
Abstract
We surveyed road kills occurring along a 26-km stretch of a major national road (Portugal) in two different years: 1996 and 2005. For analysis purposes, we divided the data into seven vertebrate groups: amphibians, reptiles, carnivores, prey mammals (shrews, moles, rodents, rabbits and hares), hedgehogs, owls and passerines. Main factors influencing vertebrate road casualties were evaluated using redundancy analysis and variance partitioning techniques, focusing on three sets of variables: land cover, landscape metrics and spatial location. We also took into account meteorological conditions and changes in traffic intensity specific to each of the surveyed years. The percentage of variance explained by the explanatory variables was greater in 1996 (67.5%) than in 2005 (48.1%). Many variables influencing road kill incidence were common to both years. The most significantly associated factor was the distance to the Natural Park of Serra de São Mamede (NPSSM): road kills decreased steadily as our survey moved south, away from the NPSSM border. Moreover, an increased incidence of road losses occurred in forested areas, such as montado and traditional olive groves. As 2005 was a climatically drier year, additional variance factors became prominent, including the distance to water reservoirs, suggesting a greater influence of water availability. Traffic flow increased by almost 150% from 1996 to 2005, which may explain the overall increase in road kills, with the notable exception of the amphibian group, whose road fatalities incidences decreased approximately sixfold. We expect that our survey will provide a comprehensive understanding of the most critical factors currently influencing vertebrate road fatalities and aid in improving the effectiveness of mitigation measures to reduce them.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
One of the most visible effects of roads on wildlife is road kill, a major threat to biodiversity conservation (Forman 1998; Forman and Alexander 1998; Trombulak and Frissell 2000; Sherwood et al. 2002; Forman et al. 2003; Coffin 2007). Roads can affect all types of life form on Earth, from small invertebrates, such as slugs, to large animals, like moose and brown bears (Smith-Patten and Patten 2008). Indeed, several ecological researchers have pointed out that roads are one of the main causes of modern-day vertebrate population decline and in the decrease of viability across generations (Crooks and Sanjayan 2006; Ament et al. 2008). This may be especially true for small mammals, on which barrier effects include the reluctance to cross roads, thereby leading to local extinctions (Rico et al. 2007; McGregor et al. 2008). Among larger and rarer species like the Iberian lynx (Lynx pardinus), road kills are the principal cause of death among cubs in Doñana (southwest Spain; Ferreras et al. 1992). In Britain, carnivores like badgers lose up to 66% of all post-emergent cubs and adults near urban zones (Clarke et al. 1998). In addition, the principal non-natural cause of death in otters in Britain is road fatalities, which has contributed to the general population decline of this species (Philcox et al. 1999). In Spain, barn owl populations decreased by 70% over a 10-year period, and road casualties are considered the principal cause of these deaths (Fajardo 2001). Research by Fahrig et al. (1995) suggests that the presence of roads and the increase in traffic intensity may have contributed to the global decline in amphibians, as well. Several studies have revealed a negative correlation between the amphibian anuran species' relative abundance and traffic density (Fahrig et al. 1995; Carr and Fahrig 2001; Hels and Buchwald 2001), between anuran pond occupancy and road density (Vos and Chardon 1998), and between species richness in breeding sites and paved road density (Findlay et al. 2001; Eigenbrod et al. 2008). Vertebrates remain the most studied group in road mortality research, not only because of their size but also because they comprise flagship species (Forman et al. 2003).
Understanding the principle factors underlying vertebrate road fatalities, and their relationship to natural seasonal variations that occur among vertebrate groups, will provide ecological managers with the knowledge necessary to reduce collision rates and road impact upon wildlife (Saeki and Macdonald 2004; Ramp et al. 2005; Seiler 2005; Conard and Gipson 2006). This will be accomplished by determining and describing the location of road kill aggregations (Clevenger et al. 2003; Ramp et al. 2006), which tend to be linked to specific habitats and landscape types within the road vicinity (Forman and Alexander 1998; Caro et al. 2000; Gomes et al. 2008).
Like other ecological phenomena, road kill incidence rates are mediated by numerous factors that act at a multitude of organisational levels and along a wide range of spatial and temporal scales (Cushman and McGarigal 2002). Variation partitioning is a quantitative statistical method by which the variation in response variable(s) can be decomposed into independent components, thereby reflecting the relative importance of different groups of explanatory variables and their joint effects (Borcard et al. 1992; Heikkinen et al. 2004). We used this approach to understand the influence of land cover, landscape metrics and spatial location on vertebrate road loss patterns within a Mediterranean environmental context. The partitioning method is versatile, as it can be used in a variety of univariate and multivariate settings, and with either linear or unimodal response methods (Cushman and McGarigal 2002).
The main goals of our study were (1) to describe vertebrate road kill patterns, during the two surveyed years along a section of a major national road in central Portugal; (2) to quantify the relative importance of land cover, landscape characteristics and spatial location influencing road kills and to evaluate the main factors individually; and (3) to compare road kill patterns with respect to variations in climate and traffic intensity.
Study area
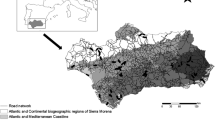
This study took place along a 26-km stretch of a main road (IP2) located in the Portalegre district, in central Portugal. The road itself is comprised of two paved lanes along its entire length and runs from the central-west border of the Natural Park of Serra de São Mamede (NPSSM) in Portalegre southward to the small village of Monforte (Fig. 1). After our field work was concluded, the area surrounding Monforte was classified as a Special Protection Zone (DR, 1ª Série, nº 40, 26 February 2008).

Location of the studied national road stretch (IP2), near the Natural Park of Serra de São Mamede (NPSSM), Portugal (obtained from GIS program Arcview 3.2)
Located near the Spanish border, this region is dominated by smooth landscape, except for the mountain topography of the Natural Park, which reaches 1,024 m above sea level.
The immediate road vicinity is composed of a characteristic Mediterranean agro-silvo-pastoral system of cork (Quercus suber) and holm oak (Quercus rotundifolia) woodlands (hereafter referred to as montado) and open land used as pasturelands, meadows, extensive agriculture and olive groves. Road topography varies slightly along the 26-km stretch, being 30.8% level, 48.8% buried and 20.4% raised relative to the immediately adjacent area.
The climate is Mediterranean with warm, dry summers alternating with cold, rainy winters. However, 2005 was an extremely dry year, with very low annual precipitation levels (IM 2005). The NPSSM is considered an Atlantic biogeographic island embedded in a Mediterranean-type matrix. This biogeographic crossroad enables the coexistence of several species from both regions and contributes to a uniquely high level of biodiversity inside the Park and its surrounding areas.
Roughly 2 years prior to 1996, the road stretch studied was enlarged, so that by 1996, it supported a moderate traffic volume of 2,965 vehicles per day (IEP 2000). By 2005, the daily traffic volume had reached 6,950 vehicles per day, and the area had become one of high traffic intensity (EPE 2005). Despite the massive increase in mean daily traffic volume, traffic peaks occurred in August of both years.
Methods
Road kill survey
Vertebrate road kills were surveyed by car, travelling at an average speed of 20 km/h, every 2 weeks from January through December in 1996 and again in 2005 (for a total of 26 road samples per year). All vertebrate mortalities were collected and identified by species level in loco, whenever possible, or by analysis in the laboratory of skin, scales, feathers or hairs depending upon the taxonomic group. We also obtained the geographic coordinates location of all road fatalities using a global positioning system unit combined with land cover maps and detailed maps (1:2,000) of road profiles. Remains were removed from the road to avoid double counting.
Sampling units
For analysis purposes, we divided the studied road into 52 segments, each 500-m long; and for every ecological group considered, we used the number of road losses registered as a response variable in multivariate analysis. A buffer 500-m wide was created around each segment, and we obtained a total of 52 rectangular polygons (≈50 ha), hereafter referred as sampling units (SU), on which explanatory variables were computed and comparing tests made. We choose a 500-m buffer based upon the average road effects defined for birds by Forman and Deblinger (2000) and Forman et al. (2002), for amphibians by Eigenbrod et al. (2008) and for reptiles by Boarman and Sazaki (2006). We also believe that 500-m segments along the road are a reasonable length for the implementation of road kill mitigation measures.
Explanatory variables
Each of the 52 SU was characterised for the 48 explanatory variables used in the present study. Variables were clustered into three groups: land cover, landscape metrics and spatial coordinates (Table 1). Detailed land cover maps were obtained through the interpretation of 2003 aerial photographs, complemented with fieldwork surveys. Comparing 1995 aerial photographs with those obtained in 2003 revealed that only minor changes had occurred. Based upon this evidence, we decided to use the same land cover map for both years under investigation. Land cover types include pasturelands, forests (comprising montado, old olive groves, and pines and eucalyptus plantations), urban zones, aquatic areas (rivers and water reservoirs), shrublands and roads.
We used the GIS program Arcview 3.2 (ESRI 1999) and the Patch analyst 2.2 (Eikie et al. 1999) extension to obtain the landscape metrics for each SU (please see Table 1 for details). Distances and all spatial descriptors were derived considering the midpoint of each 500-m road segment.
A very important landscape metric was identified as the distance to the NPSSM (see Table 1). This variable should be interpreted as the distance to the central western limit of NPSSM, which is an important natural area dominated by a NE–SW-oriented mountain range. The park itself is known for its high levels of humidity and rainfall and its particularly well-preserved landscapes. These landscapes are good examples of harmonious interactions between man and nature, maintaining high levels of biodiversity. Road topographic predictors included in the landscape metrics set were obtained by interpreting detailed (1:2,000) road profile maps furnished by Estradas de Portugal, SA.
The spatial set of explanatory variables (S) consisted of ten spatial variables (Table 1), including an autocovariate term (Borcard et al. 1992; Heikkinen et al. 2004) and a full third-order polynomial of x and y coordinates (nine spatial variables) so as to account for nonlinear responses:
Before calculating each polynomial term, the x and y coordinates were centred to a zero mean to reduce collinearity between the polynomial terms (Legendre and Legendre 1998). The existence of autocorrelation in all vertebrate group road kills was evaluated using Moran's I. When autocorrelation was detected, further analysis took this into account using an autocovariate term (Segurado et al. 2006).
The autocovariate term (AUTOCOV) considers the response at one road sector as a function of the responses at neighbouring sites. It was considered for each vertebrate road kill data group and for all vertebrates taken together (Augustin et al. 1998; Knapp et al. 2003). This term was computed using the following equations:
and
where w ij is the weighted distance (metres) between the 500-m road segment i centre and the centre of the neighbouring segment j, and y j is equal to the number of road kills along the i segment. The weight distance was calculated by Eq. 2, where d ij is the distance (metres) between the 500-m road segment i centre and the centre of the neighbouring segment j. All the distances used were calculated over the road network (Knapp et al. 2003).
Statistical analysis
For analytical purposes and to avoid a large number of zeros in the final matrix, road fatalities were aggregated into seven ecological groups (Zuur et al. 2007): amphibians, reptiles, passerines, owls, carnivores, prey mammals and hedgehogs. The group prey mammals includes shrews, moles, rodents, rabbits and hares, due to their small sample size, their ecological affinities concerning habitat selection (both tend to concentrate on road verges) and their importance in the trophic net. Hedgehogs were considered separately; due to their spiny body cover, they remain on roads for a longer period of time than other small mammals. Additionally, hedgehogs are considered one of the mammals most affected by road casualties all over the world (Huijser and Bergers 2000). We computed a road kill index for each ecological group and vertebrate class and for all road fatalities taken together to illustrate the frequency of road kills per 1,000 km of road surveyed by year (Clevenger et al. 2003).
To determine comprehensive road kill results and those for each ecological group, we tested (1) for the homogeneity (or heterogeneity) of the number of road casualties by road stretch (SU) and surveys for each year, using Chi-square analysis; (2) for significant differences between 1996 and 2005 in the road mortality pattern (peaks) along the road stretch and throughout the year (monthly samples), using paired Wilcoxon tests and (3) for differences between the years in the number of road causalities along the road stretch and samplings using Mann–Whitney tests (Sokal and Rohlf 1997). All comparisons were performed with SPSS 16.0 TM (SPSS Inc. 2008).
For multivariate analysis, in order to reduce multicollinearity, we removed from further analysis each variable with the lower biological meaning from any pair of variables having a Spearman correlation coefficient higher than ±0.70 (Tabachnick and Fidell 2001). Original variables were transformed to approach normality. We used logarithmic transformation on continuous variables (including response variables) and angular transformation for proportion land cover data (Zar 1999).
Variance partitioning
To evaluate the effects of each explanatory variable set on the seven ecological road kill groups, we used the variation partitioning procedure proposed by Borcard et al. (1992) extended to the three sets of variables and adapted for redundancy analysis (RDA; Liu 1997; Heikkinen et al. 2004). The choice between RDA and canonical correspondence analysis was decided after running a detrended correspondence analysis on the response matrix variables for the 52 road segments for each year's data. The length of the gradient for each year (1.497 and 2.261 for 1996 and 2005, respectively) suggested that a linear method (RDA) was more appropriate (Jongman et al. 1995; ter Braak and Smilauer 2002; Leps and Smilauer 2003).
As a first step, we ran an RDA on each set of putative explanatory variables using a manual selection option and Monte Carlo permutation tests (499 permutations; ter Braak and Smilauer 2002). Only the explanatory variables that contributed significantly (p < 0.1) and improved the fit of RDA models were retained for subsequent analysis (Borcard et al. 1992; Liu 1997; Heikkinen et al. 2004). For each RDA, we also tested the statistical significance of all axes and the sum of all canonical eigenvalues with a Monte Carlo permutation test (499 unrestricted permutations; ter Braak and Smilauer 2002; Leps and Smilauer 2003).
For each year's data, after developing single set models and identifying explanatory variables, we computed three joint models, one for each possible two-set combination. Additionally, we developed a global model that incorporated all the variables selected for each single set model.
This procedure allowed us to decompose the variance of the data into eight components: (a) pure effect of land cover, (b) pure effect of landscape metrics, (c) pure effect of spatial components, (ab) shared effect of habitat cover and landscape metrics, (ac) shared effect of habitat cover and spatial components, (bc) shared effect of landscape metrics and spatial components, (abc) shared effect of the three groups of explanatory variables and (u) unexplained variation.
The variance partitioning procedures were conducted in accordance with the methodology explained in Heikkinen et al. (2004). All multivariate analysis was performed using the program CANOCO version 4.5 (ter Braak and Smilauer 2002).
Results
Road kill data
Over the 52 road surveys (26 per year), a total of 1,352 km of road were covered. We registered 2,073 vertebrate road kills belonging to 87 species (see Appendix). However, for data analysis, we only used 1,922 vertebrate road losses belonging to 75 species, which were then aggregated into the seven ecological groups previously described after removing domestic animals and rare species that could not be included in any of those seven sets.
In 1996, we identified 63 mammals (12 species), 266 birds (29 species), 934 amphibians (10 species) and 63 reptiles (5 species) that had been killed along the road. In 2005, we registered 95 mammals (15 species), 296 birds (32 species), 159 amphibians (8 species) and 43 reptiles (8 species).
When comparing the RKI (number of road kills per 1,000 km driven) between the two studied years, it is worth mentioning that approximately six times as many amphibians and four times fewer hedgehogs were killed in 1996 than in 2005. For the remaining taxa, the RKI values were of the same order of magnitude in the 2 years (Table 2).
Figure 2 illustrates the distribution pattern of road fatalities per road kilometre for each year. In both years, there was significant heterogeneity in the number of road kills along the stretch (χ 2 = 1,148 and χ 2 = 388; df = 51; p < 0.0001, for 1996 and 2005), with road kills tending to be concentrated along the road's northern section, nearest to the Natural Park. In fact, along the first 10 km, which correspond to 38% of the random expected mortality, we found 61% of the road kills for 1996 (1.7 times the expected number) and 52% of the road kills for 2005 (1.4 times the expected number); moreover, this stretch included the main mortality peaks registered each year.

Road kills registered along the national road section (kilometres) on both studied years. Black and grey dashed lines correspond to the average road kills per kilometre in 1996 and 2005, respectively (obtained from Microsoft Excel)
The road kill aggregation peaks along the road were not statistically different between the two surveyed years (Z = −1.194; N = 52 (500 m) segments; p = 0.233, Wilcoxon test; Fig. 2), or within each year (Z = −1.399; N = 52 samples; p = 0.162, Wilcoxon test; Fig. 3).

Comparisons of the monthly road kills for ecological groups, between years: (a) 1996 and (b) 2005. The total number of road kills per month is on the top of each column (obtained from Microsoft Excel)
Significant differences in all vertebrate (Z = −3.417; p = 0.001, Mann–Whitney test), amphibian (Z = −5.753; p < 0.0001, Mann–Whitney test) and hedgehog (Z = −2.352; p = 0.019, Mann–Whitney test) road fatalities (Fig. 2) were identified between surveys. Hedgehogs were the only ecological group which demonstrated significant differences between samplings (Z = −2.956; p = 0.003, Mann–Whitney test; Fig. 3).
Concerning temporal variation, road casualties peaked in rainy months (usually in autumn), revealing the high number of fatalities of amphibians on these occasions. This pattern was particularly marked in 1996 (Fig. 3), the year with higher rainfall. In both years, the summer months usually exhibited the least mortality. In drier months, when losses of amphibians were scarce, road kills generally were higher in 2005 than 1996 (Fig. 3).
Variation partitioning
Among the 48 initial variables, 23 were used for further multivariate analysis, after removing collinear descriptors (see Table 1). Initially, we used the autocovariate term to account for the autocorrelation observed within groups and in the overall mortality. However, this term was excluded during exploratory analysis, due to the high correlation with distance to the NPSSM, which was easier to explain from a biological point of view.
The variables other crops, pastureland and shrubland were significant in 1996 RDA (Table 3), altogether capturing 22.9% of the explained variance in vertebrate road kills (Fig. 4). From the landscape metrics set, distance to the NPSSM, distance to shrubland, distance to forest, length of dirt roads and distance to montado with shrubland (Table 3) were considered significant, explaining 51.3% of the variance (Fig. 4). In the spatial set, the variables selected were longitude coordinates and longitude and latitude coordinate interaction, capturing 24.9% of the variance in data.

Results of variation decomposition for the total ecological vertebrate groups (EVG), shown as fractions of variation explained. Variation of the EVG matrix is explained by three groups of explanatory variables: LC (land cover), LM (landscape metrics) and S (spatial variables), and U is the unexplained variation; a, b and c are unique effects of habitat, landscape factors and spatial variables, respectively, while ab, ac, bc and abc are the components indicating their joint effects (obtained from Microsoft Power Point)
In 2005, other crops, pastureland and urban areas were the variables selected on the land cover set RDA (Table 4), capturing 14.5% of the explained variance in road kills. In the landscape metrics set, four variables were considered significant, distance to the NPSSM, distance to forest, distance to montado with shrubland and distance to water reservoirs (Table 4), capturing 33.0% of the variance. In this year, two spatial group variables were included in the RDA: longitude coordinates and longitude and latitude coordinate interaction, capturing 9.1% of the variance. When considered altogether, the variables selected captured 67.5% of the variance in 1996 and 48.1% in 2005.
The pure land cover effect was almost twice as great in 1996 (5.5%) as in 2005 (2.9%). Another interesting result was the small negative value of two fractions of variance, suggesting synergism between land cover and spatial coordinates in 1996 and landscape metrics and spatial coordinates in 2005 (Liu 1997; Legendre and Legendre 1998). The joint effects of the three variables groups were four times higher in 1996 (8.8%) as in 2005 (2.2%).
Redundancy analysis for 1996
The Monte Carlo test revealed that the first canonical partial axis (F = 61.073, p = 0.002) and all canonical axes together (F = 8.517, p = 0.002) were highly significant. Considering the vertebrate group–environment relationship in 1996, the first two partial RDA axes together captured 95.7% of all the extracted variance (88.6% and 7.1%, respectively). The triplot graph (Fig. 5) shows that, in all groups, road mortality was negatively related to distance to NPSSM, this relationship being particularly defined for prey mammals, passerines and Reptilia. Moreover, the higher mortality of prey mammals, Reptilia and Amphibia was associated with an increase in the area of other crops. Proximity to forests (a decrease in distance to forest) and smaller area of pasturelands promote mortality among prey mammals, passerines, Reptilia and Amphibia. The presence of shrubs was highly associated with owls and hedgehogs fatalities.

Ordination triplot depicting the first two axes of the environmental (total) variables from partial redundancy analysis of the species assemblages in 1996. Environmental variables (land cover, landscape metrics and spatial; grey colour) are represented by solid lines and their acronyms (see Table 1). Ecological groups' locations (black colour) are represented by dashed arrows and their name (Table 4). Road samples are symbolised by black triangles. D_FOREST distance to forest, D_NPSSM distance to Natural Park of Serra São Mamede, D_OAK_SHRUBS distance to oak woodlands with shrubs, D_SHRUBS distance to shrubland, L_DIRT_ROADS length of dirt roads, O_CROPS other crops, PAST pastureland, X longitude coordinates, XY longitude and latitude (coordinates) interaction (obtained from Canoco 4.5)
Wild carnivores mortality tended to increase near forest patches (decreasing with distance to forests), in areas of montado with shrubland and a lower density of dirt roads, and at lower longitudes and to decrease in pastureland areas. Table 3 outlines every variable selected during RDA analysis and its conditional effects. Distance to NPSSM was the strongest predictor of vertebrate mortality. This importance also is stressed in the triplot where it is the variable with the longest arrow.
Redundancy analysis for 2005
The Monte Carlo test for the global RDA performed with all the descriptors revealed that the first canonical partial RDA axis and all canonical axes combined were highly significant (F = 21.002, p = 0.002 and F = 3.883; p = 0.002, respectively). The first two partial axes accounted for 89.3% of all extracted variance (70.4% and 18.9%, respectively). The triplot (Fig. 6) for the 2005 data also reveals the strong negative association between distance to NPSSM and the mortality of prey mammals, passerines, hedgehogs and owls, similar to 1996. On the other hand, wild carnivore causalities present a slightly positive relationship with this variable. Reptilia mortality seems to be less closely related to distance to NPSSM, as it was in 1996. For the Amphibia and Reptilia, the graph shows that increases in casualties tended to occur near water reservoirs and forest patches, with smaller pastureland cover, and at lower longitudes. Other crops as cover appear to influence mortality patterns in the same way as they did in 1996. In 2005, owls sustained an increased number of fatalities near montado with shrubland areas (shorter distance to oak with shrubs). However, for this vertebrate group, and to a lesser extent for hedgehogs, urban areas and longitude and latitude coordinate interaction now presented a strong positive association with road kills.

Ordination triplot depicting the first two axes of the environmental (total) variables from partial redundancy analysis of the species assemblages in 2005. See details in legend of Fig. 5. D_DAM distance to water reservoirs, URBAN urban areas (obtained from Canoco 4.5)
In 2005, the strongest predictor of road fatalities again was distance to NPSSM, with the greatest value for conditional effects (Table 4).
When comparing the results between the 2 years, we noticed that, in the global models, a similar number of variables were selected in each case (ten vs. nine), suggesting a good balance within the statistical analysis. From all the variables selected, seven descriptors remain common to the models for both years: other crops, pastureland, distance to NPSSM, distance to forest, distance to oak with shrubs, longitude and interaction between longitude and latitude. In order to evaluate the predictive power of these variables, we reran the models with each year's data using only the seven common variables (results not shown). We verified that only a minor part of the explained variance was lost (7% in 1996 and 4% in 2005), which strengthens the argument that these common variables influence vertebrate road losses independently of the year (and also of climatic conditions and traffic), and so must be particularly useful for road managers as predictors of fauna fatalities on roads.
We also reran the models from 1 year with the road kill data from the other year, as a way to evaluate model adequacy and robustness (results not shown). Both models demonstrated analogous relationship patterns among road casualties and explanatory variables, and a similar proportion of variance explained as in the original models (47% for the 1996 model with data from 2005 and 62% for the 2005 model with data from 1996). These results suggest that the original models perform well.
Discussion
Global patterns of mortality
Information concerning overall vertebrate road kills is scarce, so that comparisons between our results and other studies are difficult to discuss meaningfully. However, we can state that our road kill numbers are higher—except for mammals—than the ones recorded by Lodé (2000) in Western France, who reported 294 amphibian, 11 reptile, 267 avian and 435 mammalian road kills per 1,000 km driven (see Table 2 for results). Concerning studies that considered different vertebrate groups, Hell et al. (2005) reported 60 mammalian and 72 avian road casualties per 1,000 km in Slovenia, values that are much lower than ours for the same groups (Table 2). Clevenger et al. (2003) reported five mammalian, five avian and 0.74 amphibian road fatalities per 1,000 km in Canada. Finally, Grilo et al. (2009) identified 470 road losses of wild carnivores per 1,000 km in south Portugal. However, they surveyed highways (260 km) and national roads (314 km), and the methodology they used was similar to ours only with the latter. We believe that besides differences in sampling protocols among these studies, our results may reflect the unusually high numbers of individuals and diversity of species that occur in our surveyed area, which is embedded at a crossroads between Atlantic and Mediterranean biogeographic regions located within a hotspot of global and national biodiversity (Myers et al. 2000).
Looking to the temporal distribution of the road casualties in each studied ecological group, the results presented here mainly reflect ecological demands and life history traits, like breeding phenology and dispersal activity (Philcox et al. 1999; Clevenger et al. 2003; Erritzoe et al. 2003). This is particularly true for the most frequently killed vertebrates, the amphibians and passerines. Amphibians are extremely affected by road mortality during their breeding season which, in Mediterranean regions, takes place in autumn and spring (Fig. 3) when rainy events are concentrated (Hels and Buchwald 2001). Passerine road kills occurred mostly from April to September, corresponding to the breeding and dispersal periods of the juveniles in southern Europe (Erritzoe et al. 2003). Road kill patterns in this group also may reflect the greater abundance of food near the roads during summer months, because of agricultural crops. Nevertheless, at least a proportion of passerine road fatalities also may be associated with the higher traffic flow that occurs during summer holidays (Erritzoe et al. 2003).
We identified highly similar spatial and temporal road kill patterns in both years (Figs. 2 and 3). This suggests that the sites where most vertebrates tend to cross the road are not random and can be predicted on the basis of surrounding environmental characteristics, despite changes in climatic and traffic conditions that occurred between 1996 and 2005 (see further sections). Animals show some fidelity to landscape corridors, especially inside their home ranges. Consequently, road kill aggregations may persist over generations (Clevenger et al. 2003).
Land cover, landscape metrics and spatial effects on spatial road kill patterns
We conducted our study to improve our understanding in the effects of land cover, landscape metrics and spatial influences on road casualties. To our knowledge, ours was the first study to use variance partitioning methodologies to evaluate the relative influence of these descriptors sets on vertebrate road kill patterns. Moreover, it is the first to have compared road fatalities along the same stretch of road between two time periods several years (9 years) apart.
The overall results of the variance partition analyses performed for the 2 years demonstrate similar tendencies over the 2 years, despite differences in the proportion of total variance explained. Moreover, seven variables were common to both years' models, and models incorporating just these common variables exhibit analogous results to the original ones, suggesting that these variables are particularly useful for the prediction of road kills on similar roads crossing the same types of environment.
Landscape metrics always were the most important set, followed by spatial features, explaining vertebrate road fatalities. In both years, land cover was the set that explained road fatality patterns least. This is a surprising result, because we would expect a greater influence of the land cover set. Indeed, some studies implicate habitat as having the highest influence on road casualties (e.g. Clevenger et al. 2003) because animals tend to cross roads near optimum habitat patches. However, due to habitat fragmentation, the remaining patches become smaller and more isolated, promoting longer movement among patches when animals are looking for high quality habitats, increasing the probability of being hit by a car outside that optimum habitat (Forman et al. 2003; Alexander et al. 2005; Eigenbrod et al. 2008; Shepard et al. 2008; Grilo et al. 2009).
Regarding the land cover variables, other crops were associated with an increase in casualties among prey mammals and passerines (Figs. 5 and 6) in both years. Small mammals living in Mediterranean areas, such as rodents, shrews, moles and rabbits and many small birds, often are favour by small orchards, vineyards and vegetable gardens, which provide water, shelter and food. One would expect road fatalities to be enhanced if these cultures are located in the vicinity of a road (Bennett 1990; Bellamy et al. 2000; Bautista et al. 2004; Rytwinski and Fahrig 2007). The positive association between owls and prey mammals road losses obtained in both years (Figs. 5 and 6) suggests that owl road fatalities may, at least, partially reflect an attraction to the road vicinity due to higher prey availability. Here, they will be exposed to vehicles while flying from one perch to another (Hernandez 1988; Gomes et al. 2008). In fact, a study recently conducted on another road in southern Portugal has found a higher concentration of rabbits and other small mammals on road verges relative to surrounding areas (Marques et al., unpublished data).
Distance to NPSSM was the most important of all of the landscape metrics variables, being negatively correlated with the majority of the vertebrate fatalities in both years. This association may reflect the distance to a mountain range in the NPSSM, where higher rainfall and humidity provide greater water availability than in surrounding flat areas. In a Mediterranean context, water supply is critical for most vertebrates, particularly during the summer months, although amphibians chiefly are affected. Probably, the elevated number of road kills that occurred in the SU closer to the Park was related to the expected southwards-decreasing water gradient (Figs. 5 and 6). These results reveal the higher abundances of amphibians near the Park, as elevated numbers of these species are expected in moist areas (Beja and Alcazar 2003; Mazerolle et al. 2005; Benayas et al. 2006; Eigenbrod et al. 2008).
Carnivore road casualties were the least strongly associated with distance to NPSSM. In 2005, this group exhibited a different pattern than the one registered in 1996. In fact, in 2005, most road losses of wild carnivores took place along the final section of the road, away from the Park but in proximity to the largest stream with a well-preserved riparian corridor (Ribeira Grande) that crosses the study area. Virgós (2001) and Matos et al. (2008) emphasised the importance of riparian woodlands for carnivores, especially when crossing open lands, as in our study area. This effect must be particularly enhanced in dry years when animals actively seek water resources. The most important factors affecting carnivore road fatalities may be related to physiological constraints, mainly due to habitat and/or environmental limitations. In 2005, the dry conditions might have caused carnivores to gather nearest to riparian areas, increasing their probability of getting killed when those areas are located near roads. On the other hand, as water availability was not a limiting factor in 1996, the presence of optimal habitat (e.g. oak woodlands) for carnivores scattered along the roadside may explain the dispersed distribution of road casualties of these species during that year (Fig. 5; Grilo et al. 2009).
In addition, it is worth mentioning that none of the descriptors related to the road profile included in the landscape metrics set were selected as significant in either year. Clevenger et al. (2003) showed that roadside topography strongly influences small vertebrate road kills, especially those with low vagility. These differences in the results may reflect the fact that our studied road crosses a flattened landscape and presents only small profile dissimilarities along the entire surveyed stretch.
The relatively large influence of the spatial set of variables on road losses is in agreement with the spatial autocorrelation found in our data for both years. Despite the great influence of latitude, revealed by the importance of distance to NPSSM, longitude and the interaction of longitude with latitude also were important. Highest interaction values also occurred near the NPSSM (higher latitudes and longitudes), which may explain the strong positive relationship of this variable with mortality in several vertebrate groups (Figs. 5 and 6). This effect was slightly more prominent in 1996, revealing a greater spatial structure among road kills during this year. The spatial effects also may incorporate the effect of other factors—such as local resources, predation and competition—that may affect species abundance and or space use, which also may define the locations of vertebrate road casualties (Legendre 1993). However, it was not possible to assess the relative importance of these factors with our data.
What caused the difference between 1996 and 2005?
Despite of the differences in the percentage of explained variances, the models for each year are similar and have in common seven explanatory variables. However, two major changes occurred between 1996 and 2005: (1) a strong decrease in rainfall and (2) a high increase in traffic volume. Most animals have a high physiological dependence on water, this being particularly strong for amphibians as they depend entirely upon it to complete their life cycles (Cushman 2006). Amphibians also show episodic massive road fatalities, usually coincident with migrations to and from spawning sites that can take place within a single night (Clevenger et al. 2003; Cushman 2006).
A dry year, like 2005, affects amphibians life cycles, inhibiting their reproduction and therefore limiting their movements (Benayas et al. 2006; Cushman 2006), which in turn lowers road kill risk. Nevertheless, not only amphibians should be affected by weather. Extreme meteorological conditions, such as the severe drought verified in Portugal in 2005, may have exerted a sizeable impact upon the population density and space use of most species as a response to restricted resources, particularly water (Vermeulena and Opdam 1995; Erritzoe et al. 2003; Araújo et al. 2006). The demand for water in dry conditions must explain the greater importance of distance to water reservoirs in structuring vertebrate road fatalities in 2005. This is particularly obvious for amphibians, but also for carnivores as previously explained.
In 1996, the abundance of water over the entire study area was not a limiting factor, as the rivers and little streams kept water running throughout the year. The extremely dry conditions experienced in 2005 may have contributed to the higher mortality of most vertebrate groups, relative to 1996 values, except for amphibians and reptiles. The moister conditions and more abundant vegetation growth on verges, versus surrounding areas, translate into an increase in the availability of cover and food, which attracts animals to the roadside, increasing their likelihood of being hit (Bellamy et al. 2000; Sherwood et al. 2002; Erritzoe et al. 2003).
On the other hand, traffic load and speed also are important factors determining road losses (Kaczensky et al. 2003; Baker et al. 2004; Jaeger et al. 2005; Grilo et al. 2009), and they also might have played a role in the higher mortality of many vertebrate groups recorded in 2005. Indeed, along the studied stretch of road, traffic increased roughly 150% between 1996 and 2005. Most species or groups seemed to be particularly vulnerable to this traffic increase. For instance, four times the hedgehog road fatalities occurred in 2005 as in 1996 (Huijser and Bergers 2000), and twice as many owls were killed (Fajardo 2001). Jaarsma et al. (2006) warmed of the effect of higher speeds, which may increase the chance of an animal being hit by a car while attempting to cross a road. This may explain the increase in avian and mammalian road casualties verified in 2005 (Erritzoe et al. 2003; Van Langevelde and Jaarsma 2004). Additionally, the road had been enlarged roughly 2 years prior to 1996. This roadside invasion, which resulted in the destruction of verges, may have temporarily decreased the density of hedgehogs and rabbits on verges (Rondinini and Doncaster 2002; Bautista et al. 2004), thereby lowering their probability of being killed during the first year of our study. Finally, we also should consider that the traffic load should be greater near Portalegre, due to the daily influx of people from the surrounding villages who work there, which also may have contributed to the higher number of road kills for all groups along the first 10 km of the studied stretch of road (Fig. 2). Originally, our goal concerning comparisons between 1996 and 2005 was to evaluate the effect of traffic intensification on vertebrate road fatalities. The large differences in rainfall between these 2 years confounded our results and did not allow for the identification and discussion of any pure traffic effect on road fatalities.
Conclusions
This investigation provided the first results and discussion regarding the relative roles of land cover, landscape metrics and spatial effects, through variance partitioning, on all terrestrial vertebrate class road kill patterns, taken together, within a Mediterranean context. We have demonstrated that landscape metrics are the most important group of factors influencing vertebrate road kills. This means that, when choosing road corridors, the landscape structure, and not only its land cover, must be taken into account as a whole if we aim to effectively reduce the risk of fauna fatalities. The high density of road mortality found in our study justifies the urgency of implementing mitigation measures on existing roads. Hotspots of mortality on the road must be identified on the basis of field surveys and modelling. Mitigation measures must take place for each vertebrate group and must be defined specifically taking into account species behaviour and ecological needs (Caro et al. 2000; Underhill and Angold 2000; Iuell et al. 2003; Grilo et al. 2008).
In both models, 9 years apart, the same seven variables influenced road mortality, despite traffic enhancement and a drier climate in the latter year. As explained in detail above, these predictors appear to be ecologically relevant for most of the vertebrate groups we studied, which might mean that, to be effective, mitigation measures should focus on these predictors.
Notwithstanding these main contributions, more investigation is needed to disentangle the pure effects of individual factors (e.g. road characteristics, landscape features, traffic volume and speed and weather conditions) on fauna road mortality patterns. We hope that our results will provide insight into prioritising and choosing the best options when several become available.
References
Alexander SM, Waters NM, Paquet PC (2005) Traffic volume and highway permeability for a mammalian community in the Canadian Rocky Mountains. Can Geogr 49:321–331
Ament R, Clevenger AP, Yu O, Hardy A (2008) An assessment of road impacts on wildlife populations in U.S. National Parks. Environ Manage 42:480–496
Araújo MB, Thuiller W, Pearson RG (2006) Climate warming and the decline of amphibians and reptiles in Europe. J Biogeogr 33:1712–1728
Augustin NH, Mugglestone MA, Buckland ST (1998) The role of simulation in spatially correlated data. Environmetrics 9:175–196
Baker PJ, Harris S, Robertson CPJ, Saunders G, White PCL (2004) Is it possible to monitor mammal population changes from counts of road traffic casualties? An analysis using Bristol's red foxes Vulpes vulpes as an example. Mammal Rev 34(1):115–130
Bautista LM, García JT, Calmaestra RG, Palacín C, Martín CA, Morales MB, Bonal R, Viñuela J (2004) Effects of weekend road traffic on the use of space by raptors. Conserv Biol 18(3):726–732
Beja P, Alcazar R (2003) Conservation of Mediterranean temporary ponds under agricultural intensification: an evaluation using amphibians. Biol Conserv 114:317–326
Bellamy PE, Shore RF, Ardeshir D, Treweek JR, Sparks TH (2000) Road verges as habitats for small mammals in Britain. Mammal Rev 30(2):131–139
Benayas JMR, De La Montaña E, Belliure J, Eekhout XR (2006) Identifying areas of high herpetofauna diversity that are threatened by planned infrastructure projects in Spain. J Environ Manage 79:279–289
Bennett AF (1990) Habitat corridors and the conservation of small mammals in a fragmented forest environment. Landscape Ecol 4:109–122
Boarman WI, Sazaki M (2006) A highway's road–effect zone for desert tortoises (Gopherus agassizii). J Arid Environ 65:94–101
Borcard D, Legendre P, Drapeau P (1992) Partialling out the spatial component of ecological variation. Ecology 73:1045–1055
Cabral MJ, Almeida J, Almeida PR, Dellinger T, Ferrand de Almeida N, Oliveira ME, Palmeirim JM, Queiroz AI, Rogado L, Santos-Reis M (2005) Livro vermelho dos vertebrados de Portugal. Instituto da Conservação da Natureza, Lisboa
Caro TM, Shargel JA, Stoner CJ (2000) Frequency of medium-size mammals road kills in an agricultural landscape in California. Am Mid Nat 144:362–369
Carr L, Fahrig L (2001) Effect of road traffic on two amphibian species of differing vagility. Conserv Biol 15:1071–1078
Clarke GP, White PCL, Harris S (1998) Effects of roads on badger Meles meles populations in south-west England. Biol Conserv 86:117–124
Clevenger AP, Chruszcz B, Gunson KE (2003) Spatial patterns and factors influencing small vertebrate fauna road-kill aggregations. Biol Conserv 109:15–26
Coffin AW (2007) From roadkill to road ecology: a review of the ecological effects of roads. J Transp Geogr 15:396–406
Conard JM, Gipson PS (2006) Spatial and seasonal variation in wildlife–vehicle collisions. Prairie Nat 38(4):251–260
Crooks KR, Sanjayan M (2006) Connectivity conservation. Cambridge University Press, UK
Cushman SA (2006) Effects of habitat loss and fragmentation on amphibians: a review and prospectus. Biol Conserv 128:231–240
Cushman SA, McGarigal K (2002) Hierarchical, multi-scale decomposition of species–environment relationships. Landscape Ecol 17:637–646
Eigenbrod F, Hecnar SJ, Fahrig L (2008) The relative effects of road traffic and forest cover on anuran populations. Biol Conserv 141:35–46
Eikie PC, Rempel RS, Carr AP (1999) Patch analyst user’s manual. A tool for quantifying landscape structure. NWST technical manual TM–002. Northwest Science and Technology, Ontario
EPE (2005) Recenseamento do tráfego – Portalegre. Estradas de Portugal, Lisboa
Erritzoe J, Mazgajski TD, Rejt L (2003) Bird casualties on European roads—a review. Acta Ornithol 28(2):77–93
ESRI (1999) ArcView GIS 3.2 ESRI. Environmental Systems Research Institute, New York
Fahrig L, Pedlar J, Pope S, Taylor P, Wegner J (1995) Effect of road traffic on amphibian density. Biol Conserv 75:177–182
Fajardo I (2001) Monitoring of non-natural mortality in the barn owl (Tyto alba), as an indicator of land use and social awareness in Spain. Biol Conserv 97:143–149
Ferreras P, Aldama JJ, Beltran JF, Delibes M (1992) Rates and causes of mortality in a fragmented population of Iberian lynx Felis pardina Temminck, 1824. Biol Conserv 61:197–202
Findlay CS, Lenton J, Zheng LG (2001) Land-use correlates of anuran community richness and composition in southeastern Ontario wetlands. Ecoscience 8:336–343
Forman RTT (1998) Road ecology: a solution for the giant embracing us. Landsc Ecol 13:iii–v
Forman RTT, Alexander LE (1998) Roads and their major ecological effects. Annu Rev Ecol Syst 29:207–231
Forman RTT, Deblinger RD (2000) The ecological road–effect zone of a Massachusetts (USA) suburban highway. Conserv Biol 14:36–46
Forman RTT, Reineking B, Hersperger AM (2002) Road traffic and nearby grassland bird patterns in a suburbanizing landscape. Environ Manage 29(6):782–800
Forman RTT, Sperling D, Bissonette JA, Clevenger AP, Cutshall CD, Dale VH et al (2003) Road ecology: science and solutions. Island Press, Washington
Gomes L, Grilo C, Silva C, Mira A (2008) Identification methods and deterministic factors of owl roadkill hotspot locations in Mediterranean landscapes. Ecol Res 24:355–370
Grilo C, Bissonette J, Santos-Reis M (2008) Response of carnivores to existing highway culverts and underpasses: implications for road planning and mitigation. Biodivers Conserv 17:1685–1699
Grilo C, Bissonette J, Santos-Reis M (2009) Spatial-temporal patterns in Mediterranean carnivore road casualties: consequences for mitigation. Biol Conserv 142:301–313
Heikkinen RT, Luoto M, Virkkala R, Rainio K (2004) Effects of habitat cover, landscape structure and spatial variables on the abundance of birds in an agricultural–forest mosaic. J App Ecol 41:824–835
Hell P, Plavý R, Slamečka J, Gašparík J (2005) Losses of mammals (Mammalia) and birds (Aves) on roads in the Slovak part of the Danube Basin. Eur J Wildl Res 51:35–40
Hels T, Buchwald E (2001) The effect of road kills on amphibian populations. Biol Conserv 99:331–340
Hernandez M (1988) Road mortality of the little owl (Athene noctua) in Spain. J Raptor Res 22:81–84
Huijser MP, Bergers PJM (2000) The effect of roads and traffic on hedgehog (Erinaceus europaeus) populations. Biol Conserv 95:111–116
IEP (2000) Recenseamento de tráfego de 1995 e 1996 e Planos de Estradas. Instituto de Estradas de Portugal, Lisboa
Iuell B, Bekker GJ, Cuperus R, Dufek J, Fry G, Hicks C, Hlavac V, Keller VB, Rosell C, Sangwine T, Torslov N, Wandall B le Maire (eds) (2003) Cost 341 habitat fragmentation due to transportation infrastructure. Wildlife and traffic: a European handbook for identifying conflicts and designing solutions. European Co-operation in the Field of Scientific and Technical Research
IUCN (2007) 2007 IUCN Red List of Threatened Species. Available at http://www.iucnredlist.org
Jaarsma CF, Van Langevelde F, Botma H (2006) Flattened fauna and mitigation: traffic victims related to road, traffic, vehicle, and species characteristics. Transp Res D 11:264–276
Jaeger JAG, Bowman J, Brennan J, Fahrig L, Bert D, Bouchard J, Charbonneau N, Frank K, Gruber B, von Toschanowitz KT (2005) Predicting when animal populations are at risk from roads: an interactive model of road avoidance behaviour. Ecol Model 185:329–348
Jongman RHG, ter Braak CJF, van Tongeren EFR (1995) Data analysis in community and landscape ecology. Cambridge University Press, Cambridge
Kaczensky P, Knauer F, Krže B, Jonozovič M, Adamič M, Gossow H (2003) The impact of high speed, high volume traffic axes on brown bears in Slovenia. Biol Conserv 111:191–204
Knapp RA, Matthews KR, Preisler HK, Jellison R (2003) Developing probabilistic models to predict amphibian site occupancy in a patchy landscape. Ecol Appl 13(4):1069–1082
Legendre P (1993) Spatial autocorrelation: trouble or a new paradigm? Ecology 74:1659–1673
Legendre P, Legendre L (1998) Numerical ecology, second English ed. Elsevier Science BV, Amsterdam
Leps J, Smilauer P (2003) Multivariate analysis of ecological data using CANOCO. Cambridge University Press, Cambridge
Liu Q (1997) Variation partitioning by partial redundancy analysis (RDA). Environmetrics 8:75–85
Lodé T (2000) Effect of a motorway on mortality and isolation of wildlife populations. Ambio 29:163–166
Matos HM, Santos MJ, Palomares F, Santos-Reis M (2009) Does riparian habitat condition influence mammalian carnivore abundance in Mediterranean ecosystems? Biodivers Conserv 18:373–386
Mazerolle M, Huot M, Gravel M (2005) Behaviour of amphibians on the road in response to car traffic. Herpetologica 61(4):380–388
McGregor RL, Bender BJ, Fahrig L (2008) Do small mammals avoid roads because of the traffic? J App Ecol 45:117–123
Myers N, Mittermeier RA, Mittermeier CG, da Fonseca GAB, Kent J (2000) Biodiversity hotspots for conservation priorities. Nature 403:853–858
Philcox CK, Grogan AL, Macdonald DW (1999) Patterns of otter (Lutra lutra) road mortality in Britain. J App Ecol 36:748–761
Ramp D, Caldwell J, Edwards KA, Warton D, Croft DB (2005) Modelling of wildlife fatality hotspots along the snowy mountain highway in New South Wales, Australia. Biol Conserv 126:474–490
Ramp D, Wilson VK, Croft DB (2006) Assessing the impacts of roads in peri-urban reserves: road-based fatalities and road usage by wildlife in the Royal National Park, New South Wales, Australia. Biol Conserv 129:348–359
Rico A, Kindlmann P, Sedláček F (2007) Barriers effects of roads on movements of small mammals. Folia Zool 56(1):1–12
Rondinini C, Doncaster CP (2002) Roads as barriers to movement for hedgehogs. Funct Ecol 16:504–509
Rytwinski T, Fahrig L (2007) Effects of road density on abundance of white-foot-mice. Landscape Ecol 22:1501–1512
Saeki M, Macdonald DW (2004) The effects of traffic on the raccoon dog (Nyctereutes procyonoides viverrinus) and other mammals in Japan. Biol Conserv 118:559–571
Segurado P, Araújo MB, Kunin WE (2006) Consequences of spatial autocorrelation for niche-based models. J App Ecol 43:433–444
Seiler A (2005) Predicting locations of moose–vehicle collision in Sweden. J Appl Ecol 42:371–382
Shepard DB, Dreslik MJ, Jellen BC, Phillips CA (2008) Roads as barriers to animal movement in fragmented landscapes. Anim Conserv 11:288–296
Sherwood B, Cutler D, Burton J (eds) (2002) Wildlife and roads: the ecological impact. Imperial College Press, London
Smith-Patten BD, Patten MA (2008) Diversity, Seasonality, and context of mammalian roadkills in the southern Great Plains. Environ Manage 41:844–852
Sokal RR, Rohlf JR (1997) Biometry: the principles and practice of statistic in biological research, 3rd edn. W. H. Freeman and Company, Nova York
SPSS (2008) SPSS for Windows 16.0. (Software for PC). SPSS Inc, Chicago
Tabachnick BG, Fidell LS (2001) Using multivariate statistics. Allyn and Bacon, Boston
ter Braak CJF, Smilauer P (2002) CANOCO reference manual and CanoDraw for Windows user's guide: software for canonical community ordination version 4.5. Microcomputer power, Ithaca
Trombulak SC, Frissell CA (2000) Review of ecological effects of roads on terrestrial and aquatic communities. Conserv Biol 14:18–30
Underhill JE, Angold PG (2000) Effects of roads on wildlife in an intensively modified landscape. Environ Rev 8:21–39
Van Langevelde F, Jaarsma CF (2004) Using traffic flow theory to model traffic mortality in mammals. Landscape Ecol 19:895–907
Vermeulena HJW, Opdam FM (1995) Effectiveness of roadside verges as dispersal corridors for small ground-dwelling animals: asimulation study. Landsc Urban Plan 31:233–248
Virgós E (2001) Relative value of riparian woodlands in landscapes with different forest cover for medium-sized Iberian carnivores. Biodivers Conserv 10:1039–1049
Vos CC, Chardon JP (1998) Effects of habitat fragmentation and road density on the distribution pattern of the Moor frog Rana arvalis. J App Ecol 35:44–56
Zar JH (1999) Biostatistical analysis, 4th edn. Prentice Hall, Inc, Upper Saddle River
Zuur AF, Leno EN, Smith GM (2007) Analysing ecological data. Springer, UK
Acknowledgements
We are thankful to several investigators who helped in different stages of road survey: Carmo Silva, Eduardo Sequeira, Hugo Casco, Nuno Soares, Ondina Giga and Sandra Alcobia. We thank Mafalda Costa for helping in data analysis. We are also grateful to Estradas de Portugal SA for traffic data and to Ana Galantinho by the first English revue. We are grateful to the two anonymous reviewers for their valuable comments. This study was supported by Unity of Biology Conservation, University of Évora and Institute for the Conservation of Nature and Biodiversity.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by H. Kierdorf
Appendix
Appendix
Rights and permissions
About this article
Cite this article
Carvalho, F., Mira, A. Comparing annual vertebrate road kills over two time periods, 9 years apart: a case study in Mediterranean farmland. Eur J Wildl Res 57, 157–174 (2011). https://doi.org/10.1007/s10344-010-0410-0
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10344-010-0410-0