
Abstract
Managing ecosystems to ensure the provision of multiple ecosystem services is a key challenge for applied ecology. Functional traits are receiving increasing attention as the main ecological attributes by which different organisms and biological communities influence ecosystem services through their effects on underlying ecosystem processes. Here we synthesize concepts and empirical evidence on linkages between functional traits and ecosystem services across different trophic levels. Most of the 247 studies reviewed considered plants and soil invertebrates, but quantitative trait–service associations have been documented for a range of organisms and ecosystems, illustrating the wide applicability of the trait approach. Within each trophic level, specific processes are affected by a combination of traits while particular key traits are simultaneously involved in the control of multiple processes. These multiple associations between traits and ecosystem processes can help to identify predictable trait–service clusters that depend on several trophic levels, such as clusters of traits of plants and soil organisms that underlie nutrient cycling, herbivory, and fodder and fibre production. We propose that the assessment of trait–service clusters will represent a crucial step in ecosystem service monitoring and in balancing the delivery of multiple, and sometimes conflicting, services in ecosystem management.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The recent emphasis on ecosystem services as a currency to value ecosystems and promote their sustainable use (Millennium Ecosystem Assessment 2005) has drawn attention to the ways in which different organisms contribute to the delivery of ecosystem services (Kremen 2005; Díaz et al. 2007; Suding et al. 2008; Luck et al. 2009). While the current demand for ecosystem services is growing rapidly (Beier et al. 2008; Loring et al. 2008; Carpenter et al. 2009), uncertainty remains as to how to manage the provision of multiple, and sometimes conflicting, ecosystem services. How these challenges are met will play a major role in determining the ecological, economic, and cultural future of the planet (Bennett and Balvanera 2007).
The ability of an ecosystem to provide multiple services, first, consists in allocating relevant ecosystem properties to each service and identifying which organisms, or groups of organisms, control these properties (Bengtsson 1998; Kremen 2005). Next, it is necessary to identify the key characteristics and mechanisms by which these organisms affect ecosystem properties (Grime 2001; Eviner and Chapin 2003; Hooper et al. 2005; Luck et al. 2009). This task is being undertaken in an increasing number of studies, such as those identifying the functional roles of pollinators (Kremen et al. 2007), particular plant groups as nitrogen fixers (Spehn et al. 2002), soil or sediment engineers (Wardle et al. 2004; Boyero et al. 2007) and different soil microbial and faunal groups (Bailey et al. 2002; Heemsbergen et al. 2004; Wall 2004). Furthermore, empirical studies and syntheses have identified functional traits (Box 1) as the key mechanism by which single species (Luck et al. 2009) and groups of species (Lavorel and Garnier 2002; Díaz et al. 2004, 2007; Hooper et al. 2005) influence ecosystem properties.
There is thus a growing consensus that the quantification of the kind, range, and relative abundance of functional traits in biotic communities (Box 1) can greatly contribute to ecosystem service assessment and management (Petchey and Gaston 2006; Díaz et al. 2007; Fox and Harpole 2008). However, relevant studies and concepts are scattered in the literature and across disciplines (Eviner and Chapin 2003; Heemsbergen et al. 2004; Hooper et al. 2005). In particular, as ecosystem processes relevant to important ecosystem services are often associated with the activities of organisms at several trophic levels (Bardgett and Wardle 2003; Kremen et al. 2007), trait approaches across disciplines need to be integrated.
This review takes on these challenges and is the first study to compile and synthesize the scattered information available connecting functional traits across different groups of organisms to a wide range of ecosystem processes and services. It goes beyond previous syntheses of biodiversity effects on ecosystem processes (Hooper et al. 2005; Balvanera et al. 2006; Cardinale et al. 2006; Kremen et al. 2007) by introducing the concept of trait–service clusters, which result from the multiple associations between traits and services across different trophic levels. Identifying consistent clusters of traits and services could greatly enhance our theoretical understanding and capacity for managing biodiversity for the provision of multiple ecosystem services.
Data compilation
We compiled peer reviewed literature published up to 2007 documenting links between functional traits and ecosystem services, and the underlying ecosystem processes, for the widest possible range of organisms and disciplines. We only dealt with studies that explicitly considered functional traits and/or functional groups of organisms, and excluded publications that simply reported the effects of species diversity or composition. The literature survey included: (i) a search in ISI Web of Science using various key words (“ecosystem* service*”, “ecosystem* process*”, “ecosystem* function*”, trait*, guild*, “functional diversity”, “mass ratio hypothesis”, “niche complementarity”; the same keywords were also combined with specific individual ecosystem services and processes); and (ii) a compilation of studies based on the specific literature knowledge of the different authors whose expertise collectively covers a broad range of organisms and ecosystems. This allowed inclusion in the database of those studies that actually meet the review criteria but that do not mention the above keywords explicitly.
Overall, in the final data compilation, the number of studies initially considered was then restricted based on different criteria: only those studies demonstrating statistically significant associations between traits and ecosystem processes and/or services, based on quantitative data, were considered (as opposed to verbal statements). These filters clearly restricted the number of studies considered, within the number found in the literature, to only those ones illustrating an effect of traits on a given ecosystem process. Therefore, the frequency of reported trait–service associations is a first indication of their relevance (i.e. how often a given relationship was found to be statistically significant), as well as of the ease of obtaining associated measurements or the prevalence of the hypothesis from the literature. In this approach, the magnitude of each trait–process relationship was not taken into account to avoid biased comparisons across different methodological approaches in different studies and also because this was often not explicitly indicated. Double counting of a given trait–process relationship based on the same experimental evidence was also avoided. We are aware that, most likely, our review does not include all existing studies in the literature on the link between traits and ecosystem functions. We are, however, confident that that is the first and most complete attempt to synthesize this very scattered information. Even if a limited number of studies could be missing, we are convinced that it is a excellent representation of the knowledge that it is available in the literature.
In identifying trait–service associations we followed a two-stage approach. Traits were first associated with given ecosystem processes and then single or multiple processes associated with different ecosystem services were identified (Quétier et al. 2007; Appendix). This approach was necessary to deal with the great majority of studies showing effects of traits on those ecosystem processes that control different ecosystem services, but that do not mention explicitly the relevance of traits for ecosystem services (the concept of which is more recent than much of the older literature that we considered). Although links between ecosystem processes and services are not always trivial, ecosystem services and their changes can be reasonably estimated by a set of specific processes that are routinely measured in ecological studies (Chan et al. 2006; Díaz et al. 2007). For example, Quétier et al. (2007) showed that ecosystem services identified by stakeholders, such as fodder production and the maintenance of soil fertility, can be associated with specific ecosystem processes, such as plant green biomass and litter production, soil water fluxes, and litter decomposition rate. In our approach, we also combined the ‘regulating’ and ‘supporting’ ecosystem service categories identified by the Millennium Ecosystem Assessment (2005) because, in most cases, a clear objective distinction between the two categories was not possible (see, e.g. Brauman et al. 2007). Then, ecological processes such as herbivory and seed dispersal, initially recognized in the Millennium Ecosystem Assessment as ecosystem services, were here considered as ecosystem processes affecting several services (e.g., biological control, decomposition, food production; see also Carpenter et al. 2009).
In total, 247 references documenting trait effects on ecosystem properties were compiled in a database (original database available on request; see Appendix for detailed results) in which each entry (row) was classified according to 10 criteria (columns): (1) the nature of the study (a review or primary data publication); (2) the category of ecosystem service assessed (provisioning, cultural and regulating/supporting services); (3) the specific ecosystem service assessed; (4) the ecosystem process underlying the service; (5) the level of organization at which traits were assessed (i.e. species, functional groups, whole community; see details below and Box 1); (6) the specific traits; (7) whether traits were assessed in combination with other traits or individually; (8) the relationship of traits with ecosystem processes and services (an increase in trait value/abundance improved or was detrimental to the process, or the relationship was not identifiable); (9) the organisms providing the service; and (10) the ecosystem type. The 247 studies resulted in a database with 548 entries because studies assessing multiple trait–service combinations were included as multiple rows (as, often, studies detect the effects of more than one trait on a given ecosystem process).
For the level of organization at which functional traits effects were documented (criterion 5) we distinguished different categories: (5a) species trait; (5b) functional group (Box 1); (5c) community dominant trait, i.e. locally abundant traits or functional groups resulting in different ecosystem effects (Grime 1998; Díaz et al. 2007; Garnier et al. 2007; Box 1); and (5d) community trait range, which includes studies analysing effects of functional divergence (as defined by Mason et al. 2005; Lepš et al. 2006) or functional group richness (Díaz and Cabido 2001; Hooper et al. 2005; Balvanera et al. 2006; see Box 1). These four categories were not exclusive; for instance, a case considering functional group richness would feature in the table under both functional group (5b) and trait range levels (5d).
Links between traits and services: spread of existing evidence
Relationships between traits and services have been documented for a range of organisms and ecosystems (Fig. 1). These results show the state of research and the existing bias towards specific ecosystems and organisms, therefore highlighting the areas for which the greatest research need still exists. Most of the proven trait–service associations correspond to plant communities and, to a lesser extent, to soil and freshwater organisms and ecosystems. Within organism groups, several traits have been reported, mostly structural traits in plants (e.g. plant growth form, tissue chemistry, leaf morphology, canopy size and architecture), ecophysiological traits in microbes and behavioural traits in animals (feeding habit, substrate type, nesting activity, mobility, environmental and temporal niche; Appendix). Easily measurable structural traits have been identified for plants, allowing the production of shortlists of key traits and standardized protocols to assess them (Cornelissen et al. 2003). However, such shortlists need further progress for other organisms, such as soil fauna (Faber 1991; Bradford et al. 2002; Rutgers et al. 2009), soil microbes (Heemsbergen et al. 2004), freshwater and marine ecosystems (Dolédec et al. 1999; Gerino et al. 2003) and pollinators (Kremen et al. 2007; Moretti et al. 2009). The morphological characteristics of animals most frequently associated with particular ecosystem properties were body size, relative growth rate and mandibular structure (Appendix).

Distribution of trait–service relationship information by organism and system type (number, i.e. No., and percentage of entries in the database as found in the literature are shown—note each study recorded could generate more than one entry in the database, see “Data compilation”). Entries not included in the system categorization (right figure; 15% of the total number of entries) refer to meta-analysis studies
The processes most often reported as underlying trait–service associations were rates of decomposition and mineralization, nutrient retention and sedimentation, net primary productivity (in the cases of fodder and fibre), evapotranspiration and herbivory (Appendix, Fig. 2). The assessment of other ecosystem processes and services, and especially the provisioning and cultural ones, has been based on functional trait approaches to a much lesser extent (Appendix). While evidence for trait-based relationships is accumulating for these services as well (Turpie and Joubert 2004; Quétier et al. 2007), further research on this is needed.

Distribution of trait–service relationship information by ecosystem processes assessed. Quantitative axis refers to number of entries in the database (log scale). * Cultural services not directly provided by a single ecosystem process
Components of trait effects
Functional trait effects on ecosystem processes were most frequently reported at the levels of functional groups and dominant trait values in a community, followed by those assessing traits of single species (Fig. 3). Studies showing a significant effect of the range of trait values present in a community (e.g. functional group richness and functional divergence; Box 1) on ecosystem services were less common and mostly referred to processes based on primary productivity, nutrient cycling, pollination, and in particular their maintenance through time (i.e. stability and resilience; sensu Lepš et al. 1982). The low number of studies documenting effects of the range of trait values as compared to those indicating effects of the dominant functional traits or groups suggests, in principle, more support for the mass ratio hypothesis (Grime 1998; Garnier et al. 2007) than for the niche complementarity hypothesis (Trenbath 1974; Tilman et al. 1997; Box 1). However, definitive conclusions cannot be drawn until the comparative roles of these two components are assessed over a large number of studies, including organisms other than plants. Overall, our review supports the development of frameworks incorporating different functional dimensions of biodiversity in ecosystem service assessments (Díaz et al. 2007; Mokany et al. 2008).

Levels at which the collected studies assessed the relationship between traits and ecosystem services (see “Data compilation” and Box 1 for details). No. = number of entries in the database
A clear directional relationship between functional traits and the delivery of services (either a beneficial or detrimental effect of given traits) could be established in the majority of the cases considered (69%; Appendix). The relationship was more complex in the remaining cases, for example when significant effects of traits were shown without clear specification as to their direction (e.g., when functional group composition effects were reported without explaining which particular group was mostly associated with a given process). The relevance of these directional effects of traits can have important implications for ecosystem management (see below in “Managing clusters and trade-offs of services”).
Multiple trait–service associations
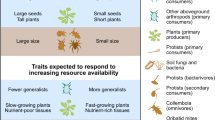
Within organism groups, combinations of key traits were important in controlling a range of ecosystem processes and services (Appendix). In particular, leaf chemistry and morphology, canopy and root architecture in plants, body size and feeding habit in soil invertebrates (Fig. 4), appeared to simultaneously influence a range of ecosystem services. While the idea of simultaneous control from a combination of traits on a given process has been previously suggested for plants (Grime et al. 1997; Lavorel and Garnier 2002; Eviner and Chapin 2003; Díaz et al. 2004, 2007) the concept could be expanded further and applied across trophic levels and ecosystem services. Indeed, only 45% of cases considered the combined effect of various traits on ecosystem service delivery and just 2% considered more than one trophic level (mostly plants together with pollinators, soil invertebrates or microbes). The understanding of multiple linkages between traits and services should thus benefit greatly from further research on the functional consequences of multitrophic trait combinations on ecosystem service delivery.

Most commonly reported plant and invertebrate traits and their involvement in multiple ecosystem service delivery. Larger arrow thickness for a given trait–service relationship indicates associations with more entries in the database (i.e., more statistically significant associations found in the literature). The assessment within this framework of multiple trait–services association with field data is thought to reduce uncertainties in the management of multifunctional ecosystems. SLA specific leaf area. * Body size can be often related to difference in feeding habit. ** The aesthetic value refers to multiple cultural services linked to land stewardship
The assessment of multiple ecosystem services: identifying trait–service clusters
Progress in the management of multiple ecosystem service provision can be attained by identifying clusters of ecologically-related services (Bennett and Balvanera 2007). The multiple associations between traits and services (Appendix, Fig. 4) offer a unique opportunity to develop such an approach, by allowing the assessment of combined biotic effects on the simultaneous delivery of multiple services. To illustrate such identification of trait–service clusters and synthesize the main concepts emerging from this review, we subjected our database to a multivariate analysis (Fig. 5). The results of this analysis are by no means applicable to the management of given ecosystems since they stem from studies conducted in different habitats and contexts. Still, they illustrate how combinations of functional traits across trophic levels control the provision of multiple ecosystem services and can be used to identify trait–service clusters. Similar approaches should be developed and validated in particular habitats and regions for identifying consistent trait–service clusters that are relevant for ecosystem management (see next section).

Association between traits and ecosystem processes including different organism types. Using Detrended Correspondence Analysis (DCA), ecosystem processes (cases) were ordinated on the basis of traits (variables). Ecosystem processes (full circles) that are similar in terms of the traits that predict them (different symbols for traits of different organisms) appear close to each other on the ordination plane (in DCA, the proximity of two points reflect contingency). Only processes with >10 entries in the review (see also Fig. 2) were considered. DCA axes 1 and 2 explain 25 and 14% of the total variability, respectively; the ordination was run on CANOCO 4.5 (Lepš and Šmilauer 2003 for details). Not all traits are shown for clarity reasons (traits displayed are those that showed the higher loads in DCA)
In our example, several trait–service clusters can be identified, highlighting the extent and current breadth of knowledge on trait–service associations. The analysis discriminated the pollination and biocontrol services provided by invertebrates and plant traits (right hand side of Fig. 5), from the water purification service mediated by a range of aquatic organisms (bottom left) and a third, much larger, cluster of functions and associated services (top left). The first two groups are intuitive for most ecologists and highlight the utility of subjecting multiple trait–service associations, such as via multivariate analyses. The third cluster revealed how various traits for plants and soil organisms show close associations with carbon and nutrient cycling (through effects on carbon sequestration and decomposition), herbivory and productivity on one hand, and water flow and soil and sediment formation on the other. Examples showing strong associations between traits from different types of organism in controlling a range of processes and services are accumulating (Berg et al. 2001; Wall 2004; Kremen et al. 2007), especially for traits of plants and soil organisms that underlie nutrient cycling, herbivory, and fodder and fibre production (Bardgett and Wardle 2003; Cornelissen et al. 2003; Díaz et al. 2004; Hattenschwiler and Gasser 2005). These examples represent a first step towards the integration of multiple traits effects across trophic levels.
Managing clusters and trade-offs of services
Our review aims at showing the potential of multiple trait–service clusters as a basis for practical ecosystem management and decision-making. In this section, we will consider the patterns emerging from the database and the specific references in the surveyed literature to illustrate three potential applications of trait–service clusters: managing trade-offs of services associated to traits (1) within one trophic level and (2) across multiple trophic levels; (3) monitoring clusters of services at different spatial scales. We highlight future research directions necessary to create, validate and launch ecosystem services monitoring frameworks in the light of these patterns.
Trait–service clusters within one trophic level
The first set of examples demonstrates, for a given trophic level, the potential for trait–service clusters to allow monitoring and management of ecosystem services trade-offs. In particular, trait–service clusters underlie possible positive or negative associations between services depending on the same functional traits. Given vegetation types, i.e. characterized by different plant traits, have beneficial effects on some services and detrimental effects on others (Chan et al. 2006; Nelson et al. 2009). For instance, the same trait configuration in plant communities that improves fodder production is likely to reduce soil carbon sequestration (Wardle et al. 2004; Dijkstra et al. 2006; Klumpp et al. 2009) and might impede services linked to aesthetic and cultural values (Hodgson et al. 2005; Quétier et al. 2007). Similar patterns emerge from other systems where key functional traits are associated with multiple ecosystem functions (Engelhardt 2006 for aquatic plants; Seeber et al. 2006 for decomposers; Wojdak and Mittelbach 2007 for pond grazers).
In this context, it is apparent from our exercise that while the services delivered by several key traits often coincided, the direction and number of statistically significant associations found for each trait–service association varied (e.g. Appendix, Fig. 4), providing a first indication of the trait’s overall relevance. While these observations remain to be tested within comparable experimental designs using standardized methodology for trait measurements (e.g. Wardle et al. 1998; Garnier et al. 2007), they do open a new area of research at the interface of land change science and fundamental ecology. At present, for example, studies explicitly quantifying the relative influence of various traits on multiple ecosystem functions are extremely rare (but see Díaz et al. 2007; Mokany et al. 2008).
We foresee that assessing the multiple directional links between traits and services will represent a key tool for quantifying ecosystem service delivery. To actively achieve this goal, new data analyses approaches should be developed. Ideally, if we consider Fig. 4 as a hypothetical cluster of traits associated with a set of services for a given ecosystem, then data analysis should assess both the direction and the extent of trait associations with specific ecosystem services. This would give arrows in the figure the ability to represent the statistical strength of a given trait–service relationship. The approach (Fig. 4) can be further expanded by considering how different components of trait effects (e.g. ‘dominant traits’ and ‘functional divergence’, see Box 1) underlie a given trait–service association. This analysis should lead to retaining the most parsimonious set of biotic and abiotic factors that are associated with clusters of related services. The first analyses in this direction show how traits can greatly support the prediction of multiple ecosystem services within the context of land cover change (Díaz et al. 2007). Similar approaches should be developed and validated in different habitats and regions for analysing consistent trait–service clusters that are relevant to ecosystem management.
Trait–service clusters across trophic levels
The second set of examples explores the potential of expanding trait–service cluster analyses across trophic levels (see examples in Appendix and Fig. 5). Figure 4 might provide also a basic analytical framework to test for the relative effects of multiple traits across trophic levels on ecosystem services. To date, the few existing studies show that the traits act mostly synergistically or as a ‘cascade’ across trophic levels. Predominantly synergistic effects occur when ecosystem services are determined by the coupled action of two or more trophic levels. For instance, processes linked to soil fertility and carbon sequestration are affected by the functional composition of both plants and soil fauna (Hattenschwiler and Gasser 2005; Zimmer et al. 2005). Also, pollination is regulated by the interaction of plant and insect traits (Fontaine et al. 2006; Albrecht et al. 2007). In multitrophic cascades, one trophic level alters the functional composition of an associated trophic level, thereby modifying the delivery of a given service. For example, changes in the proportion of functionally different riparian plant or macrophyte species can markedly alter the functional composition of leaf shredders, thus driving changes in water purification and services linked to the type and abundances of fish (Graca 2001; Meerhoff et al. 2003). Such consequences of the traits of organisms from one trophic level on other trophic levels, and the net consequences of this for ecosystem service delivery, could be demonstrated by testing for multiple organism effects. In particular, path analysis could be applied to provide estimates of the magnitude and significance of hypothesised causal connections between trophic levels and ecosystem services.
Trait–service clusters across spatial scales
Finally, the third set of examples analyses the potential of trait–service clusters to improve the quantification of ecosystem services at different spatial scales, which currently is still largely lacking (Millennium Ecosystem Assessment 2005; Chan et al. 2006; Beier et al. 2008; Nelson et al. 2009). To improve the management of multiple ecosystem services and inform land-use decisions, it is necessary to consider the spatial scale at which biotic effects are primarily acting. Our review highlights that the strength of trait–service associations might depend on the spatial scale at which the effect traits operate and that at which the services are delivered. For instance, leaf traits were more frequently reported in relation to services at smaller scales (i.e. local fertility), whilst broader-scale services, such as water and climate regulation, were more often linked to canopy traits (Fig. 4). This observation warrants more attention in order to close the gap between local and regional assessments of ecosystem service delivery (e.g., Chan et al. 2006; Beier et al. 2008; Nelson et al. 2008, 2009). It also reinforces the case for testing the relative strength of multiple trait–service associations, consistent with the framework presented in Fig. 4.
Conclusions
The impact of biodiversity changes on ecosystem service delivery can be assessed by identifying the key characteristics through which organisms affect ecosystem properties. Our study indicates that the type, the range and especially the relative abundance of functional traits in biotic communities exert a significant control over different ecosystem services across a range of organisms and ecosystems. Although the evidence so far is biased towards particular ecosystems, trophic levels and services, the approach is flexible enough to be applied more widely. Our findings also indicate that several individual traits simultaneously affect the delivery of multiple services, and that single services often depend on multiple traits, resulting in clusters of associated traits and services. These clusters can provide a common basis for managing biological communities and for the provision of bundles of services. A major remaining conceptual and methodological challenge stems from the fact that trade-offs and positive or negative feedbacks are likely within these clusters, e.g. through conflicting services that are associated with similar functional traits. Given that ecosystem processes and services rely on the combined action of different organisms, our understanding of the linkages between traits and services would also benefit from further research on the functional interactions among different trophic levels.
References
Albrecht M, Duelli P, Muller C, Kleijn D, Schmid B (2007) The Swiss agri-environment scheme enhances pollinator diversity and plant reproductive success in nearby intensively managed farmland. J Appl Ecol 44:813–822
Bailey VL, Smith JL, Bolton H (2002) Fungal-to-bacterial ratios in soils investigated for enhanced C sequestration. Soil Biol Biochem 34:997–1007
Balvanera P, Pfisterer AB, Buchmann N, He JS, Nakashizuka T, Raffaelli D, Schmid B (2006) Quantifying the evidence for biodiversity effects on ecosystem functioning and services. Ecol Lett 9:1146–1156
Bardgett RD, Wardle DA (2003) Herbivore-mediated linkages between aboveground and belowground communities. Ecology 84:2258–2268
Beier CM, Patterson TM, Chapin FS (2008) Ecosystem services and emergent vulnerability in managed ecosystems: a geospatial decision-support tool. Ecosystems 11:923–938
Bengtsson J (1998) Which species? What kind of diversity? Which ecosystem function? Some problems in studies of relations between biodiversity and ecosystem function. Appl Soil Ecol 10:191–199
Bennett EM, Balvanera P (2007) The future of production systems in a globalized world. Front Ecol Environ 5:191–198
Berg M, de Ruiter P, Didden W, Janssen M, Schouten T, Verhoef H (2001) Community food web, decomposition and nitrogen mineralisation in a stratified Scots pine forest soil. Oikos 94:130–142
Boyero L, Pearson RG, Bastian M (2007) How biological diversity influences ecosystem function: a test with a tropical stream detritivore guild. Ecol Res 22:551–558
Bradford MA, Jones TH, Bardgett RD, Black HIJ, Boag B, Bonkowski M, Cook R, Eggers T, Gange AC, Grayston SJ, Kandeler E, McCaig AE, Newington JE, Prosser JI, Setala H, Staddon PL, Tordoff GM, Tscherko D, Lawton JH (2002) Impacts of soil faunal community composition on model grassland ecosystems. Science 298:615–618
Brauman KA, Daily GC, Duarte TK, Mooney HA (2007) The nature and value of ecosystem services: an overview highlighting hydrologic services. Annu Rev Environ Resour 32:67–98
Cardinale BJ, Srivastava DS, Duffy JE, Wright JP, Downing AL, Sankaran M, Jouseau C (2006) Effects of biodiversity on the functioning of trophic groups and ecosystems. Nature 443:989–992
Carpenter SR, Mooney HA, Agard J, Capistrano D, DeFries RS, Díaz S, Dietz T, Duraiappah AK, Oteng-Yeboah A, Pereira HM, Perrings C, Reid WV, Sarukhan J, Scholes RJ, Whyte A (2009) Science for managing ecosystem services: beyond the Millennium Ecosystem Assessment. Proc Natl Acad Sci USA 106:1305–1312
Chan KMA, Shaw MR, Cameron DR, Underwood EC, Daily GC (2006) Conservation planning for ecosystem services. PLoS Biol 4:2138–2152
Cornelissen JHC, Lavorel S, Garnier E, Díaz S, Buchmann N, Gurvich DE, Reich PB, ter Steege H, Morgan HD, van der Heijden MGA, Pausas JG, Poorter H (2003) A handbook of protocols for standardised and easy measurement of plant functional traits worldwide. Aust J Bot 51:335–380
Díaz S, Cabido M (2001) Vive la difference: plant functional diversity matters to ecosystem processes. Trends Ecol Evol 16:646–655
Díaz S, Hodgson JG, Thompson K, Cabido M, Cornelissen JHC, Jalili A, Montserrat-Marti G, Grime JP, Zarrinkamar F, Asri Y, Band SR, Basconcelo S, Castro-Diez P, Funes G, Hamzehee B, Khoshnevi M, Perez-Harguindeguy N, Perez-Rontome MC, Shirvany FA, Vendramini F, Yazdani S, Abbas-Azimi R, Bogaard A, Boustani S, Charles M, Dehghan M, de Torres-Espuny L, Falczuk V, Guerrero-Campo J, Hynd A, Jones G, Kowsary E, Kazemi-Saeed F, Maestro-Martinez M, Romo-Diez A, Shaw S, Siavash B, Villar-Salvador P, Zak MR (2004) The plant traits that drive ecosystems: evidence from three continents. J Veg Sci 15:295–304
Díaz S, Lavorel S, de Bello F, Quétier F, Grigulis K, Robson M (2007) Incorporating plant functional diversity effects in ecosystem service assessments. Proc Natl Acad Sci USA 104:20684–20689
Dijkstra FA, Hobbie SE, Reich PB (2006) Soil processes affected by sixteen grassland species grown under different environmental conditions. Soil Sci Soc Am J 70:770–777
Dolédec S, Statzner B, Bournard M (1999) Species traits for future biomonitoring across ecoregions: patterns along a human-impacted river. Freshw Biol 42:737–758
Engelhardt KAM (2006) Relating effect and response traits in submersed aquatic macrophytes. Ecol Appl 16:1808–1820
Eviner VT, Chapin FS (2003) Functional matrix: a conceptual framework for predicting multiple plant effects on ecosystem processes. Annu Rev Ecol Evol Syst 34:455–485
Faber JH (1991) Functional classification of soil fauna—a new approach. Oikos 62:110–117
Fontaine C, Dajoz I, Meriguet J, Loreau M (2006) Functional diversity of plant-pollinator interaction webs enhances the persistence of plant communities. PLoS Biol 4:129–135
Fox JW, Harpole WS (2008) Revealing how species loss affects ecosystem function: the trait-based price equation partition. Ecology 89:269–279
Garnier E, Lavorel S, Ansquer P, Castro H, Cruz P, Dolezal J, Eriksson O, Fortunel C, Freitas H, Golodets C, Grigulis K, Jouany C, Kazakou E, Kigel J, Kleyer M, Lehsten V, Lepš J, Meier T, Pakeman R, Papadimitriou M, Papanastasis VP, Quested H, Quétier F, Robson M, Roumet C, Rusch G, Skarpe C, Sternberg M, Theau JP, Thebault A, Vile D, Zarovali MP (2007) Assessing the effects of land-use change on plant traits, communities and ecosystem functioning in grasslands: a standardized methodology and lessons from an application to 11 European sites. Ann Bot 99:967–985
Gerino M, Stora G, Francois-Carcaillet F, Gilbert F, Poggiale JC, Mermillod-Blondin F, Desrosiers G, Vervier P (2003) Macro-invertebrate functional groups in freshwater and marine sediments: a common mechanistic classification. Vie Milieu 53:221–231
Graca MAS (2001) The role of invertebrates on leaf litter decomposition in streams—a review. Int Rev Hydrobiol 86:383–393
Grime JP (1998) Benefits of plant diversity to ecosystems: immediate, filter and founder effects. J Ecol 86:902–910
Grime JP (2001) Plant strategies, vegetation processes and ecosystem properties. Wiley, Chichester, p 416
Grime JP, Thompson K, Hunt R, Hodgson JG, Cornelissen JHC, Rorison IH, Hendry GAF, Ashenden TW, Askew AP, Band SR, Booth RE, Bossard CC, Campbell BD, Cooper JEL, Davison AW, Gupta PL, Hall W, Hand DW, Hannah MA, Hillier SH, Hodkinson DJ, Jalili A, Liu Z, Mackey JML, Matthews N, Mowforth MA, Neal AM, Reader RJ, Reiling K, RossFraser W, Spencer RE, Sutton F, Tasker DE, Thorpe PC, Whitehouse J (1997) Integrated screening validates primary axes of specialisation in plants. Oikos 79:259–281
Harrington R, Anton C, Dawson TP, de Bello F, Feld CK, Haslett JR, Kluvánkova-Oravská T, Kontogianni A, Lavorel S, Luck GW, Rounsevell MDA, Samways MJ, Settele J, Skourtos M, Spangenberg JH, Vandewalle M, Zobel M, Harrison PA (2010) Ecosystem services and biodiversity conservation: concepts and a glossary. Biodivers Conserv. doi:10.1007/s10531-010-9834-9
Hattenschwiler S, Gasser P (2005) Soil animals alter plant litter diversity effects on decomposition. Proc Natl Acad Sci USA 102:1519–1524
Heemsbergen DA, Berg MP, Loreau M, van Haj JR, Faber JH, Verhoef HA (2004) Biodiversity effects on soil processes explained by interspecific functional dissimilarity. Science 306:1019–1020
Hodgson JG, Montserrat-Marti G, Tallowin J, Thompson K, Díaz S, Cabido M, Grime JP, Wilson PJ, Band SR, Bogard A, Cabido R, Caceres D, Castro-Diez P, Ferrer C, Maestro-Martinez M, Perez-Rontome MC, Charles M, Cornelissen JHC, Dabbert S, Perez-Harguindeguy N, Krimly T, Sijtsma FJ, Strijker D, Vendramini F, Guerrero-Campo J, Hynd A, Jones G, Romo-Diez A, Espuny LD, Villar-Salvador P, Zak MR (2005) How much will it cost to save grassland diversity? Biol Conserv 122:263–273
Hooper DU, Chapin FS, Ewel JJ, Hector A, Inchausti P, Lavorel S, Lawton JH, Lodge DM, Loreau M, Naeem S, Schmid B, Setala H, Symstad AJ, Vandermeer J, Wardle DA (2005) Effects of biodiversity on ecosystem functioning: a consensus of current knowledge. Ecol Monogr 75:3–35
Klumpp K, Fontaine S, Attard E, Le Roux X, Gleixner G, Soussana JF (2009) Grazing triggers soil carbon loss by altering plant roots and their control on soil microbial community. J Ecol 97:876–885
Kremen C (2005) Managing ecosystem services: what do we need to know about their ecology? Ecol Lett 8:468–479
Kremen C, Williams NM, Aizen MA, Gemmill-Herren B, LeBuhn G, Minckley R, Packer L, Potts SG, Roulston T, Steffan-Dewenter I, Vazquez DP, Winfree R, Adams L, Crone EE, Greenleaf SS, Keitt TH, Klein AM, Regetz J, Ricketts TH (2007) Pollination and other ecosystem services produced by mobile organisms: a conceptual framework for the effects of land-use change. Ecol Lett 10:299–314
Lavorel S, Garnier E (2002) Predicting changes in community composition and ecosystem functioning from plant traits: revisiting the Holy Grail. Funct Ecol 16:545–556
Lepš J, Šmilauer P (2003) Multivariate analysis of ecological data using Canoco. Cambridge University Press, Cambridge, 282 pp
Lepš J, Osbornovakosinova J, Rejmanek M (1982) Community stability, complexity and species life-history strategies. Vegetatio 50:53–63
Lepš J, de Bello F, Lavorel S, Berman S (2006) Quantifying and interpreting functional diversity of natural communities: practical considerations matter. Preslia 78:481–501
Loring PA, Chapin FS, Gerlach SC (2008) The services-oriented architecture: ecosystem services as a framework for diagnosing change in social ecological systems. Ecosystems 11:478–489
Luck GW, Harrington R, Harrison PA, Kremen C, Berry PM, Bugter R, Dawson TP, de Bello F, Díaz S, Feld CK, Haslett JR, Hering D, Kontogianni A, Lavorel S, Rounsevell M, Samways MJ, Sandin L, Settele J, Sykes MT, van den Hove S, Vandewalle M, Zobel M (2009) Quantifying the contribution of organisms to the provision of ecosystem services. Bioscience 59:223–235
Mason NWH, Mouillot D, Lee WG, Wilson JB (2005) Functional richness, functional evenness and functional divergence: the primary components of functional diversity. Oikos 111:112–118
Meerhoff M, Mazzeo N, Moss B, Rodriguez-Gallego L (2003) The structuring role of free-floating versus submerged plants in a subtropical shallow lake. Aquat Ecol 37:377–391
Millenium Ecosystem Assessment (2005) Ecosystems and human well-being: biodiversity synthesis. World Resources Institute, Washington, DC, 86 pp
Mokany K, Ash J, Roxburgh S (2008) Functional identity is more important than diversity in influencing ecosystem processes in a temperate native grassland. J Ecol 96:884–893
Moretti M, de Bello F, Roberts SPM, Potts SG (2009) Taxonomical vs. functional responses of bee communities to fire in two contrasting climatic regions. J Anim Ecol 78:98–108
Nelson JL, Zavaleta ES, Chapin FS (2008) Boreal fire effects on subsistence resources in Alaska and adjacent Canada. Ecosystems 11:156–171
Nelson E, Mendoza G, Regetz J, Polasky S, Tallis H, Cameron DR, Chan KMA, Daily GC, Goldstein J, Kareiva PM, Lonsdorf E, Naidoo R, Ricketts TH, Shaw MR (2009) Modeling multiple ecosystem services, biodiversity conservation, commodity production, and tradeoffs at landscape scales. Front Ecol Environ 7:4–11
Petchey OL, Gaston KJ (2006) Functional diversity: back to basics and looking forward. Ecol Lett 9:741–758
Quétier F, Lavorel S, Thuiller W, Davies I (2007) Plant-trait-based modelling assessment of ecosystem service sensitivity to land-use change. Ecol Appl 17:2377–2386
Rutgers M, Schouten AJ, Bloem J, van Eekeren N, de Goede RGM, Akkerhuis GAJM, van der Wal A, Mulder C, Brussaard L, Breure AM (2009) Biological measurements in a nationwide soil monitoring network. Eur J Soil Sci 60:820–832
Seeber J, Scheu S, Meyer E (2006) Effects of macro-decomposers on litter decomposition and soil properties in alpine pastureland: a mesocosm experiment. Appl Soil Ecol 34:168–175
Spehn EM, Scherer-Lorenzen M, Schmid B, Hector A, Caldeira MC, Dimitrakopoulos PG, Finn JA, Jumpponen A, O’Donnovan G, Pereira JS, Schulze ED, Troumbis AY, Korner C (2002) The role of legumes as a component of biodiversity in a cross-European study of grassland biomass nitrogen. Oikos 98:205–218
Suding KN, Lavorel S, Chapin FS, Cornelissen HC, Diaz S, Garnier E, Goldberg D, Hooper DU, Jackson ST, Navas ML (2008) Scaling environmental change through the community-level: a trait-based response-and-effect framework for plants. Glob Chang Biol 14:1125–1140
Tilman D, Knops J, Wedin D, Reich P, Ritchie M, Siemann E (1997) The influence of functional diversity and composition on ecosystem processes. Science 277:1300–1302
Trenbath BR (1974) Biomass productivity of mixtures. Adv Agron 26:177–210
Turpie J, Joubert A (2004) The value of flower tourism on the Bokkeveld Plateau—a botanical hotspot. Dev South Afr 21:645–662
Wall DH (ed) (2004) Sustaining biodiversity and ecosystem services in soils and sediments. Island Press, Washington, DC
Wardle DA, Barker GM, Bonner KI, Nicholson KS (1998) Can comparative approaches based on plant ecophysiological traits predict the nature of biotic interactions and individual plant species effects in ecosystems? J Ecol 86:405–420
Wardle DA, Bardgett RD, Klironomos JN, Setala H, van der Putten WH, Wall DH (2004) Ecological linkages between aboveground and belowground biota. Science 304:1629–1633
Wojdak JM, Mittelbach GG (2007) Consequences of niche overlap for ecosystem functioning: an experimental test with pond grazers. Ecology 88:2072–2083
Zimmer M, Kautz G, Topp W (2005) Do woodlice and earthworms interact synergistically in leaf litter decomposition? Funct Ecol 19:7–16
Acknowledgements
This work was supported by the RUBICODE Coordination Action (Rationalising Biodiversity Conservation in Dynamic Ecosystems) funded under the Sixth Framework Programme of the European Commission (Contract No. 036890). We are grateful for the critical comments from John Haslett, Fabien Quétier and several anonymous reviewers. Petr Šmilauer provided constructive suggestions on the multivariate example.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
de Bello, F., Lavorel, S., Díaz, S. et al. Towards an assessment of multiple ecosystem processes and services via functional traits. Biodivers Conserv 19, 2873–2893 (2010). https://doi.org/10.1007/s10531-010-9850-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10531-010-9850-9