete the 25-km as quickly as possible and the only feedback provided during exercise was the elapsed distance. Heart rate was recorded using short-range telemetry (Accurex Plus, Polar, Kempele, Finland). During exercise subjects wore a facemask (model 7940, Hans Rudolph, Kansas City, Missouri) connected to a two-way, non-rebreathing valve (model 2730, Hans Rudolph), and expired air was collected in Douglas bags at 5-, 10-, 15-, and 20-km intervals. Concentrations of oxygen and carbon dioxide were determined by sampling through paramagnetic and infrared transducers, respectively (Series 1440, Servomex, Crowborough, UK), which were calibrated using certified gases (BOC gases, Guilford, UK). Sample volume was determined using a dry gas meter (Harvard, Edenbridge, UK). Minute ventilation \( (\dot V_{\text{E}} ) \) is presented at BTPS, whereas \( \dot V{\text{O}}_2 \) and carbon dioxide production \( (\dot V{\text{CO}}_2 ) \) are presented at STPD. Upon completion of the test, subjects performed a 3 min cool-down at a self-selected intensity. MIP was measured prior to exercise, following the 3 min cool-down, and 15 min thereafter.
W lim–T lim relationship
The W lim–T lim relationship was determined using three separate, square-wave constant-power tests performed to the limit of volitional tolerance on an electromagnetically-braked cycle ergometer (Excalibur Sport, Lode, Groningen, The Netherlands). Power outputs were chosen to elicit exercise intolerance within each of the following time domains: 3–10 (Ex1), 10–20 (Ex2), and 20–30 (Ex3) min. Subjects adopted a spontaneous cycling cadence in order to maximise T lim (Hill 1993), although in practice the experienced cyclists participating in this study adopted their usual working cadence (approximately 80–100 rpm) and scarcely deviated from this throughout the duration of the tests. Exercise was terminated when cycling cadence could not be maintained above 60 rpm. A 3 min cool-down at 60 W was performed following exercise. MIP was assessed prior to exercise, following the 3 min cool-down, and 15 min thereafter. For each subject W lim was plotted against T lim and the slope and y-intercept of this relationship were taken to represent CP and AWC, respectively (e.g. see Bishop and Jenkins 1996; Monod and Scherrer 1965).
Training protocols
IMT was performed using an inspiratory pressure-threshold device (POWERbreathe®, Gaiam, UK). The IMT group performed 30 dynamic inspiratory efforts twice daily for 6 weeks against a pressure-threshold load of ∼50% MIP. Thereafter, subjects periodically increased the load to a level that would permit them to only just complete 30 manoeuvres. Subjects initiated each inspiratory effort from residual volume and strove to maximise tidal volume. To avoid hyperventilation and therefore hypocapnia due to the increased tidal volume, subjects adopted a reduced breathing frequency. This IMT protocol is known to be effective in eliciting an adaptive response (Griffiths and McConnell 2007; McConnell and Lomax 2006; McConnell and Sharpe 2005; Romer et al. 2002a, 2002b; Volianitis et al. 2001). The placebo group used a sham hypoxic trainer for 15 min, 5 days week−1 (Sonetti et al. 2001). The placebo device was identical to that used by the IMT group, except that the resistance spring was removed and the lower chamber was loosely packed with aquarium gravel, which was promoted to the subjects as being oxygen absorbent, thus reducing the oxygen content of inspired air and mimicking altitude exposure. Subjects were instructed to breathe normally through the device and not increase their normal breathing effort. MIP was assessed every 2 weeks during the intervention, at which time the “oxygen absorbent” gravel in the placebo device was also replaced. All subjects completed a training diary throughout the study to record training adherence and whole-body training sessions.
Statistical analyses
Statistical analyses were performed using SPSS for Windows (SPSS Inc., Chicago, IL, USA). Pre- and post-intervention results and group interactions were compared using one-way or two-way ANOVA for repeated measures and Tukey’s HSD post-hoc analysis. Pearson product-moment correlation coefficients were calculated to assess the relationship between selected variables. Statistical significance was set at P < 0.05. Values are presented as mean ± SD.
Results
Each subject’s habitual training regimen remained unchanged during the intervention. Subjects’ training diaries demonstrated excellent training compliance for both IMT (95% adherence) and placebo (97% adherence) groups.
Pulmonary function and maximal inspiratory pressure
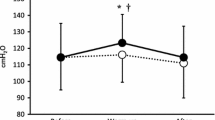
Baseline pulmonary function and MIP were all within normal limits and were not different between IMT and placebo groups (Table 1). Throughout the intervention MIP remained unchanged in the placebo group. Conversely, relative to baseline, MIP increased following 2, 4 and 6 weeks of IMT by 8.4 ± 9.1% (P < 0.05), 10.6 ± 8.3% (P < 0.01), and 17.1 ± 12.2% (P < 0.01), respectively (Fig. 1).

Maximal inspiratory mouth pressure (MIP) at baseline and after 2, 4, and 6 weeks of IMT (filled bars) and placebo (open bars) (mean ± SD). *Significantly different from baseline (P < 0.05). ** Significantly different from baseline (P < 0.01)
Time-trial performance and physiological responses
Group mean changes in time-trial performance are shown in Fig. 2. Time taken to cycle 25-km decreased by 2.66 ± 2.51% following IMT (pre vs. post:- 36.29 ± 3.64 vs. 35.33 ± 3.70 min, P < 0.05) (average power output: 274 ± 66 vs. 290 ± 71 W), whereas no change was observed following placebo (35.72 ± 1.97 vs. 35.98 ± 2.12 min, P = 0.51) (average power output: 275 ± 37 vs. 271 ± 42 W). Changes in time-trial performance were also different between groups, as indicated by a significant group × time interaction effect (P < 0.05). There was a consistent improvement in 5-km split times following IMT with significant group × time interaction effects being observed at 15-, 20-, and 25-km (Fig. 2). Figure 3 shows individual changes in time-trial performance for IMT and placebo subjects. The regression line of pre- vs. post-intervention time-trial performance for the placebo group is similar to the line of identity indicating no change in time-trial performance, whereas that for the IMT group was below, but parallel to, the line of identity. This reflects the improvement in time-trial performance following IMT and also indicates that the improvement was not related to baseline time-trial performance. With the exception of a significant increase in \( \dot V{\text{CO}}_2 \) at 20-km following IMT (3.17 ± 0.73 vs. 3.34 ± 0.86 l min−1), physiological responses to time-trial exercise were not changed at equal distances following IMT or placebo. Prior to the intervention the average \( \dot V_{\text{E}} ,\,\dot V{\text{O}}_2 ,\,\dot V{\text{CO}}_2 ,{\text{ RER}}, \) and heart rate during time-trial exercise in IMT and placebo groups were 126.6 ± 39.9 and 116.8 ± 22.0 l min−1, 3.65 ± 0.86 and 3.51 ± 0.36 l min−1, 3.47 ± 0.82 and 3.33 ± 0.34 l min−1, 0.95 ± 0.03 and 0.95 ± 0.05, and 176 ± 11 and 176 ± 11 beats min−1, respectively. These responses remained unchanged following IMT and placebo. HR measured upon completion of time-trial exercise was also unchanged following IMT (189 ± 15 vs. 187 ± 12 beats min−1) and placebo (189 ± 12 vs. 188 ± 12 beats min−1).

Relative changes in time-trial performance in IMT (filled bars) and placebo (open bars) groups (mean ± SD). *Significant interaction effect (P < 0.05). **Significant interaction effect (P < 0.01)

Pre- and post-intervention time-trial duration for IMT (filled circle) and placebo (open circle) subjects. Regression lines for placebo (solid line) and IMT (dashed line) groups are shown, as is the line of identity
W lim–T lim relationship
Power outputs for Ex1, Ex2, and Ex3 in IMT and placebo groups were 333 ± 74, 286 ± 63, and 281 ± 62 W, and 320 ± 37, 288 ± 28, and 272 ± 29 W, respectively. Constant-power cycling endurance times are shown in Fig. 4. Ex1, Ex2, and Ex3 cycling endurance times were unchanged following placebo. Conversely, although Ex2 endurance time was unchanged following IMT (P = 0.11), endurance times for Ex1 and Ex3 increased by 18.3 ± 15.1 and 15.3 ± 19.1%, respectively (P < 0.05), although group × time interaction effects were not significant (P = 0.08 and 0.11 for Ex1 and Ex3, respectively).

Ex1 (a), Ex2 (b), and Ex3 (c) cycling endurance pre- (filled bars) and post- (open bars) IMT and placebo (mean ± SD). *Significantly different from pre-IMT (P < 0.05)
Changes in CP and AWC are shown in Fig. 5. The relationship between work and cycling endurance time was well described by the W lim–T lim model in both IMT (R 2 = 1.000 ± 0.000 vs. 1.000 ± 0.000) and placebo (R 2 = 0.999 ± 0.002 vs. 0.999 ± 0.002) groups. There was no change in CP following either IMT or placebo, and no change in AWC following placebo. Conversely, AWC increased following IMT (P < 0.05), although a group × time interaction effect was not observed.

Critical power (CP) and anaerobic work capacity (AWC) pre- (filled bars) and post- (open bars) IMT and placebo (mean ± SD). *Significantly different from pre-IMT (P < 0.05)
Exercise-induced changes in maximal inspiratory pressure
Prior to the intervention MIP measured 3 min after time-trial exercise was reduced relative to baseline by 11.8 ± 9.6 and 10.4 ± 7.2% in IMT and placebo groups, respectively (P < 0.01). Following 15 min recovery there was, relative to baseline, a non-significant 6.6 ± 8.8% reduction in MIP in the IMT group, and a 8.2 ± 5.6% reduction (P < 0.05) in the placebo group. Prior to the intervention no decreases in MIP were observed following constant-power exercise except following Ex2 in the IMT group where 7.7 ± 6.0 and 5.9 ± 5.1% decreases (P < 0.05) in MIP were observed 3 and 15 min post-exercise, respectively. When expressed relative to pre-exercise MIP, exercise-induced decrements in MIP were unchanged following IMT and placebo.
Correlations among variables
Relative improvements in cycling time-trial performance following IMT were not correlated with relative IMT-induced changes in MIP or AWC. Increased MIP following IMT was also not correlated with changes in Ex1 and Ex2 endurance times, or increases in AWC. Conversely, although the relative increase in AWC following IMT was not correlated with changes in Ex2 (r = 0.14) or Ex3 (r = −0.42) endurance times, it was correlated with the relative improvement in Ex1 endurance time (r = 0.73, P < 0.05). Pre- and post-intervention values for CP and time-trial performance were also significantly correlated in IMT (r = −0.89 and −0.93, P < 0.01) and placebo (r = −0.69 and −0.75, P < 0.05) groups separately, and when pooled together (r = −0.82 and −0.89, P < 0.01).
Discussion
Main findings
The present study examined the effects of 6 weeks IMT upon 25-km cycling time-trial performance and the W lim–T lim relationship in competitive cyclists. The main finding was that IMT improved cycling time-trial performance to a greater extent than that observed in a sham training placebo group, and that this was accompanied by an increase in AWC.
Inspiratory muscle strength
The 17.1% increase in MIP after 6 weeks IMT is consistent with previous IMT studies (Downey et al. 2007; Edwards and Cooke 2004; Griffiths and McConnell 2007; Hanel and Secher 1991; Inbar et al. 2000; McConnell and Sharpe 2005; Wells et al. 2005), but less than the 30–50% increase reported by others (Gething et al. 2004; Guenette et al. 2006; Huang et al. 2003; Leith and Bradley 1976; Romer and McConnell 2003; Volianitis et al. 2001; Williams et al. 2002). Established training principles appear to apply to IMT (Romer and McConnell 2003), thus these discrepancies may be related, in part, to inter-study differences in IMT mode, intensity, and duration. It is also striking that baseline MIP was high in our subjects (137% predicted, Wilson et al. 1984). The scale of physiological adaptation within a system is dependent upon its baseline status (Åstrand et al. 2003), thus compared to previous studies a smaller window for adaptation may have existed in our subjects. Despite this we observed a rapid increase in MIP of 8.4% following just 2 weeks of IMT. This surprisingly swift pattern of change has been reported elsewhere with ∼14 and ∼28% increases in MIP observed after 1 and 2 weeks of IMT, respectively (Downey et al. 2007; Huang et al. 2003). Such rapid training-induced increases in limb muscle strength are usually ascribed to neural adaptations, with structural alterations becoming evident after about 6–8 weeks of training (Kraemer et al. 1996). However, the signalling pathways that elicit structural alterations in inspiratory muscles might arise earlier than anticipated because of the greater training frequency with IMT (up to 14 sessions week−1) (Kraemer et al. 1996). In support, diaphragm thickness and type II fibre size in the external intercostals increased by 8–12 and 21%, respectively, following 4–5 weeks of IMT (Downey et al. 2007; Ramírez-Sarmiento et al. 2002).
Increased MIP per se is, however, unlikely to explain IMT-induced improvements in exercise performance, since these variables do not correlate (present study; Griffiths and McConnell 2007; Guenette et al. 2006). Rather, IMT-induced alterations to the structure and function of the inspiratory muscles probably elicit systemic and perceptual repercussions that are beneficial to performance (McConnell and Romer 2004).
Time-trial performance and physiological responses
In trained cyclists using the Kingcycle variation in cycling time-trial performance lasting ∼30 min is 1.1% (Palmer et al. 1996). The 2.66% improvement in 25-km cycling time-trial performance observed following IMT exceeds half of the variation between individual performances and can thus be considered worthwhile (Hopkins et al. 1999). Similar improvements in 20- and 40-km cycling (3.8–4.6%) (Romer et al. 2002a), and 6-min rowing (1–3.5%) (Griffiths and McConnell 2007; Volianitis et al. 2001), time-trial performances have been reported in trained athletes following IMT, whereas Leddy et al. (2007) observed a 4% improvement in 4-mile running time following VIH training. The use of an identical sham hypoxic trainer to that used by Sonetti et al. (2001), which anecdotal evidence suggests fully deceived our subjects, contests the notion that placebo effects underpin the efficacy of RMT. Sonetti et al. (2001) argue that the sham hypoxic trainer is superior to the placebo regimens used in other IMT studies, which typically involve training with negligible resistance. Our findings thus suggest a genuine ergogenic effect of IMT on time-trial performance in cyclists. The absence of an ergogenic effect reported by Sonetti et al. (2001) may be because their concurrent IMT-VIH training regimen provided conflicting training stimuli, as evidenced by the modest 8% increase in MIP.
Following IMT physiological responses \( (\dot V_{\text{E}} ,{\text{ }}\dot V{\text{O}}_2 ,{\text{ }}\dot V{\text{CO}}_2 ,{\text{ and heart rate}}) \) to time-trial exercise were unchanged despite an increased power output. This is consistent with previous studies showing reduced metabolic and heart rate responses during constant-power exercise following pressure-threshold IMT (Downey et al. 2007; Gething et al. 2004; Griffiths and McConnell 2007; Romer et al. 2002a). The improvement in time-trial performance following IMT may therefore be linked to an ability to sustain a higher intensity of exercise for the same metabolic and cardiovascular demand.
W lim–T lim relationship
The IMT-induced improvement in cycling time-trial performance was not associated with an increase in CP. This finding is supported by those of McConnell and Sharpe (2005) who observed no change in maximal lactate steady-state cycling power following a similar IMT regimen. Whilst maximal lactate steady-state and CP share similar definitions intra-subject comparisons suggest that CP occurs at a slightly higher power output (Pringle and Jones 2002). Notwithstanding this it appears that adaptations resulting from IMT do not lead to an increase in maximum sustainable power output. It appears more likely that IMT-mediated increases in time-trial performance are linked, in part, to the observed 12% increase in AWC. Unlike CP, \( \dot V{\text{O}}_{2\max } , \) and ventilatory/lactate threshold, AWC is not correlated with time-trial performance (Bulbulian et al. 1986; Smith et al. 1999). Furthermore, we found no correlation between IMT-induced increases in AWC and improvements in 25-km cycling time-trial performance. Conversely, IMT-mediated increases in AWC were correlated with increases in Ex1 cycling endurance. This is perhaps unsurprising given that AWC is related to the ability to perform high-intensity exercise with a large anaerobic component (Hill 1993; Jenkins and Quigley 1993). However, although considered a limiting factor (Brandon 1995; Bulbulian et al. 1986; Fernández-García et al. 2000; Fukuba and Whipp 1999) the contribution of AWC to more prolonged endurance exercise performance is difficult to assess (Brandon 1995). This is reflected by the lack of correlation between IMT-induced increases in AWC and Ex2 and Ex3 cycling endurance. During cycling time-trial exercise lasting ∼30 min power output fluctuates above and below CP, thus AWC is utilised periodically (Tucker et al. 2006). Presumably therefore, the IMT-mediated increase in AWC would have allowed the cyclists to perform a greater volume of work above CP, although the 4.2 kJ increase in AWC alone cannot explain the 16 W increase in mean time-trial cycling power output. Other adaptations, for example perceptual changes, may have influenced the intrinsic system control mechanisms that regulate power output (c.f. Tucker et al. 2006) thereby reducing the volume of work performed below CP and increasing the time spent exercising at, or close to CP.
Our baseline AWC was broadly similar to that reported by others (Bishop and Jenkins 1996), although these values underestimate the true AWC because the W lim–T lim model does not account for the temporal lag in oxidative ATP synthesis at the onset of square-wave exercise (Morton 2006). Previous studies have also shown that strength and high-intensity interval training increase AWC independently of CP (Bishop and Jenkins 1996; Jenkins and Quigley 1993). AWC is thought to reflect finite intramuscular energy stores comprising a phosphagen pool, an anaerobic glycolytic component, and an oxygen store (Morton 2006). Increases in AWC with high-intensity training have thus been attributed to upregulation of phosphofructokinase activity and/or increased buffering capacity within the locomotor muscles (Jenkins and Quigley 1993). It is highly unlikely that IMT would stimulate such adaptations (within the locomotor muscles), thus other mechanisms must explain the increase in AWC. An attractive hypothesis resides in the potential effects of exercise-induced diaphragm fatigue on exercise performance. High-intensity exercise causes diaphragm fatigue, which may reduce limb vascular conductance via a metaboreflex and exacerbate locomotor muscle fatigue (for a review see Dempsey et al. 2006). Since IMT attenuates this metaboreflex (McConnell and Lomax 2006), the increase in AWC may be explained, in part, by improved perfusion of, and oxygen transport to, locomotor muscles and a subsequent reduction in the accumulation of fatigue-inducing intramuscular metabolites. Unfortunately, exercise-induced changes in MIP before and after IMT did not support this hypothesis. However, inferences regarding peripheral diaphragm fatigue cannot be made from MIP measures, which are unable to discriminate either fatigue in different inspiratory muscles, or peripheral and central components of fatigue. Therefore, our data do not exclude the possibility that exercise-induced diaphragm fatigue was attenuated following IMT, as was recently reported following VIH training (Verges et al. 2007). It is also possible that following IMT the same absolute decrease in MIP imparts a smaller influence on limb muscle endurance (McConnell and Lomax 2006).
Ventilatory work during heavy endurance exercise may contribute to the accumulation of metabolites (Johnson et al. 2006) that exacerbate respiratory and locomotor muscle fatigue. One of the most consistent observations following both IMT (Edwards and Cooke 2004; Griffiths and McConnell 2007; McConnell and Sharpe 2005; Romer et al. 2002b; Volianitis et al. 2001) and VIH training (Kohl et al. 1997; Leddy et al. 2007; Spengler et al. 1999; Verges et al. 2007) is a reduction in blood lactate concentration. Although the origin of this reduction remains unknown, it may reflect favourable changes in acid-base balance and/or a delay in the accumulation of fatigue-inducing metabolites, which may also partly explain the increase in AWC following IMT.
It is well known that even after exhaustive exercise locomotor muscle function is preserved and intramuscular glycogen stores are only partially depleted (Morton 2006). It follows that exercise tolerance (and presumably the capacity to deplete AWC) is not exclusively determined by physiological factors; it is likely that perceived effort and feelings of discomfort also play a significant role. Indeed, perhaps the most consistent feature of humans at the limit of exercise tolerance is the reporting of maximal ratings of perceived exertion (Noakes 2004). IMT attenuates ratings of limb discomfort and dyspnoea during exercise (McConnell and Romer 2004) and this may provide a mechanism by which AWC can be more fully exploited.
In summary, IMT-induced improvements in 25-km cycling time-trial performance in competitive cyclists are not explained by an increase in CP, but might be explained, in part, by an increase in AWC. Mechanisms underpinning an IMT-mediated increase in AWC remain unknown, but might be partly related to a reduction/delay in the accumulation of fatigue-inducing metabolites. That IMT resulted in comparable improvements in Ex1 (+18%) and Ex3 (+15%) cycling endurance disputes the notion that the efficacy of RMT is inversely proportional to the intensity of the performance evaluation test (Leddy et al. 2007; McConnell and Romer 2004).
References
Åstrand P-O, Rodahl K, Dahl HA, Stromme SB (2003) Textbook of work physiology: physiological bases of exercise, 4th edn. Human Kinetics, Leeds, pp 313–368
Bishop D, Jenkins DG (1996) The influence of resistance training on the critical power function and time to fatigue at critical power. Aust J Sci Med Sport 28:101–105
Boutellier U, Buchel R, Kundert A, Spengler C (1992) The respiratory system as an exercise limiting factor in normal trained subjects. Eur J Appl Physiol 65:347–353
Brandon LJ (1995) Physiological factors associated with middle distance running performance. Sports Med 19:268–277
Bulbulian R, Wilcox AR, Darabos BL (1986) Anaerobic contribution to distance running performance of trained cross-country athletes. Med Sci Sports Exerc 18:107–113
Cotes JE (1993) Lung function: assessment and application in medicine, 5th edn. Blackwell Scientific Publications, London, pp 496–497
Dempsey JA, Romer L, Rodman J, Miller J, Smith C (2006) Consequences of exercise-induced respiratory muscle work. Respir Physiol Neurobiol 151:242–250
Downey AE, Chenoweth LM, Townsend DK, Ranum JD, Ferguson CS, Harms CA (2007) Effects of inspiratory muscle training on exercise responses in normoxia and hypoxia. Respir Physiol Neurobiol 156:137–146
Edwards AM, Cooke CB (2004) Oxygen uptake kinetics and maximal aerobic power are unaffected by inspiratory muscle training in healthy subjects where time to exhaustion is extended. Eur J Appl Physiol 93:139–144
Fairbarn MS, Coutts KC, Pardy RL, McKenzie DC (1991) Improved respiratory muscle endurance of highly trained cyclists and the effects on maximal exercise performance. Int J Sports Med 12:66–70
Fernández-García B, Pérez-Landaluce J, Rodríguez-Alonso M, Terrados N (2000) Intensity of exercise during road race pro-cycling competition. Med Sci Sports Exerc 32:1002–1006
Fukuba Y, Whipp BJ (1999) A metabolic limit on the ability to make up for lost time in endurance events. J Appl Physiol 87:853–861
Fukuba Y, Miura A, Endo M, Kan A, Yanagawa K, Whipp BJ (2003) The curvature constant parameter of the power-duration curve for varied-power exercise. Med Sci Sports Exerc 35:1413–1418
Gething AD, Williams M, Davies B (2004) Inspiratory resistive loading improves cycling capacity: a placebo controlled trial. Br J Sports Med 38:730–736
Griffiths LA, McConnell AK (2007) The influence of inspiratory and expiratory muscle training upon rowing performance. Eur J Appl Physiol 99:457–466
Guenette JA, Martens AM, Lee AL, Tyler GD, Richards JC, Foster GE, Warburton DER, Sheel AW (2006) Variable effects of respiratory muscle training on cycle exercise performance in men and women. Appl Physiol Nutr Metab 31:159–166
Hanel B, Secher NH (1991) Maximal oxygen uptake and work capacity after inspiratory muscle training: a controlled study. J Sports Sci 9:43–52
Hill DW (1993) The critical power concept. Sports Med 16:237–254
Hill DW, Poole DC, Smith JC (2002) The relationship between power and the time to achieve \( \dot V{\text{O}}_{2\max } . \) Med Sci Sports Exerc 34:709–714
Hopkins WG, Hawley JA, Burke LM (1999) Design and analysis of research on sport performance enhancement. Sports Med 31:472–485
Huang CH, Martin AD, Davenport PW (2003) Effect of inspiratory muscle strength training on inspiratory motor drive and RREP early peak components. J Appl Physiol 94:462–468
Inbar O, Weiner P, Azgad Y, Rotstein A, Weinstein Y (2000) Specific inspiratory muscle training in well-trained endurance athletes. Med Sci Sports Exerc 32:1233–1237
Jenkins DG, Quigley BM (1993) The influence of high-intensity exercise training on the W lim–T lim relationship. Med Sci Sports Exerc 25:275–282
Johnson MA, Sharpe GR, McConnell AK (2006) Maximal voluntary hyperpnoea increases blood lactate concentration during exercise. Eur J Appl Physiol 96:600–608
Kohl J, Koller EA, Brandenberger M, Cardenas M, Boutellier U (1997) Effect of exercise-induced hyperventilation on airway resistance and cycling endurance. Eur J Appl Physiol 75:305–311
Kraemer WJ, Fleck SJ, Evans WJ (1996) Strength and power training: physiological mechanisms of adaptation. Exerc Sport Sci Rev 24:363–397
Leddy JJ, Limprasertkul A, Patel S, Modlich F, Buyea C, Pendergast DR, Lundgren CE (2007) Isocapnic hyperpnea training improves performance in competitive male runners. Eur J Appl Physiol 99:556–676
Leith DE, Bradley M (1976) Ventilatory muscle strength and endurance training. J Appl Physiol 41:508–516
Markov G, Spengler CM, Knopfli-Lenzin C, Stuessi C, Boutellier U (2001) Respiratory muscle training increases cycling endurance without affecting cardiovascular responses to exercise. Eur J Appl Physiol 85:233–239
McConnell AK, Lomax M (2006) The influence of inspiratory muscle work history and specific inspiratory muscle training upon human limb muscle fatigue. J Physiol 577:445–457
McConnell AK, Romer LM (2004) Respiratory muscle training in healthy humans: resolving the controversy. Int J Sports Med 25:284–293
McConnell AK, Sharpe GR (2005) The effect of inspiratory muscle training upon maximum lactate steady-state and blood lactate concentration. Eur J Appl Physiol 94:277–284
McMahon ME, Boutellier U, Smith RM, Spengler CM (2002) Hyperpnoea training attenuates peripheral chemosensitivity and improves cycling endurance. J Exp Biol 205:3937–3943
Monod H, Scherrer J (1965) The work capacity of a synergic muscular group. Ergonomics 8:329–338
Morgan DW, Kohrt WM, Bates BJ, Skinner JS (1987) Effects of respiratory muscle endurance training on ventilatory and endurance performance of moderately trained cyclists. Int J Sports Med 8:88–93
Morton RH (2006) The critical power and related whole-body bioenergetic models. Eur J Appl Physiol 96:339–354
Noakes TD (2004) Linear relationship between the perception of effort and the duration of constant load exercise that remains. J Appl Physiol 96:1571–1572
Palmer GS, Dennis SC, Noakes TD, Hawley JA (1996) Assessment of the reproducibility of performance testing on an air-braked cycle ergometer. Int J Sports Med 17:293–298
Poole DC, Ward SA, Gardner GW, Whipp BJ (1988) Metabolic and respiratory profile of the upper limit for prolonged exercise in man. Ergonomics 31:1265–1279
Pringle JS, Jones AM (2002) Maximal lactate steady state, critical power and EMG during cycling. Eur J Appl Physiol 88:214–226
Ramírez-Sarmiento A, Orozco-Levi M, Güell R, Barreiro E, Hernandez N, Mota S, Sangenis M, Broquetas JM, Casan P, Gea J (2002) Inspiratory muscle training in patients with chronic obstructive pulmonary disease: structural adaptation and physiologic outcomes. Am J Respir Crit Care Med 166:1491–1497
Romer LM, McConnell AK (2003) Specificity and reversibility of inspiratory muscle training. Med Sci Sports Exerc 35:237–244
Romer LM, McConnell AK, Jones DA (2002a) Effects of inspiratory muscle training on time-trial performance in trained cyclists. J Sports Sci 20:547–562
Romer LM, McConnell AK, Jones DA (2002b) Effects of inspiratory muscle training upon recovery time during high-intensity, repetitive sprint activity. Int J Sports Med 23:353–360
Sheel AW (2002) Respiratory muscle training in healthy individuals: physiological rationale and implications for exercise performance. Sports Med 32:567–581
Smith JC, Dangelmaier BS, Hill DW (1999) Critical power is related to cycling time trial performance. Int J Sports Med 20:374–378
Sonetti DA, Wetter TJ, Pegelow DF, Dempsey JA (2001) Effects of respiratory muscle training versus placebo on endurance exercise performance. Respir Physiol 127:185–199
Spengler CM, Roos M, Laube SM, Boutellier U (1999) Decreased exercise blood lactate concentrations after respiratory endurance training in humans. Eur J Appl Physiol 79:299–305
Tucker R, Bester A, Lambert EV, Noakes TD, Vaughan CL, St Clair Gibson A (2006) Non-random fluctuations in power output during self-paced exercise. Br J Sports Med 40:912–917
Verges S, Lenherr O, Haner AC, Schulz C, Spengler CM (2007) Increased fatigue resistance of respiratory muscles during exercise after respiratory muscle endurance training. Am J Physiol Regul Integr Comp Physiol 292:1246–1253
Volianitis S, McConnell AK, Koutedakis Y, McNaughton L, Backx K, Jones DA (2001) Inspiratory muscle training improves rowing performance. Med Sci Sports Exerc 33:803–809
Wells GD, Plyley M, Thomas S, Goodman L, Duffin J (2005) Effects of concurrent inspiratory and expiratory muscle training on respiratory and exercise performance in competitive swimmers. Eur J Appl Physiol 94:527–540
Williams JS, Wongsathikun J, Boon SM, Acevedo EO (2002) Inspiratory muscle training fails to improve endurance capacity in athletes. Med Sci Sports Exerc 34:1194–1198
Wilson SH, Cooke NT, Edwards RHT, Spiro SG (1984) Predicted normal values for maximal respiratory pressures in caucasian adults and children. Thorax 39:535–538
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Johnson, M.A., Sharpe, G.R. & Brown, P.I. Inspiratory muscle training improves cycling time-trial performance and anaerobic work capacity but not critical power. Eur J Appl Physiol 101, 761–770 (2007). https://doi.org/10.1007/s00421-007-0551-3
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00421-007-0551-3