
Abstract
Halophilic archaeal strain YC89T was isolated from Yuncheng salt lake in Shanxi, China. Cells from strain YC89T were short rods, lysed in distilled water, stained Gram-negative and formed red-pigmented colonies on agar plate. Strain YC89T was able to grow at 25–50 °C (optimum 37 °C), at 1.4–4.8 M NaCl (optimum 2.6–3.1 M), at 0–1.0 M MgCl2 (optimum 0.3 M) and at pH 6.0–9.5 (optimum pH 7.5). The major polar lipids are phosphatidylglycerol, phosphatidylglycerol phosphate methyl ester, sulfated mannosyl glucosyl diether and two unknown glycolipids. 16S rRNA gene analysis revealed that strain YC89T was phylogenetically related to Halorientalis persicus D108T (95.6 % nucleotide identity) and H. regularis TNN28T (95.3 % nucleotide identity). The rpoB′ gene similarities between strain YC89T and H. persicus IBRC-M 10043T and H. regularis TNN28T were 88.1 and 88.0 %, respectively. The DNA G+C content of strain YC89T was determined to be 61.3 mol%. The phenotypic, chemotaxonomic and phylogenetic properties suggested that strain YC89T (=CGMCC 1.12125T = JCM 18366T) represents a new species of Halorientalis, for which the name H. brevis sp. nov. is proposed.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Salt lakes are natural athalassohaline environments hosting diverse halophilic archaea, members of the family Halobacteriaceae within the order Halobacteriales [1, 7, 12, 15, 16]. The family Halobacteriaceae encompasses 47 genera containing over 165 species as of February 2014 [13, 14]. During our survey on halophilic archaeal diversity of an inland salt lake of China, we obtained a halophilic archaeal strain YC89T, which was most closely related to the members of Halorientalis, as judged from 16S rRNA gene sequences. The genus Halorientalis was first proposed to accommodate the species Halorientalis regularis based on two strains isolated from an artificial marine solar saltern in Eastern China [4]. Recently, H. persicus was described based on the strain isolated from an inland salt lake of China [1]. The current two members of Halorientalis are aerobic and chemoorganotrophic, using sugars as sources of carbon and energy. The major polar lipids of Halorientalis species are phosphatidylglycerol (PG), phosphatidylglycerol phosphate methyl ester (PGP-Me), sulfated mannosyl glucosyl diether (S-DGD-1) and several unknown glycolipids. In this study, we characterize strain YC89T as a new species of the genus Halorientalis, for which the name H. brevis sp. nov. is proposed.
Materials and Methods
Isolation and Cultivation of Halophilic Archaeal Strain
Strain YC89T was isolated from the brine sampled from Yuncheng salt lake at Yuncheng, Shanxi Province, China (35°00′14″N, 111°00′19″E; elevation, 323 m above sea level) and stored at 4 °C during transport to the laboratory in 2010. The pH of the brine was 7.9 and the salinity 235.6 g/L. The neutral haloarchaeal medium (NHM) was used for the isolation procedure and contained the following ingredients (g/L): yeast extract (Oxoid) 0.05, fish peptone (Sinopharm Chemical Reagent Co., Ltd.) 0.25, sodium pyruvate 1.0, KCl 5.4, K2HPO4 0.3, CaCl2 0.29, NH4Cl 0.27, MgSO4·7H2O 26.8, MgCl2·6H2O 23.0, NaCl 184.0 (pH adjusted to 7.0–7.2 with 1 M NaOH solution). The brine was serially diluted in liquid NHM medium and spread onto NHM agar plates. The inoculated plates were incubated for 3 months at 37 °C. After this initial cultivation, colonies were successively re-streaked on NHM agar plates at least three times to obtain pure colonies. The strains were routinely grown aerobically at 37 °C for 7 days in NHM and preserved at −20 °C as a suspension in NHM broth supplemented with glycerol (15 %, w/v).
Phenotypic Determination
Phenotypic tests were performed according to the proposed minimal standards for description of novel taxa in the order Halobacteriales [11]. Determination of morphology and growth characteristics, nutrition, miscellaneous biochemical tests and sensitivity to antimicrobial agents was performed as described and cited previously [3]. The halophilic archaeal strains H. regularis CGMCC 1.10123T and H. persicus IBRC-M 10043T were selected as reference strains. These reference strains were routinely grown aerobically at 37 °C in NHM medium.
Chemotaxonomic Characterization
Polar lipids were extracted using a chloroform/methanol system and analysed using one- and two-dimensional TLC, as described previously [3]. Merck silica gel 60 F254 aluminium-backed thin-layer plates were used for TLC analyses. In two-dimensional TLC, the first solvent was chloroform:methanol:water (65:25:4, by vol.), and the second solvent was chloroform:methanol:acetic acid:water (80:12:15:4, by vol.). The latter solvent mixture was also used for one-dimensional TLC. Two specific detection spray reagents, phosphate stain reagent for phospholipids and α-naphthol stain for glycolipids, were used. The general detection reagent, sulphuric acid:ethanol (1:2, by vol.), was also used to detect total polar lipids. The presence of phospholipids and glycolipids on the two-dimensional TLC was confirmed by comparing with one-dimensional TLC on which the polar lipid profile of reference strains was developed.
Phylogenetic and Genomic Analysis
Genomic DNA from halophilic archaeal strain was prepared as described previously for determination of the DNA base composition and PCR-mediated amplification experiments [5]. The 16S rRNA genes were amplified, cloned and sequenced according to the previous protocol [2]. PCR-mediated amplification and sequencing of the rpoB′ genes were performed according to Minegishi et al. [10]. Multiple sequence alignments were performed using the ClustalW program integrated in the MEGA 5 software [17]. Phylogenetic trees were reconstructed using the Maximum-Likelihood algorithm in the MEGA 5 software. Gene sequence similarity among halophilic archaea was calculated using the Pairwise-Distance computing function of MEGA 5. The DNA G+C content was determined from the mid-point value (T m) of the thermal denaturation method [8] at 260 nm with a Beckman-Coulter DU800™ spectrophotometer equipped with a high-performance temperature controller.
Results and Discussion
Morphological, Physiological and Biochemical Characteristics
Cells of strain YC89T were observed to be motile and short rod-shaped (0.5–1.0 × 1.0–2.5) when grown in NHM liquid medium (Supplementary Fig. S1). They stained Gram-negative, and the colonies were observed to be red-pigmented. Strain YC89T was able to grow at 25–50 °C (optimum 37 °C), at 1.4–4.8 M NaCl (optimum 2.6–3.1 M), at 0–1.0 M MgCl2 (optimum 0.3 M) and at pH 6.0–9.5 (optimum pH 7.5). The cells lysed in distilled water, and the minimal NaCl concentration to prevent cell-lysis was found to be 8 % (w/v). The strain was able to grow under anaerobic conditions using nitrate, DMSO and l-arginine. It was found to be positive for H2S formation and negative for indole formation. Strain YC89T did not hydrolyse starch, casein, gelatin or Tween 80. Strain YC89T was sensitive to the following antimicrobial compounds (µg per disc, unless otherwise indicated): novobiocin (30), bacitracin (0.04 IU per disc) and nitrofurantoin (300). It was resistant to the following antimicrobial compounds: rifampin (5), mycostatin (100), trimethoprim (5), erythromycin (15), penicillin G (10 IU per disc), ampicillin (10), chloramphenicol (30), neomycin (30), norfloxacin (10), ciprofloxacin (5), streptomycin (10), kanamycin (30), tetracycline (30), vancomycin (30), gentamicin (10) and nalidixic acid (30). The main phenotypic characteristics differentiating strain YC89T from H. regularis CGMCC 1.10123T and H. persicus IBRC-M 10043T are Mg2+ requirement, anaerobic growth with nitrate, arginine and DMSO, gas formation from nitrate, utilization of specific carbon sources, indole formation, hydrolysis of Tween 80, H2S formation (Table 1). More detailed results of phenotypic features of strain YC89T are given in the species description.
Chemotaxonomic Characteristics
The major polar lipids of strain YC89T are PG, PGP-Me, one glycolipids (GL2) chromatographically identical to S-DGD-1 and two unknown glycolipids (GL1 and GL3) (Supplementary Fig. S2). The phospholipid profile of strain YC89T is identical to that of H. regularis CGMCC 1.10123T. The main glycolipid composition of strain YC89T (GL1 and GL2) is chromatographically identical to the two glycolipids of H. regularis CGMCC 1.10123T. The major polar lipid composition supported classification of strain YC89T in the genus Halorientalis.
Phylogenetic Analysis
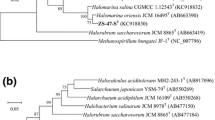
Eight complete 16S rRNA gene sequences of strain YC89T were obtained (1472 bp in length), and sequence comparisons indicated that strain YC89T has one kind of 16S rRNA gene sequence. 16S rRNA gene analysis revealed that strain YC89T was phylogenetically related to H. persicus D108T (95.6 % nucleotide identity) and H. regularis TNN28T (95.3 % nucleotide identity). These 16S rRNA gene similarities are well below the recently recommended thresholds (98.2–99.0, 98.65 %) to separate two prokaryotic species [6, 8]. Phylogenetic tree reconstructions using the maximum-likelihood (ML) algorithm revealed that strain YC89T tightly clustered with the current three members of Halorientalis (Fig. 1a). The phylogenetic position was also confirmed in the tree generated using the neighbour-joining (NJ) algorithm (Supplementary Fig. S3a).

Maximum-likelihood phylogenetic tree reconstructions based on 16S rRNA gene (a) and rpoB′ gene (b) sequences, showing the relationships between strain YC89T and related members within the family Halobacteriaceae. Bootstrap values (%) are based on 1000 replicates and are shown for branches with more 70 % bootstrap support. Bar represents expected substitutions per nucleotide position
The rpoB′ gene of strain YC89T was closely similar to the corresponding gene of H. persicus D108T (88.1 % nucleotide identity) and H. regularis TNN28T (88.0 % nucleotide identity), and the similarity values are less than the recommended threshold (86.2 %) which is used to distinguish genera [10]. In phylogenetic tree reconstruction using rpoB′ (Fig. 1b), strain YC89T tightly clustered with H. persicus D108T and H. regularis TNN28T. The phylogenetic position was also confirmed in the tree generated using the NJ algorithm (Supplementary Fig. S3b).
The 16S rRNA gene and rpoB′ gene-based phylogenetic analysis results supported the placement of strain YC89T in the genus Halorientalis.
The DNA G+C content of strain YC89T was determined to be 61.3 mol%, which is lower than those of H. regularis CGMCC 1.10123T (61.9 mol%) and H. persicus IBRC-M 10043T (62.8 mol%).
Based on these phenotypic, chemotaxonomic and phylogenetic properties, a novel species of the genus Halorientalis, H. brevis sp. nov., is proposed to accommodate strain YC89T. Characteristics that distinguish strain YC89T from the current two Halorientalis members are shown in Table 1.
Description of Halorientalis brevis sp. nov.
Halorientalis brevis (bre’vis. L. fem. adj. brevis, short)
Cells are motile, short rod-shaped (0.5–1.0 × 1.0–2.5) under optimal growth conditions and stain Gram-negative. Colonies on agar plates containing 3.1 M NaCl are red, elevated and round. The strain is chemoorganotrophic and aerobic. Growth occurs at 25–50 °C (optimum 37 °C), in the presence of 1.4–4.8 M NaCl (optimum 3.1 M NaCl), with 0–1.0 M MgCl2 (optimum 0.3 M MgCl2) and at pH 6.0–9.5 (optimum pH 7.5). Cells lyse in distilled water and the minimal NaCl concentration to prevent cell lysis is 8 % (w/uv). Catalase and oxidase are positive. Anaerobic growth occurs in the presence of nitrate, arginine and DMSO. Nitrate reduction to nitrite is observed, but gas formation from nitrate does not occur. H2S formation is positive, and indole formation is negative. Does not hydrolyse starch, casein, gelatin or Tween 80. The following substrates are utilized as single carbon and energy sources for growth: d-glucose, d-mannose, d-galactose, sucrose, glycerol, d-mannitol, d-sorbitol, acetate, pyruvate, dl-lactate, succinate, l-malate, fumarate and citrate. The following substrates are utilized as single carbon, nitrogen or energy sources for growth: l-alanine, l-arginine, l-aspartate, l-glutamate, l-lysine and l-ornithine. No growth occurs on d-fructose, l-sorbose, d-ribose, d-xylose, maltose, lactose or glycine. The major polar lipids are phosphatidic acid (PA), PG, PGP-Me, S-DGD-1 and two unknown glycolipids. The DNA G+C content of the type strain was 61.3 mol% (T m).
The type strain is YC89T (=CGMCC 1.12125T = JCM 18366T) and was isolated from Yuncheng salt lake in Shanxi, China.
References
Amoozegar MA, Makhdoumi-Kakhki A, Mehrshad M, Fazeli SAS, Spröer C, Ventosa A (2014) Halorientalis persicus sp. nov., an extremely halophilic archaeon isolated from a salt lake and emended description of the genus Halorientalis. Int J Syst Evol Microbiol 64:940–944
Cui H-L, Zhou P-J, Oren A, Liu S-J (2009) Intraspecific polymorphism of 16S rRNA genes in two halophilic archaeal genera, Haloarcula and Halomicrobium. Extremophiles 13:31–37
Cui H-L, Gao X, Yang X, Xu X-W (2010) Halorussus rarus gen. nov., sp. nov., a new member of the family Halobacteriaceae isolated from a marine solar saltern. Extremophiles 14:493–499
Cui H-L, Yang X, Gao X, Xu X-W (2011) Halobellus clavatus gen. nov., sp. nov. and Halorientalis regularis gen. nov., sp. nov., two new members of the family Halobacteriaceae. Int J Syst Evol Microbiol 61:2682–2689
Cui H-L, Yang X, Mou Y-Z (2011) Salinarchaeum laminariae gen. nov., sp. nov.: a new member of the family Halobacteriaceae isolated from salted brown alga Laminaria. Extremophiles 15:625–631
Kim M, Oh H-S, Park S-C, Chun J (2014) Towards a taxonomic coherence between average nucleotide identity and 16S rRNA gene sequence similarity for species demarcation of prokaryotes. Int J Syst Evol Microbiol 64:346–351
Liu Q, Ren M, Zhang L-L (2015) Natribaculum breve gen. nov., sp. nov. and Natribaculum longum sp. nov., halophilic archaea isolated from saline soil. Int J Syst Evol Microbiol 65:604–608
Marmur J, Doty P (1962) Determination of the base composition of deoxyribonucleic acid from its thermal denaturation temperature. J Mol Biol 5:109–118
Meier-Kolthoff JP, Göker M, Spröer C, Klenk H-P (2013) When should a DDH experiment be mandatory in microbial taxonomy? Arch Microbiol 195:413–418
Minegishi H, Kamekura M, Itoh T, Echigo A, Usami R, Hashimoto T (2010) Further refinement of Halobacteriaceae phylogeny based on the full-length RNA polymerase subunit B′ (rpoB′) gene. Int J Syst Evol Microbiol 60:2398–2408
Oren A, Ventosa A, Grant WD (1997) Proposed minimal standards for description of new taxa in the order Halobacteriales. Int J Syst Bacteriol 47:233–238
Oren A, Arahal DR, Ventosa A (2009) Emended descriptions of genera of the family Halobacteriaceae. Int J Syst Evol Microbiol 59:637–642
Oren A (2012) Taxonomy of the family Halobacteriaceae: a paradigm for changing concepts in prokaryote systematics. Int J Syst Evol Microbiol 62:263–271
Oren A (2014) Taxonomy of halophilic Archaea: current status and future challenges. Extremophiles 18:825–834
Xu W-D, Zhang W-J, Han D, Cui H-L, Yang K (2014) Halorussus ruber sp. nov., isolated from an inland salt lake of China. Arch Microbiol 197:91–95
Yuan P-P, Zhang W-J, Han D, Cui H-L (2015) Haloarchaeobius salinus sp. nov., isolated from an inland salt lake of China. Int J Syst Evol Microbiol 62:910–914
Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S (2011) MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol Biol Evol 28:2731–2739
Acknowledgments
This work was supported by the National Natural Science Foundation of China (No. 31370054), the 11th “Six Talents Peak” Project of Jiangsu Province (No. 2014-SWYY-021), the Qinglan Project of Jiangsu Province and a project funded by the Priority Academic Program Development of Jiangsu Higher Education Institutions (PAPD). We are grateful to Dr. M. A. Amoozegar for kindly providing strain Halorientalis persicus IBRC-M 10043T.
Author information
Authors and Affiliations
Corresponding author
Additional information
The GenBank/EMBL/DDBJ accession numbers for the 16S rRNA gene and rpoB ′ gene sequences of strain YC89T are JQ237120 and KJ913076, respectively.
Phase-contrast micrograph of strain YC89T, thin-layer chromatograms of strain YC89T and related members, neighbour-joining phylogenetic tree reconstructions based on 16S rRNA gene and rpoB′ gene sequences showing the relationships between strain YC89T and related members within the family Halobacteriaceae are available as supplementary materials.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Yuan, PP., Yin, S., Han, D. et al. Halorientalis brevis sp. nov., Isolated from an Inland Salt Lake of China. Curr Microbiol 71, 382–386 (2015). https://doi.org/10.1007/s00284-015-0861-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00284-015-0861-3