
Abstract
Atriplex lentiformis is a halophytic plant species used for desalination and phytoremediation. The plant tolerates abiotic constraints, such as salinity, drought, and toxic metals. It is also used as a fodder for domestic animals. It grows luxuriantly at 100–400-mM NaCl concentrations without any toxic symptoms. In the present investigation effects of biological amendments—PGPB (Plant growth-promoting bacteria—Bradyrhizobium japonicum—NCIM5350 and Pseudomonas fluorescens—NCIM2100), organic manure (OM), and chemical amendment—Ethylene Diamine Tetra Acetic acid (EDTA) on Atriplex lentiformis were explored in cadmium- and nickel-contaminated soil. Heavy metal resistance and plant growth-promoting traits of PGPB were also analyzed. Augmentation with a combination of both PGPB and OM, A. lentiformis displayed maximum uptake of Ni (45.67 mg kg−1 in roots; 24.68 mg kg−1 in shoots) and Cd (14.15 mg kg−1 in roots; 7.19 mg kg−1 in shoots). Highest Ni uptake in shoots was observed under the EDTA amendment (25.33 mg kg−1). Metal uptake by A. lentiformis under NCIM2100 was greater than NCIM5350 for both Cd and Ni (10.57 and 43.87 mg kg−1). Among all the amendments highest metal uptake was recorded under bio-organic treatments (PGPB1 + PGPB2 + OM) for both Cd and Ni (14.15 and 45.67 mg kg−1), respectively. The results showed that this association has significantly improved the plant height, biomass, chlorophyll, MDA (Malondialdehyde) content, and the activity of antioxidative enzymes (CAT, APX, and SOD) which exhibited a positive correlation with metal uptake at 1% level of significance and the potency of synergistic impact of microbial consortium, while EDTA reduced the growth of the plant. Metal uptake under EDTA was also much lower than biological amendments. Higher metal values in roots establishes A. lentiformis as a phytostabilizer thus indicating its suitability as a safer forage. Biological amendments-based phytoremediation holds great promise and could be used in future to give further impetus to the antioxidative defense, phytoremedial potential, and growth of this and other important forage plants.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Environmental pollution effectuated by inorganic contaminants like heavy metals (HM) has encountered increasing attention and awareness worldwide. The problem of HM contamination is all over the world and with the day by day increasing magnitude of pollution, the intractable and pertinacious character of HM has become a serious threat to the environment as it affects the lives of both plants and animals, causing serious diseases in humans (Wani et al. 2018; Kubier et al. 2019). Recently, American Agency for the Toxic Substances and Disease Registry (ATSDR 2019) has listed out arsenic (first place), lead (second place), mercury (third place), and cadmium (seventh place) in their annual list. Pollution from HM and metalloids, like As, Co, Ni, Cu, Zn, Cd, Hg, and Pb, among others, can out-turn inconspicuous toxic consequences (Zaid et al. 2020). Some of these metal ions are essential for many physiological pathways (Ni, Zn, Co, Cu) in living beings while others have no such biological functions. These HM are bio-accumulative and may slowly enter the next trophic level in the progression of a food chain over a certain period. Worldwide, industries like smelting of metalliferous surface finishing, electric appliances, installation of aerospace and atomic plants, leather, metallurgy, mining, use of inorganic fertilizers and pesticides, application of sewage sludge in lands, wrong agricultural practices, inappropriate waste disposal, and some military operations have directly, obliquely, or incidentally released ample and noticeable amount of toxic HMs into the environment with an alarming perilous influence on both ecological and anthropoid health, preponderantly in developing countries (Toth et al. 2016; Ahmad et al. 2022). Irrigation of crop with water containing industrial waste and sewage sludge have increased the concentration of the HM in agricultural soil at toxic level (Woldetsadik et al. 2017). Sewage sludge, manure, and limes have been characterized as one of the prime causes of Cd contamination in agricultural soil. As Cd is a highly water-soluble metal exhibiting considerably high level of toxicity even at very low concentrations, United States Environmental Protection Agency (USEPA) categorized Cd as an extremely threatening and toxic pollutant to all life forms, with carcinogenicity in humans (group B1). Cd toxicity results in the building of free radicals like reactive oxygen species (ROS) which cause oxidative damage to the cell and muddle the redox equilibrium of the cell with a concomitant decrease in the biosynthesis of enzymatic and non-enzymatic antioxidants (Zaid et al. 2019a, b).
However, unlike Cd, nickel (Ni) is an essential trace element (17th) for plant growth and physiological system. Ni can affect plant health both positively and negatively, depending on the concentration present in the growth medium. It can be toxic and possibly carcinogenic for human beings; constituting almost all types of soils, covering a diversified range of climatic ecosystems. Ni contamination has been found in at least 872 of the 1,662 sites identified for hazardous waste that has been recommended for incorporation on the U.S. Environmental Protection Agency (EPA) National Priorities List (NPL) (HazDat 2006). Toxicity of Ni in the soil can slow down the plant’s enzymatic machinery such as respiration, photosynthesis, plants water status (transpiration, leaf succulence), and the capability of antioxidative defense in response to the production of ROS (Gill and Tuteja 2010). The toxic effects of higher concentrations of Ni have been reported at multiple levels, including suppression of mitotic activity (Rao and Sresty 2000), photosynthesis, decrease in plant growth, and inhibition of nitrogen metabolism (Zaid et al. 2019a, b).
HM-induced generation of ROS in plants decimate the physiological functions of the plant and strikes almost all the prime cellular organelles and components such as cell membrane, mitochondria, vacuoles, chloroplasts, nucleic acids, enzymes, lipids, and protein (Ishtiyaq et al. 2018; Wani et al. 2018). Yellowing of leaves (chlorosis), necrosis, and deformed/reduced growth are some visual indications of Ni and Cd toxicity.
To mitigate the HM toxicity, plants have evolved a counter antioxidative defense mechanism composed of many enzymatic and non-enzymatic antioxidants, such as catalase (CAT), superoxide dismutase (SOD), ascorbate peroxidase (APX), glutathione (GSH), glutathione reductase (GR), guaiacol peroxidase (POX), proline, carotenoids, flavonoids, and tocopherols, that instigate efficient and stalwart scavenging of ROS (Ishtiyaq et al. 2021). As land and water are some of the most influential and crucial natural resources, they require attentive and vigilant management for HM pollution in order to attain the United Nations’ Sustainable Development Goals.
Many factors such as low biomass production at HM concentration, sensitivity to multi-metals, poor growth, and shallow root system limited the phytoremediation efficiency of the plants. Hence, to improve the phytoremediation efficiency of the plant at higher metal concentration is a promising approach, which may be attained by utilizing biological amendments (PGPB—Plant growth-promoting bacteria), chelating agents (EDTA—Ethylenediaminetetraaceticacid), and organic manure (OM).
The most common implies the use of chelating agents, such as EDTA, which enhances the metal bioavailability (Cui et al. 2015). However, they may remain persistent in the environment and result in groundwater contamination. Large quantities of organic alteration, such as organic manure, compost, urban solid wastes, and biosolids, have been used as a source of nutrients or as a conditioner to enhance the physical properties and fertility of soils (Adekiya et al. 2020). Modification of organic matter can alter the pH of the soil and thus have an indirect effect on the bioavailability of HM (Kelvin et al. 2020).
As plants and soil microbes (PGPB) have a strong complementary relationship, phytoremediation based on the combined use of plants and PGPB is currently a research hotspot (Kong and Glick 2017; Mesa-Marin et al. 2020). When PGPB are used as a bioinoculant, they increased biomass and root growth of the plant through nutrient recycling, stabilizing the soil structure, and modulating the bioavailability and toxicity of HMs (Ahemad 2019). To date, the majority of PGPB-assisted HM remediation and plant growth studies have been conducted with glycophytes, such as Thlaspi caerulescens (Whiting et al. 2001), Sedum plumbizincicola (Ma et al. 2016), Pongamia pinnata (Yu et al. 2017), Napier grass (Wiangkham and Prapagdee 2018), and Festuca arundinacea (Seniyat and Lesley 2019). However, to the best of our knowledge, effects of amendments like PGPB, OM, and EDTA on phytoremediation potential of Atriplex lentiformis grown under HM stress have not been explored previously. Hence, physiological and biochemical response of A. lentiformis under PGPB, OM, and EDTA needs to be discoursed.
Atriplex lentiformis (Torr.) S.Wats., a halophytic plant is known to exhibit highly developed tolerance mechanism under varied pressed conditions like ionic and osmotic pressure of salinity, excess of toxic ions, cold, and xeric environment with the capability to maintain a propitious water potential gradient to guard their cellular structures (Meyer 2005; Soliz et al. 2011). The additional attributes of high forage production (with considerably high content of protein) especially in view of large semi-arid regions in India with very poor vegetation cover and extremes of biotic pressure due to excessive grazing by a very heavy population of cattle load makes this species as an ideal one in this context. This species also has an ability to restore degraded agricultural lands and soil erosion prevention. However, very few studies are available with regard to the metal accumulation capacity of A. lentiformis for phytoremediation of contaminated soil.
In the current investigation, a halophytic species (Atriplex lentiformis) has been explored for the remediation of Cd and Ni with two PGPB (Pseudomonas fluorescens and Bradyrhizobium japonicum), EDTA and OM for the possible optimization of its growth, physiology, defense, and metal uptake ability. The reason of using Pseudomonas and Bradyrhizobium for the alleviation of the taken problem has been done after extensive research and validation of characteristics like Cd and Ni resistance capacity, many plant growth-promoting traits, and compatibility with each other, as documented by Al-Dhabi et al. 2019, Alsohim 2020; and Zeffa et al. 2020, respectively. Through this research work, possibility of developing a viable and self-sustainable phytoremedial—cum—safe forage supply system is being explored using A. lentiformis especially under the pressing situations.
Materials and Methods
PGPR Material, Compatibility Assessment, and Characterization of Plant Growth-Promoting Traits
Two PGPR strains (Bradyrhizobium japonicum—PGPB1 and Pseudomonas fluorescens—PGPB2) were procured from National Chemical Laboratory, Pune, India in freeze-dried cultures and stored at − 20 °C. Both cultures (NCIM5350 and NCIM2100) were revived according to the specified and optimum incubation conditions prescribed for each strain. The identity of 16S rRNA sequence of both strains was done by performing a resemblance against the GenBank database (http://www.ncbi.nih.gov/BLAST). Existing 16S rRNA gene sequences (from the National Center for Biotechnology Information, NCBI GenBank database) of approximately related bacteria were used to construct the phylogenetic tree by the neighbor-joining method using MEGA6.0 software (Saitou and Nei 1987; Tamura et al. 2013). Cd and Ni resistances of both PGPB strains were tested by streaking them on King’s B medium (for PGPB1) and Tryptone Yeast extract agar (for PGPB2) supplemented with a standard aqueous solution of Cd (Merck-119777) and Ni (Merck-119792) of 400-ppm concentration and incubated at 30 °C. After 4 days of incubation, single colonies were picked from the Petri plates and sub-cultured to get pure cultures. Stock cultures were made in nutrient broth containing 50% (wt/vol) glycerol and stored at − 80 °C.
After the resistance test, both PGPR isolates were tested for plant growth-promoting traits, like ACC (1-aminocyclopropane-1-carboxylate) deaminase, siderophore, IAA (Indole acetic acid) production, phosphate solubilization, and available phosphate. ACC deaminase activity was determined by the modified protocol of Honma and Shimomura (1978) and Penrose and Glick (2003). Individual colonies of each PGPB strain were inoculated in nutrient broth test tubes (10 ml) and kept for 3 days in a shaker (120 rpm) at 30 °C. After centrifugation (6000 rpm) for 10 min, bacterial cells were collected and washed with 1% NaCl solution. Collected cells were again added to 10 ml of nutrient broth containing 5-mM ACC. Number of mmol of α-ketobutyrate produced by the reaction was measured by spectrophotometer at 540 nm using a standard curve of α-ketobutyrate. Both qualitative and quantitative assays were corroborated for siderophore production in both PGPR strains by Chrome Azurol S (CAS) Assay (Schwyn and Neilands 1987). The formation of bright zone with yellow fluorescent color by the culture in the medium confirmed the production of siderophore. IAA production was determined by Gorden and Paleg (1957) method using lysogeny broth (LB) supplemented with 0.01% wt/vol L tryptophan as the precursor of IAA. IAA concentration (μg/ml) was calculated by the standard curve of known concentration of IAA solution. Phosphate solubilization by both PGPBs was identified by Katznelson and Bose (1959) protocol and marked positive (clear halo around colonies against an opaque background) and negative (colonies without halo). Sperberg’s hydroxyapatite broth (20 ml) was used for quantitative estimation of phosphate solubilization and was carried out in Erlenmeyer flasks inoculated with PGPB (500 mL inoculum with ~ 2 × 108 CFU/mL). Sterilized medium without PGPB inoculum was treated as control. After incubation for 5 days at 28 °C, the supernatant was harvested by centrifugation at 10,000 rpm for 10 min and used to assay the available phosphorus (P) (Olsen and Sommers 1982). The amount of solubilized phosphorus was calculated using the standard curve of orthophosphate. The available P content was expressed in mg/ml.
Both PGPR strains were tested for their compatibility before applying them to plants. Compatibility of both PGPR was tested between themselves by streaking one PGPR on one side and the other PGPR perpendicularly up to the test PGPR (as cross-streak). The growth was visually observed after 3 days and recorded as positive or negative.
Plant Material and Experimental Design
Certified seeds of Atriplex lentiformis were procured from the United States Department of Agriculture—Agriculture Research Service (USDA-ARS), Washington State University, USA, and Forest Research Institute, Jodhpur, Rajasthan, India. The seeds were first sterilized in 0.1% HgCl2 and then soaked in petri plates containing double distilled water or bacterial suspensions having 108 CFU ml−1 (according to the treatment plan) for 3 h, after this seeds were kept in dark and allowed to germinate. A greenhouse pot experiment was set up in the botanical garden of Department of Botany, St. John’s College, Agra (27.18° N 78.02° E), India. The experimental soil was collected from the botanical garden of the college (sandy loam-sand 60–80%, silt 10–25%, clay 8–15%) with pH 7.18 ± 0.51, electrical conductivity (EC 1:2.5) 1.04 ± 0.23 ds/m, moisture content 8.32 ± 0.59% and 1.10 ± 0.1%, 20.00 ± 0.7 kg/ha, 320.65 ± 5.4 kg/ha, 616 ± 15 kg/ha of organic carbon, available P2O5, available nitrogen, and available K2O, respectively. Sodium (Na), calcium–magnesium (Ca + Mg), and potassium (K) content of soil was 730 ± 5, 4.4 ± 0.5, and 23 ± 4 mEq l−1, respectively. Each plastic pot was filled with autoclaved and air-dried 4-kg soil and artificially spiked with respective salts of CdCl2.5H2O and NiSO4.6H2O as a source of HMs depending upon the treatment to be given. Pot soil was thoroughly mixed and kept aside for 14 days in order to stabilize. After emergence of the first five true leaves, seedlings were again soaked in bacterial suspension for 2 h as per the treatment plan and were transferred manually in pots having 4 kg soil and placed into a growth chamber with a 16/8-h light/dark cycle, day/night temperatures of 27/22 °C, and 70–80% relative humidity. The pots were dampened with distilled water once per day. After 10 days, 5 ml of PGPB inoculum (106–108 CFU ml−1), EDTA (5 mmol/kg soil), and 250 g of organic manure (with 40–50% organic matter) were injected near the rhizosphere of seedlings in accordance with the treatments. The treatments selected were as follows: 1. Control 0 ppm HM; 2. Cd25 ppm; 3. Cd50 ppm; 4. Cd25 ppm + PGPB1; 5. Cd25 ppm + PGPB2; 6. Cd25 ppm + OM; 7. Cd25 ppm + EDTA; 8. Cd25 ppm + PGPB1 + PGPB2 + OM; 9. Ni50 ppm; 10. Ni100 ppm; 11. Ni50 ppm + PGPB1; 12. Ni50 ppm + PGPB2; 13. Ni50 ppm + OM; 14. Ni50 ppm + EDTA; and 15. Ni50 ppm + PGPB1 + PGPB2 + OM. All treatments were maintained in triplicates, including the control and kept in randomized block design. At the end of the study, i.e., 120 DAT (days after treatment), pots were dismantled and each plant was washed in deionized water and divided into root and shoot and analyzed for metal content. The soil was also tested to find the residual metal content. In addition, chlorophyll, proline content, lipid peroxidation, root length, shoot length, biomass, and antioxidative enzyme activity of plants were also measured.
Biochemical Analysis
Chlorophyll content of leaf was estimated as per the method given by Lichtenthaler (1987). The amount of proline in plants was calculated according to the method given by Bates et al. (1973). Lipid peroxidation was expressed as MDA (malondialdehyde) content and was measured by Heath and Packer (1968) method.
Antioxidative Enzyme Assay
The samples of the plant leaves were first processed by washing (2–3 times) in distilled water. One gram of leaf sample was crushed in liquid nitrogen using a mortar and pestle with 10 ml of 0.1-M phosphate buffer (pH 7) containing 0.5-mM EDTA in case of SOD and CAT and 0.5-mM EDTA and 1-mM ascorbic acid in case of APX. The filtrate was passed through 4 layers of cheesecloth followed by centrifugation at 16,000 rpm for 20 min at 4 °C, and the supernatant was used as an enzyme extract to measure the antioxidant activities. Bradford assay was used to determine the soluble protein content in the enzyme extract. Bovine serum albumin (BSA) was used as the standard.
Catalase (EC 1.11.1.6) activity was measured as a decrease in absorbance (240 nm) according to the method of Aebi (1984). Reaction mixture (3 ml) containing 50 µl of enzyme extract with 1.5 ml of 0.1 M phosphate buffer (pH 7), 0.5 ml of 75 mM H2O2 (Hydrogen peroxide), and distilled water to make up the volume was used to measure the enzyme activity by comparing decomposed amount of H2O2 (initial reading–final reading) with a standard curve drawn with known concentrations of H2O2 and expressed as µmol H2O2 consumed/min/mg protein (extinction coefficient = 37.5 mM−1 cm−1).
Ascorbate peroxidase (EC 1.11.1.11) activity was determined at 290 nm for a period of 30 s as per the method given by Nakano and Asada (1987) and expressed as µmol ascorbic acid oxidized min−1 mg protein−1. The reaction was started with the addition of H2O2 to the reaction mixture containing phosphate buffer (50 mM), EDTA (0.5 mM), and ascorbic acid (3 mM).
Superoxide dismutase (EC 1.15.1.1) activity was estimated by measuring the inhibition of photochemical reduction of nitroblue tetrazolium dye (NBT) by the enzyme as a decrease in absorbance (at 560 nm) according to Dhindsa et al. (1981). Methionine (13.33 Mm), NBT (75 µM), EDTA (0.1 mM), 50-mM buffer, 50-mM Na2CO3, and 0.1-ml enzyme with water (to make up a final volume of 3 ml) was used as a reaction mixture. The reaction was started by keeping the tubes in light of 15-W fluorescent lamp with 2-μM riboflavin for 15 min followed by placing of the tubes in dark to stop the reaction. Control was used as a complete reaction mixture (colored) without enzyme, while a non-irradiated mixture was used as a blank.
Heavy Metal Analysis
After 120 DAT, soil, root, and shoot samples were collected and dried in an oven at 80 °C for 48 h followed by a microwave-assisted wet digestion method with 3 mL HNO3 (69%, Merck) + 9 mL HCl (30%, Merck) for 0.5 g soil, and 5 mL HNO3 (69%, Merck) + 2 mL H2O2 (30%, Merck) for 0.5-g plant sample. The filtrate was analyzed for metal (Cd, Ni) by graphite furnace atomic absorption spectrophotometry using a Solaar M2–Thermo Unicam instrument. Both blank and standard reference materials (Virginia tobacco leaves CTA-VTL-2, Polish Certified Reference Material, and NIST2709–San Joaquin Soil) were included for quality assurance. The recovery rates for the elements analyzed were 89% for Ni and 93% for Cd.
Statistical Analysis
The experiment and all the tests were performed in triplicate. Data recorded for the calculation of plant growth, photosynthetic pigments, proline, lipid peroxidation, metal accumulation, and antioxidative enzyme activities were analyzed using one-way ANOVA (SigmaPlot 11.0) and the mean differences were detected using Tukey’s LSD test (α < 0.05 level). Pearson’s coefficient of Correlation was calculated at a significance level of p ≤ 0.05 and p ≤ 0.01.
Results
Identification, Compatibility, Screening, and Characterization Of PGPB
The PGPB strains (NCIM5350 and NCIM2100) used in this study were phylogenetically identified by 16S rRNA sequencing and showed close resemblance with Bradyrhizobium japonicum (PGPB1) and Pseudomonas fluorescens (PGPB2), respectively (Table 1). The obtained gene sequences were lined up with NCBI (National Center for Biotechnology Information) database and were handed down for the formation of a phylogenetic tree (Fig. 1). Both the strains of PGPB were found to be compatible in cross-streak assay as they did not inhibit the growth of each other and were able to grow simultaneously in the same Petri plate without showing any inhibition zone. Screening experiment results of PGPB1 and PGPB2 for Cd and Ni (up to 400 ppm) resistance exhibited their tolerance toward both the metals as clear colonies were obtained after incubation on King’s B media and Tryptone Yeast extract agar, respectively, supplemented with the respective HM.

Phylogenetic tree based on partial 16S rRNA gene of selected plant growth-promoting metal-tolerant bacterial strain showing similarity with Pseudomonas fluorescens and Bradyrhizobium japonicum (accession numbers are in parentheses)
In addition to metal tolerance, PGPB1 and PGPB2 also exhibited some plant growth-promoting traits (Table 2). Both PGPB produce fluorescent yellowish orange bright colonial zones that confirmed the production of siderophores. The PGPB2 was found to produce ACC Deaminase (82.4 nmol α-ketobutyrate/mg protein/hour) while PGPB1 showed negative results for the same. IAA production was exhibited by both the PGPBs. Both of them showed good phosphate solubilization efficiency. The strains did not show any disease symptoms in A. lentiformis and were found to be highly compatible with standard bioinoculants.
Plant Growth Response
Treatments with both the metals (Cd and Ni) significantly affected the growth parameters of A. lentiformis like root length, shoot length, and biomass which were in positive correlation with metal concentration (Figs. 2 and 3). Plants treated with Cd experienced more toxicity (low shoot, root length, and biomass) as compared to Ni treatments (Table 3). A number of toxicity symptoms like rolling of leaves, chlorosis, and stunted growth were observed visually throughout the experiment in plants treated with Cd. Plants treated with PGPB displayed better growth than control even in the presence of metals or in comparison to EDTA or OM. The application of biological amendments (PGPB1 + PGPB2 + OM) were found to be the most effective treatment to overcome the stress generated by the toxicity of Cd/Ni. They significantly promoted plant growth as compared to the plants treated with metals only (Cd25, Ni50). Maximum biomass was observed in plants treated with Cd25 + PGPB1 + PGPB2 + OM (38.54 g) and Ni50 + PGPB1 + PGPB2 + OM (40.84 g) indicating the synergistic impact of both PGPBs (Table 3).

Relationship of Cd uptake with catalase activity (a); superoxide dismutase activity (b); peroxidase activity (c); shoot length (d); root length (e); and biomass (f)

Relationship of Ni uptake with catalase activity (a); superoxide dismutase activity (b); peroxidase activity (c); shoot length (d); root length (e); and biomass (f)
Chlorophyll, MDA, and Proline Content
Data presented in Fig. 4 show a greater reduction in the total chlorophyll content of leaves due to Cd as compared to Ni stress. Inoculation with Pseudomonas fluorescens and Bradyrhizobium japonicum was found to be the most effective treatment for scaling down the adverse effects of Cd and Ni on chlorophyll, although EDTA and OM also improved the chlorophyll content as compared to the non-inoculated plants. The data showed that the order of chlorophyll reduction for both the metals was as follows: 10.2 and 18.2% under 25- and 50-ppm concentration of Cd, whereas 5.2 and 17.3% under 50- and 100-ppm concentration of Ni.

Effect of heavy metals, amendments, and PGPB on chlorophyll content in A. lentiformis
Lipid peroxidation in leaves of A. lentiformis measured as MDA content is presented in Fig. 5. It was significantly increased due to Cd and Ni stress. Maximum MDA content was observed in Cd50 (10.12 µg/g/FW) and Ni100 (9.45 µg/g/FW) treatments which were about twofold higher than control. However, in treatments with amendments, a significant decrease was observed especially in Cd25 + PGPB1 + PGPB2 + OM and Ni50 + PGPB1 + PGPB2 + OM treatments. A. lentiformis with a high MDA level indicate high lipoxygenase activity.

Effect of heavy metals, amendments, and PGPB on MDA content in A. lentiformis
Accumulation of stress indicating osmolyte (proline) was found to be notably affected under metal stress. An increase up to 90.3% (Cd25 ppm), 135% (Cd50 ppm), 42.3% (Ni50 ppm), and 124.6% (Ni100 ppm) was reported, respectively (Fig. 6). Similar to MDA, proline content also showed a gradual decrease at all time intervals, in plants treated with amendments and PGPB.

Effect of heavy metals, amendments, and PGPB on proline in A. lentiformis
Antioxidative Defense System, Amendments, PGPB, and Heavy Metal Uptake
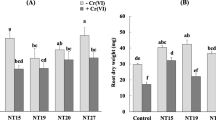
All the antioxidative enzymes that had been measured at 120 DAT showed remarkable variation in accordance with various treatments and amendments. Antioxidative activity was found to be in markedly significant correlation with metal uptake (for both the metals) at 1% level of significance (Table 3 and 4). Application of all the amendments and PGPB increased the metal uptake and improved the overall plant health. Among all the treatments highest fluctuation in antioxidative activity was found in Cd25 + PGPB1 + PGPB2 + OM and Ni50 + PGPB1 + PGPB2 + OM treatments (Fig. 7).

The activities of ROS antioxidant enzymes (CAT, SOD, and APX activity) in A. lentiformis. Each value is a mean ± SD of replicates (n = 4); values followed by the same superscript letter in each bars are not significantly different from each other (Tukey’s LSD; p ≤ 0.001)
All the three enzymes showed a positive interconnection with metal accumulation and both PGPBs, as test plants with high metal accumulation (Cd25 + PGPB1; Cd25 + PGPB2; Cd25 + PGPB1 + PGPB2; Ni50 + PGPB1; Ni100 + PGPB2; Ni50 + PGPB1 + PGPB2) also showed highest enzyme activities displaying the influence of PGPB1 and PGPB2 (Fig. 7). Order of treatment efficiency affecting metal uptake was as follows: Cd25 + PGPB1 + PGPB2 + OM > Cd25 + PGPB1 > Cd25 + PGPB2 > Cd25 + EDTA > Cd25 + OM for Cd and Ni50 + PGPB1 + PGPB2 + OM > Ni50 + PGPB1 > Ni50 + PGPB2 > Ni50 + EDTA > Ni50 + OM for Ni. A. lentiformis showed phytostabilization of Cd and Ni as the roots of the plant exhibit greater potential for accumulating given metals especially under the ascendancy of Pseudomonas fluorescens over Bradyrhizobium japonicum. Twofold increase in Cd uptake was measured in roots (Cd25 + PGPB1 + PGPB2 + OM) with highest CAT activity thus Cd stabilization by plant showed strong interdependence with CAT activity with a correlation coefficient value of 0.879. Cd uptake reached maximum (67%) in Cd25 + PGPB1 + PGPB2 + OM treatment and minimum (48.2%) for Cd25 + EDTA. Thus amply demonstrating the edge of the biological inoculants (PGPB) over the chemicals (EDTA). A linear relationship has been observed between the CAT activity and metal uptake in regression analysis with R2 value of 0.889 for Cd and 0.768 for Ni. Similar trends were also reported for SOD and APX.
Increased metal uptake exhibited a strong dependence on antioxidative defense activity of the plant; however, the ratios of changes in enzyme concentration differ for both the metals. Correlation between Cd treatments and CAT activity was most prominent as compared to the other enzymes (Table 4).
Discussion
In the present study amendment-based phytoremediation strategy for HMs was worked out based upon certain selection criteria like capability to improve the plant growth, the bioavailability of HM in soils, and the selection of an appropriate plant species with the potential to withstand stress and characteristics required for metal uptake. Hence, this study was carried out on Atriplex lentiformis with two metal-resistant PGPBs, OM, and EDTA. It was found that Atriplex lentiformis has the potential to remediate HM from the soil especially when its potential is combined with PGPBs, OM, and EDTA. This plant is equipped with inherent specific and robust mechanism of salinity tolerance and can survive over a wide range of stress-producing abiotic factors that seems to analog and turn-up mechanisms that can confer HM tolerance (Al-Aqeel and Vinod 2016; Bulent et al. 2019).
PGPBs are the plant-associated free-living, soil-borne bacteria, which have the ability to enhance the plant growth by facilitating the soil nutrient availability, stimulating root growth, cell division, suppressing the HM-induced toxicity and plant pathogens, and improving induction of systemic resistance. PGPB1 and PGPB2 strains used in the present study were phylogenetically identified by 16S rRNA sequencing and showed close resemblance with Bradyrhizobium japonicum (PGPB1) and Pseudomonas fluorescens (PGPB2). Further, screening experiment results of PGPB1 and PGPB2 for Cd and Ni (up to 400 ppm) resistance showed their tolerance toward both the metals as clear colonies were obtained after incubation on King’s B media and Tryptone Yeast extract agar, respectively, supplemented with the respective HM. Mechanisms which impart resistance to HM in bacterial species include deposition of metal in the cell wall and vacuole, accumulation, and modification of toxic metal into less toxic form (Tang et al. 2018). Both the strains were found to be positive for oxidase, CAT, and citrate. Chellaiah (2018) demonstrated Pseudomonas for its Cd-resistant capability (up to 500 ppm) and reported that the PGPB was able to maintain its plant growth-promoting traits up to 200-ppm Cd concentrations in soil. P. fluorescens and B. japonicum have the ability for Ni sequestration and plant growth promotion (Seneviratne et al. 2016). They can directly enhance the metal uptake through modifying metal bioavailability in the rhizosphere by altering soil pH, chelator release (e.g., organic acids, siderophores), and oxidation/reduction reactions. Egamberdieva et al. (2017) explored the coordination between Bradyrhizobium and Pseudomonas and found that they can synergistically improve the tolerance level of the plant by changing the architecture of root system and promote nitrogen and phosphorus acquisition with increased nodule formation, thus this study supports the results of compatibility assay for Bradyrhizobium and Pseudomonas. PGPB are known to boost soil fertility by the production of siderophores and phytohormones and mitigate the ethylene-regulated strain by synthesizing ACC deaminase and enhance plant stress tolerance to drought, salinity, and HM toxicity (Jahanian et al. 2012; Maxton et al. 2018; Gupta and Pandey 2019; Kang et al. 2019; Orozco-Mosqueda et al. 2020). The use of PGPB possessing the measured multiple plant productive properties as well as metal resistance and detoxifying characters came out to be an assured, cost effective, and environment supporting HM bioremediating tool.
In this experiment, Cd toxicity significantly reduced root and shoot length along with reduced biomass. This may be attributed to the inhibition of mitotic activity of cells due to Cd exposure, which leads to reduced root length and dry biomass (Gratão et al. 2009). Cd exposure causes osmotic stress in plants by lowering water content, stomatal conductance, and transpiration rate, therefore results in physiological damage (Rizwan et al. 2016; Zaid et al. 2022). Cd also interferes with the uptake and transport of P, K, Mg, and Ca (Nazar et al. 2012). However, the application of inoculants of both the PGPB with OM were found to be the most effective amendment in overcoming stress generated by the toxicity of Cd/Ni thereby significantly promoting plant growth as compared to the metal-treated plant (Cd25, Pb50). Similar results were also reported by Tank and Saraf (2009) on selected five strains of microbes testing their potential as plant growth promoters, on the basis of their phosphate solubilization ability, IAA, siderophore, HCN (Hydrogen cyanide) production, and bio-control potentials. The results suggested that the use of these PGPB can enhance plant growth in Ni- and Cd-spiked soil and can remediate them from contaminated sites. Maria et al. (2010) studied the interaction of Atriplex nummularia with halotolerant and bioprospect nitrogen-fixing bacteria. The results from this study indicated that the test PGPBs when injected at the seedling level besides seed treatments and could produce desirable effects like fixing nitrogen and promoting growth under HM stress, thus exhibiting their high adaptability and efficiency in shorter duration.
These microorganisms can directly enhance the phytoremediation cycle through modifying metal bioavailability by altering soil pH, chelator release (e.g., organic acids, siderophores), and oxidation /reduction reactions. The present study is also consistent with previous findings of Treesubsuntorn et al. (2018) who documented that Bacillus subtilis and B. cereus, when inoculated to Cd-exposed Oryza sativa plants led to higher root and shoot biomass. This may be due to the development of plant growth hormone (IAA) by augmented microbes that control the hormones within plant tissues and make them adjust to environmental stresses. A study conducted by Egamberdieva et al. (2017) explored the coordination between Bradyrhizobium and Pseudomonas and found that they can synergistically improve the tolerance level of the plant by changing the architecture of root system and promote nitrogen and phosphorus acquisition with increased nodule formation.
Plant antioxidant system is severely altered by HM stress, through the generation of ROS molecules, such as superoxide radicals, hydrogen peroxide, and hydroxyl radical, which cause oxidative damage leads to cell death (Del Río et al. 2003; Zaid and Wani 2019). Under these circumstances PGPB strains can protect plant from ROS induced oxidative damage by the reduction of ROS generation by the production of various enzymatic and non-enzymatic antioxidants, therefore regulate the ROS level in the plant (Karthik et al. 2016). Inoculation of Pseudomonas sp. CPSB21 decreased the ROS generation in Helianthus annuus (sunflower) and Solanum lycopersicum (tomato) by increased production of enzymatic antioxidants, such as SOD and CAT (Gupta et al. 2018). Likewise, in the present investigation, PGPBs enhanced the plant metal tolerance level, which can be corroborated with the modulation of the antioxidative enzymes that play a vital role in resisting oxidative damage as an adaptive strategy of the plant for survival under metal stress thereby providing an impetus to plant biomass, root length, shoot length, and overall growth. These results confirmed the ability of Atriplex to grow well in Cd/Ni-contaminated soil.
Increased accumulation of HMs like Ni and Cd by hyperaccumulators plants such as Brassica napus and Brassica juncea was reported when the plants were inoculated with Bacillus sp. (Zaidi et al. 2006). The results demonstrated that the synchronized use of both microorganisms and plants for soil remediation resulted in faster and more efficient cleaning of the polluted sites (Weyens et al. 2009).
HM toxicity can result in a reduction of chlorophyll content either by inhibition of chlorophyll biosynthesis or acceleration of its degradation (Gopal and Rizvi 2008); this was in conformity with the findings presented in Fig. 4. A decrease in the content of chlorophyll pigment due to Cd/Ni has also been reported by Szopiński et al. 2019. The possible reason for the reduction of the photosynthetic pigment may be ascribed to the fact that excess metal hampers the uptake of Mg and Fe (Piccini and Malavolta 1992). However, the inoculation of both PGPB significantly improved the chlorophyll content (Fig. 4). The synergistic impact of both PGPB strains with OM under Cd and Ni contamination resulted in highest increase of chlorophyll content by 24.4 and 34.5%, respectively, as compared to single strain inoculum for both Cd/Ni treatments. Chlorophyll content in Cd25 + PGPB1 + PGPB2 + OM and Ni50 + PGPB1 + PGPB2 + OM was 1.3 and 1.4 times higher than observed under Cd25 and Ni50, respectively. Inoculation of Klebsiella pneumoniae in Vigna mungo enhanced the levels of chlorophyll under Cd stress (Dutta et al. 2018). It was reported by Rizvi et al. (2019) that Azotobacter chrococcum, when augmented with Cu- and Pb-exposed Zea mays plants, enhanced the chlorophyll contents. Increase in chlorophyll upon PGPB inoculation could be associated with the change of microbial population in the rhizosphere of the plant and the synthesis of various growth substances like IAA, siderophore, phosphate solubilization (improved acquisition of iron, nitrogen, phosphorus, and essential minerals), reduced oxidative damage, and ethylene production by them.
In the present investigation, the imposition of Cd/Ni induced a significant increase in MDA and H2O2 contents in Atriplex plants showing the role of excess toxic metals in oxidative stress. When toxic metal ions enter the cell they react with H2O2 to form free radicals (OH− in a Haber–Weiss and Fenton reactions) and damage the plant cell by initiating non-specific lipid peroxidation (MDA synthesis) and leading to the synthesis of hydroperoxyl fatty acids (Garnier et al. 2006). In contrast, a substantial decrease in the content of MDA and H2O2 has been observed in plants treated with bacterial inoculation, which is the indication that a better protective mechanism exists in bacterial-inoculated plants as they have developed various defense mechanisms to scavenge free radicals and peroxides. The rise in the cellular level of H2O2 may be due to higher MDA contents (Rowe and Abdel-Magid 1995). Combined application of PGPB with OM decreased the MDA content, which was in congruence with the earlier studies, correlating the regulation of lipid peroxidation to better stress tolerance mechanisms (Liu et al. 2013) and reflects the ameliorative ability of selected PGPB and OM to oxidative stress.
Osmotic adjustment under HM stress is an adaptation mechanism operated by both halophytes and glycophytes in order to maintain their water balance (Flowers and Colmer 2008). Besides the accumulation of HMs and its sequestration in the vacuole, the osmotic balance between vacuole and cytoplasm in response to HM stress is through the synthesis of organic solutes to retain the stability of the proteins in cells (Zhang et al. 1999). Plants synthesize a variety of organic solutes, such as proline, glycine betaine, and soluble sugars, which are collectively known as osmolytes. These are accumulated in high concentrations in cells without disturbing cellular biochemistry and functions (Cushman 2001). They protect subcellular structures, mitigate oxidative damage caused by free radicals and maintain the enzyme activities under stress environment (Ahmad et al. 2019; Yokoi et al. 2002). Proline accumulation is an adaptive strategy of the plant to various abiotic stresses, and it plays a significant role in the detoxification of ROS, stabilization of proteins and protein complexes (Suprasanna et al. 2014; Slama et al. 2015; Nazir et al. 2019). Application of amendments under Cd/Ni stress resulted in reduction of proline in A. lentiformis, which manifests improved physiology, reduced oxidative stress, and adaptation toward HM toxicity acting as a sensing or signaling code for lesser proline accumulation in plant. Similarly, decrease in plant proline contents upon PGPB inoculation has been associated with lower membrane damage (MDA and H2O2) in maize plants under HM by Islam et al. (2014). The proline accumulation is frequently reported in halophytic plants exposed to HM stress and has been correlated with a plants capacity to tolerate and adapt to HM stress (Errabii et al. 2007; Slama et al. 2008). Besides proline, HM stress also causes increased accumulation of glycine betaine in halophytic species Sesuvium portulacastrum (Lokhande et al. 2010). Wang and Showalter (2004) reported higher accumulation of glycine betaine in Atriplex prostrate, which plays a positive role in maintenance of membrane integrity and stability under HM stress. Furthermore, proline accumulation also has been suggested to have a role in detoxification of ROS (Szabados and Savoure 2009).
One of the main limits of phytoextraction is the low solubility and availability of HM for root uptake. Chelating agents used to extract metals from soils, among chelators EDTA is regarded as the most effective in solubilizing soil-bound HMs (Nascimento et al. 2006). Moreover, EDTA has been shown to increase HM movement to roots via mass flow or diffusion, enhance HM uptake, and trigger root to shoot translocation of HM (Nascimento et al. 2006).
In the present investigation, we tested the metal tolerance and accumulation ability by the halophyte species A. lentiformis. We also assess EDTA efficiency for improvement of Cd/Ni by this species. Our results indicated that A. lentiformis do not exhibit any toxicity symptom, such as chlorosis, necrosis, or reduced growth. This confirms its strong tolerance to Cd/Ni as already shown by similar studies (Van Engelen et al. 2007; Zaier et al. 2010). On the other hand, considering the low bioavailability of Cd/Ni in soil, we suggested that increased mobility of Cd/Ni by EDTA could enhance the potential of metal accumulation in the shoot. The result obtained in our study was consistent with the findings of others studies showing that the enhancement of HM availability in soils by the addition of EDTA improves metal phytoextraction (Liphadzi and Kirkham 2006; Van Engelen et al. 2007).
Antioxidant enzymes are considered to be the most important defense system toward oxidative stress caused by HMs (Weckx and Clijsters 1996). Activities of CAT, SOD, and APX are reported to increase under oxidative stress (Gill and Tuteja 2010). Furthermore, HMs are believed to cause oxidative damage to plants through the production of ROS, which cause damage to biomolecules, such as membrane lipids and proteins. Besides these, excess Cd and Ni indirectly induce oxidative stress by interrupting the equilibrium between ROS production and detoxification (Ishtiyaq et al. 2018). Under such conditions, scavenging of O2− by SOD and H2O2 decomposition by APX and CAT is primarily responsible for the maintenance of cellular redox state. APX activity exhibited maximum range of variation and showed a linear relationship with SOD (8.12 U/min/mg protein) and CAT (17.15 U/min/mg protein). APX is an indispensable component of the ascorbate–glutathione pathway required to scavenge H2O2 and to maintain the redox state of the cell (Tripathi et al. 2009). APX is mainly produced in the chloroplast of the plant cell, therefore, the higher activity of APX seems to help leaf cells to sustain their redox potential and decrease ROS production, thereby preventing PSII damage.
The bacterial SODs play an essential role in their survival in the rhizosphere by facilitating the removal of free radicals (Wang et al. 2007). On average, the activity of SOD increased in the plants when A. lentiformis was inoculated with PGPB1 and PGPB2. Plants showed up to 48.5 and 45.8% increase in the SOD activities (in the case of Cd) and up to 10.6 and 13% increase (for Ni), while plants co-inoculated with (Bradyrhizobium + Pseudomonas + OM) showed an increase up to 32.2 and 15.8% under Cd and Ni stress, respectively.
A variety of tolerance mechanisms have evolved in halophytic plants against HM ions, which allow plants to survive while accumulating high concentrations of HMs. (Baker and Walker 1990; Cobbett and Goldsbrough 2002). Moreover, effective phytoremediation depends on the bioavailability of HMs which can be greatly influenced by bioaugmentation of some metal-resistant bacteria. PGPB can simultaneously promote plant growth and accelerate the process of metal remediation. Saleh and Saleh (2006) used biological inoculation technology, on the host cowpea (Vigna sinensis) in pot cultures with Zn (0.0–1000 mg/kg dry soil) and Cd (0.0–100 mg/kg dry soil). They found that micro-symbionts significantly increased dry weight, root:shoot ratios, leaf number, and area, plant length, leaf pigments, total carbohydrates, and N and P content of infected plants as compared to non-infected controls at all levels of HM concentrations. This study revealed that all three amendments were able to enhance the metal uptake and accumulation in A. lentiformis and provided impetus to the physiology in a healthier way. A. lentiformis showed a significant decrease in growth parameters upon high dosage of Ni (100 ppm) and Cd (50 ppm) as HM-contaminated soils often show negative repercussions on plant growth and scarcity of nutrients (Wan et al. 2012).
The researchers also indicated that the mechanisms used by PGPB in the remediation of HM-contaminated soils may rely entirely on the species of PGPB and plant involved in the process. For example, the specific response of plant tissues toward HM accumulation under the influence of PGPB may be consistent with previous studies of lentil plants where the uptake of Ni and Zn was higher in the root as compared to shoot and grain (Wani et al. 2008). Likewise, another study found that inoculation with Brevi bacillus spp. decreased the Zn uptake in Trifolium repens (Vivas et al. 2006). Seyed et al. (2018) investigated the effect of siderophore-producing PGPB strains—Bacillus safensis FO-036b (T) and Pseudomonas fluorescens inoculation on Helianthus annuus (sunflower) growth and metals accumulation. In which PGPB inoculations solubilize and increased the Zn and Pb accumulation in test plant. Similarly, Amjad et al. (2017) and Vartika et al. (2016) reported that inoculation of siderophore-producing PGPR strains significantly increased the accumulation of Zn, Pb, and Fe in their respective host plants.
The results obtained in this work revealed that HMs with agronomic supplements such as (PGPB, OM, and EDTA) reduce the oxidative stress in A. lentiformis, mainly when plant inoculated with Bradyrhizobium japonicum (PGPB1) and Pseudomonas fluorescens (PGPB2) or co-inoculated with Bradyrhizobium japonicum and Pseudomonas fluorescens (Fig. 7). These symbiotic pairs are likely to possess a higher efficiency in their nodule antioxidative systems for maintaining lower nodule H2O2 levels against Cd/Ni stress (Rodrigues et al. 2013). Similar results have been reported in Trigonella foenum-graecum co-inoculated with Ensifer meliloti and Bacillus exposed to moderate and severe drought (Barnawal et al. 2013) and in cowpea plants co-inoculated with Bradyrhizobium, Puccinia graminis, and P. durus (Egamberdieva et al. 2013). Our results also suggest that Atriplex plants inoculated with Bradyrhizobium (PGPB1) and Pseudomonas (PGPB2) showed greater oxidative protection followed by OM and EDTA, respectively. An increase in CAT activity has been shown as a measure of antioxidant defense in halophytes (Lokhande et al. 2013).
The mechanisms used by PGPB in the remediation of HM-contaminated soils may rely entirely on the species of PGPB and plants involved in the process. Plant adaptation to HM-induced stress is controlled by cascades of molecular networks. Plant genomes contain a large number of genes with specific expression pattern in response to HM uptake and their transport to specific part. For every essential metal a particular gateway (metal transporter) is present for its entry in plant. This metal transporter allows only that particular metal ion or its close homolog to pass through it. However, due to the structural similarity of HMs with other essential nutrients, often, these HMs are transferred to root with the help of membrane transporter proteins. So far, several genes and gene families have been identified in plants, which play a key role in metal transportation and accumulation. Several metal transporters gene families have been identified in Arabidopsis thaliana which are directly or indirectly involved in HM transport and accumulation, namely ZIP (Zinc-regulated transporter—ZRT, Iron regulated transporter, IRT), Natural Resistance-Associated Macrophage Protein (NRAMP), ABC transporter, Cation Diffusion Facilitator (CDF), Heavy Metal ATPase (HMA), Cation proton exchanger (CAX), Heavy Metal ATPase (HMA), Low-affinity Cation Transporters (LCT), Copper transporter (COPT), and metal tolerance protein (MTP). (Nakanishi et al. 2006; Sasaki et al. 2012; Takahashi et al. 2012). IRT and NRAMP family genes are involved in the uptake of HMs to root from soil (Sasaki et al. 2012). HMA gene families and AtALS3 gene facilitates HM loading in shoot from root through xylem (Takahashi et al. 2012). In general, to cope up with HM toxicity, plants activate the metal assimilation pathway by increasing transcription of related genes.
PGPB-assisted phytoremediation studies conducted in the recent past concluded that PGPB inoculation influences the HM uptake in the plant by regulating the gene expression pattern of major metal transporter gene families (Ghassemi and Mostajeran 2018; Jebara et al. 2018). Further, PGPB inoculations play a vital role in the expression of growth and metabolic process-related genes expression, which systematize the plant’s growth (biomass, leaf surface area, and lateral root formation), physiology, and biochemical expressions (Ambreetha et al. 2018). Khanna et al. (2019) observed that inoculation of PGPR strains significantly enhance the antioxidant system of Lycopersicon esculentum (tomato) by up-regulating mRNA expression of SOD, POD, and PPO genes under Cd stress. In the present investigation, a similar generalization may be proposed for Atriplex to understand the response of HM transport and assimilation pathway under the influence of PGPB inoculation.
Saleh and Saleh (2006), used biological inoculation technology, on the host plant cowpea (Vigna sinensis) in pot cultures with Zn (0.0–1000 mg kg−1 dry soil) and Cd (0.0–100 mg kg−1 dry soil), and they found that micro-symbionts significantly increased dry weight, root:shoot ratios, leaf number, leaf area, plant length, leaf pigments, total carbohydrates, nitrogen, and phosphorus content of infected plants as compared to non-infected controls at all levels of HM concentrations. The present study revealed that all three amendments were able to enhance the metal uptake and accumulation in A. lentiformis and provide impetus to physiology in a healthier way. A. lentiformis showed a significant decrease in growth parameters upon high dosage of Ni100 ppm and Cd50 ppm as HM-contaminated soils often show negative repercussions on plant growth and scarcity of nutrients (Wan et al. 2012).
APX are indispensable components of the ascorbate–glutathione pathway required to scavenge H2O2 and are produced mainly in chloroplasts and maintain the redox state of the cell (Tripathi et al. 2009). Therefore, the higher activity of APX seems to help leaf cells to sustain their redox potential and decrease ROS production, thereby preventing PSII damage. Enhanced APX activity is mainly associated with an adaptive mechanism to increase the level of ROS content produced by HM exposure. Karthikeyan et al. (2007) reported an increase in the activities of antioxidant enzymes such as CAT, APX, and SOD due to the treatment with diazotrophic bacteria, such as Azospirillum and Azotobacter. These findings may be correlated with the present outcomes where inoculation with Pseudomonas fluorescens (PGPB2) and Bradyrhizobium japonicum (PGPB1) resulted in enhanced activities of antioxidative enzymes that might protect the photosynthetic machinery from oxidative damages and finally enhanced photosynthetic rate that results in increased biomass production.
An effective phytoremediation depends upon the bioavailability of HMs which can be greatly influenced by bioaugmentation of some metal-resistant bacteria. PGPB can simultaneously promote plant growth and accelerate the process of metal remediation. Use of Pseudomonas species for remediation of HMs like Pb, Cd, Ni, and Cr have been documented (Karimpour et al. 2018). Pseudomonas and Bradyrhizobium inoculum can increase the bioavailability and mobilization of HM (Dary et al 2010). The results of the current investigation suggest that a major benefaction of PGPB for better growth and tolerance of A. lentiformis was the stimulation of antioxidative enzymes which enabled the selected halophyte to overcome all the pernicious symptoms of HM accumulation. Within a range of salt concentrations optimal for growth, the sequestration of saline ions into the vacuoles results in increased succulence of the plant’s vegetative parts which is a common characteristic of the halophytes (commonly called as halosucculence) (Short and Colmer 1999). Succulence minimizes the toxic effect of excessive ion accumulation and is associated with accretion of osmotically active solutes for maintenance of cell turgor pressure (Luttge and Smith 1984).
The data on adaptability of the plant exposed to various abiotic factors reveal that A. lentiformis sustains its growth by sequestration of HMs into the vacuoles to maintain the osmotic balance between vacuole and cytoplasm. The exact physiological adaptation to HM stress and fate of accumulated metal in A. lentiformis whether it is subcellular localized or sequestered by metallothionine or phytochelatin or proline is yet be elucidated. However, the ability of this plant to take up Cd and Ni, from contaminated soils with its maximum uptake in roots followed by shoot makes A. lentiformis, a potential phytoremediator. The feature of A. lentiformis to accumulate high amount of HMs in its tissues may be exploited for reducing HM levels in the potential agriculture soil and in the arid and semi-arid regions by repetitive cultivation and harvesting of plant in these areas.
Conclusion
Plant growth-promoting bacteria like Bradyrhizobium japonicum and Pseudomonas fluorescens could be used successfully to promote plant growth, physiology, antioxidative defense, and uptake of metal from soil. Synergistic interactions between the consortia of Pseudomonas fluorescens, Bradyrhizobium japonicum, and OM with Atriplex lentiformis significantly improved the growth, antioxidative defense, and metal uptake than these PGPBs alone. Thus it can be concluded that inoculating the rhizosphere soils with selected metal-tolerant bacteria along with OM can be a sustainable, economical and eco-friendly option to elevate bioavailable metal concentration in the soil for plant uptake and thereby improving overall phytoremedial potential of the test plant. Activity of antioxidative enzymes was found to have a positive correlation on Ni and Cd uptake by A. lentiformis. Remediation of contaminated soils is a cumbersome and slow process that requires long periods of time to be effective. Therefore, direct use of contaminated sites with appropriate candidate species in association with biological amendments is likely to be more efficient method in order to remediate such lands. Consequently, the production of safe animal forages from contaminated soils was also one of the aims of this research especially in view of the stressed environment, sparse vegetation cover, and heavy load of biotic stress found therein. Atriplex species being equipped with excellent halophytic attributes and enormous forage potential can efficiently serve the purpose, especially in view of being identified as an efficient phytostabilizer. This process can be accelerated by implementing biological amendment-based phytoremediation, thus offering a self-sustaining and long-term solution for such habitats. Other species of Atriplex can also be screened along with different biological amendment combinations to further enhance the amplitude of phytoremediation prospects. Further, in spite of the information available on the physiological and biochemical basis of tolerance to HM stress, an intensive research needs to be focused toward understanding the molecular basis of metal tolerance in Atriplex, is warranted which could provide an additional resource for the improvement of HM stress tolerance in forage and other crops.
Data Availability
Not applicable.
References
Adekiya AO, Ejue WS, Olayanju A (2020) Different organic manure sources and NPK fertilizer on soil chemical properties, growth, yield and quality of okra. Sci Rep 10:16083
Aebi H (1984) Catalase in Vitro. Methods Enzymol 105:121–126
Agency for Toxic Substances and Disease Registry ATSDR (2019) Substance priority list. Atlanta, GA: U.S. Department of Health and Human Services, Public Health Service
Ahemad M (2019) Remediation of metalliferous soils through the heavy metal resistant plant growth promoting bacteria: paradigms and prospects. Arab J Chem 12(7):1365–1377. https://doi.org/10.1016/j.arabjc.2014.11.020
Ahmad B, Zaid A, Sadiq Y, Bashir S, Wani SH (2019) Role of selective exogenous elicitors in plant responses to abiotic stress tolerance. In: Hasanuzzaman M, Hakeem K, Nahar K, Alharby H (eds) Plant abiotic stress tolerance. Springer, Cham, pp 273–290. https://doi.org/10.1007/978-3-030-06118-0_12
Ahmad B, Zaid A, Zulfiqar F, Bovand F, Dar TA (2022) Nanotechnology: A novel and sustainable approach towards heavy metal stress alleviation in plants. Nanotechnol Environ Eng. https://doi.org/10.1007/s41204-022-00230-8
Al-Aqeel H, Vinod K (2016) Heavy metal uptake efficiency of Alfalfa, Barley, Indian Mustard and Atriplex from contaminated desert soil. Int J of Tropi Agri 34:2403–2406
Al-Dhabi NA, Esmail GA, Mohammed GAK, Valan AM (2019) Optimizing the management of cadmium bioremediation capacity of metal-resistant Pseudomonas sp. Strain Al-Dhabi-126 isolated from the industrial City of Saudi Arabian environment. Int J Environ Res Public Health 16:4788. https://doi.org/10.3390/ijerph16234788
Alsohim AS (2020) Influence of Pseudomonas fluorescens mutants produced by transposon mutagenesis on in vitro and in vivo biocontrol and plant growth promotion. Egypt J Biol Pest Control 30:19. https://doi.org/10.1186/s41938-020-00220-5
Ambreetha S, Chinnadurai C, Marimuthu P, Balachandar D (2018) Plant-associated Bacillus modulates the expression of auxin-responsive genes of rice and modifies the root architecture. Rhizosphere 5:57–66
Amjad A, Di G, Amanullah M, Fang M, Ronghua L, Feng S, Ping W, Zengqiang Z (2017) Streptomyces pactum assisted phytoremediation in Zn/Pb smelter contaminated soil of Feng County and its impact on enzymatic activities. Sci Rep 7:46087
Baker AJM, Walker PL (1990) Ecophysiology of metal uptake by tolerant plants, heavy metal tolerance in plants. In: Shaw AJ (ed) Evolutionary aspects. CRC, Boca Raton, pp 155–177
Barnawal D, Maji D, Bharti N, Chanotiya CS, Kalra A (2013) ACC deaminase-containing Bacillus subtilis reduces stress ethylene-induced damage and improves mycorrhizal colonization and rhizobial nodulation in Trigonella foenum-graecum under drought stress. J Plant Growth Regul 32:809–822. https://doi.org/10.1007/s00344-013-9347-3
Bates L, Waldren RP, Teare ID (1973) Rapid determination of free proline for water-stress studies. Plant Soil 39:205–207
Bulent T, Nuri A, Houcine A, Reguieg Y (2019) Phytoremediation potential of Atriplex canescens (Pursh) Nutt and Nicotiana tabacum grown in heavy metal contaminated soil. International Conference on Food, Nutrition and Agriculture (ICFNA19). Istanbul (Turkey). Conference Book.
Chellaiah E (2018) Cadmium (heavy metals) bioremediation by Pseudomonas aeruginosa: a minireview. Appl Water Sci 8:154
Cobbett C, Goldsbrough P (2002) Phytochelatins and metallothioneins, roles in heavy metal detoxification and homeostasis. Ann Rev Plant Biol 53:159–182
Cui LM, Wang YG, Gao L, Hu LH, Yan LG, Wei Q, Du B (2015) EDTA functionalized magnetic graphene oxide for removal of Pb(II), Hg(II), and Cu(II) in water treatment: adsorption mechanism and separation property. Chem Eng J 281:1–10
Cushman JC (2001) Osmoregulation in plants: implications for agriculture. Amer Zool 41:758–769. https://doi.org/10.1093/icb/41.4.758
Del Río LA, Sandalio LM, Altomare DA, Zilinskas BA (2003) Mitochondrial and peroxisomal manganese superoxide dismutase: Differential expression during leaf senescence. J Exp Bot 54:923–933. https://doi.org/10.1093/jxb/erg091
Dary M, Chamber-Pérez MA, Palomares AJ, Pajuelo E (2010) “In situ” phytostabilization of heavy metal polluted soils using Lupinus luteus inoculated with metal resistant plant-growth-promoting rhizobacteria. J Hazard Mater 177:323–330
Dhindsa RS, Plumb-Dhindsa P, Thorpe TA (1981) Leaf senescence correlated with increased levels of membrane permeability and lipid peroxidation and decreased levels of superoxide dismutase and catalase. J Exp Bot 32:93–101
Dutta P, Karmakar A, Majumdar S, Roy S (2018) Klebsiella pneumonia (HR1) assisted alleviation of Cd (II) toxicity in Vigna mungo: a case study of biosorption of heavy metal by an endophytic bacterium coupled with plant growth promotion. Euro-Medi J Environ Integ 27:1–10. https://doi.org/10.1007/s41207-018-0069-6
Egamberdieva D, Berg G, Lindström K, Räsänen LA (2013) Alleviation of salt stress of symbiotic Galega officinalis L (Goat’s Rue) by co-inoculation of rhizobium with root colonizing Pseudomonas. Plant Soil 369:453–546
Egamberdieva D, Wirth S, Jabborova D, Leena AR, Liao H (2017) Coordination between Bradyrhizobium and Pseudomonas alleviates salt stress in soybean through altering root system architecture. J Plant Interact 12:100–107. https://doi.org/10.1080/17429145.2017.1294212
Errabii T, Gandanou CB, Essalmani H, Abrini J, Idamor M, Senhaji NS (2007) Effect of NaCl and mannitol induced stress on sugarcane (Saccharum sp.) callus cultures. Acta Physiol Plant 29:95–102. https://doi.org/10.1007/s11738-006-0006-1
Flowers TJ, Colmer TD (2008) Salinity tolerance in halophytes. New Phytol 179:945–963. https://doi.org/10.1111/j.1469-8137.2008.02531.x
Garnier L, Françoise SP, Patrice T, Jean-Pierre A, Jean-pierre B, Raoul R, Jean-luc M (2006) Cadmium affects tobacco cells by a series of three waves of reactive oxygen species that contribute to cytotoxicity. Plant Cell and Environ 29:1956–1969. https://doi.org/10.1111/j.1365-3040.2006.01571.x
Ghassemi HR, Mostajeran A (2018) TASOS1 and TATM20 genes expression and nutrient uptake in wheat seedlings may be altered via excess cadmium exposure and inoculation with Azospirillum brasilense sp7 under saline condition. Appl Ecol Environ Res 16:1797–1817
Gill SS, Tuteja N (2010) Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol Biochem 48:909–930
Gopal R, Rizvi AH (2008) Excess lead alters growth, metabolism, and translocation of certain nutrients in radish. Chemosphere 70:1539–1544. https://doi.org/10.1016/j.chemosphere.2007.08.043
Gorden SA, Paleg LG (1957) Observations on the quantitative determination of indole acetic acid. Physiol Plant 10:39–47. https://doi.org/10.1111/j.1399-3054.1957.tb07608.x
Gratão PL, Monteiro CC, Rossi ML, Martinelli AP, Peres LEP, Medici LO, Lea PJ, Azevedo RA (2009) Differential ultrastructural changes in tomato hormonal mutants exposed to cadmium. Environ Exp Bot 67:387–394. https://doi.org/10.1016/j.envexpbot.2009.06.017
Gupta P, Rani R, Chandra A, Kumar V (2018) Potential applications of Pseudomonas sp. (strain CPSB21) to ameliorate Cr6þ stress and phytoremediation of tannery effluent contaminated agricultural soils. Sci Rep 8:4860
Gupta S, Pandey S (2019) ACC deaminase producing bacteria with multifarious plant growth promoting traits alleviates salinity stress in french bean (Phaseolus vulgaris) plants. Front Microbiol 10:1506. https://doi.org/10.3389/fmicb.2019.01506
HazDat (2006) HazDat database: ATSDR’s Hazardous Substance Release and Health Effects Database. Atlanta, GA: Agency for Toxic Substances and Disease Registry. www.atsdr.cdc.gov/hazdat.html.
Heath RL, Packer L (1968) Photoperoxidation in isolated chloroplasts I-Kinetics and stoichiometry of fatty acid peroxidation. Arch Biochem Biophys 125:189–198
Honma M, Shimomura T (1978) Metabolism of 1-aminocyclopropane-1-carboxylic acid. Agr Biol Chem 42:1825–1831. https://doi.org/10.1080/00021369.1978.10863261
Ishtiyaq S, Kumar H, Varun M, Kumar B, Paul MS (2018) Heavy metal toxicity and antioxidative response in plants: an overview: responses, tolerance, and remediation. In: Hasanuzzaman M, Nahar K, Fujita M (eds) Plants under metal and metalloid stress. E-Publishing Inc. Springer Intern Publishing, Switzerland, pp 77–106
Ishtiyaq S, Kumar H, Clement OO, Varun M, Paul MS (2021) Role of secondary metabolites in salt and heavy metal stress mitigation by halophytic plants: An overview. In: Hasanuzzaman M, Prasad MNV (eds) Handbook of bioremediation. E-Publishing Inc. Academic Press, Cambridge, pp 307–321
Islam F, Yasmeen T, Ali Q, Ali S, Arif MS, Hussain S, Rizvi H (2014) Influence of Pseudomonas aeruginosa as PGPR on oxidative stress tolerance in wheat under Zn stress. Ecotoxicol Environ Saf 104:285–293. https://doi.org/10.1016/j.ecoenv.2014.03.008
Jahanian A, Chaichi MR, Rezaei K, Rezayazdi K, Khavazi K (2012) The effect of plant growth-promoting rhizobacteria (PGPR) on germination and primary growth of Artichoke (Cynara scolymus). Int J Agric Crop Sci 4:923–929
Jebara SH, Chiboub M, Jebara M (2018) Antioxidant responses and gene level expressions of Sulla coronaria inoculated by heavy metals resistant plant growth promoting bacteria under cadmium stress. In: Kallel A et al (eds) Recent advances in environmental science from the euro-mediterranean and surrounding regions, advances in science, technology & innovation. Springer International Publishing, Berlin. https://doi.org/10.1007/978-3-319-70548-4_106,2018
Kang SM, Shahzad R, Bilal S, Khan AL, Park YG, Lee KE, Lee IJ (2019) Indole-3-acetic-acid and ACC deaminase producing Leclercia adecarboxylata MO1 improves Solanum lycopersicum L. growth and salinity stress tolerance by endogenous secondary metabolites regulation. BMC Microbiol 19(1):80
Karimpour M, Ashraf SD, Taghavi K, Mojtahedi A, Roohbakhsh E, Naghipour D (2018) Adsorption of cadmium and lead onto live and dead cell mass of Pseudomonas aeruginosa: a dataset. Data Brief 18:1185–1192. https://doi.org/10.1016/j.dib.2018.04.014
Karthik C, Oves M, Thangabalu R, Sharma R, Santhosh SB, Arulselvi PI (2016) Cellulosimicrobium funkei-like enhances the growth of Phaseolus vulgaris by modulating oxidative damage under chromium (VI) toxicity. J Adv Res 7:839–850
Karthikeyan B, Jaleel CA, Gopi R, Deiveekasundaram M (2007) Alterations in seedling vigour and antioxidant enzyme activities in Catharanthus roseus under seed priming with native diazotrophs. J Zhejiang Univ Sci 8:453–457. https://doi.org/10.1631/jzus.2007.B0453
Katznelson H, Bose B (1959) Metabolic activity and phosphate-dissolving capability of bacterial isolates from wheat roots, rhizosphere, and non-rhizosphere soil. Can J Microbiol 5:79–85. https://doi.org/10.1139/m59-010
Kelvin RP, Víctor MC, Julio CAO, Warren RR (2020) Bioavailability and solubility of heavy metals and trace elements during composting of cow manure and tree litter. Appl Envir Soil Sci. https://doi.org/10.1155/2020/5680169
Khanna K, Jamwal VL, Kohli SK, Gandhi SG, Ohri P, Bhardwaj R, Allah EFA, Hashem A, Ahmad P (2019) Plant growth promoting rhizobacteria induced Cd tolerance in Lycopersicon esculentum through altered antioxidative defense expression. Chemosphere 217:463–474. https://doi.org/10.1016/j.chemosphere.2018.11.005
Kong Z, Glick BR (2017) The role of plant growth-promoting bacteria in metal phytoremediation. Adv Microb Physiol 71:97–132. https://doi.org/10.1016/bs.ampbs.2017.04.001
Kubier A, Wilkin RT, Pichler T (2019) Cadmium in soils and groundwater: a review. Appl Geochem 108:10438. https://doi.org/10.1016/j.apgeochem.2019.104388
Lichtenthaler HK (1987) Chlorophylls and carotenoids: pigments of photosynthetic biomembranes. Meth Enzymol 148:350–382. https://doi.org/10.1016/0076-6879(87)48036-1
Liphadzi MS, Kirkham MB (2006) Availability and plant uptake of heavy metals in EDTA-assisted phytoremediation of soil and composted biosolids. S Afr J Bot 72:391–397
Liu X, Wang C, Su Q (2013) Screening for salt tolerance in eight halophyte species from yellow river delta at the two initial growth stages. ISRN Agronomy 1:1–8. https://doi.org/10.1155/2013/592820
Lokhande VH, Nikam TD, Suprasanna P (2010) Differential osmotic adjustment to iso-osmotic salt and PEG stress in vitro in the halophyte Sesuvium portulacastrum L. J Crop Sci Biotechnol 13(4):251–256. https://doi.org/10.1007/s12892-010-0008-9
Lokhande VH, Mulye K, Patkar R, Nikam TD, Suprasanna P (2013) Biochemical and physiological adaptations of the halophyte Sesuvium portulacastrum L (Aizoaceae) to salinity. Arch Agron Soil Sci 59:1373–1391. https://doi.org/10.1080/03650340.2012.712207
Luttge U, Smith JAC (1984) Structural, biophysical, and biochemical aspects of the role of leaves in plant adaptation to salinity and water stress. In: Staples RC, Toenniessen GH (eds) Salinity tolerance in plants: strategies for crop improvement. Wiley, New York, pp 125–150
Ma Y, Oliveira RS, Freitas H, Zhang C (2016) Biochemical and molecular mechanisms of plant-microbe-metal interactions: relevance for phytoremediation. Front Plant Sci 7:918. https://doi.org/10.3389/fpls.2016.00918
Maria BGSF, Luana CC, Andreza RBF, Júlia KS, Karen CFS (2010) Isolation of Atriplex nummularia-associated halotolerant bacteria and bioprospecting by nitrogen-fixing bacteria in saline-sodic soil. 19th World Congress of Soil Science, Soil Solutions for a Changing World, Brisbane, Australia. pp. 1–4
Maxton A, Singh P, Masih SA (2018) ACC deaminase-producing bacteria mediated drought and salt tolerance in Capsicum annuum. J Plant Nutr 41(5):574–583
Mesa-Marín J, Pérez-Romero JA, Redondo-Gómez S, Pajuelo E, Rodríguez-Llorente ID, Mateos-Naranjo E (2020) Impact of plant growth promoting bacteria on Salicornia ramosissima ecophysiology and heavy metal phytoremediation capacity in estuarine soils. Frontiers in Microbiol. https://doi.org/10.3389/fmicb.2020.553018
Meyer R (2005) Atriplex lentiformis. In: Fire Effects Information System [Online] U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory (Producer). http://www.fs.fed.us/database/feis.
Nakanishi H, Ogawa I, Ishimaru Y, Mori S, Nishizawa NK (2006) Iron deficiency enhances cadmium uptake and translocation mediated by the Fe2+ transporters OsIRT1 and OsIRT2 in rice. Soil Sci Plant Nutr 52:464–469
Nakano Y, Asada K (1987) Purification of ascorbate peroxidase in spinach chloroplasts: its inactivation in ascorbate depleted medium and reactivation by monodehydroascorbate radical. Plant Cell Physiol 28:131–140. https://doi.org/10.1093/oxfordjournals.pcp.a077268
Nascimento CWA, Amarasiriwardena D, Xing B (2006) Comparison of natural organic acids and synthetic chelates at enhancing phytoextraction of metals from a multi-metal contaminated soil. Environ Pollut 140:114–123
Nazar R, Iqbal N, Masood A, Khan MIR, Syeed S, Khan NA (2012) Cadmium toxicity in plants and role of mineral nutrients in its alleviation. Am J Plant Sci 3:1476–1489. https://doi.org/10.4236/ajps.2012.310178
Nazir F, Hussain A, Fariduddin Q (2019) Interactive role of epibrassinolide and hydrogen peroxide in regulating stomatal physiology, root morphology, photosynthetic and growth traits in Solanum lycopersicum L. under nickel stress. Environ Exp Bot 162:479–495
Olsen SR, Sommers LE (1982) Determination of available phosphorus. In: Page AL, Miller RH, Keeney DR (eds) Methods of soil analysis. E-Publishing Inc. American Society of Agronomy, Madison, pp 403–430
Orozco-Mosqueda MC, Glick BR, Santoyo G (2020) ACC deaminase in plant growth-promoting bacteria (PGPB): an efficient mechanism to counter salt stress in crops. Microbiol Res 235:126439. https://doi.org/10.1016/j.micres.2020.126439
Penrose DM, Glick BR (2003) Methods for isolating and characterizing ACC deaminase containing plant growth-promoting rhizobacteria. Plant Physiol 118:10–15. https://doi.org/10.1034/j.1399-3054.2003.00086.x
Piccini D, Malavolta E (1992) Effect of nickel on two common bean cultivars. J Plant Nut 15:2343–2350. https://doi.org/10.1080/01904169209364478
Rao KVM, Sresty TVS (2000) Antioxidative parameters in the seedlings of pigeon pea (Cajanus cajan L.) Millspauga in response to Zn and Ni stress. Plant Sci 157:113–128. https://doi.org/10.1016/s0168-9452(00)00273-9
Rizvi A, Ahmed B, Zaidi A, Khan MS (2019) Bioreduction of toxicity influenced by bioactive molecules secreted under metal stress by Azotobacter chroococcum. Ecotoxicol 28:302–322. https://doi.org/10.1007/s10646-019-02023-3
Rizwan M, Ali S, Abbas T, Rehman MZ, Hannan F, Keller C, Al-Wabel MI, Ok YS (2016) Cadmium minimization in wheat: a critical review. Ecotoxicol Environ Safety 130:43–53. https://doi.org/10.1016/j.ecoenv.2016.04.001
Rodrigues AC, Bonifacio A, Antunes JEL, Silveira JAG, Figueiredo MVB (2013) Minimization of oxidative stress in cowpea nodules by the interrelationship between Bradyrhizobium spp and plant growth-promoting bacteria. Appl Soil Ecol 64:245–251. https://doi.org/10.1016/j.apsoil.2012.12.018
Rowe DR, Abdel-Magid IM (1995) HandBook of waste water reclamation. CRC Press Inc, Boca Raton, p 550
Saitou N, Nei M (1987) The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol Biol Evol 4:406–425. https://doi.org/10.1093/oxfordjournals.molbev.a040454
Saleh M, Saleh AG (2006) Increased heavy metal tolerance of cowpea plants by dual inoculation of an arbuscular mycorrhizal fungi and nitrogen-fixer Rhizobium bacterium. Afr J Biotechnol 5:133–142
Sasaki A, Yamaji N, Yokosho K, Ma JF (2012) Nramp5 is a major transporter responsible for manganese and cadmium uptake in rice. Plant Cell 24:2155–2167
Schwyn B, Neilands JB (1987) Universal chemical assay for the detection and determination of siderophores. Anal Biochem 160:47–56. https://doi.org/10.1016/0003-2697(87)90612-9
Seneviratne M, Gunaratne S, Bandara T, Weerasundara L, Rajakaruna N, Seneviratne G, Vithanage M (2016) Plant growth promotion by Bradyrhizobium japonicum under heavy metal stress. S Afri J of Bot 105:19–24. https://doi.org/10.1016/j.sajb.2016.02.206
Seniyat LA, Lesley CB (2019) Effect of plant growth-promoting bacterium; Pseudomonas putida UW4 inoculation on phytoremediation efficacy of monoculture and mixed culture of selected plant species for PAH and lead spiked soils. Int J of Phytorem 21:200–208. https://doi.org/10.1080/15226514.2018.1501334
Seyed MM, Babak M, Hossein MH, Hoseinali A, Ali AZ (2018) Root-induced changes of Zn and Pb dynamics in the rhizosphere of sunflower with different plant growth promoting treatments in a heavily contaminated soil. Ecotoxicol Environ Saf 147:206–216
Short DC, Colmer TD (1999) Salt tolerance in the halophyte Halosarcia pergranulata subsp. pergranulata. Annu Bot 83:207–213. https://doi.org/10.1006/anbo.1998.0812
Slama I, Ghnaya T, Savouŕe A, Abdelly C (2008) Combined effects of long-term salinity and soil drying on growth, water relations, nutrient status and proline accumulation of Sesuvium portulacastrum. C R Biol 331:442–451. https://doi.org/10.1016/j.crvi.2008.03.006
Slama I, Abdelly C, Bouchereau A, Flowers T, Savoure A (2015) Diversity, distribution, and roles of osmoprotective compounds accumulated in halophytes under abiotic stress. Ann Bot 115:433–447. https://doi.org/10.1093/aob/mcu239
Soliz D, Edward G, Seaman RB, Yoklic M, Nelson SG, Brown P (2011) Water consumption, irrigation efficiency and nutritional value of Atriplex lentiformis grown on reverse osmosis brine in a desert irrigation district. Agric Ecosyst Environ 140:473–483. https://doi.org/10.1016/j.agee.2011.01.012
Suprasanna P, Rai AN, Kumari HP, Kumar SA, Kishor KPB (2014) Modulation of proline: implications in plant stress tolerance and development. In: Anjum NA, Gill SS, Gill R (eds) Plant adaptation to environmental change. CABI Publishers, Wallingford, pp 68–93
Szabados L, Savoure A (2009) Proline: a multifunctional amino acid. Trends Plant Sci 15:89–97. https://doi.org/10.1016/j.tplants.2009.11.009
Szopiński M, Sitko K, Gieroń Ż, Rusinowski S, Corso M, Hermans C, Verbruggen N, Małkowski E (2019) Toxic effects of Cd and Zn on the photosynthetic apparatus of the Arabidopsis halleri and Arabidopsis arenosa pseudo-metallophytes. Front Plant Sci 10:748. https://doi.org/10.3389/fpls.2019.00748
Takahashi R, Shimaru Y, Shimo H, Ogo Y, Senoura T, Nishizawa NK, Nakanishi H (2012) The OsHMA2 transporter is involved in root-to-shoot translocation of Zn and Cd in rice plant. Cell Environ 35:1948–1957
Tamura K, Stecher G, Peterson D, Filipski A, Kumar S (2013) MEGA6: molecular evolutionary genetics analysis version 6.0. Mol Biol Evol 30:2725–2729
Tang X, Zeng G, Fan G, Zhou M, Tang L, Zhu J, Wan J, Huang D, Chen M, Xu P, Zhang C, Lu Y, Xiong W (2018) Chromosomal expression of CadR on Pseudomonas aeruginosa for the removal of Cd (II) from aqueous solutions. Sci Tot Environ 636:1335–1361. https://doi.org/10.1016/j.scitotenv.2018.04.229
Tank N, Saraf M (2009) Enhancement of plant growth and decontamination of nickel spiked soil using PGPR. J Basic Microbiol 49:195–204. https://doi.org/10.1002/jobm.200800090
Toth G, Hermann T, Da Silva MR, Montanarella L (2016) Heavy metals in agricultural soils of the European Union with implications for food safety. Environ Pollution 88:299–309. https://doi.org/10.1016/j.envint.2015.12.017
Treesubsuntorn C, Dhurakit P, Khaksar G, Thiravetyan P (2018) Effect of microorganisms on reducing cadmium uptake and toxicity in rice (Oryza sativa L.). Environ Sci Pollut Res 25:25690–25701
Tripathi RP, Singh B, Bisht SS, Pandey J (2009) L-Ascorbic acid in organic synthesis: an overview. Curr Org Chem 13:99–122. https://doi.org/10.2174/13852720978719379.2
Van Engelen DL, Sharpe-Pedler RC, Moorhead KK (2007) Effect of chelating agents and solubility of cadmium complexes on uptake from soil by Brassica juncea. Chemosphere 68:401–408
Vartika M, Antriksh G, Parvinder G, Simranjeet S, Nasib S, Praveen G, Joginder S (2016) Synergistic effects of Arbuscular mycorrhizal fungi and plant growth promoting rhizobacteria in bioremediation of iron contaminated soils. Int J Phytorem 18:697–703
Vivas A, Biro B, Ruiz-Lozano JM, Barea JM, Azcon R (2006) Two bacterial strains isolated from a Zn-polluted soil enhance plant growth and mycorrhizal efficiency under Zn toxicity. Chemosphere 52:1523–1533
Wan Y, Luo S, Chen J, Xiao X, Chen L, Zeng G, Liu C, He Y (2012) Effect of endophyte-infection on growth parameters and Cd-induced phytotoxicity of Cd-hyperaccumulator Solanum nigrum L. Chemosphere 89:743–750. https://doi.org/10.1016/j.chemosphere.2012.07.005
Wang LW, Showalter AM (2004) Cloning and salt-induced, ABA independent expression of choline mono-oxygenase in Atriplex prostrate. Physiol Plant 120:405–412. https://doi.org/10.1111/j.0031-93172004.00247.x
Wang Y, Wang H, Yang CH, Wang Q, Mei R (2007) Two distinct manganese-containing superoxide dismutase genes in Bacillus cereus: Their physiological characterizations and roles in surviving in wheat rhizosphere. FEMS Microbiol Lett 272:206–213. https://doi.org/10.1111/j.1574-6968.2007.00759.x
Wani P, Khan M, Zaidi A (2008) Effect of metal-tolerant plant growth-promoting Rhizobium on the performance of pea grown in metal amended soil. Ach Environ Contam Toxicol App Pharmacol 55:33–42. https://doi.org/10.1007/s00244-007-9097-y
Wani W, Masoodi KZ, Zaid A, Wani SH, Shah F, Meena VS, Wani SA, Mosa KA (2018) Engineering plants for heavy metal stress tolerance. Rendiconti Lincei. Scienze Fisiche e Naturali. 29:709–723
Weckx JEJ, Clijsters HMM (1996) Oxidative damage and defense mechanisms in primary leaves of Phaseolus vulgaris as a result of root assimilation of toxic amounts of copper. Physiol Plant 96:506–512
Weyens N, Van der Lelie D, Taghavi S, Newman L, Vangronsveld J (2009) Exploiting plant-microbe partnerships to improve biomass production and remediation. Trends Biotechnol 27:591–598. https://doi.org/10.1016/j.tibtech.2009.07.006
Whiting SN, de Souza MP, Terry N (2001) Rhizosphere bacteria mobilize Zn for hyperaccumulation by Thlaspi caerulescens. Envirn Sci and Technol 35:3144–3150. https://doi.org/10.1021/es001938v
Wiangkham N, Prapagdee B (2018) Potential of Napier grass with cadmium-resistant bacterial inoculation on cadmium phytoremediation and its possibility to use as biomass fuel. Chemosphere 201:511–518. https://doi.org/10.1016/j.chemosphere.2018.03.039
Woldetsadik D, Drechsel P, Keraita B, Itanna F, Gebrekidan H (2017) Heavy metal accumulation and health risk assessment in wastewater-irrigated urban vegetable farming sites of Addis Ababa. Ethiopia Int J Food Contam 4:9. https://doi.org/10.1186/s40550-017-0053-y
Yokoi S, Bressan RA, Hasegawa PM (2002) Genetic engineering of crop plants for abiotic stress. In: Iwanaga M (ed) Salt stress tolerance of plants, The Japan International Centre for Agricultural Sciences (JIRCAS) Working Report No. 23, Tsukuba: Japan International Centre for Agricultural Sciences Publishing, Japan, pp 25–33
Yu X, Li Y, Zu C, Cui Y, Xiang Q, Gu Y, Zhau K, Zhang X, Penttinen P, Chen Q (2017) Pongamia pinnata inoculated with Bradyrhizobium liaoningense PZHK1 shows potential for phytoremediation of mine tailings. App Microbiol Biotechnol 101:1739–1751. https://doi.org/10.1007/s00253-016-7996-4
Zaidi S, Usmani S, Singh BR, Musarrat J (2006) Significance of Bacillus subtilis strain SJ-101 as a bioinoculant for concurrent plant growth promotion and nickel accumulation in Brassica juncea. Chemosphere 64:991–997
Zaier H, Ghnaya T, Lakhdar A, Baioui R, Ghabriche R, Mnasri M, Sghair S, Lutts S, Abdelly C (2010) Comparative study of Pb phytoextraction potential in Sesuvium portulacastrum and Brassica juncea: tolerance and accumulation. J Hazard Mater 183:609–615
Zaid A, Mohammad F, Fariduddin Q (2019a) Plant growth regulators improve growth, photosynthesis, mineral nutrient and antioxidant system under cadmium stress in menthol mint (Mentha arvensis L.). Physiol Mol Bio Plants 26:25–39
Zaid A, Mohammad F, Wani SH, Kadambot MH, Siddique, (2019b) Salicylic acid enhances nickel stress tolerance by up-regulating antioxidant defense and glyoxalase systems in mustard plants. Ecotoxicol Environ Saf 180:575–587
Zaid A, Wani SH (2019) Reactive oxygen species generation, scavenging and signaling in plant defense responses. In: Jogaiah S, Abdelrahman M (eds) Bioactive molecules in plant defense signaling in growth and stress. Springer, Cham, pp 111–132. https://doi.org/10.1007/978-3-030-27165-7_7
Zaid A, Bhat ZA, Wani SH (2020) Influence of Metalloids and Their Toxicity Impact on Photosynthetic Parameters of Plants. In: Deshmukh R, Tripathi DK, Guerriero G (eds) Metalloids in Plants: Advances and Future Prospects. John Wiley, New York
Zaid A, Mohammad F, Siddique HM (2022) Salicylic acid priming regulates stomatal conductance, trichome density and improves cadmium stress tolerance in Mentha arvensis L. Front Plant Sci. https://doi.org/10.3389/fpls.2022.895427
Zeffa DM, Fantin LH, Koltun A, Andre LMO, Maria PBAN, Marcelo CG, Leandro SAG (2020) Effects of plant growth-promoting rhizobacteria on co-inoculation with Bradyrhizobium in soybean crop: a meta-analysis of studies from 1987 to 2018. Peer J 8:7905. https://doi.org/10.7717/peerj.7905
Zhang JS, Xie C, Li ZY, Chen SY (1999) Expression of the plasma membrane H+ ATPase gene in response to salt stress in a rice salt tolerant mutant and its original variety. Theoret Appl Genet 99:1006–1011
Acknowledgements
This study had the support of Fundação para a Ciência e Tecnologia (FCT), through the strategic Project UIDB/04292/2020 granted to MARE. We also thank University Grants Commission, New Delhi for providing funding (Grant sanction F. no. 43-100/2014 SR) for the work. Harsh Kumar is grateful for the fellowship under the project. Authors are thankful to the Principal, St. John’s College, Agra, India for providing the necessary facilities for conducting this research.
Funding
This study had the support of Fundação para a Ciência e Tecnologia (FCT), through the strategic Project UIDB/04292/2020 granted to MARE and also the University Grants Commission, New Delhi for providing funding (Grant sanction F. no. 43-100/2014 SR) for the work.
Author information
Authors and Affiliations
Contributions
HK conceptualized the study and designed the methodology of the experiment. HK, SI, and MV carried out the experiment, monitor the growth stages, and collected data. HK, SI, and MV were involved in the data analysis and drafting of manuscript. MV analyzed the data statistically and interpreted the results. PJCF and MSP supervised the overall study and reviewed the manuscript draft before submission.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Ethical Approval
The study has no ethical issue problem related to any person or organization.
Consent to Participate
Not applicable.
Consent for Publication
The author agreed freely to publish the present work in this journal.
Additional information
Handling Editor: M. Iqbal R. Khan.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Kumar, H., Ishtiyaq, S., Favas, P.J.C. et al. Effect of Metal-resistant PGPB on the Metal Uptake, Antioxidative Defense, Physiology, and Growth of Atriplex lentiformis (Torr.) S.Wats. in Soil Contaminated with Cadmium and Nickel. J Plant Growth Regul 42, 3868–3887 (2023). https://doi.org/10.1007/s00344-022-10853-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00344-022-10853-5