
Abstract
Several studies have reported that improvements in endurance performance following respiratory muscle training (RMT) are associated with a decrease in blood lactate concentration ([Lac]B). The present study examined whether pressure threshold inspiratory muscle training (IMT) elicits an increase in the cycling power output corresponding to the maximum lactate steady state (MLSS). Using a double-blind, placebo-controlled design, 12 healthy, non-endurance-trained male participants were assigned in equal numbers to an experimental (IMT) or sham training control (placebo) group. Cycling power output at MLSS was initially identified using a lactate minimum protocol followed by a series of constant power output rides (2.5% increments) of 29.5 min duration; MLSS was reassessed following six weeks of IMT or sham IMT. Maximum inspiratory mouth pressure increased significantly (26%) in the IMT group, but remained unchanged in the placebo group. The cycling power output corresponding to MLSS remained unchanged in both groups after the intervention. After IMT, [Lac]B decreased significantly at MLSS power in the IMT group [−1.17 (1.01) mmol l−1 after 29.5 min of cycling; mean (SD)], but remained unchanged in the placebo group [+0.37 (1.66) mmol l−1]. These data support previous observations that IMT results in a decrease in [Lac]B at a given intensity of exercise. That such a decrease in [Lac]B was not associated with a substantial (>2.5%) increase in MLSS power is a new finding suggesting that RMT-induced increases in exercise tolerance and reductions in [Lac]B are not ascribable to a substantial increase in the ‘lactate threshold’.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Recent well-controlled studies have demonstrated that specific respiratory muscle training (RMT) prolongs exercise tolerance at fixed intensities of exercise above the lactate threshold (Boutellier and Piwko 1992; Boutellier et al. 1992; Markov et al. 2001), and improves time trial performance (Romer et al. 2002a; Volianitis et al. 2001; see McConnell and Romer (2004) for review). However, the mechanisms that underlie the observed improvements in whole-body performance remain poorly understood. The available evidence indicates that performance improvement after RMT occurs without any accompanying increase in maximal oxygen uptake (Boutellier et al. 1992; Markov et al. 2001; Romer et al. 2002a;). However, a number of studies have reported a significant reduction in the circulating concentration of lactate [Lac]B at equivalent intensities of exercise after RMT (Boutellier and Piwko 1992; Boutellier et al. 1992; Romer et al. 2002b; Spengler et al. 1999). This reduction in [Lac]B has been observed after both inspiratory threshold loading (a strength-training stimulus) and isocapnic hyperpnoea (an endurance-training stimulus). The two most recent studies (Romer et al. 2002b, strength; Spengler et al. 1999, endurance) have demonstrated a significant association between post-RMT changes in blood [Lac]B and performance improvement, such that up to 52% of the variation in performance was accounted for by the changes in [Lac]B (Romer et al. 2002b). However, the authors of neither study implied any causation on the basis of the association. Notwithstanding this, the association suggests that the balance between the appearance and disappearance of lactate, or something related to it, is linked strongly to improved exercise performance following RMT.
An increase in the exercise intensity corresponding to the onset of [Lac]B accumulation would provide a plausible explanation for improved whole body performance post-RMT. Spengler et al. (1999) observed a (qualitative) rightward shift in the relationship between exercise intensity and [Lac]B following 4 weeks of isocapnic hyperpnoea training. Such changes are reminiscent of those that follow a period of whole body exercise training and might explain the ergogenic effect of RMT. However, the protocol employed by Spengler et al. (30-W increments every 2 min) was designed primarily to elicit maximal O2 uptake, rather than the individual lactate threshold of each participant; indeed, the latter was not even reported. In a study examining the relationship between stage duration and work increment size, Stockhausen et al. (1997) observed that, with a 30-W increment, stage duration of at least 4 min was required in order to accurately determine the lactate threshold. Furthermore, Smith and Jones (2001) have argued that if one wishes to obtain a true and precise estimate of the exercise intensity associated with the transition between a raised, but stable [Lac]B, and a [Lac]B that increases continuously with time, the maximum lactate steady state (MLSS) must be derived. The MLSS is defined as ‘the highest blood lactate concentration and work load that can be sustained over time without continual lactate accumulation’ (Billat et al. 2003). The main reason for not using MLSS routinely is that it is extremely time consuming, requiring multiple visits to the laboratory in order to titrate the exercise intensity at which MLSS occurs (testing at each intensity requires a 30-min fixed intensity bout of exercise). Thus, although Spengler et al.’s (1999) data suggest that there may be an increase in the intensity of exercise associated with the onset of [Lac]B accumulation post-RMT (rightward shift of the relationship between exercise intensity and [Lac]B), this issue has not been addressed directly, and certainly not using a methodology that is capable of precisely identifying an RMT-induced improvement in the exercise intensity at which a steady-state of [Lac]B can be maintained, i.e. the MLSS intensity.
Thus, the purpose of the present study was to examine the influence of 6 weeks of pressure threshold inspiratory muscle training (IMT) upon the [Lac]B and cycling power output at the maximum lactate steady state (MLSS). We hypothesise that IMT will (1) decrease [Lac]B at MLSS (confirming previous observations), and (2) increase the power output corresponding to MLSS (providing an explanation for the lowered [Lac]B following RMT).
Methods
Participants
After local ethics committee approval and written informed consent, 12 recreationally active, non-smoking (self-reported) participants volunteered to participate in the study. Descriptive characteristics of the participants are presented in Table 1.
General design
Participants were allocated in equal numbers to either an experimental inspiratory muscle training group (IMT) or a sham training control group (placebo) using a double blind, placebo controlled design. Participants were familiarised thoroughly with all test procedures during their first laboratory visit. All procedures were conducted in accordance with ethical standards of the Committee on Human Experimentation at the host institution and with the Helsinki Declaration of 1975.
Procedure
Pre-test preparation
Participants were asked to refrain from exercise and from consuming alcohol and caffeine for 24 h prior to testing. On a test day participants were asked to report to the laboratory at a similar (±1 h) time of day to minimise any diurnal effects and to refrain from eating for a 2-h period before testing.
Blood sampling and analysis
Arterialised mixed venous blood was sampled from an indwelling 21-G cannula positioned in a dorsal hand vein. Continuous arterialisation was maintained by placing the hand in water at 43°C for 10 min prior to cannulation. During exercise the area around the cannula was heated with an infra-red lamp (skin temperature was kept at around 40°C and was measured using a thermistor). Samples were drawn into a heparinised 1-ml syringe and were shaken vigorously. Whole blood samples were analysed immediately for [Lac]B, partial pressure of CO2 (PCO2), O2 saturation (SaO2) and pH using a Nova Stat Profile 9 (Nova Biomedical, Waltham, Mass., USA). Between samples the cannula was flushed with approximately 2 ml of saline. Residual fluids were drawn into a 5-ml syringe prior to sampling.
Measurement of pulmonary ventilation and gas exchange
Pulmonary ventilation and gas exchange were measured during all constant power cycling tests. Respiratory airflow was measured using an ultrasonic phase-shift flow meter interfaced with the participant via a mouth piece (Birmingham Flowmetrics, Birmingham, UK). Gas concentrations were sampled at the mouth via a catheter approximately 2 m in length and were measured continuously by mass spectrometer (Airspec MGA 2000; Biggin Hill, Kent, UK) with a sampling frequency of 50 Hz. The flow meter and mass spectrometer signals were processed and integrated using dedicated software to give gas exchange and ventilatory parameters (Labview 3; National Instruments, Austin, Tex., USA). All instruments were calibrated immediately prior to each test.
Inspiratory muscle strength
Respiratory muscle strength was assessed using a portable hand-held mouth pressure meter (Precision Medical, Pickering, North Yorks., UK) for the determination of maximum static inspiratory pressure (MIP). Participants performed ten maximal efforts at 30-s intervals from residual volume (Maillard et al. 1998), with the highest value recorded.
Assessment of MLSS
All exercise was performed using an electromagnetically braked cycle ergometer (Excalibur Sport, Lode, Groningen, The Netherlands). A lactate minimum protocol, based on that described by Tegtbur et al. (1993) was used to provide a initial starting point for the estimation of MLSS. The lactate minimum power output derived from this protocol had previously been shown (unpublished results) to be in good agreement with MLSS (lactate minimum power was a mean of 10.5 W lower than MLSS power with limits of agreement of 26 W and −5 W). The required elevation of [Lac]B was achieved using a maximal incremental cycle test in which power output was increased by a constant increment every 15 s. The magnitude of this increment was based upon the subject’s size and training status; using our experience, we estimated the likely peak power output of the subject. This was then used to derive a power output ramp rate that would achieve the predicted peak power output within 10 min. The test was followed by an 8-min period of active recovery (cycling at 60 W), after which, five stages of exercise of 4 min duration were performed each separated by 60 s of recovery, during which a 1-ml blood sample was drawn for immediate analysis. The power outputs for the five stages were based on a percentage of the maximum power output attained during the incremental test (50%, 55%, 60%, 65% and 70%). A second-order polynomial function was fitted to the relationship of [Lac]B and power output. The asymptote of the resulting curve (the lactate minimum) was determined through differentiation of the quadratic equation.
A minimum of two constant power tests of 29.5 min duration were used to derive the MLSS power output (the initial power output being that derived from the lactate minimum test). Blood sampling began at minute 10 and was continued at 1.5-min intervals. When determining MLSS, a linear regression was performed between [Lac]B and time ([Lac]B did not stabilise until minute 13 and this was therefore the first value used during this analysis). The following equation was used to establish whether the gradient of the linear regression was significantly different to zero:
where b is the gradient of the line, Sx is the standard deviation of x (time), and Syx is the standard error of the estimate. If the gradient was statistically significantly different to zero, and positive, power was reduced by 2.5%; if the gradient did not meet these criteria the power was increased by 2.5%. The MLSS was defined as the highest power output at which the gradient of the linear regression was not positive.
After the 6-week period of IMT, participants performed at least two constant intensity exercise tests. The first test was performed at the power output equivalent to the pre-IMT MLSS; if [Lac]B did not increase over time (using the previously described criteria) the power output for the second test was increased by 2.5%; if [Lac]B did increase with time, the power output was reduced by 2.5%. This process was repeated until the power output equivalent to MLSS was identified.
Inspiratory muscle training
The principle investigators were blinded to the training condition of each participant. The IMT group performed 30 dynamic inspiratory efforts twice daily for 6 weeks against a pressure-threshold load equivalent to 50% of MIP, a protocol known to be effective in eliciting an adaptive response (Romer and McConnell 2003; Romer et al. 2002a, 2002b; Volianitis et al. 2001). The placebo group trained using 60 slow protracted breaths once daily for 6 weeks at 15% MIP, a protocol known to induce minimal changes in inspiratory muscle function (Romer and McConnell, 2003; Romer et al. 2002a, 2002b, Volianitis et al. 2001). Loading characteristics of the IMT device (POWERbreathe; Leisure Systems International, Southam, UK) have been documented previously (Caine and McConnell 2000). Following the initial setting of training loads, participants in the IMT group were instructed by an independent observer to periodically increase the load to a level that would permit them to only just complete 30 manoeuvres; the placebo group were not given these instructions. All participants were told they were participating in a study to compare the influence of strength (IMT group) versus endurance (placebo group) protocols and, as a consequence, were blinded to the true purpose of the study and the expected outcomes. Participants were instructed to cease training 48 h prior to post-IMT trials. All participants completed IMT diaries throughout the study.
Data analyses
Repeated measures ANOVA was used to test for between group effects due to “treatment” (IMT or placebo) and within group effects due to “time” (pre- versus post-intervention) on each of the dependent variables. Mauchly’s sphericity test was used to check for homogeneity of variance and homogeneity of covariance. Violations of the assumption of sphericity were corrected using the Greenhouse-Geisser adjustment. Planned pairwise comparisons were made using paired t-tests.
An alpha level of 0.05 was chosen a priori to represent statistical difference. All statistical analyses were performed using SPSS v10.0 for Windows (SPSS, Chicago, Ill., USA). All data are presented as mean (SD).
Results
Maximum inspiratory pressure
The adherence to the 6-week IMT programme was good; 83.7 (11%) of the prescribed training sessions were performed. There was a significant interaction of ‘treatment’ (IMT or placebo) and ‘time’ (pre- or post-IMT) (P<0.01). Paired t-tests indicated a statistically significant increase in MIP after the intervention for the IMT group (26%; P<0.01), but not the control group (8%; see Table 2).
Maximum lactate steady state
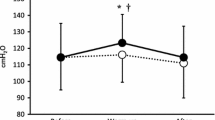
The power output corresponding to MLSS was identical in all participants in the placebo group and in five participants in the IMT group. Power output was reduced by 2.5% for the remaining participant in IMT group in order to achieve MLSS (see Figs. 1, 2).

Placebo group’s maximum lactate steady-state (MLSS) cycling power pre- and post-intervention (n=6)

IMT group’s MLSS cycling power pre- and post-intervention (n=6)
Mean data for blood measures of PCO2, SaO2, pH and [Lac]B are reported in Table 2. There was no statistically significant difference either within or between groups for PCO2, SaO2, or pH. However, for [Lac]B the interaction of ‘treatment’ (IMT or placebo) and ‘time’ (pre- or post-IMT) was only just short of statistical significance (P<0.056). In addition, the effect size for the difference in [Lac]B pre- versus post-IMT was calculated to be 0.96, i.e. a large effect (Cohen 1988). In combination, these observations suggest that there was a genuine ‘treatment’ and ‘time’ interaction, justifying post-hoc t-tests. Paired t-tests indicated a statistically significant reduction in mean [Lac]B (minutes 23.5–29.5; see Table 2) after the intervention for the IMT group (P<0.01), but not the control group.
Figures 3 and 4 depict the [Lac]B whilst cycling at MLSS for each group. The difference in [Lac]B between the pre- and post-IMT tests increased during the latter stages of the test and was largest in the IMT group [pre- to post-difference=1.17 (1.01) mmol l−1 versus +0.37 (1.66) mmol l−1 in the IMT and placebo group, respectively] at the end of the test (29.5 min).

Placebo group mean [Lac]B whilst cycling at MLSS pre- and post-intervention. Error bars=1 SD (n=6)

Pressure threshold inspiratory muscle training (IMT) group mean [Lac]B whilst cycling at MLSS pre- and post-intervention. Error bars=1 SD (n=6)
There were no differences in minute ventilation, O2 uptake, CO2 production, tidal volume, respiratory frequency, duty cycle, or peak or mean inspiratory flow rate in either group post-intervention (Table 3).
Discussion
Our data confirm previous observations that IMT reduces the [Lac]B during sub-maximal constant intensity exercise (Boutellier and Piwko 1992; Boutellier et al. 1992; Romer et al. 2002b; Spengler et al. 1999). We had originally hypothesised that this might be due to an increase MLSS power, but this did not change post-IMT. Previous studies have indicated that there is a correlation between the magnitude of RMT-induced reduction in [Lac]B and the magnitude of improvements in exercise tolerance (Romer et al. 2002b; Spengler et al. 1999). This might imply a direct ‘cause and effect’ relationship between the ‘lactate threshold’ and exercise tolerance. However, our observation that there was no substantial change in MLSS power (>2.5%) does not support this.
Our statistical treatment of the [Lac]B data requires justification. The P value of the interaction effect of treatment (IMT versus placebo) and time (pre- versus post-IMT) upon the [Lac]B during post-IMT exercise at MLSS power was 0.056, which exceeds the a priori alpha level of 0.05. However, it was accompanied by an effect size of 0.96. According to Cohen (1988), effect sizes >0.8 are considered ‘large’. Furthermore, the lower [Lac]B post-RMT is consistent with those of previous studies (Boutellier and Piwko 1992; Boutellier et al. 1992; Romer et al. 2002b; Spengler et al. 1999), and the within group effect was highly significant in the IMT group (P<0.01). Thus, we believe that we would be committing a type 2 error if we were to consider the change in [Lac]B in the IMT group to be non-significant. Accordingly, we are confident that the reduction in [Lac]B that we observed in the IMT group was genuine.
At first sight, our observation that IMT does not increase the power output at MLSS would appear contrary to the findings of Spengler et al. (1999). These authors observed a (qualitative) rightward shift in the relationship of [Lac]B and power output following RMT, a change that is normally suggestive of an increase in the ‘lactate threshold’. For reasons stated in our introduction, the use of incremental protocols to derive the exercise intensity associated with [Lac]B accumulation represents a compromise between validity and practicality. Accordingly, we believe that the data from the present study have greater validity than those of Spengler et al. (1999), who did not, after all, set out to examine the issue of ‘lactate threshold’ or MLSS power specifically.
The power output corresponding to MLSS has been shown to be sensitive to the effects of endurance training, but few studies have focused on the value of [Lac]B at this exercise intensity (see Billat et al. 2003 for a review). Only one study has examined the effect of training on this value. Billat et al. (2004) demonstrated that endurance training improved running velocity at MLSS by just 3.4%, but increased time to the limit of tolerance (Tlim) at MLSS from 44 to 63 min (50%). In contrast to our own observations, Billat et al. (2004) noted no change in the [Lac]B at the running velocity corresponding to MLSS. Methodological differences between Billat et al.’s study and our own mean that the data should be interpreted with caution. For example, the method of deriving the MLSS, [Lac]B at MLSS and the training stimulus (whole body endurance training) all differed. It is also important to note that Billat et al.’s subjects showed a small, but significant increase in maximal oxygen uptake (3%) post-intervention. If confirmed, Billat et al.’s (2004) data appear to suggest that impressive improvements in Tlim could be achieved with comparatively small improvements in MLSS power (around 3%). In our study, the resolution of the MLSS power was ±2.5% (~6 W), and the MLSS power was identical in all subjects but one, pre- and post-intervention. Thus, although we are confident that we could have detected a change in MLSS power >2.5%, we cannot exclude the possibility that a change of <2.5% arose as a result of the IMT. Since Billat et al.’s (2004) recent study tentatively suggests that changes in the exercise intensity corresponding to MLSS as small as 3.4% may result in substantial improvements in Tlim, the possibility remains that a small shift in MLSS may contribute to RMT-induced improvements in performance. However, there is currently a dearth of information on the interrelationship of MLSS power improvement and enhanced exercise performance (Billat et al. 2003), so the functional relevance of small changes in MLSS power remains unknown. Our data indicate that future studies of the effect of IMT upon MLSS power must be designed to detect changes of <2.5%.
Increases in the exercise intensity corresponding to MLSS, and indeed rightward shifts in the lactate threshold, following whole body endurance training have been attributed to training-induced adaptations occurring within locomotor muscles. These include increases in mitochondrial density, oxidative enzyme activity and capillarisation (Holloszy and Coyle 1984). Similar adaptations have been observed in the inspiratory muscles of animal models after whole body endurance training (Bigard et al. 1992; Powers et al. 1992a) and high intensity interval training (Powers et al. 1992b). In contrast to the relative abundance of data on the effects of whole body training upon the metabolic properties of inspiratory muscle, to our knowledge, there are no data available regarding the nfluence of specific RMT upon respiratory muscle oxidative capacity, in either animals or human beings. Animal studies suggest that the diaphragm shows metabolic adaptations to whole body training that are similar to those observed in locomotor muscles, albeit smaller in magnitude (Powers et al. 1995). This being the case, it is reasonable to suppose that the inspiratory muscles also show metabolic adaptations to specific resistance training that are similar to locomotor muscles. Indeed, this similarity has recently been confirmed from a functional (force/velocity) perspective (Romer and McConnell 2003).
In a study of the influence of limb muscle strength training upon whole-body exercise performance, Marcinik et al. (1991) observed both an increase in time to the limit of tolerance at 75% V̇O2 max, and a lowering of [Lac]B at equivalent cycling intensities following training. They also noted a 12% increase in the lactate threshold using an incremental protocol. Other studies (Hoff et al. 1999) have also observed improvements in time to the limit of tolerance after strength training (Hoff et al. 1999; Hickson et al. 1988), but they have failed to replicate Marcinik et al.’s (1991) finding of a change in the lactate threshold. These studies also reported reductions in the [Lac]B at end exercise (limit of tolerance). Under conditions where time to the limit of tolerance has increased, it is reasonable to suppose that isotime measurements of [Lac]B during exercise would show at least equivalent reductions (10–23%) to those seen at end exercise. These data therefore suggest that there may be a reduction in [Lac]B during exercise after limb muscle strength training and that this is accompanied by an increase in the time to the limit of tolerance during whole-body exercise.
The mechanism for the improvements in exercise performance after limb strength training remains unknown, but does not appear to be secondary to changes in muscle enzyme activities; with the exception of lactate dehydrogenase (Tesch et al. 1987), muscle enzyme activities have been shown to decrease following strength training (Tesch et al. 1990). The data from limb muscle strength training studies are consistent with the findings of the present study (with the exception of Marcinik et al.’s (1991) finding of a change in the lactate threshold, which appears to be unique to their subjects), as well as those from other RMT studies that have observed improved whole body exercise performance and lowered [Lac]B after RMT (Boutellier and Piwko 1992; Boutellier et al 1992; Romer et al. 2002b; Spengler et al. 1999).
The IMT-induced lowering of [Lac]B is difficult to account for, but possible explanations have been put forward previously. Spengler et al. (1999) suggested that it was due to an improvement in the inspiratory muscles’ ability to metabolise lactate. Elsewhere (Wetter and Dempsey 2000), it has been suggested that training-induced adaptations of the inspiratory muscles may affect [Lac]B either through increased rates of lactate clearance, and/or decreased rates of production. Evidence from animal models supports the notion that the inspiratory muscles may be net consumers of lactate during exercise (Fregosi and Dempsey 1986, Manohar and Hassan 1990, 1991), and that the lactate dehydrogenase activity of the rat diaphragm increases following endurance training (Bigard et al. 1992). However, our training stimulus was strength orientated, which has not been shown to increase muscle enzyme activities in limb muscles (Tesch et al. 1987, 1990), so alternative explanations must be sought.
There is emerging evidence that membrane bound monocarboylate transporters (MCT) of lactate are increased in response to training (Dubouchaud et al. 2000). It has been argued that isoforms MCT1 and MCT4 facilitate the exchange of lactate between cells (the ‘lactate shuttle’), and that MCT1 is also involved in intracellular shuttling of lactate (Dubouchaud et al. 2000). The majority of studies that have examined the influence of training upon the muscle content of MCTs have examined the influence of endurance training. However, one study has examined the influence of isolated knee-extensor training (Pilegaard et al. 1999). After training, maximum voluntary isometric force was 16% higher in the subjects’ trained leg than in the untrained, control leg. Peak and mean power output, and an index of leg endurance were also highest in the trained leg. In concert with these functional changes was an increase in sarcolemmal lactate/hydrogen ion transport capacity and an increased content of MCT1 (70%) and MCT4 (33%) protein in the trained muscle. The increase in MCT content was also accompanied by other changes that were consistent with a training-induced improvement in sarcolemmal lactate/hydrogen ion transport capacity. This was the first study to demonstrate that isolated, high intensity resistance-type training could induce improvement in lactate/hydrogen ion transport capacity. The training stimulus in Pilegaard et al.’s study (1999) is not dissimilar to that used in the present study. Thus, it is possible that IMT may generate a training stimulus that induces increases in the inspiratory muscle content of MCT1 and MCT4.
But how might increasing the inspiratory muscle content of MCT1 and MCT4 result in a lower circulating [Lac]B? Short-term training models (7 days) have been used to differentiate effects due to muscle fibre adaptations from those due to changes in the number or activity of transporters. Using this approach it has been shown that MCT1 content increases, rate of lactate appearance in the blood is unchanged, and that [La]B and muscle lactate concentration are lower (Phillips et al. 1995; Bonen et al. 1998). The MCT1 and MCT4 transporters can shuttle lactate into, as well as out of cells; thus, in a muscle such as the diaphragm that has been suggested to be a net consumer of lactate during exercise (Fregosi and Dempsey 1986; Manohar and Hassan 1990, 1991), it is possible that an increase in MCT1 and MCT4 content improves the transport of lactate from the circulation, lowering circulating [Lac]B. Although the diaphragm represents a relatively small mass of muscle (around 0.5% of total body mass, according to Wetter and Dempsey 2000), it must be remembered that the accessory muscles of respiration also make a substantial contribution to exercise ventilation. In addition, the high oxidative capacity of the inspiratory muscles must also be borne in mind; there is evidence that respiratory muscles are capable of demanding as much as 16% of available cardiac output during strenuous exercise (Harms and Dempsey 1999). This suggests that their high capillarity and oxidative capacity may create a greater metabolic potential than their relatively small mass might predict.
In summary, our data support previous evidence that at a given intensity of exercise (in this study, MLSS power), circulating [Lac]B is lowered following IMT. Our data are sufficient to exclude as a mediator of the ergogenic effect of RMT, any substantial increases in the power output corresponding to MLSS, but our design cannot exclude the possibility that an increase in MLSS power of less that 2.5% occurred after IMT. The functional relevance of such small changes in MLSS power remains untested, as there is currently a lack of information on the interrelationship of MLSS power improvement and enhanced exercise performance (Billat et al. 2003). Nonetheless, our data suggest that future studies of the effect of IMT upon MLSS power will require designs that are capable of discriminating very subtle changes in MLSS power. The mechanisms underlying the IMT-induced reduction in [Lac]B at MLSS power requires further study, and the role of inspiratory muscle MCTs may provide an interesting avenue for this.
References
Bigard AX, Brunet A, Serrurier B, Guezennec CY, Monod H (1992) Effects of endurance training at high-altitude on diaphragm muscle properties. Pflugers Arch 422:239–244
Billat VL, Sirvent P, Py G, Koralsztein JP, Mercier J (2003) The concept of maximal lactate steady state: a bridge between biochemistry, physiology and sport science. Sports Med 33:407–426
Billat V, Sirvent P, Lepretre PM, Koralsztein JP (2004) Training effect on performance, substrate balance and blood lactate concentration at maximal lactate steady state in master endurance-runners. Pflugers Arch 447:875–883
Bonen A, McCullagh KJ, Putman CT, Hultman E, Jones NL, Heigenhauser GJ (1998) Short-term training increases human muscle MCT1 and femoral venous lactate in relation to muscle lactate. Am J Physiol 274:E102–107
Boutellier U, Piwko P (1992) The respiratory system as an exercise limiting factor in normal sedentary subjects. Eur J Appl Physiol 64:145–152
Boutellier U, Buchel R, Kundert A, Spengler C (1992) The respiratory system as an exercise limiting factor in normal trained subjects. Eur J Appl Physiol 65:347–353
Caine MP, McConnell AK (2000) Development and evaluation of a pressure threshold inspiratory muscle trainer for use in the context of sports performance. J Sports Eng 3:149–159
Cohen J (1988) Statistical power analysis for the behavioural sciences. Academic Press, New York
Dubouchaud H, Butterfield GE, Wolfel EE, Bergman BC, Brooks GA (2000) Endurance training, expression, and physiology of LDH, MCT1, and MCT4 in human skeletal muscle. Am J Physiol 278:E571–579
Fregosi RF, Dempsey JA (1986) Effects of exercise in normoxia and acute hypoxia on respiratory muscle metabolites. J Appl Physiol 60:1274–1283
Harms CA, Dempsey JA (1999) Cardiovascular consequences of exercise hyperpnea. Exerc Sport Sci Rev 27:37–62
Hickson RC, Dvorak BA, Gorostiaga EM, Kurowski TT, Foster C (1988) Potential for strength and endurance training to amplify endurance performance. J Appl Physiol 65:2285–2290
Hoff J, Helgerud J, Wisloff U (1999) Maximal strength training improves work economy in trained female cross-country skiers. Med Sci Sports Exerc 31:870–877
Holloszy JO, Coyle EF (1984) Adaptations of skeletal muscle to endurance exercise and their metabolic consequences. J Appl Physiol 56:831–838
Maillard JO, Burdet L, Melle G van, Fitting JW (1998) Reproducibility of twitch mouth pressure, sniff nasal inspiratory pressure, and maximal inspiratory pressure. Eur Respir J 11:901–905
Marcinik EJ, Potts J, Schlabach G, Will S, Dawson P, Hurley BF (1991) Effects of strength training on lactate threshold and endurance performance. Med Sci Sports Exerc 23:739–743
Markov G, Spengler CM, Knopfli-Lenzin C, Stuessi C, Boutellier U (2001) Respiratory muscle training increases cycling endurance without affecting cardiovascular responses to exercise. Eur J Appl Physiol 85:233–239
Manohar M, Hassan AS (1990) Diaphragm does not produce ammonia or lactate during high-intensity short-term exercise. Am J Physiol 259:H1185–H1189
Manohar M, Hassan AS (1991) Diaphragmatic energetics during prolonged exhaustive exercise. Am Rev Respir Dis 144:415–418
McConnell AK, Romer LM (2004) Respiratory muscle training in healthy humans: resolving the controversy. Int J Sports Med 25:284–93
Phillips SM, Green HJ, Tarnopolsky MA, Grant SM (1995) Increased clearance of lactate after short-term training in men. J Appl Physiol 79:1862–1869
Pilegaard H, Domino K, Noland T, Juel C, Hellsten Y, Halestrap AP, Bangsbo J (1999) Effect of high-intensity exercise training on lactate/H+ transport capacity in human skeletal muscle. Am J Physiol 276:E255–261
Powers SK, Criswell D, Lieu F-K, Dodd S, Silverman H (1992a) Diaphragmatic fibre type specific adaptation. Respir Physiol 89:195–207
Powers SK, Grinton S, Lawler J, Criswell D, Dodd S (1992b) High intensity exercise training-induced metabolic alterations in respiratory muscles. Respir Physiol 89:169–177
Powers SK, Wade M, Criswell D, Herb RA, Dodd S, Hussain R, Martin D (1995) Role of beta-adrenergic mechanisms in exercise training-induced metabolic changes in respiratory and locomotor muscle. Int J Sports Med16:13–18
Romer LM, McConnell AK (2003) Specificity and reversibility of inspiratory muscle adaptations following inspiratory muscle training. Med Sci Sports Exerc 35:237–244
Romer LM, McConnell AK, Jones DA (2002a) Effects of inspiratory muscle training on time trial performance in trained cyclists. J Sports Sci 20:547–562
Romer LM, McConnell AK, Jones DA (2002b) Effects of inspiratory muscle training upon recovery time during high intensity, repetitive sprint activity. Int J Sports Med 23:353–360
Smith CG, Jones AM (2001) The relationship between critical velocity, maximal lactate steady-state velocity and lactate turnpoint velocity in runners. Eur J Appl Physiol 85:19–26
Spengler CM, Roos M, Laube SM, Boutellier U (1999) Decreased exercise blood lactate concentrations after respiratory endurance training in humans. Eur J Appl Physiol 79:299–305
Stockhausen W, Grathwohl D, Burklin C, Spranz P, Keul J (1997) Stage duration and increase of work load in incremental testing on a cycle ergometer. Eur J Appl Physiol Occup Physiol 76:295–301
Tegtbur U, Busse MW, Braumann KM (1993) Estimation of an individual equilibrium between lactate production and catabolism during exercise. Med Sci Sports Exerc 25:620–627
Tesch PA, Komi PV, Hakkinen K (1987) Enzymatic adaptations consequent to long-term strength training. Int J Sports Med 8 [Suppl 1]:66–69
Tesch PA, Thorsson A, Colliander EB (1990) Effects of eccentric and concentric resistance training on skeletal muscle substrates, enzyme activities and capillary supply. Acta Physiol Scand 140:575–580
Volianitis S, McConnell AK, Koutedakis Y, McNaughton L, Backx K, Jones DA (2001) Inspiratory muscle training improves rowing performance. Med Sci Sports Exerc 33:803–809
Wetter TJ, Dempsey JA (2000) Pulmonary system and endurance exercise. In: Shephard RJ, Astrand P.-O. (eds) Endurance in sport. Blackwell, Oxford, pp 52–67
Acknowledgements
Alison McConnell declares a beneficial interest in the POWERbreathe inspiratory muscle trainer.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
McConnell, A.K., Sharpe, G.R. The effect of inspiratory muscle training upon maximum lactate steady-state and blood lactate concentration. Eur J Appl Physiol 94, 277–284 (2005). https://doi.org/10.1007/s00421-004-1282-3
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00421-004-1282-3