
Abstract
Roads have important effects on wildlife, such as natural habitat fragmentation and degradation and direct killing of fauna, which leads to reductions in wildlife population size. We focused on a principal road in Northwest Argentina to test for the effect of seasonality and landscape features on the composition of road-killed wildlife. We conducted regularly scheduled road trips during the dry and wet seasons. We recorded the presence or absence of a vegetation curtain or hedge along the road. We measured land use by remote sensing in a 500 m buffer along the road. We compared the abundance of animals killed between seasons (dry and wet) for different taxonomic groups (mammals, birds and reptiles) and for different origins (domestic and native). We built linear mixed models to test the effect of landscape features on the abundance of killed animals. Two hundred and ninety-three individuals were killed, belonging to 35 species; 75.8 % were native and 24.2 % domestic species. The majority of animals killed were mid-sized mammals. More animals were killed during the dry season. The most important factors to explain the wildlife road-killing were the season and the proportion of agricultural landscape. The composition of the killed animals changed with the season. The proportion of agricultural landscape incremented the number of killed birds and mammals during both seasons, without affecting reptiles. The ratio of wild to domestic animals killed was dependent on the season. This study sets a precedent as the first in road ecology in Northwest Argentina and should be taken into account for road planning and regulation.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Direct and indirect effects of roads jeopardize species and even entire ecosystems (World Resource Institute 1990; Forman and Alexander 1998). Roads act as physical and behavioural barriers to animals (Carr and Fahrig 2001). They affect otherwise continuous habitats by reducing their quantity and quality, provoking their alignment and causing edge effects (Brown and Lomolino 1998; Clevenger et al. 2001; Jaeger and Fahrig 2004). Indirectly, roads promote uncontrolled urban development along their course (Environment Protection Authority 1999). Roads affect the movement and distribution of a diversity of animal taxa, including mammals, birds, reptiles and amphibians (Carvalho and Mira 2011). Most obvious direct impacts are wildlife-vehicle collisions (WVC’s) causing mortality in animals, reducing population size and diminishing effective movement between populations for some species (Carr and Fahrig 2001), whilst alien species could take advantage of roads by spreading more quickly.
Some factors affecting WVCs are related to the fauna’s inherent characteristics: abundance (Seiler 2005), density (Joyce and Mahoney 2001; Dussault et al. 2006), and taxonomic group; amphibians, reptiles, mid-sized mammals, birds and small mammals are susceptible to different types of roads (Forman and Alexander 1998). Factors related to road characteristics are presence/absence of roadsides or barriers (Gunson et al. 2009) and vehicle speed limits (Seiler 2005; Danks and Porter 2010). Landscape-related predictors should also be considered; for example, the risk of VWC increases when the road bisects a level terrain (Gunson et al. 2011), in addition to the road itself (Clevenger et al. 2003; Kanda et al. 2006; Danks and Porter 2010; Jancke and Giere 2011; Garriga et al. 2012). Also, climatic conditions affect the impact range of these factors (Clevenger et al. 2003; Shepard et al. 2008); for example, temperature seasonality regulates animal population densities and behaviour (Rolandsen et al. 2011).
Differences in composition and species abundance of roadkills are related to season (Myers et al. 2008) and to habitat selection (Zhang et al. 2013). Roads change the spatial configuration and surroundings of the landscape. These changes in landscape configuration are associated with mean patch size, edge density, and core area and therefore roadkills could be spatially affected. Research on WVCs has demonstrated that they are spatially clustered in ‘hotspots’ (Puglisi et al. 1974; Hubbard et al. 2000; Clevenger et al. 2001; Joyce and Mahoney 2001). Landscape spatial patterns are expected to play an important role in determining roadkill locations and rates (Forman and Alexander 1998), as the presence of wildlife tends to be linked to specific habitats and adjacent land-use types.
Roads can affect wildlife by direct mortality, which is especially important for threatened species, but can also change population numbers and genetic flow by isolation. Many indirect effects of roads are cumulative and involve changes in community structure and ecological processes that are not well understood. Also, roads should be monitored as they have long-term effects that cause deterioration in ecosystems (Noss 1990). The expansion of the road network is one of the major threats to tropical biodiversity (Teixeira et al. 2016). Therefore there is a need to improve and integrate science and environmental licensing to mitigate wildlife mortality on roads.
In this study we present the first results on the composition of animals killed on the principal, basically north–south orientated, paved road, of 255 km in length, in the premontane forest of Yungas in Northwest Argentina. We examine the species composition of road-killed animals and we analyse the effect of spatial factors, i.e., surrounding landscape and roadway features and the seasonal pattern (wet and dry season) of wildlife-vehicle collisions. Finally, we relate our findings to management guidance. Season can have an effect on WVC in opposite ways; winter can increase the number of WVC by causing food scarcity, which increments animal movements, especially for ungulates (Gunderson and Andreassen 1998; Myers et al. 2008; Meisingset et al. 2013). Animal movements could also increase during the mating season and therefore also increment roadkills in spring, especially for reptiles (Southwood and Avens 2010; Van der Ree et al. 2015). Winter corresponds to the dry season in the study area, which could increase the effect on species even more due to water stress on animals. We therefore expect a higher number of roadkills in winter (dry season) for mammals; and for reptiles we expect a higher death rate in spring, corresponding to the wet season. Forest adjacent to roads increases the number of ungulate collisions and agriculture and urban areas surrounding roads decreases the number of collisions (Gunson et al. 2011). Therefore, in road segments surrounded by more natural vegetation offering habitat for native species, we expect more native species to be killed.
Methods
Study Area
Neotropical cloudy forests or Andean Yungas are a unique ecosystem associated with mountains, ranging from Venezuela to Argentina (Churchill et al. 1995; Myers et al. 2000). The subtropical Yungas are distributed in southern Bolivia and north-western Argentina (19°–29°S), occupying an area of 56,000 km2. Yungas extend from 400–1200 to 3400 m.a.s.l. and can be divided into three altitudinal vegetation belts with their typical flora and fauna (Cabrera and Willink 1973; Beck 1988; Brown and Grau 1993; Brown et al. 2001; Ferro and Barquez 2009). The lowest altitudinal level is occupied by the ‘lower montane forest’ or ‘premontane forest’, the second belt corresponds to the ‘upper montane cloud forest’ or ‘montane humid forest’, and the third forest belt corresponds to the ‘subalpine cloud forest’ or ‘upper montane forest’. In the southern limit they form a sort of peninsula of high biodiversity in relation to the surrounding biomes of the southern cone of South America (Chaco in the east and High Andes to the west). The Yungas ecosystem is considered vulnerable (Dinerstein et al. 1995). In Argentina, it is the most diverse ecoregion for mammals with 123 species (Barquez et al. 2006). Yungas are extremely seasonal with discontinuous precipitation regimes (Richter 2008) and 80 % of rain falls during summer (Brown et al. 2001). Rainfall ranges from 700 mm to 2000 mm/year, depending on altitude (Brown et al. 2001). At high elevations, this ecosystem has, in general, a relatively good conservation status due to its relief that hinders access and extractive activities. But the low and flat areas of the premontane forest suffer from deforestation and landscape transformation (Malizia et al. 2012). It is also at low altitudes where roads were traced to avoid difficult relief and landslides that occur frequently at more susceptible higher elevations. Remarkably, this first elevation band could also be the richest in animal species diversity (Ojeda et al. 2008) and could have higher abundances of rare and threatened species than higher elevations (Chalukian et al. 2009; Cuyckens 2013). Although a growing literature in the field of road ecology suggests that WVC can be major sources of vertebrate mortality and thus potentially limit wildlife populations (Aresco 2005), thereby requiring appropriate management, no road ecology studies exist in Northwest Argentina.
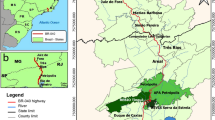
In the study area road density is low, with only two roads existing: the national road No 9 which transforms into the national road No 34 and the provincial road No 50. This study includes parts of the national road No 9, 34 and provincial road No 50. These paved roads extend 255 km through premontane forests of Yungas from the capital city of Salta to the city of Orán, near the international border with Bolivia (Fig. 1). The road track crosses 10 minor cities in the Argentine provinces of Jujuy and Salta, summing approximately 640,000 habitants, and we estimated a traffic volume of 4000 vehicles per day, with a high percentage of trucks (approximately 20 %). The busiest part of the road is in the surroundings of the city of Güemes because of the intersection between the roads No 9 and No 34 and more quiet near the city of Oran. The road was paved in the 1970’s but traffic increased since then. It crosses cultivated and urban areas and semi-natural vegetation of premontane forest. The road has 2 lanes and is 9 m in width, with a two-metre-wide grassy shoulder or road side along the entire trace, occasionally with a vegetation curtain or hedge.

Location of the road Salta–Orán and detail of the landscape classification in a 500 m buffer along the 2 km strip of the road used for analysis
Data Collection
We conducted 22 systematic road trips by pick-up truck along the entire length of the road (255 km); 13 during the dry season (May to September 2000) and 9 during the wet season (September 2000 to May 2001) with an interval of 16 ±30 days. Data collection was carried out by two observers (apart from the driver). We conducted trips during different times of the day, with a speed between 50 and 100 km/h and we varied direction of the road; travelling intercalated from south to north and from north to south. We georeferenced each road-killed animal and we identified it to the species level when possible, otherwise to genus or family. We grouped each animal by class as mammal, bird, reptile or amphibian and by origin into domestic or native (all wild species were native).
We split the road into segments of 1, 2 and 5 km in length for analysis, characterizing each segment by: (1) curvature of the road, (2) hedge presence, and (3) land use. We used curvature as a proxy of vehicle potential velocity; we used 5 classes (discrete quantitative variables) according to the highest angle of each segment: the higher the curvature class, the lower the potential velocity. We recorded the presence or absence of a hedge, considering a hedge as a single line of closely (max 1.5 m) spaced trees.
To characterize land use we created a buffer around the road to both sides and we calculated the proportion of different land uses. As for different species the effects of roads act on different scales (Jackson and Fahrig 2015) we tried buffer areas of 500, 750 and 1000 m, but proportions of each land use category did not change. We classified land use with three variables: agricultural, natural vegetation and urban areas. Agricultural areas belonged to cultivated areas; such as tobacco, soybean and other crops; and grasslands of anthropic origin. Natural vegetation areas belonged to the Yungas eco-region, and urban areas were cities and minor towns. To characterize the composition of the different types of landscape in each road segment we selected two Landsat TM images covering the study area (Path-Row: 231–276 and 231–277) from December 2000, the same year as that of the road data collection. We performed a supervised classification with SOPI vs. 2.5.1 image processing software using the Maximum Likelihood algorithm. Agriculture and Forest polygons were generated in Google Earth and used as ground truth data for the supervised classification. We used a total of 160 polygons; 70 % of these were used for the classification per se and the other 30 % for the evaluation of the classification. Urban areas were digitalised by visual interpretation. The overall accuracy of the classification was 97.7 %. Our analysis indicate that in the area about 68 % had agricultural use, 29 % were natural lands, 2.2 % were urban areas and the remainder 1.2 % of the area was miscellaneous (Fig. 1).
Data Analysis
To compare the number of killed animals per trip between seasons we used a t-test. We compared the abundance of animals killed per season (dry and wet seasons), for different taxonomic groups (mammals, birds and reptiles) and for different origins (domestic and native), with a χ 2 test which allows considering different sampling efforts between seasons when analysing relative frequencies. To evaluate if the abundance of animals killed was related to the characteristics of the road and surrounding landscape (curvature, hedge presence and land use) we conducted linear mixed models. We modeled the number of killed animals per segment with a generalized mixed model, function glmer from the package lme4 (Bates et al. 2014). We used this function because we found no auto-correlation among segments (acf function). We used the Poisson distribution to model the abundance of roadkills and curvature of the road; hedge presence and land use were considered as fixed factors while trips were considered as a random factor (Zuur et al. 2009). Agricultural and natural vegetation presented a strong negative correlation (Pearson Correlation Coefficient = −0.91; p < 0.0001) so we only kept “percentage of agriculture” and “percentage of urban areas” as variables for the model. We performed these analyses with 4 data–subsets of animals killed: birds, reptiles, native mammals and domestic mammals, split by season (wet and dry). We used the R (vs 3.1.3) software for all statistical analysis (R Development Core Team 2012).
Results
During this study we recorded 293 killed individuals, belonging to 35 species, 9 families and 6 orders (Fig. 2); 75.8 % correspond to native species and 24.2 % to domestic species. Also, three animals were not identified and excluded from all analysis. Of the animals killed 67.0 % were mammals, 24.7 % of them were birds and 8.3 % were reptiles. No amphibians were found. Among the native species we found 126 individuals of mammals (56.7 %), 72 birds (32.4 %), and 24 reptiles (10.8 %), while all domestic animals were mammals. Some species (34 %) were found only once. Among killed birds, raptors were the dominant group (88 %, 18 individuals). Reptiles were sporadically found; almost 92 % of them were snakes and just 8 % were lizards. We found 5 snake species including 2 vipers and three unknown species (Fig. 2).

Numbers of killed animals on a 255 km road in northwest Argentina during 13 trips during the dry season (May to September 2000) and 9 in the wet season (September 2000 to May 2001). We also found 13 snakes, 13 mammals and 5 birds (one dove) which we were unable to identify
The average number of animals killed per trip was not statistically different during the dry and wet season (14.2 vs. 11.1, t = 0.56, p = 0.577). However, the ratio of wild to domestic animals killed was dependent on season (χ 2 = 4.37, p = 0.036; Fig. 3a), with more wild animals killed during the wet season than expected (8 individuals more than expected). Also, within taxonomic groups, the amount of killed animals was strongly dependent on the season (χ 2 = 24.29, p < 0.001; Fig. 3b); birds appeared equally in the dry and wet season, mammals were 3-fold more abundant in the dry than in the wet season, while reptiles were 4-fold more abundant in the wet season than in the dry one. Some species were exclusive of either the dry or the wet season (Online Supplementary Material 1).

Abundances of domestic and wild animals killed during the wet and dry season a for different taxonomic groups and b along a 255 km road in north western Argentina
As the different length of segments and different buffer areas generated the same results, we only present models resulting from the 2 km segments and 500 m buffer area. The mixed model analysis showed a positive effect of the amount of agricultural landscape on bird mortality during wet and (marginally) dry seasons. Agricultural areas also affected native mammals only during the wet season and domestic mammals during the dry and (marginally) wet seasons (Table 1). We did not find any other significant relationship with city surface cover, curvature (related to potential velocity of vehicles) nor hedge presence.
Discussion
The composition of road-killed animals showed an inverted J-shaped curve (Fig. 2), so the ‘community’ of killed animals was composed of few species represented by a lot of individuals and most species by few individuals. No small rodents or amphibians were found, probably due to their small body size and multiple run-overs resulting in their quick disappearance (Santos et al. 2011). This underscores the importance of sampling frequency (Bager and da Rosa 2011) and additionally of driving order; if one always sets out on the road early in the morning and from the same place, it is more likely to find small carcasses in the first segments. Therefore, we changed the hour and direction of driving when we could. Also, the observation speed is important, but as we were covering more than 200 km, we had to balance between accuracy and cover, and a slower speed was not possible.
The majority of killed animals are mid-sized mammals, which are especially susceptible to this type of road; a two-lane, high-speed road (Forman and Alexander 1998), suggesting the possibility that they might avoid collisions if the road were narrower or drivers drove slower. But, as we do not have any correction data for survey detectability or carcass removal, nor any information about species abundance near the road, it is possible that species with higher mobility and lower reproduction rates are more susceptible to roads (Rytwinski and Fahrig 2011). This is important for management as these species would recover more slowly if populations were drastically diminished. The most common species killed was the grey fox (Lycalopex gymnocercus), while the other native canid, the crab-eating fox (Cerdocyon thous), was among the less abundant species killed. Grey foxes and crab-eating foxes are morphologically and ecologically similar, with the difference that grey foxes are diurnal while crab-eating foxes are nocturnal, with peaks of activity after dusk and before dawn (Di Bitetti et al. 2009). During dusk and dawn, collision chances could be low (Vieira and Port 2007) or contradictorily higher (Haikonen and Summala 2001), so crab-eating foxes could be over or under-represented. Although the crab-eating fox is typical of premontane forest in Yungas, it seems to be naturally much rarer than the grey fox (G.A.E.C., unpublished data) and therefore less killed than the grey fox.
The fifth abundant species was again a native species, Geoffroy’s cat (Leopardus geoffoyi). This species takes advantage of anthropogenic habitat (Pereira et al. 2012; Cuyckens et al. 2016) and could therefore advance on roads and adjacent transformed habitat with more kills as a consequence. The other felid species, Pampa’s cat (L. colocolo), was found once. This species is vulnerable in Argentina (Aprile et al. 2012) and, in contrast to Geoffroy’s cat, we estimate it to be extremely rare in low altitudes of Yungas; this species is usually found at higher elevations (>1500 m.s.al.), so its presence on the road was unexpected. The only rodent species was the coypus (Myocastor coypus) which is relative big in size (up to 10 kg); it was found twice. It is extremely rare in Yungas, so recording it twice is a relatively high number. Therefore, although this species is of Least Concern on a global and Argentinean level (Ojeda et al. 2015), we consider it locally of conservation concern. Remarkable was the encounter of the Argentinean Boa (Boa constrictor occidentalis), an endangered species (Giraudo et al. 2012) with an unclear taxonomic status, which therefore should be monitored to determine if more encounters occur and specimens should picked up for collection.
The most common birds were scavengers. This was not surprising as these are more susceptible to road mortality because of their carrion feeding habits taking advantage of carcasses on the road (Stoner 1925; Bennett 1990; Saunders and Hobbs 1991). It has been reported that aerial predators actively scavenge along road networks (Hubbard et al. 2012). But, the higher abundance of scavengers among bird roadkills may only reflect higher carcass detectability and lower carcass removal than for smaller birds. We did not find endangered raptors killed; the most common species found were the Black Vulture (Coragyps atratus) and Crested Caracara (Caracara plancus); both species are of Least Concern and the latter is commonly observed posing along the road side.
During the dry season more domestic and native animals were killed; domestic animals are especially sensitive to drought as they probably have fewer adaptations in their non-native environment. We found an effect of season on native mammals, which were more killed in the dry season. Mammals have to cover higher distances to satisfy their water requirements. For carnivores, water availability is an important factor limiting distribution (Valeix et al. 2009). The number of collisions was highest in the dry season, which corresponds to winter in the study area; winter is generally the lean period in terms of food availability, and scarcity of food affects the movement of native species. Deer are especially vulnerable to season as they make seasonal movements, but we did not find deer species in our study (Mazama sp.), indicating that the dry season implies a higher risk for other mammals as well (Smith-Patten and Patten 2008). Seasonal differences (and relief) are of the most important ecological factors shaping communities in Yungas.
Reptiles were most frequently killed during the wet season (80 %), agreeing with our suppositions. Reptiles encounter roads when making seasonal movements for foraging, breeding, and overwintering (Southwood and Avens 2010; Van der Ree et al. 2015). The wet season coincides with their reproduction season in the study area. The preponderance of snakes over lizards probably mirrors contrasting reaction and movement skills or could be an effect of their smaller body size. Among reptiles and amphibians, lizards are faster, because of their legs, and they are better adapted to life on land. Also, the Argentinean red tegu is common in premontane forest and Chaco and, because of its relative bigger body mass; it is conserved for longer periods of time when killed. The difference in abundance of roadkills between lizards and snakes is not supported by other studies. Reptiles were also the most common group in another study in Argentina (Attademo et al. 2011), but the eco-region in question was characterized by wetlands, increasing the natural presence of this group. This study also marked spring/summer as a factor that increases road-kills as we did, although this eco-region is not seasonally marked as is Yungas.
Contradictory to our hypotheses we found more killed animals, including domestic animals, when the proportion of agricultural areas was higher. Agricultural areas affected the number of mammals and birds, but not of reptiles. An increasing proportion of agricultural areas increased the number of birds killed; this could be related to killings of ground-dwelling birds such as gallinaceous birds and owls, ground-nesters and scavengers. These species are also habitat generalists and associated to open habitats. Agricultural areas increased the number of VWCs in the study area and roadkills were lower in natural areas. We did not find an effect of hedge presence on abundance of VWC’s in our study area. Roach and Kirkpatrick (1985) also found that hedges did not affect roadkills, but stated that trees near to roads were more used by birds than grassland areas. This could be related to the findings of Jacobson (2005) who discovered that hedges had a negative impact on roadkills of birds (more birds killed in the presence of a hedge). We did not find any effect on bird species, probably because the alternative of hedge in our study area is not grassland, but could also be natural areas with trees.
The absence of deer (Mazama gouazoubira and M. americana), present in the study area, in our roadkill data, is remarkable seeing as deer and other ungulates are commonly killed by vehicle collisions (Olson et al. 2014). One possibility is that these species have developed behavior to avoid collisions. Roads can change the behavior of animals; for example, reindeer in Norway started roaming parallel to the road (Panzacchi et al. 2013); and avoidance behavior in caribous was detected (Dyer et al. 2002). But, we suggest that they were lifted up by drivers for meat, as these species are commonly hunted as game meat (Altrichter 2005). The problem of survey detectability and carcass removal generally arises in roadkill studies, so we do not know how much of the data was effectively collected by us. For example, we did not find ocelot (Leopardus pardalis) killed on the road but we saw a vehicle driver lift up a felid species (most probably an ocelot) he had previously killed.
The effects of other linear structures such as trails, pipelines, and seismic lines on wildlife are similar in nature to those of roads (Jalkotzy et al. 1997). Sporadically, we found tapirs (Tapirus terristris) drowned in a water-channel in the study area (P. P. pers. obs.) and not on the road, maybe due to channel banks challenging big species such as tapir to getting out of the water, suggesting water channels to be a bigger threat for this particular species. The effects of these two linear structures in the study area could interact and should be considered together.
Cows, sheep, goats and even a horse were killed during our sampling period. Domestic grazers graze freely in rural and natural areas (no enclosures) in the study area. Grazing animals could be attracted to roadsides because of the presence of tender herbs, especially after mowing, and ones near the road could be easily killed. In addition to negatively impacting wildlife, collisions with big wild or domestic animals can cause economic and social costs associated with vehicle towing and repair; human injuries and fatalities; and accident mitigation, support and investigation. For example, Conover et al. (1995) estimated that more than one million deer–vehicle collisions occur annually in the United States, costing over $1.1 billion in repair and resulting in 29,000 human injuries and 211 human fatalities. No such estimates exist in Yungas or Argentina.
Two factors came out as most important for animals killed on road: the season on the composition of the killed animals and the proportion of agricultural landscape on the abundance of road-killed birds and native and domestic mammals. So, when studying different taxa of wildlife, seasonality should be taken into account differentially. Population and movement studies on wildlife should consider measurements of variables both during wet and dry seasons. Yungas are highly seasonal and temporal variation in rainfall is important to wildlife. The dry season increases movements in search for water sources and the wet season is when mating is concentrated and therefore is associated with a higher risk for VWC. Climate change could enhance seasonal differences even more, so interactions of roads with climate change are also important. In this study we used a short time scale to avoid changes in environmental variables.
Of the potential components that affect WVCs we found one habitat and one season-related factor as important. Other factors could be; human causes: traffic volume (Gu et al. 2011), attitude of drivers, road composition (paved or not), guardrail and barrier installation, lighting, road widening and improvements (Clevenger et al. 2001); animal-related causes: density or behavior, relationship between population size and distance to road, as well as population dynamics of killed animals (Groot Bruinderink and Hazebroek 1995); and environmental factors such as weather conditions and the moment of the day (Haikonen and Summala 2001). For large carnivores, road density on a regional scale could be more important (Thiel 1985). In northern countries snow depth was crucial (Rolandsen et al. 2011); in Yungas, it could be important to identify and measure which seasonal factor constrains animal movement and habitat use; in mountainous areas as our study area, animals carry out altitudinal movements following the season, so animal movement studies, home range studies etc. are needed.
Mitigation remains challenging as results from one area cannot be extrapolated to another and the effect of roads varies between species and groups of species. For example, birds present several challenges compared to other vertebrates as they do not use wildlife over-crossings; and scavengers are killed while foraging on roadkill (Jacobson 2005). In this study most birds were scavengers, so we suggest that if mitigation actions for the other groups are successful then also the road-killed scavengers will diminish as less forage will be available on the road. For our study area, we suggest two types of mitigation actions. For domestic animals we need to encourage social conscience in owners of domestic animals, as collisions cause economic costs and are also a danger for drivers. Domestic animals should be more controlled, enclosing dogs and cats, and especially farm animals such as cows, horses, goats and sheep. Also, drivers should consider diminishing their speed near towns, seeing as not only animals can be hit but also humans. Native animals are of a bigger complication. Ecopassages are expensive and, as we could not (yet) detect hotspots, we do not encourage their implementation yet, if the roads maintain their current properties. In this study we could not identify hotspots of roadkills, suggesting that season might be more important. As agricultural areas had an influence on roadkills it may be more useful to maintain natural areas along the road. So, based on these findings and our updated knowledge of the areas surrounding the roads, we suggest that the segment from the crossroads, where the No 66 road meets the No 34 road (its southern part) until ‘Bananal’, a small town further north, receive full protection. Spending money on hedges might not be useful, as we did not discover any influence of them. Complementary, although speed did not come out as an important factor in our study area, based on the characteristics of the road (high-speed, two lanes), warning and maximum velocity signs and radars to control speed could be placed. We also suggest that, before any ‘improvement’ on the road is made (for example, making it wider or installing new pavement), a previous study or environmental impact analysis should be carried out. A double goal (awareness–raising and information–generation) could be achieved by inviting the general public to contribute information of killed animals, for example by a cellphone app that indicates the exact roadkill location or by an Internet form; examples of this are: <http://impactorutasecobiouy.blogspot.com.ar/p/blog-page.html> and <http://www.wildlifemapping.org/>. Our map of roadkills could be made available online and updated with this information.
This study provides information on numbers and species road-killed in Northwest Argentina for the first time, which is valuable data. The methods employed here were useful for the objectives of our study, but not for small animals such as amphibians, rodents or lizards. For those groups, frequency of road trips must be increased, probably shortening the trajectory surveyed. Analyzing satellite images with adequate software reduces field work and allows concentrating on the detection of roadkills. Road ecology is a relatively new discipline in Argentina. Published roadkill data are difficult to find, so this study together with Attademo et al. (2011) and Malizia et al. (1998) for birds, are the first studies in the country. We encourage more studies of this type and the development of road ecology induced by national funding in the country. Also, new and existing scientific information should be made available to decision makers, scientists and urban developers who, together, should work more closely when planning or improving a (new) road.
Apart from mitigation on existing roads, prevention of impacts of new roads or improvements of existing roads (such as paving) is important. Previous to tracing roads, populations studies should be done in order to design roads that reduce their impact on the environment (Jones 2000; Roger et al. 2011). In the study area virtually no population studies exist; we therefore encourage populations studies on endangered species in the area, such as the jaguar (Panthera onca), in critical danger (Aprile et al. 2012) , and the white-lipped peccary (Tayassu peccary), in danger (Chalukian et al. 2012). Both need huge areas for their development, and higher road density could affect negatively their already critically small populations. The causes of the absence of deer, tapirs and bigger carnivores might be important, as they could be related to decreasing populations of these species. The human population, economy, and consequently road density are growing in Northwest Argentina, for this reason, landscape and urban planning is absolutely necessary. For example, at least two roads are planned in the area; one to connect Bermejo with Los Toldos, in the extreme north of Salta; and one west-east directed, connecting two eco-regions: High Andes with Yungas (form Santa Ana to Valle Colorado in the Jujuy province). These roads will fragment a highly biodiverse and geomorphologically unstable eco-region. Also, vehicle volume has increased since our study (from our estimated 4000 per day to 13000 vehicles per day in the most transited part, in the city of Güemes). So, studies of landscape ecology and fragmentation and corridor planning and implementation should avoid impacts on an already highly-threatened eco-region, allowing, at the same time, safer transportation for people. This eco-region, although threatened and highly fragmented, still harbours important species, such as top predators (jaguar), engineers of ecosystems (tapirs and white-lipped peccaries) and accomplishes important ecological services, such as protecting watersheds amongst others. For these reasons we believe that investing in its conservation and minimizing the effects of roads and other human impacts is an investment for the future.
References
Altrichter M (2005) The sustainability of subsistence hunting of peccaries in the Argentine Chaco. Biol Cons 126:351–362. doi:10.1016/j.biocon.2005.06.024
Aprile G, Cuyckens E, De Angelo C, et al. (2012) Familia: Felidae. In: Ojeda RA, Chillo V, Diaz Isenrath GB (eds) Libro rojo de Mamíferos amenazados de la Argentina, SAREM, Argentina, pp 92–101.
Aresco M (2005) The effect of sex-specific terrestrial movements and roads on the sex ratio of freshwater turtles. Biol Cons 123:37–44. doi:10.1016/j.biocon.2004.10.006
Attademo AM, Peltzer PM, Lajmanovich RC et al. (2011) Wildlife vertebrate mortality in roads from Santa Fe Province, Argentina. Rev Mex Biodivers 82:915–925
Bager A, da Rosa C (2011) Influence of sampling effort on the estimated richness of road-killed vertebrate wildlife. Environ Manage 47:851–858. doi:10.1007/s00267-011-9656-x
Barquez RM, Díaz MM, Ojeda RA, Sociedad Argentina para el Estudio de los Mamíferos (2006) Mamíferos de Argentina : sistemática y distribución. SAREM, Tucumán, Argentina
Bates D, Mächler M, Bolker B, Walker S (2014) Fitting linear mixed-effects models using lme4.
Beck S (1988) Las regiones ecológicas y las unidades fitogeográficas de Bolivia. In: Morales CB (ed) Manual de Ecología, La Paz, pp 233–267.
Bennett AF (1990) Habitat corridors and the conservation of small mammals in a fragmented forest environment. Landscape Ecol 4:109–122
Brown AD, Grau HR (1993) La naturaleza y el hombre en las Selvas de montaña. Proyecto GTZ - Desarrollo Agroforestal en Comunidades Rurales del Noroeste, Argentino, Salta
Brown AD, Grau HR, Malizia LR, Grau A (2001) Argentina. In: Kapelle M, Brown AD (eds) Bosques nublados del Neotrópico, INBio, San José, Costa Rica, pp 623–659.
Brown JH, Lomolino MV (1998) Biogeography, 2nd edn. Sinauer Asociates, Inc., Sunderland, Massachusetts, USA
Cabrera AL, Willink A (1973) Biogeografía de América Latina. Secretaría General de la Organización de los Estados Americanos, Washington DC
Carr LW, Fahrig L (2001) Effect of road traffic on two amphibian species of differing vagility. Conserv Biol 15:1071–1078
Carvalho F, Mira A (2011) Comparing annual vertebrate road kills over two time periods, 9 years apart: a case study in Mediterranean farmland. Eur J Wildl Res 57:157–174. doi:10.1007/s10344-010-0410-0
Chalukian S, De Bustos S, Lizárraga L, et al. (2009) Plan de acción para la conservación del tapir (Tapirus terrestris) en Argentina.
Chalukian S, de Bustos S, Quiroga V et al. (2012) Orden Artiodactyla. Familia Tayassuidae. Libro rojo de Mamíferos amenazados de la Argentina. SAREM (Sociedad Argentina para el Estudio de los Mamíferos), Buenos Aires, Argentina, pp 117–120
Churchill SP, Balslev H, Forero E, Luteyn JL (1995) Biodiversity and conservation of neotropical montane forests. In: Proceedings of the symposium, the New York Botanical Garden, 21.26 June 1993. p xiv + 702.
Clevenger AP, Chruszcz B, Gunson K (2001) Drainage culverts as habitat linkages and factors affecting passage by mammals. J Appl Ecol 38:1340–1349
Clevenger AP, Chruszcz B, Gunson KE (2003) Spatial patterns and factors influencing small vertebrate fauna road-kill aggregations. Biol Conserv 109:15–26
Conover MR, Pitt WC, Kessler KK et al. (1995) Review of human injuries, illnesses, and economic losses caused by wildlife in the United States. Wildl Soc Bull 23:407–414
Cuyckens GAE (2013) Distribución geográfica y conservación de los félidos presentes en Argentina y las Yungas a través de modelos de distribución de especies. Tesis doctoral, Universidad Nacional de Salta.
Cuyckens GAE, Pereira JA, Trigo TC et al. (2016) Refined assessment of the geographic distribution of Geoffroy’s cat (Leopardus geoffroyi) (Mammalia: Felidae) in the Neotropics. J Zool 298:285–292. doi:10.1111/jzo.12312
Danks ZD, Porter WF (2010) Temporal, spatial, and landscape habitat characteristics of moose-vehicle collisions in western Maine. J Wildl Manage 74:1229–1241. doi:10.1111/j.1937-2817.2010.tb01243.x
Di Bitetti MS, Di Blanco YE, Pereira JA et al. (2009) Time partitioning favors the coexistence of sympatric crab-eating foxes (Cerdocyon thous) and pampas foxes (Lycalopex gymnocercus). J Mammal 90:479–490
Dinerstein E, Olson DM, Graham DJ et al. (1995) Una evaluación del estado de conservación de las eco-regiones terrestres de América Latina y el Caribe. Banco Mundial, Washington, DC
Dussault C, Poulin M, Courtois R, Ouellet J-P (2006) Temporal and spatial distribution of moose-vehicle accidents in the Laurentides Wildlife Reserve, Quebec, Canada. Wildl Biol 12:415–425. doi:10.2981/0909-6396(2006)12[415:TASDOM]2.0.CO;2
Dyer SJ, O’Neill JP, Wasel SM, Boutin S (2002) Quantifying barrier effects of roads and seismic lines on movements of female woodland caribou in northeastern Alberta. Can J Zool 80:839–845. doi:10.1139/z02-060
Environment Protection Authority (1999) Environmental criteria for road traffic noise. Environment Protection Authority, Chatswood, N.S.W
Ferro LI, Barquez RM (2009) Species richness of nonvolant small mammals along elevational gradients in Northwestern Argentina. Biotropica 41:759–767. doi:10.1111/j.1744-7429.2009.00522.x
Forman RT, Alexander LE (1998) Roads and their major ecological effects. Annu RevEcol System 29:207–231
Garriga N, Santos X, Montori A et al. (2012) Are protected areas truly protected? The impact of road traffic on vertebrate fauna. Biodivers Conserv 21:2761–2774. doi:10.1007/s10531-012-0332-0
Giraudo AR, Arzamendia V, Bellini GP et al. (2012) Categorización del estado de conservación de las Serpientes de la República Argentina. Cuad Herpetol 26:303–326
Groot Bruinderink GWTA, Hazebroek E (1995) Ungulate traffic collisions in Europa. Conserv Biol 10:1059–1067
Gu H, Dai Q, Wang Q et al. (2011) Factors contributing to amphibian road mortality in a wetland. Curr Zool 57:768–774
Gunderson H, Andreassen HP (1998) The risk of moose Alces alces collision: a predictive logistic model for moose-train accidents. Wildl Biol 4:103–110
Gunson KE, Clevenger AP, Ford AT et al. (2009) A comparison of data sets varying in spatial accuracy used to predict the occurrence of wildlife-vehicle collisions. Environ Manage 44:268–277. doi:10.1007/s00267-009-9303-y
Gunson KE, Mountrakis G, Quackenbush LJ (2011) Spatial wildlife-vehicle collision models: a review of current work and its application to transportation mitigation projects. J Environ Manage 92:1074–1082. doi:10.1016/j.jenvman.2010.11.027
Haikonen H, Summala H (2001) Deer–vehicle crashes extensive. peak at 1 hour after sunset. Am J Prev Med 21:209–213
Hubbard KA, Chalfoun AD others ( (2012) An experimental evaluation of potential scavenger effects on snake road mortality detections. Herpetol Conserv Biol 7:150–156
Hubbard MW, Danielson BJ, Schmitz RA (2000) Factors influencing the location of deer-vehicle accidents in Iowa. J Wildl Manag 64:707. doi:10.2307/3802740
Jackson HB, Fahrig L (2015) Are ecologists conducting research at the optimal scale? Is research conducted at optimal scales? Glob Ecol Biogeogr 24:52–63. doi:10.1111/geb.12233
Jacobson SL (2005) Mitigation measures for highway-caused impacts to birds. General technical report PSWGTR-191 US Department of Agriculture, Forest Service, Albany, California pp 1043–1050.
Jaeger JAG, Fahrig L (2004) Effects of road fencing on population persistence. Conservation Biology 18:1651–1657
Jalkotzy MG, Ross PI, Nasserden MD (1997) The Effects of Linear Developments on Wildlife: a Review of Selected Scientific Literature. Canadian Association of Petroleum Producers, Arc Wildlife Services, Ltd., Calgary, Canada
Jancke S, Giere P (2011) Patterns of otter Lutra lutra road mortality in a landscape abundant in lakes. Eur J Wildl Res 57:373–381
Jones ME (2000) Road upgrade, road mortality and remedial measures: impacts on a population of eastern quolls and Tasmanian devils. Wildl Res 27:289–296
Joyce TL, Mahoney SP (2001) Spatial and temporal distributions of moose-vehicle collisions in Newfoundland. Wildl Soc Bull 29:281–291
Kanda LL, Fuller TK, Sievert PR (2006) Landscape associations of road-killed Virginia opossums (Didelphis virginiana) in central Massachusetts. Am Midl Nat 156:128–134
Malizia LR, Aragón R, Chacoff P, Monmany AC (1998) ¿Son las rutas una barrera para el desplazamiento de las aves? El caso de la reserva provincial La Florida (Tucumán, Argentina). El Hornero 15:10–16
Malizia LR, Pacheco S, Blundo C, Brown AD (2012) Caracterización altitudinal, uso y conservación de las Yungas Subtropicales de Argentina. Ecosistemas 21:53–73
Meisingset EL, Loe LE, Brekkum Ø et al. (2013) Red deer habitat selection and movements in relation to roads. J Wildl Manag 77:181–191. doi:10.1002/jwmg.469
Myers N, Mittermeier RA, Mittermeier CG et al. (2000) Biodiversity hotspots for conservation priorities. Nature 403:853–858
Myers WL, Chang WY, Germaine SS, et al. (2008) An analysis of deer and elk-vehicle collision sites along state highways in Washington State.
Noss R (1990) Indicators for monitoring biodiversity: a hierarchical approach. Conserv Biol 4:355–364
Ojeda RA, Barquez RM, Stadler J, Brandl R (2008) Decline of mammal species diversity along the Yungas Forest of Argentina. Biotropica 40:515–521. doi:10.1111/j.1744-7429.2008.00401.x
Ojeda R, Bidau C, Emmons L (2015) Myocastor coypus. In: The IUCN Red List of Threathened Species. http://www.iucnredlist.org/details/14085/0. Accessed 6 Apr 2015.
Olson DD, Bissonette JA, Cramer PC et al. (2014) Monitoring wildlife-vehicle collisions in the information age: how smartphones can improve data collection. PLoS ONE 9:e98613. doi:10.1371/journal.pone.0098613
Panzacchi M, Van Moorter B, Strand O (2013) A road in the middle of one of the mast wild reindeer migration routes in Norway: crossing behaviour and threats to conservation. Rangifer 33:15–26
Pereira JA, Walker RS, Novaro AJ (2012) Effects of livestock on the feeding and spatial ecology of Geoffroy’s cat. J Arid Environ 76:36–42. doi:10.1016/j.jaridenv.2011.08.006
Puglisi MJ, Lindzey JS, Bellis ED (1974) Factors associated with highway mortality of white-tailed deer. JWildl Manag 38:799. doi:10.2307/3800048
R Development Core Team (2012) The R project for statistical computing. Viena, Austria.
Richter M (2008) Tropical mountain forests - distribution and general featurres. In: Gradstein SR, Homeier J, Gansert D (eds) The tropical mountain forests - Patterns and processes in a biodiversity hotspot, Göttingen Centre for Biodiversity and Ecology, pp 7–24.
Roach GL, Kirkpatrick RD (1985) Wildlife use of roadside woody plantings in Indiana. Transport Res Rec 1016:11–15
Roger E, Laffan SW, Ramp D (2011) Road impacts a tipping point for wildlife populations in threatened landscapes. Popul Ecol 53:215–227. doi:10.1007/s10144-010-0209-6
Rolandsen CM, Solberg EJ, Herfindal I et al. (2011) Large-scale spatiotemporal variation in road mortality of moose: is it all about population density?. Ecosphere 2:art113. doi:10.1890/ES11-00169.1
Rytwinski T, Fahrig L (2011) Reproductive rate and body size predict road impacts on mammal abundance. Ecol Appl 21:589–600
Santos SM, Carvalho F, Mira A (2011) How long do the dead survive on the road? Carcass persistence probability and implications for road-kill monitoring surveys. PLoS ONE 6:e25383. doi:10.1371/journal.pone.0025383
Saunders DA, and Hobbs RJ (eds) (1991) Nature conservation 2, the role of corridors. Surrey Beatty & Sons in Association with Western Australian Laboratory of the Commonwealth Scientific and Industrial Research Organization, Division of Wildlife Ecology, Western Australian Dept. of Conservation and Land Management : Western Australian Roadside Conservation Committee : Western Australian Main Roads Dept., Chipping Norton, N.S.W
Seiler A (2005) Predicting locations of moose-vehicle collisions in Sweden: Predicting locations of moose-vehicle collisions. J Appl Ecol 42:371–382. doi:10.1111/j.1365-2664.2005.01013.x
Shepard DB, Dreslik MJ, Jellen BC, Phillips CA (2008) Reptile road mortality around an oasis in the Illinois corn desert with emphasis on the endangered eastern Massasauga. Copeia 2008:350–359. doi:10.1643/CE-06-276
Smith-Patten B, Patten M (2008) Diversity, seasonality, and context of mammalian roadkills in the southern great plains. Environ Manage 41:844–852. doi:10.1007/s00267-008-9089-3
Southwood A, Avens L (2010) Physiological, behavioral, and ecological aspects of migration in reptiles. J Comp Physiol [B] 180:1–23. doi:10.1007/s00360-009-0415-8
Stoner D (1925) The toll of the automobile. Science 61:56–57
Teixeira FZ, Coelho IP, Lauxen M et al. (2016) The need to improve and integrate science and environmental licensing to mitigate wildlife mortality on roads in Brazil. Trop Conserv Sci 9:34–42
Thiel RP (1985) Relationship between Road densities and wolf habitat suitability in Wisconsin. Am Midl Nat 113:404. doi:10.2307/2425590
Valeix M, Loveridge AJ, Davidson Z et al. (2009) How key habitat features influence large terrestrial carnivore movements: waterholes and African lions in a semi-arid savanna of north-western Zimbabwe. Landscape Ecol 25:337–351. doi:10.1007/s10980-009-9425-x
Van der Ree R, Smith DJ, Grilo C (2015) Handbook of road ecology. John Wiley and Sons, USA
Vieira EM, Port D (2007) Niche overlap and resource partitioning between two sympatric fox species in southern Brazil. J Zool 272:57–63. doi:10.1111/j.1469-7998.2006.00237.x
World Resource Institute (1990) World resources 1990-1991: a guide to the global environment. Oxford University Press, New York, USA
Zhang M, Liu Z, Teng L (2013) Seasonal habitat selection of the red deer (Cervus elaphus alxaicus) in the Helan Mountains, China. Zoologia (Curitiba) 30:24–34. doi:10.1590/S1984-46702013000100003
Zuur AF, Ieno EN, Walker NJ et al. (2009) Mixed effects models and extensions in ecology with R. Springer, New York
Acknowledgments
The authors thank Greenpeace-Argentina and Grupo Yaguareté for logistical support. We are thankful to S. de Bustos, J. Gato and D. Vazquez for data collection and to J. Osorio and M. Zietsman for English revision. We thank three anonymous reviewers and the editor.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of Interest
The authors declare that they have no conflict of interest.
Electronic supplementary material
Rights and permissions
About this article

Cite this article
Cuyckens, G., Mochi, L., Vallejos, M. et al. Patterns and Composition of Road-Killed Wildlife in Northwest Argentina. Environmental Management 58, 810–820 (2016). https://doi.org/10.1007/s00267-016-0755-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00267-016-0755-6