
Abstract
Environmental flows represent a legal mechanism to balance existing and future water uses and sustain non-use values. Here, we identify current challenges, provide examples where they are important, and suggest research advances that would benefit environmental flow science. Specifically, environmental flow science would benefit by (1) developing approaches to address streamflow needs in highly modified landscapes where historic flows do not provide reasonable comparisons, (2) integrating water quality needs where interactions are apparent with quantity but not necessarily the proximate factor of the ecological degradation, especially as frequency and magnitudes of inflows to bays and estuaries, (3) providing a better understanding of the ecological needs of native species to offset the often unintended consequences of benefiting non-native species or their impact on flows, (4) improving our understanding of the non-use economic value to balance consumptive economic values, and (5) increasing our understanding of the stakeholder socioeconomic spatial distribution of attitudes and perceptions across the landscape. Environmental flow science is still an emerging interdisciplinary field and by integrating socioeconomic disciplines and developing new frameworks to accommodate our altered landscapes, we should help advance environmental flow science and likely increase successful implementation of flow standards.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Societies exploit the benefits of freshwater and place extensive stress on aquatic ecosystems. River modification was documented as early as 5000 years ago via civilization of ancient Egypt and Mesopotamia (Wootton 1990). Since that time, aquatic ecosystems have been channelized, dammed, dredged, leveed, and pumped to reduce flooding, promote river commerce, exploit hydropower and meet industrial, agricultural, and municipal needs. Although human needs are a necessary component of water use, the extensive pressures on our global aquatic systems have become clear with approximately 20 % of all freshwater fishes now listed as threatened or endangered due to alteration and destruction of lotic systems (Moyle and Leidy 1992; Naiman et al. 2002).
Aquatic ecosystems are extremely diverse but are threatened by human landscape alteration. Freshwaters systems, despite not only covering a small proportion of the Earth’s surface, are extremely ecologically diverse (Dudgeon et al. 2006) but also some of the most heavily affected by human activity (Malmqvist and Rundle 2002). River regulation has resulted in over half of global freshwater resources being used by humans (Jackson et al. 2001), while more than 50 % of the world’s large rivers are fragmented by dams (Nilsson et al. 2005). Streamflow alteration to freshwater systems, combined with additional threats, has negatively affected coastal ecosystems (e.g., estuaries, salt marshes, and mangrove forests). Over three billion people (around 40 % of the world’s population) live within 100 km of a coastline with growth rates estimated at 16 % per decade (Millennium Ecosystem Assessment 2008). In addition to the direct effects of concentrated human populations, coastal ecosystems are profoundly influenced by damming and other inland anthropogenic activities (Montagna et al. 2002).
Anthropogenic threats to aquatic ecosystems are generally organized into five main categories: overexploitation, pollution, habitat degradation and loss of connectivity, invasive species, and flow modification (see Fig. 1, Dudgeon et al. 2006). The relative importance of these factors is spatially variable and differs among species (Richter et al. 1997). There is also recognition that multiple stressors often interact to affect aquatic organisms (Miller et al. 1989). The legacy of human alteration of freshwater systems is extensive (Downs and Gregory 2004), and although there has been progress in addressing certain issues (e.g., water quality improvements in some developed nations; Allan and Flecker 1993), future changes in climate, population growth, and continued land-use alteration are likely to further exacerbate stress on aquatic ecosystems (Sala et al. 2000). In response to these pressures, freshwater organisms are exhibiting higher extinction rates than their terrestrial counterparts (Ricciardi and Rasmussen 1999; Revenga et al. 2000; Jenkins 2003), with certain species groups disproportionally affected (e.g., Strayer et al. 2004; Dudgeon et al. 2006).

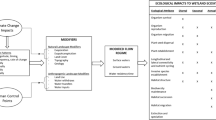
Major topics where research advances would benefit environmental flow (e-flow) science: existing environmental frameworks and implications for highly modified landscapes, interactions of water quality and quantity, invasive species, identifying the non-use economic values, and the spatial distributions of attitudes and perceptions of environmental flows. The primary research recommendations are listed in bullets next to each topic
The relationship between flow regime and the aquatic biota is pervasive and multifaceted, to the extent that discharge has been described as a ‘master variable’ controlling stream ecosystems and driving ecological processes (Fig. 1, Hart and Finelli 1999; Power et al. 1995). Both directly and indirectly, the flow regime affects other regulatory factors such as water quality and temperature thereby regulating ecological integrity (Poff et al. 1997). Discharge is a key driver of physical habitat providing a template determining both taxonomical and functional community structure (Poff and Allan 1995; Bunn and Arthington 2002; Bêche et al. 2006). Disturbance events promote habitat complexity and patchiness through lateral and longitudinal connectivity, and natural fragmentation (Lake 2000). Sediment and solutes are transported through the system creating spatial and temporal gradients in physicochemical conditions. Discharge patterns maintain the river channel (Gordon et al. 2004) and shape the transfer of energy throughout the river system longitudinally (e.g., River Continuum Concept, Vannote et al. 1980), laterally (e.g., Flow Pulse Concept, Junk et al. 1989), and vertically (e.g., Hyporheic Corridor Concept, Stanford and Ward 1993). Discharge also stimulates biotic processes like nutrient uptake and decomposition (Doyle et al. 2005) and predator–prey interactions (Hart and Finelli 1999).
There is an obvious link between aquatic biota and human use via fishing but there are also various non-use activities affected by environmental flow decisions including recreation (e.g., canoeing, birding). In North America, fishing generates $100 billion each year to local economies (American Sportfishing Association 2011) and these economic returns have arguably been one of the most common recreational values quantified relative to environmental flows or water use (Loomis et al. 2003; Katz 2006). In European rivers, the link between high flows, upstream migration (Solomon and Sambrook 2004; Thorstad et al. 2008), and recreational angling catches of Atlantic salmon Salmo salar (Alabaster 1970; Aprahamian and Ball 1995) has been highlighted, driving an interest in using artificial water releases to stimulate upstream movement (e.g., Thorstad and Heggberget 1998; Alfredsen et al. 2012). However, many other river recreation activities are growing in popularity with considerable overlap in participation. For example, Nadel (2005) indicated that >82 million USA residents hunted, fished, and viewed wildlife and many of the activities were in combination. In 2003, an estimated 72 million Americans participated in recreation boating activities and over $200 million was spent on the purchase of canoes and kayaks (Nadel 2005). It is anticipated that canoeing alone will increase substantially by 2050 (Bowker et al. 1999). The perceived quality of these activities will be affected by environmental flow designations. Brown et al. (1991) found that of 25 reviewed studies (primarily related to canoeing and kayaking), all found recreational quality increased with discharge until some threshold was reached where quality declined. The thresholds reached varied with activity and experience level of the recreationist. As river and lake recreation increases, these activities may benefit local economies, particularly in rural areas. For example, the development of recreation trails is a form of tourism with substantial economic benefit. The Northern Forest Canoe Trail of New England, USA, attracted 90,000 visitors in 2006 alone and generated approximately 12 million dollars for local economies (Pollock et al. 2012). The growing popularity and value of these activities will no doubt contribute to the increasing complexity of environmental flow decisions, particularly as pressures increase on our limited aquatic resources.
Future changes in climate and population growth will amplify stress on global water resources (Duda and El-Ashry 2000; Sheffield and Wood 2008; Vörösmarty et al. 2010) making environmental flows an important component of management strategies. Environmental flows represent a legal mechanism that is used to balance providing for existing water uses, meeting the needs of future uses, and maintaining aquatic ecosystems and other non-use values (e.g., recreation). Although the legal framework varies geographically, environmental flows are defined as flows necessary to sustain aquatic ecosystems and the ‘human livelihoods and well-being that depend on these ecosystems’ (Brisbane Declaration 2007). The seemingly simple question of how much water should be left in an aquatic system is compounded by a multitude of societal demands, policies, and needs for advances in our understanding of various ecological interactions. The objective of this paper was to identify the major threats or areas where scientific advancement would provide the most benefit to successful implementation of environmental flows. We identified five major topics where research would be needed to advance environmental flow science: existing environmental frameworks and implications for highly modified landscapes, interactions of water quality and quantity, invasive species, identifying the non-use economic values, and the spatial distributions of attitudes and perceptions of environmental flows (Fig. 1).
Existing Environmental Frameworks
An extensive list of environmental flow frameworks has emerged with the developing discipline of environmental or instream flow science. The Instream Flow Council places the techniques in three basic categories: standard setting, incremental, and monitoring/diagnostic, although some hybrids certainly exist (Annear et al. 2004). Standard setting techniques (e.g., Tennant method) are often policy-driven and set ‘rules’ to define flow limits (Annear et al. 2004). They are advantageous because of simplicity and cost; however, there are many disadvantages including failure to maintain existing fisheries (Stalnaker 1993; Stalnaker and Wick 2000). Incremental methods (e.g., physical habitat simulation) are site-specific field evaluations of alternative flow regimes. Incremental approaches rely on measurements of river ecohydrological conditions at incremental discharges to analyze river discharge-stage-ecological (biotic and abiotic) relationships (Annear et al. 2004). These methods are some of the most widely applied but can sometimes be costly and time-consuming (Annear et al. 2004) and have resulted in mixed outcomes when evaluated. Gore and Nestler (1988) provide an overview of some common criticisms. Lastly, monitoring methods (e.g., Range of Variability Approach) assess temporal changes in biotic or abiotic conditions related to flow alteration (Annear et al. 2004). The methods vary on the focus of the specific resource component and have been categorized by Annear et al. (2004): hydrology, biology, geomorphology, connectivity, and water quality.
Each of the environmental flow frameworks may be applicable under a variety of societal and geographic settings; however, the relatively recent move toward regional water planning has emphasized a process that includes hydrologic properties, stream classification, and biological responses. This framework is based on the idea accepted by the ecological community that organisms have evolved with the natural flow regime (Poff et al. 1997) and that the relationships between organisms and flow provide a basis for our understanding of how flow alteration will affect the biological community. This led to the development of a subdiscipline focusing on stream classification based on hydrologic properties (e.g., Belmar et al. 2011; Olden et al. 2012; Reidy Liermann et al. 2012).
Streams are classified by similar hydrologic properties as a foundation for applying streamflow recommendations across classes of streams (i.e., negating the need for site-specific flow evaluations unless a follow-up is needed or desired). The ecological limit of flow alteration (Poff et al. 2010) is a holistic planning effort (Richter et al. 2012) that was developed to provide quantitative information to support environmental flows at a regional scale. In addition to stream classification, this approach defines the ‘natural flow regime’ for the region, assesses the level of alteration to hydrologic conditions, and establishes univariate flow-ecology relationships as a foundation for societal and policy discussions (Poff et al. 2010).
Two recent criticisms of the aforementioned process include: (1) univariate relationships may be insufficient to develop predictable flow-ecology relationships due to other environmental factors (e.g., morphology and temperature; McManamay et al. 2013) and the confounding relationship among multiple flow components (Davis and Brewer 2014) and (2) questionable application to rivers and/or regions that have a highly altered landscape (e.g., Great Plains, Davis and Brewer 2014), and locations where uncertainty is high and physicochemical conditions are extremely unique (e.g., tailwaters; McManamay and Bevelhimer 2013). This leads to the inevitable conclusion that comparing the flow regime to ‘natural’ conditions may not be appropriate or feasible in many highly altered landscapes of the world and represents an area where future research efforts would benefit river managers. Often, anthropogenic development pre-dates available flow data, requiring modeling approaches to extrapolate from nearby unregulated catchments (e.g., the River Tay, UK, Birkel et al. 2014). Further, in many cases, historical or pre-disturbance flows are not an option, because water volume is limited, overallocated, or is serving multiple purposes (e.g., hydropower and recreation). For example, water allocation in Lake Tenkiller, Oklahoma is over 100 % allocated; therefore, current water law would prevent anything resembling the natural flow regime in the lower river. In addition, the regulatory context may be so complex and multifaceted that recommendations on the basis of the natural flow regime are impractical or target the wrong regulatory entity (e.g., least impactful stressor; Pearsall et al. 2005).
Needs for Highly Altered Landscapes
The science of environmental flows is intended to balance the multidimensional nature of shared water resources in a human-dominated world. Even still, as human population growth continues and water supplies become limited, environmental flow science will be challenged to ensure river flows sustain aquatic ecosystems within highly altered landscapes. In part, this is due to the scale of the problem (i.e., the global extent of hydrologic alteration).
Dramatic alterations to waterbodies and their surrounding landscape have left very few rivers in their natural hydrologic state (Fig. 2). For example, 56 % of stream miles within the USA have habitats that are estimated to be at least moderately degraded and 27 % is estimated to be at least highly degraded (NFHP 2010). River flows have either been altered directly (i.e., channelization and impoundment) or indirectly (i.e., landscape modification). Over 50 % of the global landscape has been transformed or degraded as a result of human activity (Vitousek et al. 1986). Irrigation is the largest consumer of water around the world (Postel et al. 1996); however, other major sources of water loss through land transformation, such as deforestation, are increasingly recognized as significant contributors to the global water cycle (Gordon et al. 2004). Within urban landscapes, extensive impervious surfaces decrease infiltration of precipitation, leading to episodic high storm runoff followed by extremely low, or even intermittent, base flows (Brown et al. 2009). Global estimates of the percent of large rivers regulated by dams range from 50 %, on average, to as high as 77 % for specific regions (Nilsson et al. 2005; Lehner et al. 2011). Large dams tend to stabilize the natural extremities of hydrographs by diminishing flood flows, elevating baseflows, and reducing overall variability; thus, extensive dam regulation is likely to have homogenized regionally distinct streamflow patterns (Poff et al. 2010), potentially leading to losses in continental-wide aquatic biodiversity (Moyle and Mount 2007). Moreover, the effects of flow regulation of river systems are cumulative and certainly affect our estuaries and marine systems (Wilber 1994; Christensen et al. 1998; Livingston et al. 2000). We conclude that hydrologic alterations are widespread and will likely increase in the future regardless of the cause (direct or indirect alteration).

Altered landscape conditions in the conterminous USA. Cumulative disturbance index represents potential risk to fish habitats (NFHP 2015). The right inset depicts two major factors contributing to the index: cumulative number of upstream dams from NFHP (2015) (top inset) and urban and agricultural landscapes from MRLC (2016) (bottom inset)
Although the global state of hydrologic alteration substantiates the need for streamflow protection, the ubiquitous nature of highly modified landscapes poses fundamental challenges for environmental flow science. First, determining environmental flow measures, either as recommendations for improvement or as thresholds for protection, is primarily based on our knowledge of natural ecosystem behavior. However, according to the summary provided above, natural ecosystems are becoming rare. More importantly, rivers draining altered landscapes are likely to function and behave differently than natural systems (Brown et al. 2009). For example, peak flood events are periodic, but essential to maintaining floodplain-channel interactions and scouring sediment in natural streams; however, peak floods of similar magnitude may occur more frequently in highly urbanized watersheds and exacerbate poor habitat conditions by scouring limited quantities of wood or gravel substrates (Brown et al. 2009). Potential environmental flow recommendations for urbanized systems would be to dampen or eliminate peak flows and increase baseflows as to artificially stabilize the system. For environmental flow science to adapt, we need to understand the functionality, processes, and pathways for maintaining or restoring some desired degree of ecological integrity in highly altered landscapes.
One of the major challenges of environmental flow protection and enhancement is identifying quantifiable objectives, such as key ecological targets and flow requirements needed to sustain those targets (Richter et al. 2003). These flow requirements may represent adequate ranges of sustainability (Richter 2009) or thresholds, beyond which, ecological impairment is expected (Richter et al. 2012). Out of convenience, the natural flow regime has been used as the objective, assuming that this represents both ecological needs and flow requirements. However, identifying ecosystem needs requires intimate knowledge of the dependency of ecosystem function on flows in altered and unaltered systems, and this knowledge gap has been addressed in various settings through workshops based on regional expertise (King et al. 2008). In the absence of such information, Richter et al. (2012) proposed a presumptive standard ranging from 10 to 20 % hydrologic alteration, beyond which ecological functions may be impaired. This is highly suitable for regional planning in areas of limited a priori exploitation; however, it still relies on the natural flow regime as the baseline to measure percent alterations. Highly altered landscapes, however, may already exceed the presumptive standard, as evidenced by a large number of streams with >25 % hydrologic alteration in the USA (Carlisle et al. 2011) and the large areas of Africa, Iberia, eastern Europe, and eastern and south-central Asia ‘heavily’ or ‘severely’ affected by river regulation (Grill et al. 2015). Under these conditions, two options exist: (1) either landscape conditions are improved and/or dams are removed or (2) environmental flows must be identified that relate to the sustainability of a few fundamental ecosystem components, indicative of functioning ecosystems. Furthermore, environmental flow prescriptions may not necessarily be informed by the natural flow regime, but be catered to maximize conditions for the target ecosystem components identified, as the natural flow regime may create more harm than benefit. Due to complex stressors and balancing multiple objectives, Bayesian belief networks provide a suitable means to support decision making regarding a few ecosystem components in light of limited information (Webb et al. in press).
Another consideration is that in highly altered landscapes, channel morphology has been modified to such an extent that historical flows may not be beneficial, but potentially harmful (Jackson and Pringle 2010). A recent tendency in environmental flow science has been to recommend flows strictly on the basis of hydrologic records without proper cognizance of the dimensions or morphological condition of the stream channel. Globally, floodplains support a range of human uses (Tockner and Standford 2002); for example, in Europe, approximately 50 % of the population inhabits former floodplain areas (Tockner et al. 2009). To support this human use, channelization and levee construction have disconnected rivers from their floodplain; thus, floods of historical magnitude not only cause destruction of human infrastructure, but also massive losses of limiting habitat features (e.g., large woody debris, Roni et al. in press). In the River Elbe, Germany, reduction in floodplain storage due to urbanization has been linked to increased flood peak water levels at lower discharge in comparison to historic flood events (Becker and Grünewald 2003). River channels of the Great Plains, USA have changed from widespread, braided channels to much more narrow and incised channels that are often disconnected from the floodplains due to bed degradation, upstream dams, encroachment by salt cedar Tamarix spp., and adjacent land-use practices. The resulting morphology of the stream channels lacks the capacity to carry historical flow magnitudes (see Fig. 3). Additionally, impoundments trap sediments and bedload; thus, large discharges from dams that exceed transport capacities are likely to eliminate fine substrates unless accompanied by substrate supplementation (Kondolf 1997; Fox et al. 2016). One of the most publicized large-scale restoration projects is the Trinity River Restoration Program, California, a multifaceted restoration to rehabilitate salmon spawning habitats in a diverted stream below Trinity Dam, U.S. Bureau of Reclamation (Trush et al. 2000). A major success has been the implementation of environmental flows that mimic the natural seasonality of the pre-dam flow regime; however, the full magnitude of historical flows has not been implemented due to existing diversions (Trush et al. 2000), but also to avoid flushing valuable substrates downstream (Wilcock et al. 1996b). Furthermore, to ensure adequate salmonid spawning habitat and that the channel would match the new flow regime’s capacity, channel reconfiguration and gravel augmentation were also required (Wilcock et al. 1996a). Although regional evaluations of ecological relationships with altered hydrology can assist in developing water policies for states or basins (Poff et al. 2010), knowledge of stream channel-hydraulic interactions are needed when implementing environmental flows within specific streams. Fortunately, a multitude of frameworks are available to aid in this process (Annear et al. 2004).

Photograph comparison of the Canadian River (west of Norman, Oklahoma, USA) in the 1920’s (top panel) and the current channel (2015) near the same location (bottom panel, N 37° 45.017′ W 093° 15.984′). The photograph in the upper panel was reproduced with permission from the Research Division of the Oklahoma Historical Society
To date, the field of environmental flows has primarily focused on the water required to sustain ecosystems within individual stream reaches. Although this scope has expanded to consider larger scale dynamics and regional water policies in recent years (e.g., Poff et al. 2010), environmental flow science might benefit from greater interoperability between catchment-scale models (e.g., SWAT) and reach-specific water needs. Constraints on the availability of water to support aquatic ecosystems are likely to stem from strong human demands occurring in the watershed upstream. Environmental flow science will progressively benefit from a synergy between watershed models at basin scales in conjunction with hydraulic modeling of flow-channel dynamics. In addition, environmental flow science has understandably focused on protection and prevention through the development of policies, with most proactive restoration approaches limited to regulatory contexts supported by legislation, such as dam relicensing. Given the widespread extent of altered landscape conditions, coarse-scale research that identifies opportunities to maximize flow enhancement and restoration would be beneficial.
Interactions of Water quality and Quantity
Water quality and quantity are among the key components of environmental flows (Acreman and Ferguson 2010), and have considerable, often interactive, effects on aquatic systems. Although flow magnitude is just one component of the natural flow regime (Poff et al. 1997), the resulting effects on water quality and other processes have implications for ecological integrity (see Fig. 1, Poff et al. 1997). Existing frameworks for understanding effects of environmental flows at local (e.g., PHABSIM, Milhous and Waddle 2012) to regional (e.g., ecological limits of hydrologic alteration, Poff et al. 2010) scales (see also Tharme 2003) may not be suitable in all locations (e.g., tailwaters, McManamay and Bevelhimer 2013).
In arid and highly populated regions, to name a few, baseflow conditions may be sustained via treated sewage effluent. The wastewater discharged into these streams may support ecosystems that would otherwise not exist because of water limitations (Luthy et al. 2015). However, relying on effluent to support these ecosystems can have serious consequences. It is not unusual for polychlorinated biphenyls, polyaromatic hydrocarbons, pesticides, pharmaceuticals, personal care products, metals, and radionuclides to be present following treatment of sewage (Hughes et al. 2014). Wastewater discharges often contain constituents that have the potential to alter fish behavior (e.g., Fluoxetine, fathead minnow Pimephales promelas, Weinberger and Klaper 2014), reproductive fitness (e.g., steroidal estrogens, common roach Rutilus rutilus, Hamilton et al. 2014) and larval survival and juvenile production (e.g., steroid estrogen, fathead minnow, Schwindt et al. 2014). Wastewater reuse solutions to water shortages (i.e., reclamation of wastewater for repeated irrigation and industrial use) are increasingly used to conserve water resources (Levine and Asano 2004). While these practices augment water supplies and may reduce pollution, they pose alternative threats for lotic systems where treated wastewater may be the primary source of discharge during portions of the year. In Israel where almost all water is allocated for human uses, recycling of wastewater provides a mechanism for river recovery (Juanico and Friedler 1999; Tal 2006). However, a wastewater-derived baseflow reduces a river’s capacity to absorb further pollution loading (Juanico and Friedler 1999). In the Canadian River, Oklahoma, wastewater discharge currently supplies much of the baseflow during the summer months and is one of the last remaining strongholds for the federally threatened Arkansas River shiner Notropis girardi (Worthington et al. 2014). Major declines in baseflow lead to the loss of longitudinal connectivity that over time has important consequences for the persistence of many riverine populations (Bunn and Arthington 2002). This effect is particularly evident with repeated stream drying in fragmented stream systems of the Great Plains (Perkin et al. 2014). Decreased flows in arid streams exacerbate the influence of non-native predators on fish populations, perhaps by creating less favorable habitat conditions for flow-adapted native species, though the possible mechanisms remain unclear (Propst et al. 2008). Lastly, diminished baseflows change the dynamic between surface water and groundwater interactions (Hynes 1983; Gordon et al. 2004), especially for hyporheic flows that create thermal refugia for fish populations (Power et al. 1999; Sutton et al. 2007). Loss of these refugia created via groundwater interactions has been related to the decline of several fishes (e.g., Arkansas Darter, Etheostoma cragini, Eberle and Stark 2000).
Water quality includes chemical and thermal characteristics and is an important but often neglected component of environmental flows (Nilsson and Renöfält 2008; Olden and Naiman 2010). Decreased water quality is one of many consequences of human-altered flow regimes (Abromovitz 1996; Collier et al. 1996). Water quantity reductions concentrate solutes, leading to increased salinity (Bradley et al. 1990) among other constituents. High pulse flows are capable of flushing waste and pollutants following prolonged low-flow periods (Postel and Richter 2003). Heating capacity, and thus stream temperature, is inherently linked to the volume of water in a system (Caissie 2006). Low flows can lead to more frequent high-temperature events, which can result in reproductive stress in freshwater mussels (Galbraith and Vaughn 2009), and exceed critical temperatures for fishes (Sinokrot and Gulliver 2000) leading to bioenergetic loss (Elliott 1976) or even death (Fry 1947; Brett 1956). Despite the effects of water temperature on ecological patterns, the thermal regime has been insufficiently considered in the context of environmental flows (Olden and Naiman 2010). The thermal effects of dams contribute to the unpredictability of tailwater systems and represent a challenge for environmental flow assessments, particularly given the lack of suitable frameworks for these ecosystems.
The location of water released from a dam or diversion alters the thermal regime thereby affecting downstream biota. Failure to consider temperature effects of dam releases may lead to failure to replicate natural ecological processes (King et al. 1998). Altered thermal gradients below dams can extend for hundreds of kilometers downstream (Ellis and Jones 2013). Increased water temperatures below dams have led to accelerated migrations and earlier spawning of sockeye salmon Oncorhynchus nerka in the Pacific Northwest (Quinn et al. 1997), while hypolimnetic cool water releases resulted in reduced growth, later smoltification and lower production of Atlantic salmon and anadromous brown trout Salmo trutta in the River Surna, Norway (Saltveit 1990). Temperature gradients caused by dam releases also influence spatial patterns of gizzard shad Dorosoma cepedianum spawning in the Savannah River, between Georgia and South Carolina (Paller and Saul 1996). Alternatively, decreases in stream temperatures may extirpate native species from these locations (Haxton and Findlay 2008; Olden and Naiman 2010). For example, hypolimnetic releases have led to the development of tailwater trout fisheries below many dams in systems that were naturally considered to be ‘warmwater’ (e.g., lower Illinois River, Oklahoma, Guadalupe River below Canyon Dam, Texas). Temperature, however, is not the only physicochemical parameter altered downstream of dams and diversions.
Water releases may create additional water quality constraints in tailwater systems (i.e., the stream segment downstream of a dam). Hypolimnetic releases often result in downstream anoxia and it can be particularly difficult to engineer structures at the dam that may improve these conditions (e.g., lower Illinois River, Oklahoma). In addition, water releases are still necessary to improve downstream conditions, often a source of political and economic debate, particularly where reservoir waters are over 100 % allocated. Improvements in water quality at these locations will need to address water quantity using a framework that differs from traditional environmental flow approaches (e.g., water quality analysis simulation program modeling, Y. Zhou, unpublished data).
The interaction of reduced water quality and quantity has also severely altered the integrity of coastal ecosystems worldwide. Coastal ecosystems are unique brackish environments that form an interface between freshwater and marine environments. The ecosystem services provided to humans by coastal habitats include high productivity (Millennium Ecosystem Assessment 2008), nutrient cycling (Basset et al. 2013), reduced magnitude of flood events (Temmerman et al. 2013), and diminished intensity of storm surges (Barbier and Enchelmeyer 2014). Coastal ecosystems often also exhibit high species diversity (Basset et al. 2013), harbor endemic biota (Marrack et al. 2015), and form important nursery habitat for fish species (Bertelli and Unsworth 2014). Coastal habitats are among the most threatened ecosystems on the planet. For example, at least 35 % of mangrove forests worldwide have been destroyed (Millennium Ecosystem Assessment 2008). In addition to the large human populations in coastal areas that have affected ecosystem dynamics through excessive use of freshwater and changes in land-use practices (Montagna et al. 2002; Millennium Ecosystem Assessment 2008), inland human activities can also impair coastal habitats. Nutrients and pesticides from intensive agricultural activities that enter inland streams and rivers through runoff ultimately reach coastal areas, leading to reduced water quality with detrimental effects to biota. For example, nitrogen loading in coastal ecosystems has doubled worldwide, which has caused severe eutrophication (Millennium Ecosystem Assessment 2008) and large hypoxic zones (Ritter and Montagna 1999; Montagna and Froecshke 2009). Alternatively, coastal ecosystems also rely on inlands flows for the delivery of both freshwater and sediment. Damming and other diversions have decreased streamflow delivery rates to coastal areas by 30 % worldwide (Millennium Ecosystem Assessment 2008). Reduced freshwater inflow to coastal ecosystems alters their unique environmental characteristics that form the freshwater-marine transition. The reduction of freshwater delivery to coastal habitats can reduce ecosystem metabolism, thus lowering productivity (Russell et al. 2006). Furthermore, reduced freshwater inflow to coastal ecosystems has shifted the salinity gradient with more saline waters found further inland (Millennium Ecosystem Assessment 2008). Few brackish water organisms are tolerant of high salinity and the inland intrusion of seawater can profoundly alter the structure of coastal habitat communities (Sklar and Browder 1998). The large spatial scales that influence coastal ecosystems present unique challenges for both research and policy development (Montagna et al. 2002).
Because coastal habitats are heavily influenced by upland activities, proper ecosystem functioning requires effective environmental flow standards across large spatial scales. Increased awareness of a global hydrologic cycle has led to the realization that environmental flow plans need to be extended from local to regional scales. Some states in coastal areas have established environmental flow standards (e.g., Texas, Opdyke et al. 2014); however, the development of multi-state environment flow policies remains in its infancy and data to establish flow-ecology relationships across broad scales are severely lacking (Poff and Matthews 2013). In addition, strategies to increase freshwater inflow rates to coastal ecosystems must accommodate human needs to gain political support (Montagna et al. 2002). Researchers have proposed flexible broad-scale environmental flow plans that can be implemented in a manner that will meet ecological objectives while avoiding economic loss and land-use conflicts (e.g., Overton et al. 2014; Pang et al. 2014; Yang and Yang 2014), including plans catered to coastal habitats. For example, Sun et al. (2015) proposed an environmental flow framework that examines spatial and temporal patterns of inland streamflow on estuarine habitats while considering human land-use requirements. Unfortunately, widespread implementation and evaluation of proposed environmental flow standards remain a major hurdle (Davies et al. 2014). An additional challenge for developing effective environmental flow standards for coastal ecosystems is their unique environmental characteristics. Since they form the freshwater-marine interface and are influenced by both ocean and inland flow patterns, the ecological response to freshwater delivery is complex and non-linear in nature (Montagna et al. 2002). Simply increasing freshwater delivery rates will not restore proper ecosystem functioning of coastal habitats without consideration of other elements of the flow regime (e.g., timing and duration of flows, Koster et al. 2016).
Increasing water demands require innovative solutions to balance human needs with ecological needs, given the interplay between water quantity and quality. Targeted studies to understand potential enhancement of water quality while maintaining water quantity and meeting human demands have shown promise. For example, methods have been developed to improve water quality below dams with either minimal (Krause et al. 2005) or beneficial (Hayes et al. 1998) effects on hydropower operations. Managing for multiple goals in tailwater systems (e.g., sport fisheries, biodiversity, and habitat enhancement) is another issue of interest. How do we balance human water demands with conservation of imperiled species, particularly in water-stressed regions? Ultimately, understanding which components of water quality and quantity are manageable will be essential to determining appropriate trade-offs for meeting human and ecosystem needs. More importantly, policy development across political boundaries will be necessary to ensure coastal regions and some interstate large river systems benefit from environmental flow standards.
Invasive Species
The link between flow alteration and changes in aquatic communities is well established (Bunn and Arthington 2002; Poff and Zimmerman 2010 and references therein), although the magnitude and direction of the response are often difficult to predict (see section above on existing environmental frameworks). A modified flow regime often allows for the establishment of non-native aquatic and riparian species (Stanford et al. 1996; Bunn and Arthington 2002; Poff and Zimmerman 2010). This operates across a continuum with successful establishment of non-native species greatest in systems with the highest levels of human disturbance (Moyle and Light 1996a, b). This has resulted in several authors suggesting restoration of the natural flow regime as a mechanism for safeguarding native species while excluding introduced species (Poff et al. 1997; Marchetti and Moyle 2001; Brown and Ford 2002; Lytle and Poff 2004). However, it is generally accepted that flow regime manipulation in isolation is not a panacea for the restoration and conservation of native aquatic species (Saunders et al. 1998; Englund and Filbert 1999; Propst et al. 2008).
The multitude of threats to freshwater biodiversity rarely operates independently (Fig. 1, Dudgeon et al. 2006). For example, the spread of Tamarix species in the USA has been linked to the reduction of high-flow events following river regulation, once established Tamarix stands drive sediment aggregation and channel narrowing further altering the hydrological regime and instream habitat (see Tickner et al. 2001). Therefore, feedbacks between individual threats may result in potential unintended consequences from restoration, such as restored flow conditions benefiting non-native species (Marks et al. 2010). Although examples of flow regime restoration leading to the reestablishment of native species are apparent in the literature (e.g., Scoppettone et al. 2005; Kiernan et al. 2012), both positive and negative ecological feedbacks have been observed and predicted (Reich et al. 2010). The restoration of a more natural flow regime in New Zealand streams is predicted to allow non-native brown trout Salmo trutta to colonize areas that provide refuges for the native roundhead galaxias Galaxias anomalus potentially leading to the species’ extirpation (Leprieur et al. 2006). Likewise, increased lateral connectivity in the Willamette River, Oregon due to re-establishment of high flows is predicted to negatively affect Oregon chub Oregon ichthys crameri by increasing dispersal of non-native species into isolated floodplain populations (Scheerer 2002). In some circumstances, the management of non-native species may oppose the goals of restoring of the natural flow regime. Intentional fragmentation has been used as a mechanism to stop the expansion of several non-native species (see Table 1 in Rahel 2013). For example, within Iowa certain dams could be retained to provide a barrier to the upstream movement of Asian carp (e.g., big head carp, Hypophthalmichthys nobilis and silver carp, Hypophthalmichthys molitrix) (Hoogeveen 2010), despite restoration of long free-flowing reaches with naturally variable hydrologic regimes being highlighted for the conservation of native minnow species in the wider Missouri River basin (e.g., Dieterman and Galat 2004; Quist et al. 2004). Our challenge moving forward will be to maintain or increase hydrological connectivity while minimizing the spread of certain taxa (Rahel 2013).
Biotic responses to restoring the natural flow regime are dependent on the interaction between hydrology, geomorphology, and ecological traits of the in situ species (Shafroth et al. 2010). In the San Juan River of Colorado, New Mexico, and Utah, reservoir releases designed to mimic historic spring discharge resulted in little change in the abundance of introduced species particularly among small-bodied, short-lived, fecund taxa (Propst and Gido 2004). For management projects, the particular flow regime components modified (e.g., magnitude, timing) are critical to the ecological response. In the Colorado River, Arizona, high magnitude, short duration flood pulses released from the Glen Canyon dam in March and November limited Tamarix establishment; conversely, a smaller, prolonged event in May followed by a steady summer discharge, designed to increase humpback chub Gila cypha recruitment, produced widespread Tamarix establishment as the flow coincided with the species’ seed release (Mortenson et al. 2012). Likewise, the timing of managed flood inundation in Macquarie Marshes, Australia was implicated in stimulating recruitment of non-native species at the expense of native fauna (Rayner et al. 2015). It is pertinent that prior to undertaking flow manipulation, managers have a firm understanding of the phenology and flow requirements of both native and introduced species to determine the potential implications of restoration actions. This is highlighted by the fact that few studies have weighed the relative contributions of flow regime restoration and non-native species removal to the conservation of native biodiversity. However, Marks et al. (2010) demonstrated that flow restoration had a smaller effect on native fish species abundance in comparison to removal of non-native species, which was achieved for a greatly reduced economic cost. The authors suggest that in areas where non-native species are still established flow restoration had no effect (Marks et al. 2010). To both minimize the persistence and spread of non-native species, while promoting conservation of native taxa, a structured approach that considers the likely response of all species within the community to flow regime restoration is warranted. In many areas, such an approach may be limited by a lack of basic knowledge of species’ ecological and life-history requirements in relation to flow.
Another concern of invasive species as a component of environmental flows is evaluating their effects on hydrological alteration, especially related to their growth forms and evapotranspiration potentials. For example, giant cane Arundo donax transpires 69.3 million m3 of water per year on the Santa Ana River, California compared to an estimated 23.1 million m3 that would be consumed by native vegetation (Giessow et al. 2011). Giant cane is expansive across the lower Rio Grande Valley, Texas and along with water hyacinth Eichhornia crassipes and hydrilla Hydrilla verticillata contributed to the Rio Grande River, Texas ceasing to flow to the Gulf in 2001 (Rister et al. 2011). Further, in the Mar Chiquita coastal lagoon, Argentina, the presence of the non-native reef-building polychaete Ficopomatus enigmaticus altered both the sediment transport and water flow in the lagoon (Schwindt et al. 2004).
Non-use Economic Value
Although the capitalist economic system encourages the overexploitation of natural resources (‘tragedy of the commons’; Hardin 1968), monetary value can still be derived by conserving ‘nature.’ In fact, the economic value of ecosystem services may equal or exceed the world’s gross domestic product (Costanza et al. 2014). At the local scale, the conservation of natural resources may provide economic benefits that exceed 100 times the cost (Balmford et al. 2002) and an investment in biodiversity can promote economic growth via tourism (Freytag and Vietze 2013). For example, birdwatching (a completely non-consumptive activity) generates over $100 billion in revenue each year in the United States (USFWS 2013). Recreational fishing also provides over $100 billion each year to state and local economies through sales of licenses, equipment, bait, food, etc. (American Sportfishing Association 2011). Sustainable recreational fishing ensures economic benefit in future years, thus providing a tangible value for environmental flows. For example, fishing for Guadalupe bass, Micropterus treculii in the Edwards Plateau ecoregion, Texas, USA generates over $75 million and 776 jobs in a sparsely populated region (Thomas et al. 2014). Likewise, angling for Atlantic salmon and sea trout in England and Wales attracts visitors to rural areas, supporting jobs and contributing to the local economy (Peirson et al. 2001; Aprahamian et al. 2010). However, game fishes make up only a small percentage of freshwater fish species and recreational fishing qualifies as a use of water resources (albeit much less exploitative than commercial fishing). The lack of ‘charismatic megafauna’ among freshwater biota makes conservation efforts challenging for non-game species. Therefore, improved public support of environmental flows requires economic value to be associated with ecosystem services provided by aquatic organisms and their habitats. Meeting these challenges require an effective collaboration of ecologists, social scientists, and economists. Two interrelated areas that need to be addressed to facilitate the implementation of environment flows are (1) better educating the general public about the value of biodiversity and (2) accurate economic valuations of non-use.
Ecosystem services are benefits provided to people by nature through direct functions and/or esthetics. Freshwater fishes provide numerous benefits to the general well-being of humans through nutrient cycling, algae control, and food web stability (see Holmlund and Hammer 1999). Since aquatic vegetation is often seasonal or ephemeral, fish are often analogous to trees in freshwater systems by playing an important role in long-term nutrient storage (Allen and Hoekstra 1992). Aquatic plants and invertebrates also provide ecosystem services via improved water quality (de Bello et al. 2010). However, the long-term advantages to human prosperity through the conservation of biodiversity are largely (almost exclusively) overlooked when installing impoundments or increasing water withdrawals. Despite an inherent economic importance, ecosystem services can only be valued through knowledge. Individuals with expertise in outreach are essential to providing the general public with a basic, yet holistic, understanding of freshwater ecosystems. Better awareness of the ‘big picture’ will not only increase public support, but will also improve the effectiveness of non-use valuation methods through better-educated responses to surveys. The Environmental Protection Agency’s (EPA) Healthy Watershed Initiative is an example of a proactive approach incorporating ecosystem services, public health and stakeholder involvement for protecting watersheds and their aquatic resources (EPA 2016).
An accurate valuation of the non-use of natural resources is challenging through conventional economic assessments. Kahn (2005) outlines three methods to assess the value of non-market goods. (1) Revealed preference techniques provide a value of an environmental amenity through market value. For example, if two properties are identical with the exception of one being located near a pristine stream and the other a degraded stream, the difference in market value of the properties reveals a value of water quality. (2) Benefit transfer techniques use values from other studies and apply them to a current problem. For instance, economic losses in one area resulting from decreased fishing due to decreased water quality can be used to project losses in another area. (3) Stated preference techniques use surveys to elicit an individual’s willingness to pay for a stated environmental quality. The most common stated preference for valuing ecosystem services is contingent valuation. For example, Hejazi et al. (2013) determined that users of a freeway would pay $77 million annually (mean of $1.84 per household) to conserve natural resources along the route (primarily for esthetic value). Loomis et al. (2000) used contingent valuation to determine that households living along the Platte River in Colorado would cumulatively contribute at least $19 million annually to restoration efforts. Contingent valuation methods have been used to evaluate the non-market value of both wetlands (e.g., Brouwer et al. 1999; Grammatikopoulou and Olsen 2013) and streams (e.g., Brown and Duffield 1995; Berrens et al. 2000; Barak and Katz 2015). Contingent valuation can also be used to value water for recreation versus consumption (e.g., Loomis 2012; Loomis and McTernan 2014). The accuracy of contingent valuation is often criticized and requires skilled economists to implement effective surveys (Kahn 2005). In particular, the value of threatened and endangered species appears to have increased over time but we need more consistent measures of those values to avoid criticism (Richardson and Loomis 2009). Nevertheless, contingent valuation allows a monetary value (i.e., a number) to be associated with non-market goods such as environmental flows (Diamond and Hausman 1994; Franco and Luiselli 2013).
Human Dimensions
Traditionally, studies addressing human perceptions of the natural environment assumed that responses are distributed evenly across the landscape of interest; however, more recent evidence has suggested that environmental concerns or perceptions by people represent clustered patterns on the landscape. These spatially correlated patterns were influenced by a variety of factors including population density, demographics, socioeconomics (Brody et al. 2005), and proximity to the environmental concern in question (Brody et al. 2004). For example, there was a positive relationship between the income of farmers and their attitude toward ecosystem services (Poppenborg and Koellner 2013). Brody et al. (2004) found perceptions of water quality to relate to driving distance to the water body. These results have significant policy implications in that new water policies would be expected to be perceived differently by the community depending on the perceptions of the first stakeholder groups engaged (Brody et al. 2005). Understanding the distribution of attitudes and perceptions of water policies appears to be an important prerequisite to gaining support for advancing environmental flow science and for successfully implementing new environmental policies. Underlying attitudes and perceptions will also provide some insight to the knowledge base of different constituencies and sectors; increasing research shows natural resource/environmental literacy of the public is low (Koyle 2005).
Some of the spatial variation in people’s perception and viewpoint of the environment is related to regional dissimilarities in water law and climate. For example, in the more arid western USA, the doctrine of prior appropriation assigns priorities to water rights based on the timing of permit execution and must meet the definition of ‘beneficial uses.’ Traditionally, ‘beneficial uses’ required water to be diverted out of the stream channel which prevented environmental flows from becoming established in many areas (Clayton 2009). However, many states have expanded the meaning of ‘beneficial use’ to include the environment and have relaxed the diversion requirement (Schempp 2009). In areas where overallocation of resources has occurred, buying back water for the environment has been the most acceptable method of reallocating water, particularly in the USA and Australia (Lane-Miller et al. 2013). In the more humid eastern USA, riparian rights allow for ‘reasonable water use’ to adjacent landowners but also provide designations for instream uses in navigable waters such as swimming, boating, and fishing. However, even in these regions of greater precipitation, the attitudes associated with environmental flows and associated pressures on existing policy may diverge during times of drought (Loch et al. 2010). However, our knowledge of patch distributions of changing attitudes and perceptions is limited, making it difficult to move policies forward in some watersheds or regions.
Certain geographic factors have been well recognized in determining environmental perceptions (e.g., urban versus rural environments, Tremblay and Dunlap 1978) though temporal changes in these trends have also been acknowledged (e.g., Fortmann and Kusel 1990). Spatial distributions of changing perceptions, particularly in individual catchments, have been much less studied. However, Brody et al. (2005) studied the perceptions and drivers of community views of water quality in small catchments in south Texas and found four distinct groups of perceptions. It is unclear what spatial scale is most appropriate for examining changing perceptions of water quantity and how that scale might align with other disciplines and existing frameworks associated with environmental flows. However, because of the increase in geospatial data and spatial modeling capabilities, it is reasonable to incorporate these approaches into the social sciences and integrate these results with other environmental flow components (e.g., ecology, hydrology, and economics). Understanding the spatial changes in values, perceptions, and attitudes of environmental flows may assist in better development of the ecological questions (e.g., organism focus) and how and when stakeholders are integrated into the process of environmental flow planning. Lastly, as with non-use, the role of social scientists may be important to reduce spatial variation in attitudes and perceptions by better educating the general public about current and future environmental issues.
Conclusions
The future of environmental flow science will rely heavily on collaboration across disciplines to move forward with the aforementioned challenges. Environmental flow science will benefit from this collaboration by (1) providing a better understanding of the ecological needs of native species to offset the often unintended consequences of benefiting non-native species, (2) developing approaches to address streamflow needs in highly modified landscapes where historic flows do not provide reasonable comparisons, (3) integrating water quality needs where interactions are apparent with quantity but not necessarily the proximate factor of the ecological degradation, and (4) providing more obvious ecological study targets and locations by understanding the attitudes and perceptions of local stakeholders. Successful implementation of environmental flow policies will benefit from increasing our understanding of the spatial distribution of attitudes and perceptions across the landscape so that we can better engage the correct stakeholders in the appropriate spatial context. Further, improving our understanding of the non-use economic value will create a metric of comparison for natural resources that will (1) better appeal to city and regional developers and (2) allow for targeted implementation of objectives in regions with highly modified landscapes (e.g., developing environmental flows for persistence of remnant fish populations between reservoirs). We recognize that the interdisciplinary collaboration required to move forward will be difficult (see NRC 2004); however, scientists working in environmental flow science already conduct interdisciplinary studies (e.g., ecological limits of hydrologic alteration). The research momentum could be redirected to address additional components (i.e., altered landscapes with no potential for historic flows) and better integrate the socioeconomic disciplines. Combined, we believe these efforts would increase the overall success of environmental flow science and implementation.
References
Abromovitz JN (1996) Imperiled waters, impoverished future: the decline of freshwater ecosystems. Worldwatch Paper 128, Washington
Acreman MC, Ferguson AJD (2010) Environmental flows and the European Water Framework Directive. Freshw Biol 55:32–48
Alabaster JS (1970) River flow and upstream movement and catch of migratory salmonids. J Fish Biol 2:1–13
Alfredsen K, Harby A, Linnansaari T, Ugedal O (2012) Development of an inflow-controlled environmental flow regime for a Norwegian river. River Res Appl 28:731–739
Allan JD, Flecker AS (1993) Biodiversity conservation in running waters. Bioscience 43:32–43
Allen TFH, Hoekstra TW (1992) Towards a unified ecology. Columbia University Press, New York
American Sportfishing Association (2011) Sportfishing in America. http://asafishing.org/uploads/2011_ASASportfishing_in_America_Report_January_2013.pdf. Accessed 23 March 2015
Annear T, Chisholm I, Beecher H, Locke A, Aarrestad P, Coomer C, Estes C, Hunt J, Jacobson R, Jöbsis G, Kauffman J, Marshall J, Mayes K, Smith G, Wentworth R, Stalnaker C (2004) Instream flows for riverine resource stewardship, revised edn. Instream Flow Council, Cheyenne
Aprahamian MW, Ball M (1995) Influence of river flow on rod catch of Atlantic salmon, Salmo salar L., from the lower River Derwent, north-west England. Fish Manag Ecol 2:75–86
Aprahamian MW, Hickley P, Sheilds BA, Mawle GW (2010) Examining changes in participation in recreational fisheries in England and Wales. Fish Manag Ecol 17:93–105
Balmford A, Burner A, Cooper P, Costanza R, Farber S, Green RE, Jenkins M, Jefferiss P, Jessamy V, Madden J, Munro K, Myers N, Naeem S, Paavola J, Rayment M, Rosendo S, Roughgarden J, Trumper K, Turner RK (2002) Economic reasons for conserving wild nature. Science 297:950–953
Barak B, Katz D (2015) Valuing instream and riparian aspects of stream restoration—a willingness to tax approach. Land Use Policy 45:204–212
Barbier EB, Enchelmeyer BS (2014) Valuing the storm surge protection service of US Gulf Coast wetlands. J Environ Econ Policy 3:167–185
Basset A, Elliot M, West RJ, Wilson JG (2013) Estuarine and lagoon biodiversity and their natural goods and services. Estuar Coast Shelf S 132:1–4
Bêche LA, Mcelravy EP, Resh VH (2006) Long-term seasonal variation in the biological traits of benthic-macroinvertebrates in two Mediterranean-climate streams in California, USA. Freshw Biol 51:56–75
Becker A, Grünewald U (2003) Flood risk in Central Europe. Science 300:1099
Belmar O, Velasco J, Martinez-Capel F (2011) Hydrological classification of natural flow regimes to support environmental flow assessments in intensively regulated Mediterranean rivers, Segura River Basin (Spain). Environ Manage 47:992–1004
Berrens RP, Bohara AK, Silva CL, Brookshire D, McKee M (2000) Contingent values for New Mexico instream flows: with tests of scope, group-size reminder and temporal reliability. J Environ Manage 58:73–90
Bertelli CM, Unsworth RKF (2014) Protecting the hand that feeds us: seagrass (Zostera marina) serves as commercial juvenile fish habitat. Mar Pollut Bull 83:425–429
Birkel C, Soulsby C, Ali G, Tetzlaff D (2014) Assessing the cumulative impacts of hydropower regulation on the flow characteristics of a large Atlantic salmon river system. River Res Appl 30:456–475
Bowker J, English D, Cordell H (1999) Projections of outdoor recreation participation to 2050. Outdoor recreation in American life: a national assessment of demand and supply trends. Sagamore, Champagne, pp 323–351
Bradley PM, Kjerfve B, Morris JT (1990) Rediversion salinity change in the Cooper River, South Carolina: ecological implications. Estuaries 13:373–379
Brett JR (1956) Some principles in the thermal requirements of fishes. Q Rev Biol 31:75–87
Brisbane Declaration (2007) Environmental flows are essential for freshwater ecosystem health and human well-being. In: 10th international rivers symposium and international environmental flows conference, Brisbane, Australia
Brody SD, Highfield W, Alston L (2004) Does location matter? Measuring environmental perceptions of creeks in two San Antonio watersheds. Environ Behav 36:229–250
Brody SD, Highfield W, Peck BM (2005) Exploring the mosaic of perceptions for water quality across watersheds in San Antonio, Texas. Landscape Urban Plan 73:200–214
Brouwer R, Langford IH, Bateman IJ, Turner RK (1999) A meta-analysis of wetland contingent valuation studies. Reg Environ Change 1:47–57
Brown TC, Duffield JW (1995) Testing part-whole valuation effects in contingent valuation of instream flow protection. Water Resour Res 31:2341–2351
Brown LR, Ford T (2002) Effects of flow on the fish communities of a regulated California river: implications for managing native fishes. River Res Appl 18:331–342
Brown TC, Taylor JG, Shelby B (1991) Assessing the direct effects of streamflow on recreation: a literature review. J Am Water Resour As 27:979–989
Brown LR, Cuffney TF, Coles JF, Fitzpatricdk F, McMahon G, Steuer J, Bell AH, May JT (2009) Urban streams across the USA: lessons learned from studies in 9 metropolitan areas. J N Am Benthol Soc 28:1051–1069
Bunn SE, Arthington AH (2002) Basic principles and ecological consequences of altered flow regimes for aquatic biodiversity. Environ Manage 30:492–507
Caissie D (2006) The thermal regime of rivers: a review. Freshw Biol 51:1389–1406
Carlisle DM, Wolock DM, Meador MR (2011) Alteration of streamflow magnitudes and potential ecological consequences: a multi-regional assessment. Front Ecol Environ 9:264–270
Christensen JD, Monaco ME, Battista TA, Klein CJ, Livingston RJ, Woodsum G, Galeprin B, Huang W (1998) Potential impacts of reduced freshwater inflow on Apalachicola Bay, FL oyster (Crassostrea virginica) populations: coupling hydrologic and biologic models. NOAA/NOS Strategic Environmental Assessments Division, Silver Spring
Clayton JA (2009) Market-driven solutions to economic, environmental, and social issues related to water management in the western USA. Water 1:19–31
Collier M, Webb RH, Schmidt JC (1996) Dams and rivers: a primer on the downstream effects of dams. USGS Circular 1126, Tucson
Costanza R, de Groot R, Sutton P, van der Ploeg S, Anderson SJ, Kubiszewski I, Farber S, Turner RK (2014) Changes in the global value of ecosystem services. Global Environ Chang 26:152–158
Davies PM, Naiman RJ, Warfe DM, Pettit NE, Arthington AH, Bunn SE (2014) Flow-ecology relationships: closing the loop on effective environmental flows. Mar Freshw Res 65:133–141
Davis M, Brewer SK (2014) Gulf Coast Prairie Landscape Conservation Cooperative regional hypotheses of ecological responses to flow alteration. U.S. Department of Interior, Fish and Wildlife Service, Cooperator Science Series FWS/CSS-108-2014, Washington
De Bello F, Lavorel S, Diaz S, Harrington R, Cornelissen JHC, Bardgett RD, Berg MP, Cipriotti P, Feld CK, Hering D, da Silva PD, Potts SG, Sandin L, Sousa JP, Storkey J, Wardle DA, Harrison PA (2010) Towards an assessment of multiple ecosystem processes and services via functional traits. Biodivers Conserv 19:2873–2893
Diamond PA, Hausman JA (1994) Contingent valuation: is some number better than no number? J Econ Perspect 8:45–64
Dieterman DJ, Galat DL (2004) Large-scale factors associated with sicklefin chub distribution in the Missouri and Lower Yellowstone Rivers. Trans Am Fish Soc 133:577–587
Downs PW, Gregory KJ (2004) River channel management: towards sustainable catchment hydrosystems. Hodder Arnold, London
Doyle MW, Stanley EH, Strayer DL, Jacobson RB, Schmidt JC (2005) Effective discharge analysis of ecological processes in streams. Water Resour Res 41:W11411. doi:10.1029/2005WR004222
Duda AM, El-Ashry MT (2000) Addressing the global water and environment crises through integrated approaches to the management of land, water and ecological resources. Water Int 25:115–126
Dudgeon D, Arthington AH, Gessner MO, Kawabata Z, Knowler DJ, Leveque C, Naiman RJ, Prieur-Richard A, Soto D, Stiassny MLJ, Sullivan CA (2006) Freshwater biodiversity: importance, threats, status and conservation challenges. Biol Rev 81:163–182
Eberle ME, Stark WJ (2000) Status of the Arkansas Darter in south-central Kansas and adjacent Oklahoma. Prairie Nat 32:103–113
Elliott JM (1976) The energetics of feeding, metabolism and growth of brown trout (Salmo trutta L.) in relation to body weight, water temperature and ration size. J Anim Ecol 45:923–948
Ellis LE, Jones NE (2013) Longitudinal trends in regulated rivers: a review and synthesis within the context of the serial discontinuity concept. Environ Rev 21:136–148
Englund RA, Filbert RB (1999) Flow restoration and persistence of introduced species in Waikele Stream, O’ahu. Micronesica 32:143–154
Environmental Protection Agency (EPA) (2016) Healthy watersheds: protecting aquatic systems through landscape approaches. https://www.epa.gov/hwp. Accessed 5 April 2016
Fortmann L, Kusel J (1990) New voices, old beliefs: forest environmentalism among new and long-standing rural residents. Rural Sociol 55:214–232
Fox GA, Sheshukov A, Cruse R, Kolar RL, Guertault L, Gesch KR, Dutnell RC (2016) Reservoir sedimentation and upstream sediment sources: perspectives and future research needs on streambank and gully erosion. Environ Manage. doi:10.1007/s00267-016-0671-9
Franco D, Luiselli L (2013) A procedure to analyse the strategic outliers and the multiple motivations in a contingent valuation: a case study for a concrete policy purpose. Int J Soc Econ 40:246–266
Freytag A, Vietze C (2013) Can nature promote development? The role of sustainable tourism for economic growth. J Environ Econ Policy 2:16–44
Fry FEJ (1947) Effects of the environment on animal activity. University of Toronto Press, Toronto
Galbraith HS, Vaughn CC (2009) Temperature and food interact to influence gamete development in freshwater mussels. Hydrobiologia 636:35–47
Giessow J, Casanova J, Leclerc R, MacArthur R, Fleming G, Giessow J (2011) Arundo donax (giant reed): distribution and impact report. Agreement No. 06-374-559-0, State Water Resources Control Board, California Invasive Plant Council (Cal-IPC)
Gordon ND, McMahon TA, Finlayson BL, Gippel CJ, Nathan RJ (2004) Stream hydrology: an introduction for ecologists, 2nd edn. Wiley, West Sussex
Gore JA, Nestler JM (1988) Instream flow studies in perspective. Regul River 2:93–101
Grammatikopoulou I, Olsen SB (2013) Accounting protesting and warm glow bidding in Contingent Valuation surveys considering the management of environmental goods–an empirical case study assessing the value of protecting a Natura 2000 wetland area in Greece. J Environ Manage 130:232–241
Grill G, Lehner B, Lumsdon AE, MacDonald GK, Zarfl C, Liermann CR (2015) An index-based framework for assessing patterns and trends in river fragmentation and flow regulation by global dams at multiple scales. Environ Res Lett 10:015001
Hamilton PB, Nicol E, De-Bastos ESR, Williams RJ, Sumpter JP, Jobling S, Stevens JR, Tyler CR (2014) Populations of a cyprinid fish are self-sustaining despite widespread feminization of males. BMC Biol 12:1. doi:10.1186/1741-7007-12-1
Hardin G (1968) The tragedy of the commons. Science 162:1243–1248
Hart DD, Finelli CM (1999) Physical-biological coupling in streams: the pervasive effects of flow on benthic organisms. Ann Rev Ecol Syst 30:363–395
Haxton TJ, Findlay CS (2008) Meta-analysis of the impacts of water management on aquatic communities. Can J Fish Aquat Sci 65:437–447
Hayes DF, Labadie JW, Sanders TG, Brown JK (1998) Enhancing water quality in hydropower system operations. Water Resour Res 34:471. doi:10.1029/97WR03038
Hejazi R, Shamsudin MN, Rahim KA, Radam A, Yazdani S, Ibrahim ZZ, Vakilpoor MH, Shamshiry E (2013) Measuring the economic values of natural resources along a freeway: a contingent valuation method. J Environ Plann Man 57:629–641
Holmlund CM, Hammer M (1999) Ecosystem services generated by fish populations. Ecol Econ 29:253–268
Hoogeveen N (2010) Solving dam problems: Iowa’s 2010 plan for dam mitigation. Iowa Department of Natural Resources, Des Moines
Hughes RM, Dunham S, Maas-Hebner KG, Yeakley JA, Harte M, Molina N, Shock CC, Kacynski VW (2014) A review of urban water body challenges and approaches: (2) mitigating effects of future urbanization. Fisheries 39:30–40
Hynes HBN (1983) Groundwater and stream ecology. Hydrobiologia 100:93–99
Jackson CR, Pringle CM (2010) Ecological benefits of reduced hydrologic connectivity in intensively developed landscapes. Bioscience 60:37–46
Jackson RB, Carpenter SR, Dahm CN, McKnight DM, Naiman RJ, Postel SL, Running SW (2001) Water in a changing world. Ecol Appl 11:1027–1045
Jenkins M (2003) Prospects for biodiversity. Science 302:1175–1177
Juanico M, Friedler E (1999) Wastewater reuse for river recovery in semi-arid Israel. Water Sci Technol 40:43–50
Junk WJ, Bayley PB, Sparks RE (1989) The flood pulse concept in river-floodplain systems. Can Spec Pub Fish Aquat Sci 106:110–127
Kahn JR (2005) The economic approach to environmental natural resources, 3rd edn. South-Western, Cincinnati
Katz D (2006) Going with the flow: preserving and restoring instream water allocations. In: Gleick P (ed) The world’s water: 2006–2007. Island Press, Washington, pp 29–39
Kiernan JD, Moyle PB, Crain PK (2012) Restoring native fish assemblages to a regulated California stream using the natural flow regime concept. Ecol Appl 22:1472–1482
King J, Cambray JA, Impson ND (1998) Linked effects of dam-released floods and water temperature on spawning of the Clanwilliam yellowfish Barbus capensis. Hydrobiologia 384:245–265
King JM, Tharme RE, De Villiers MS (2008) Environmental flow assessments for rivers: manual for the building block methodology. Water Research Commission, Report No TT 354/08, Pretoria, South Africa
Kondolf GM (1997) Hungry water: effects of dams and gravel mining on river channels. Environ Manage 21:533–551
Koster WM, Amtstaetter F, Dawson DR, Reich P, Morrongiello JR (2016) Provision of environmental flows promotes spawning of a nationally threatened diadromous fish. Mar Freshw Res. doi:10.1071/MF15398
Koyle K (2005) What ten years of NEETF/Roper research and related studies say about environmental literacy in the U.S. The National Environmental Education & Training Foundation. http://www.neefusa.org/pdf/ELR2005.pdf
Krause CW, Newcomb TJ, Orth DJ (2005) Thermal habitat assessment of alternative flow scenarios in a tailwater fishery. River Res Appl 21:581–593
Lake PS (2000) Disturbance, patchiness, and diversity in streams. J N Am Benthol Soc 19:573–592
Lane-Miller CC, Wheeler S, Bjornlund H, Connor J (2013) Acquiring water for the environment: lessons from natural resource management. J Environ Policy Plan 15:513–532
Lehner B, Lierman CR, Revenga C, Vörösmarty C, Fekete B, Crouzet P, Doll P, Endejan M, Frenken K, Magome J, Nilsson C, Robertson JC, Rodel R, Sindorf N, Wisser D (2011) High-resolution mapping of the world’s reservoirs and dams for sustainable river-flow management. Front Ecol Environ 9:494–502
Leprieur F, Hickey MA, Arbuckle CJ, Closs GP, Brosse S, Townsend CR (2006) Hydrological disturbance benefits a native fish at the expense of an exotic fish. J Appl Ecol 43:930–939
Levine AD, Asano T (2004) Recovering sustainable water from wastewater. Environ Sci Technol 38:201A–208A
Livingston RJ, Lewis FG, Woodsum GC, Niu XF, Galperin B, Huang W, Christensen JD, Monaco ME, Battista TA, Klein CJ, Howell RL (2000) Modelling oyster population response to variation in freshwater input. Estuar Coast Shelf S 50:655–672
Loch A, Bjornlund H, Kuehne G (2010) Water trade alternatives in the face of climate change. Manage Environ Qual 21:226–236
Loomis JB (2012) Comparing households’ total economic values and recreation value of instream flow in an urban river. J Environ Econ Policy 1:5–17
Loomis J, McTernan J (2014) Economic value of instream flow for non-commercial whitewater boating using recreation demand and contingent valuation methods. Environ Manage 53:510–519
Loomis J, Kent P, Strange L, Fausch K, Covich A (2000) Measuring the total economic value of restoring ecosystem services in an impaired river basin: results from a contingent valuation survey. Ecol Econ 33:103–117
Loomis JB, Quattlebaum K, Brown TC, Alexander SJ (2003) Expanding institutional arrangements for acquiring water for environmental purposes: transactions evidence for the Western United States. Int J Water Resour Dev 19:21–28
Luthy RG, Sedlak DL, Plumlee MH, Austin D, Resh VH (2015) Wastewater-effluent-dominated streams as ecosystem-management tools in a drier climate. Front Ecol Environ 13:477–485
Lytle DA, Poff NL (2004) Adaptation to natural flow regimes. Trends Ecol Evol 19:94–100
Malmqvist B, Rundle S (2002) Threats to the running water ecosystems of the world. Environ Cons 29:134–153
Marchetti MP, Moyle PB (2001) Effects of flow regime on fish assemblages in a regulated California stream. Ecol Appl 11:530–539
Marks JC, Haden GA, O’Neill M, Pace C (2010) Effects of flow restoration and exotic species removal on recovery of native fish: lessons from a dam decommissioning. Restor Ecol 18:934–943
Marrack L, Beavers S, O’Grady P (2015) The relative importance of introduced fishes, habitat characteristics, and land use for endemic shrimp occurrence in brackish anchialine pool ecosystems. Hydrobiologia 758:107–122
McManamay R, Bevelhimer M (2013) A holistic framework for environmental flows determination in hydropower contexts. Project Report, Oak Ridge National Laboratory, Oak Ridge
McManamay RA, Orth DJ, Dolloff CA, Matthews DM (2013) Application of the ELOHA framework to regulated rivers in the Upper Tennessee River basin: a case study. Environ Manage 51:1210–1235
Milhous RT, Waddle TJ (2012) Physical habitat simulation (PHABSIM) software for Windows (v.1.5.1). USGS Fort Collins Science Center, Fort Collins
Millennium Ecosystem Assessment (2008) Ecosystems and human well-being: current state and trends. Island Press, Washington
Miller RR, Williams JD, Williams JE (1989) Extinctions of North American fishes during the past century. Fisheries 14:22–38
Montagna PA, Froecshke (2009) Long-term biological effects of coastal hypoxia in Corpus Christi Bay, Texas, USA. J Exp Mar Biol Ecol 381:S21–S30
Montagna PA, Alber M, Doering P (2002) Freshwater inflow: science, policy, management. Estuaries 25:1243–1245
Mortenson SG, Weisberg PJ, Stevens LE (2012) The influence of floods and precipitation on Tamarix establishment in Grand Canyon, Arizona: consequences for flow regime restoration. Biol Invasions 14:1061–1076
Moyle PB, Leidy RA (1992) Loss of biodiversity in aquatic ecosystems: evidence from fish faunas. In: Fiedler PL, Jain SK (eds) Conservation biology. Springer, New York, pp 127–169
Moyle PB, Light T (1996a) Biological invasions of fresh water: empirical rules and assembly theory. Biol Conserv 78:149–161
Moyle PB, Light T (1996b) Fish invasions in California: do abiotic factors determine success? Ecology 77:1666–1670
Moyle PB, Mount JF (2007) Homogenous rivers, homogenous fauna. Proc Natl Acad Sci USA 104:5711–5712
Multi-Resolution Land Characteristics Consortium (MRLC) (2016) National Land Cover Database 2011 (NLCD 2011). http://www.mrlc.gov/nlcd2011.php. Accessed 7 April 2016
Nadel RE (2005) Economic impacts of parks, rivers, trails and greenways. Masters thesis, University of Michigan
Naiman RJ, Bunn SE, Nilsson C, Petts GE, Pinay G, Thompson LC (2002) Legitimizing fluvial ecosystems as users of water: an overview. Environ Manage 30:455–467
National Fish Habitat Partnership (NFHP) (2010) Through a fish’s eye: the status of fish habitats in the United States 2010. Association of Fish and Wildlife Agencies, Washington. http://static.fishhabitat.org/sites/default/files/www/fishhabitatreport_Status.pdf. Accessed 23 March 2015
National Fish Habitat Partnership (NFHP) (2015) NFHP data system. http://fishhabitat.org/content/nfhp-data-system. Accessed 7 April 2016
National Research Council (NRC) (2004) Facilitating interdisciplinary research. The National Academies Press, Washington
Nilsson C, Renöfält BM (2008) Linking flow regime and water quality in rivers: a challenge to adaptive catchment management. Ecol Soc 13:1–20
Nilsson C, Reidy CA, Dynesius M, Revenga C (2005) Fragmentation and flow regulation of the world’s large river systems. Science 308:405–408
Olden JD, Naiman RJ (2010) Incorporating thermal regimes into environmental flows assessments: modifying dam operations to restore freshwater ecosystem integrity. Freshw Biol 55:86–107
Olden JD, Kennard MJ, Pusey BJ (2012) A framework for hydrologic classification with a review of methodologies and applications in ecohydrology. Ecohydrology 5:503–518
Opdyke DR, Oborny EL, Vaugh SK, Mayes KB (2014) Texas environmental flow standards and the hydrology-based environmental flow regime methodology. Hydrol Sci J 59:820–830
Overton IC, Smith DM, Dalton J, Barchiesi S, Acreman MC, Stromberg JC, Kirby JM (2014) Implementing environmental flows in integrated water resources management and the ecosystem approach. Hydrol Sci J 59:860–877
Paller MH, Saul BM (1996) Effects of temperature gradients resulting from reservoir discharge on Dorosoma cepedianum spawning in the Savannah River. Environ Biol Fishes 45:151–160
Pang A, Sun T, Yang Z (2014) A framework for determining recommended environmental flows for balancing agricultural and ecosystem water demands. Hydrol Sci J 59:890–903
Pearsall SH, McCrodden BJ, Townsend PA (2005) Adaptive management of flows in the lower Roanoke River, North Carolina, USA. Environ Manage 35:353–367
Peirson G, Tingley D, Spurgeon J, Radford A (2001) Economic evaluation of inland fisheries in England and Wales. Fish Manag Ecol 8:415–424
Perkin JS, Gido KB, Costigan KH, Daniels MD, Johnson ER (2014) Fragmentation and drying ratchet down Great Plains stream fish diversity. Aquat Conserv 25:639–655
Poff NL, Allan JD (1995) Functional organization of stream fish assemblages in relation to hydrological variability. Ecology 76:606–627
Poff NL, Matthews JH (2013) Environmental flows in the Anthropocene: past progress and future prospects. Curr Opin Environ Sustain 5:667–675
Poff NL, Zimmerman JKH (2010) Ecological responses to altered flow regimes: a literature review to inform the science and management of environmental flows. Freshw Biol 55:194–205
Poff NL, Allan JD, Bain MB, Karr JR, Prestegaard KL, Richter BD, Sparks RE, Stromberg JC (1997) The natural flow regime. Bioscience 47:769–784
Poff NL, Richter BD, Arthington AH, Bunn SE, Naiman RJ, Kendy E, Acreman M, Apse C, Bledsoe B, Freeman MC, Henriksen J, Jacobson RB, Kennen JG, Merritt DM, O’Keeffe JH, Olden JD, Rogers K, Tharme RE, Warner A (2010) The ecological limits of hydrologic alteration (ELOHA): a new framework for developing regional environmental flow standards. Freshw Biol 55:147–170
Pollock N, Chase L, Ginger C, Kolodinsky J (2012) The Northern Forest Canoe Trail: economic impacts and implications for community development. Commun Dev J 43:244–258
Poppenborg P, Koellner T (2013) Do attitudes toward ecosystem services determine agricultural land use practices? An analysis of farmers’ decision-making in a South Korean watershed. Land Use Policy 31:422–429
Postel S, Richter B (2003) Rivers for life: managing water for people and nature. Island Press, Washington
Postel SL, Daily GC, Ehrlich PR (1996) Human appropriation of renewable fresh water. Science 271:785–788
Power ME, Sun A, Parker G, Dietrich WE, Wootton JT (1995) Hydraulic food-chain models. Bioscience 45:159–167
Power G, Brown RS, Imhof JG (1999) Groundwater and fish—insights from northern North America. Hydrol Process 13:401–422
Propst DL, Gido KB (2004) Responses of native and nonnative fishes to natural flow regime mimicry in the San Juan River. Trans Am Fish Soc 133:922–931
Propst DL, Gido KB, Stefferud JA (2008) Natural flow regimes, nonnative fishes, and native fish persistence in arid-land river systems. Ecol Appl 18:1236–1252
Quinn TP, Hodgson S, Peven C (1997) Temperature, flow, and the migration of adult sockeye salmon (Oncorhynchus nerka) in the Columbia River. Can J Fish Aquat Sci 54:1349–1360
Quist MC, Hubert WA, Rahel FJ (2004) Relations among habitat characteristics, exotic species, and turbid-river cyprinids in the Missouri River drainage of Wyoming. Trans Am Fish Soc 133:727–742
Rahel FJ (2013) Intentional fragmentation as a management strategy in aquatic systems. Bioscience 63:362–372
Rayner TS, Kingsford RT, Suthers IM, Cruz DO (2015) Regulated recruitment: native and alien fish responses to widespread floodplain inundation in the Macquarie Marshes, arid Australia. Ecohydrology 8:148–159
Reich P, McMaster D, Bond N, Metzeling L, Lake PS (2010) Examining the ecological consequences of restoring flow intermittency to artificially perennial lowland streams: patterns and predictions from the Broken–Boosey creek system in northern Victoria, Australia. River Res Appl 26:529–545
Reidy Liermann CA, Olden JD, Beechie TJ, Kennard MJ, Skidmore PB, Konrad CP, Imaki H (2012) Hydrogeomorphic classification of Washington state rivers to support emerging environmental flow management strategies. River Res Appl 28:1340–1358
Revenga C, Brunner J, Henninger N, Kassem K, Payne R (2000) Pilot analysis of global ecosystems: freshwater systems. World Resources Institute, Washington
Ricciardi A, Rasmussen JB (1999) Extinction rates of North American freshwater fauna. Conserv Biol 13:1220–1222
Richardson L, Loomis J (2009) The total economic value of threatened, endangered and rare species: an updated meta-analysis. Ecol Econ 68:1535–1548
Richter BD (2009) Re-thinking environmental flows: from allocations and reserves to sustainability boundaries. River Res Appl 25:1–12
Richter BD, Braun DP, Mendelson MA, Master LL (1997) Threats to imperiled freshwater fauna. Conserv Biol 11:1081–1093
Richter BD, Mathews R, Harrison DL, Wigington R (2003) Ecologically sustainable water management: managing river flows for ecological integrity. Ecol Appl 13:206–224
Richter BD, Davis MM, Apse C, Konrad C (2012) A presumptive standard for environmental flow protection. River Res Appl 28:1312–1321
Rister ME, Sturdivant AW, Lacewell RD, Michelsen AM (2011) Challenges and opportunities for water of the Rio Grande. J Agric Appl Econ 43:367–378
Ritter C, Montagna PA (1999) Seasonal hypoxia and models of benthic response in a Texas Bay. Estuaries 22:7–20
Roni P, Beechie TJ, Pess GR, Hanson KM (2014) Wood placement in river restoration: fact, fiction and future direction. Can J Fish Aquat Sci. doi:10.1139/cjfas-2014-0344
Russell MJ, Montagna PA, Kalke RD (2006) The effect of freshwater inflow on net ecosystem metabolism in Lavaca Bay, Texas. Estuar Coast Shelf S 68:231–244
Sala OE, Chapin FS, Armesto JJ, Berlow E, Bloomfield J, Dirzo R, Huber-Sanwald E, Huenneke LF, Jackson RB, Kinzig A, Leemans R, Lodge DM, Mooney HA, Oesterheld M, Poff NL, Sykes MT, Walker BH, Walker M, Wall DH (2000) Global biodiversity scenarios for the year 2100. Science 287:1770–1774
Saltveit SJ (1990) Effect of decreased temperature on growth and smoltification of juvenile Atlantic salmon (Salmo Salar) and brown trout (Salmo trutta) in a Norwegian regulated river. Regul River 5:295–303
Saunders JF III, Tyus HM, Poff NL, Allan JD (1998) Not a panacea. BioScience 48:427–428
Scheerer PD (2002) Implications of floodplain isolation and connectivity on the conservation of an endangered minnow, Oregon Chub, in the Willamette River, Oregon. Trans Am Fish Soc 131:1070–1080
Schempp A (2009) Western water in the 21st century: policies and programs that stretch supplies in a prior appropriation world. Environmental Law Institute, Washington
Schwindt E, Iribarne OO, Isla FI (2004) Physical effects of an invading reef-building polychaete on an Argentinean estuarine environment Estuar Coast Shelf S 59:109–120
Schwindt AR, Winkelman DL, Keteles K, Murphy M, Vajda AM (2014) An environmental oestrogen disrupts fish population dynamics through direct and transgenerational effects on survival and fecundity. J Appl Ecol 51:582–591
Scoppettone GG, Rissler PH, Gourley C, Martinez C (2005) Habitat restoration as a means of controlling non-native fish in a Mojave Desert oasis. Restor Ecol 13:247–256
Shafroth PB, Wilcox AC, Lytle DA, Hickey JT, Andersen DC, Beauchamp VB, Hautzinger A, McMullen LE, Warner A (2010) Ecosystem effects of environmental flows: modelling and experimental floods in a dryland river. Freshw Biol 55:68–85
Sheffield J, Wood EF (2008) Projected changes in drought occurrence under future global warming from multi-model, multi-scenario, IPCC AR4 simulations. Clim Dynam 31:79–105
Sinokrot BA, Gulliver JS (2000) In-stream flow impact on river water temperatures. J Hydraul Res 38:339–349
Sklar FH, Browder JA (1998) Coastal environmental impacts brought about by alterations to freshwater flow in the Gulf of Mexico. Environ Manage 22:547–562
Solomon DJ, Sambrook HT (2004) Effects of hot dry summers on the loss of Atlantic salmon, Salmo salar, from estuaries in South West England. Fish Manag Ecol 11:353–363
Stalnaker CB (1993) Fish habitat evaluation models in environmental assessments. In: Hildebrand SG, Cannon JB (eds) Environmental analysis: the NEPA experience. CRC Press, Boca Raton, pp 140–162
Stalnaker CB, Wick EJ (2000) Planning for flow requirements to sustain stream biota. In: Wohl EE (ed) Inland flood hazards: human, riparian, and aquatic communities. Cambridge University Press, Cambridge, pp 411–448
Stanford JA, Ward JV (1993) An ecosystem perspective of alluvial rivers: connectivity and the hyporheic corridor. J N Am Benthol Soc 12:48–60
Stanford JA, Ward JV, Liss WJ, Frissell CA, Williams RN, Lichatowich JA, Coutant CC (1996) A general protocol for restoration of regulated rivers. Regul River 12:391–413
Strayer DL, Downing JA, Haag WR, King TL, Layzer JB, Newton TJ, Nichols SJ (2004) Changing perspectives on pearly mussels, North America’s most imperiled animals. Bioscience 54:429–439
Sun T, Zhang H, Yang Z, Yang W (2015) Environmental flow assessments for transformed estuaries. J Hydrol 520:75–84
Sutton RJ, Deas ML, Tanaka SK, Soto T, Corum RA (2007) Salmonid observations at a Klamath River thermal refuge under various hydrological and meteorological conditions. River Res Appl 23:775–785
Tal A (2006) Seeking sustainability: Israel’s evolving water management strategy. Science 313:1081–1084
Temmerman S, Meire P, Bouma TJ, Herman PM, Ysebaert T, Vriend HJ (2013) Ecosystem-based coastal defence in the face of global change. Nature 504:79–83
Tharme RE (2003) A global perspective on environmental flow assessment: emerging trends in the development and application of environmental flow methodologies for rivers. River Res Appl 19:397–441
Thomas ZA, Arsuffi TL, Magnelia SJ (2014) Fishing warmwater streams with limited public access: angling behavior, economic impact, and the role of Guadalupe bass in a twenty-four-county region of Texas. In: Tringali MD, Long JM, Birdsong, TW Allen MS (eds) Black bass diversity: multidisciplinary science for conservation. American Fisheries Society, Bethesda, pp 322–336
Thorstad EB, Heggberget TG (1998) Migration of adult Atlantic salmon (Salmo salar); the effects of artifical freshets. Hydrobiologia 371:339–346
Thorstad EB, Økland F, Aarestrup K, Heggberget TG (2008) Factors affecting the within-river spawning migration of Atlantic salmon, with emphasis on human impacts. Rev Fish Biol Fish 18:345–371
Tickner DP, Angold PG, Gurnell AM, Mountford JO (2001) Riparian plant invasions: hydrogeomorphological control and ecological impacts. Prog Phys Geog 25:22–52
Tockner K, Standford JA (2002) Riverine flood plains: present state and future trends. Environ Conserv 29:308–330
Tockner K, Uehlinger U, Robinson CT (2009) Rivers of Europe. Academic Press, London
Tremblay KR, Dunlap RE (1978) Rural residence and concern for environmental quality: a replication and extension. Rural Sociol 43:474–491
Trush WJ, McBain SM, Leopold LB (2000) Attributes of an alluvial river and their relation to water policy and management. Proc Natl Acad Sci USA 97:11858–11863
U.S. Fish and Wildlife Service (USFWS) (2013) Birding in the United States: a demographic and economic analysis. Report 2011-1, Arlington
Vannote RL, Minshall GW, Cummins KW, Sedell JR, Cushing CE (1980) The river continuum concept. Can J Fish Aquat Sci 37:130–137
Vitousek PM, Ehrlich PR, Ehrlich AH, Matson PA (1986) Human appropriation of the products of photosynthesis. Bioscience 36:368–373
Vörösmarty C, McIntyre PB, Gessner MO, Dudgeon D, Prusevich A, Green P, Glidden S, Bunn SE, Sullivan CA, Liermann CR, Davies PM (2010) Global threats to human water security and river biodiversity. Nature 467:555–561
Webb JA, de Little SC, Miller KA, Stewardson MJ, Rutherfurd ID, Sharpe AK, Patulny L, Poff NL (in press) A general approach to predicting ecological responses to environmental flows: making best use of the literature, expert knowledge, and monitoring data. River Res Appl. doi:10.1002/rra.2832
Weinberger J, Klaper R (2014) Environmental concentrations of the selective serotonin reuptake inhibitor fluoxetine impact specific behaviors involved in reproduction, feeding and predator avoidance in the fish Pimephales promelas (fathead minnow). Aquat Toxicol 151:77–83
Wilber DH (1994) The influence of Apalachicola River flows on blue crab, Callinectes sapidus, in north Florida. Fish Bull 92:180–188
Wilcock PR, Barta AF, Shea CC, Kondolf GM, Graham Matthews WV, Pitlick J (1996a) Observations of flow and sediment entrainment on a large gravel-bed river. Water Resour Res 32:2897–2909
Wilcock PR, Kondolf GM, Graham Matthews WV, Barta AF (1996b) Specification of sediment maintenance flows for a large gravel-bed river. Water Resour Res 32:2911–2921
Wootton RJ (1990) Ecology of teleost fishes. Chapman and Hall, London
Worthington TA, Brewer SK, Grabowski TB, Mueller J (2014) Backcasting the decline of a vulnerable Great Plains reproductive ecotype: identifying threats and conservation priorities. Glob Change Biol 20:89–102
Yang W, Yang ZF (2014) Evaluation of sustainable environmental flows based on the valuation of ecosystem services: a case study for the Baiyangdian Wetland, China. J Environ Inf 24:90–100
Acknowledgments
This research is a contribution of the Oklahoma Cooperative Fish and Wildlife Research Unit (U.S. Geological Survey, Oklahoma Department of Wildlife Conservation, Oklahoma State University, and Wildlife Management Institute cooperating) and several other Big XII Universities (Kansas State University, West Virginia University, University of Texas, Austin, University of Kansas, Texas Tech University, University of Oklahoma, and Baylor University). Funding was provided by the Oklahoma Water Resources Center housed in the Division of Agricultural Sciences and Natural Resources at Oklahoma State University. Any use of trade, firm, or product names is for descriptive purposes only and does not imply endorsement by the U.S. Government. We thank Kansas University for organizing the Big XII Universities Water Workshop. We thank Garey Fox and two anonymous reviewers for providing helpful comments on an earlier draft.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Brewer, S.K., McManamay, R.A., Miller, A.D. et al. Advancing Environmental Flow Science: Developing Frameworks for Altered Landscapes and Integrating Efforts Across Disciplines. Environmental Management 58, 175–192 (2016). https://doi.org/10.1007/s00267-016-0703-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00267-016-0703-5