
Abstract
Habitat-forming species on rocky shores are often subject to high levels of exploitation, but the effects of subsequent habitat loss and fragmentation on associated species and the ecosystem as a whole are poorly understood. In this study, the effects of habitat amount on the fauna associated with mussel beds were investigated, testing for the existence of threshold effects at small landscape scales. Specifically, the relationships between mussel or algal habitat amount and: associated biodiversity, associated macrofaunal abundance and density of mussel recruits were studied at three sites (Kidd’s Beach, Kayser’s Beach and Kini Bay) on the southern and south-eastern coasts of South Africa. Samples, including mussel-associated macrofauna, of 10 × 10 cm were taken from areas with 100 % mussel cover (Perna perna or a combination of P. perna and Mytilus galloprovincialis) at each site. The amount of habitat provided by mussels and algae surrounding the sampled areas was thereafter determined at the 4.0 m2 scale. A number of significant positive relationships were found between the amount of surrounding mussel habitat and the abundances of several taxa (Anthozoa, Malacostraca and Nemertea). Likewise, there were positive relationships between the amount of surrounding algal habitat and total animal abundance as well as abundance of mussel recruits at one site, Kini Bay. In contrast, abundance of mussel recruits showed a significant negative relationship with the amount of mussel habitat at Kayser’s Beach. Significant negative relationships were also detected between the amount of mussel habitat and species richness and total abundance at Kidd’s Beach, and between amount of mussel habitat and the abundance of many taxa (Bivalvia, Gastropoda, Maxillopoda, Ophiuroidea, Polychaeta and Pycnogonida) at all three sites. No threshold effects were found, nor were significant relationships consistent across the investigated sites. The results indicate that the surrounding landscape is important in shaping the structure of communities associated with these mussel beds, with significant effects of the amount of surrounding habitat per se. The strength and the direction of habitat effects vary, however, between shores and probably with the scale of observation as well as with the studied dependent variables (e.g. diversity, abundance, mussel recruitment, species identity), indicating the complexity of the processes structuring macrofaunal communities on these shores.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Ecosystems are subject to constant change and, occasionally, drastic shifts in community structure and function to an irreversible state may occur over a short period of time (e.g. Carpenter 2001; Muradian 2001; Scheffer et al. 2001). Such shifts can be characterized by threshold values of certain independent variables beyond which the dependent variables (e.g. abundance of a species or species diversity) change abruptly. An example would be changes in habitat amount. Species require specific environmental conditions in order to survive in an area, and such conditions will generally occur in relatively discrete parts of the environment, or patches. Habitat amount can then be defined as the proportion of the environment that is habitable within the mosaic of all patches that forms a landscape (Dunning et al. 1992; Fahrig 2001, 2003; Flather and Bevers 2002). Thus, habitat amount is estimated at a landscape scale, in contrast to the patch scale, e.g. at small, medium or large landscape scales. Biodiversity is expected to depend on the amount of suitable habitat in a landscape (Fahrig 2001, 2003), but the matrix or non-habitat surrounding habitable patches can also be important and can be estimated at the same scales. This is important because qualities of the matrix and the level of habitat fragmentation can affect the biodiversity and other properties of a given patch by influencing survival and fecundity as well as migration among habitat patches (Fahrig 2001; Goodsell and Connell 2008; Matias 2013). In theory, fecundity, migration and survival in the matrix influence the occurrence and nature of a possible threshold effect in the relationship between habitat amount and abundance, with the steepness of the curve affected by habitat fragmentation (Fahrig 2001). The effects of habitat fragmentation are commonly studied in terrestrial habitats (e.g. Andrén 1994; Fuhlendorf et al. 1996). Positive and negative relationships are known to occur between biodiversity and ecosystem functioning (Naeem et al. 1994; Tilman 1996; Kraufvelin et al. 2010), though most studies suggest a positive relationship between species richness and ecosystem stability (Prins et al. 1998; Gutiérrez et al. 2003; Hooper et al. 2005; Kiessling 2005; Ieno et al. 2006; Wahl et al. 2011), so that alterations to biodiversity may cause major changes to ecosystem functioning. Despite being an important topic in terrestrial systems, few marine studies have focused on the consequences of habitat loss or fragmentation (e.g. Bell et al. 2001; Caley et al. 2001), and here the effects of habitat amount at small landscape scales are considered, including the possibility of threshold effects in the relationship between habitat amount and abundance/biodiversity.
In benthic marine systems, mussels are important through their enhancement of biodiversity by providing complex, heterogeneous habitats for a diverse range of fauna (Seed 1996; Kostylev 1996; Kostylev and Erlandsson 2001; Borthagaray and Carranza 2007). By modifying the habitat in both autogenic and allogenic ways, mussels affect nutrient levels, boundary layer characteristics, amount of organic matter and many other physical characteristics of the local environment (Seed 1996; McQuaid et al. 2000; Gutiérrez et al. 2003; Sousa et al. 2009; Zaiko et al. 2009). Within intertidal mussel beds, light intensity, temperature and water movement are reduced, while sediment accumulation and relative humidity are increased compared to neighbouring rock substrata (Menge and Branch 2000; Nicastro et al. 2012). Mussel habitats also increase the benthic surface area available for colonization (Seed 1996; Gutiérrez et al. 2003; Kostylev et al. 2005). Many microhabitats, resources and niches are thus offered by mussel beds and different species may coexist within them, contributing to the further diversification of these assemblages (Kostylev 1996; Gutiérrez et al. 2003; Kostylev et al. 2005). Earlier studies have shown that bigger patches of mussels support a higher biodiversity up to a maximum size, after which an asymptote is reached, i.e. equivalent to the species–area curve (Cain 1938; Seed 1996; Pettersson 2006; Norling and Kautsky 2008; Koivisto et al. 2011). Studies of the species–area relationship for mussel-associated invertebrates generally focus on the size of the clump examined (patch scale) (Peake and Quinn 1993), but do not consider the nature of the neighbouring habitat, i.e. the patch context. Consequently, it is not known whether a greater amount of mussel habitat surrounding a given mussel patch results in greater biodiversity, species richness, abundance or specific species compositions in the patch sampled. Additionally, it is not known whether variation in habitat amount affects the nature of possible threshold effects, as predicted by Fahrig (2001).
The South African coastline is characterized by filter feeders such as mussels, which are important for species diversity (McQuaid et al. 2000) and can be used to test the effects of habitat amount on biodiversity. The indigenous brown mussel Perna perna is an ecologically and socio-economically important species on the south and east coasts that is overexploited on parts of the east coast (Siegfried et al. 1985; Harris et al. 1998; Tunley 2009). Over-exploitation has led to extremely fragmented mussel beds and even local extinction in some areas (Dye et al. 1994; Calvo-Ugarteburu and McQuaid unpubl. in Erlandsson et al. 2011a). In such cases, understanding the links between the size and fragmentation of populations of habitat-forming species and the effects of such ecological degradation on species diversity has important socio-ecological implications. Since many studies have shown correlations between biodiversity and ecosystem functioning (Hansen and Kristensen 1998; Prins et al. 1998; Gutiérrez et al. 2003; Ieno et al. 2006), it is possible that the whole ecosystem is affected if there is a relationship between habitat amount and biodiversity.
As mussel beds decrease in size, they tend to be replaced by coralline or filamentous algae (Siegfried et al. 1985; Lasiak and Field 1995, pers. obs.). New mussel larvae must therefore often settle onto algae and are then later forced to move to the primary hard substratum as they become bigger and unable to remain attached to the algae (Erlandsson and McQuaid 2004; Erlandsson et al. 2011a). Field studies and laboratory experiments indicate that the probability of recruits being able to move successfully from macroalgae to nearby mussel beds is remarkably low (Erlandsson et al. 2008, 2011a), so that the process of primary settlement onto macroalgae followed by secondary relocation into adult beds proposed by Bayne (1964) seems not to apply in this system. The negative effect of replacing adult mussels with algae from which larvae cannot successfully recolonize the primary substratum could therefore be a key driver for maintaining an ecological state in which there are few chances for natural recovery following over-exploitation. Along the South African coast, roughly half of P. perna larvae settle in mussel beds and the other half on macroalgae (McQuaid and Lindsay 2005; Erlandsson et al. 2008, 2011b). Consequently, where the density of mussels is low, it is likely that recruitment will also be low and that, as the ratio of algal to mussel cover on the shore increases, fewer individuals will reach the recruit stage (Lasiak and Barnard 1995; Erlandsson and McQuaid 2004; Erlandsson et al. 2011b). Thus, it becomes important to determine whether the amount of mussel habitat affects recruitment of new mussels into the same population and whether threshold values exist for mussel habitat amount, under which there is a drop in biodiversity.
The relationships between the amount of habitat provided by P. perna or macroalgae (mainly the red alga Gelidium pristoides) and a range of biological variables were examined on the south and south-east coasts of South Africa at small landscape scales. Five main hypotheses were posed: (1) positive or negative relationships exist between habitat amount of mussels/algae and biodiversity or abundance of associated macrofauna (total abundance or abundance of different taxonomic groups); (2) positive relationships exist between patch size and biodiversity or abundance of associated macrofauna; (3) positive relationships exist between amount of mussel/algae habitat and mussel recruitment; (4) positive relationships exist between patch size and mussel recruitment; (5) threshold effects exist (nonlinear or partial regressions) in these relationships, with e.g. abundance or biodiversity decreasing dramatically at (and below) a certain habitat amount.
Since P. perna coexists with the invasive species Mytilus galloprovincialis in the western part of the south coast in South Africa (own observations), and some of the sampling was to take place there, the importance of the ratio between these two species was also investigated.
Materials and methods
Study sites
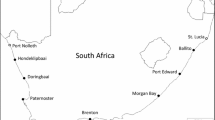
The study was carried out at three sites on the south and south-east coasts of South Africa: Kidd’s Beach, (hereafter Kidd’s 33°8,8573′S; 27°42,2104′E) and Kayser’s Beach (Kayser’s 33°12,6751′S; 27°36,7271′E), west of East London, and Kini Bay (Kini 34°1,30265′S; 25°22,7913′E), west of Port Elizabeth (Fig. 1). In contrast to shores farther east, where artisanal exploitation is intense, P. perna is abundant at these sites. All sites are exposed to strong wave action. Tides are semi-diurnal with an amplitude of ca 2 m for spring tides and ca 1 m for neap tides. Samples were collected from the mid mussel zone, where mussels form medium-sized patches interspersed with moderate to high abundance of the red alga G. pristoides and the barnacle Tetraclita serrata. Farther upshore, cover of algae and barnacles increase and mussel patches are more fragmented, while lower down, mussels create more uniform mono-layered beds, generally with 100 % cover (Dye 1998; McQuaid et al. 2000; Menge and Branch 2000).

Map showing the investigated field sites in South Africa: (from left to right) Kini, Kayser’s and Kidd’s
Sampling
Sampling was carried out during austral spring (September–October) in 2011. Two of the sites were sampled during one spring tide in September and the third (Kini) in one spring tide in October. To avoid possible natural fluctuations in biodiversity, it was necessary to complete sampling within one spring tide at each site. At each site, thirty samples were collected by scraping 10 × 10 cm quadrats placed haphazardly within patches of 100 % mussel cover. Patches were selected to provide a wide range of patch sizes in combination with a wide range of habitat amount surrounding these patches, allowing us to test our hypotheses. Mussels and all associated macrofauna were collected using a spoon and forceps and stored in 70 % ethanol until further analysis, which took place in random order. At Kidd’s and Kayser’s, adjacent samples were separated by a minimum distance of 2 m to avoid overlap in the surrounding habitat, which was estimated later. At Kini, the minimum distance between the samples was 1.5 m as fewer mussel patches were available.
The amounts of mussel and algal habitat surrounding each sample were estimated at the 4.0 m2 scale, using a 50 × 50 cm quadrat marked with crossed strings making 25 intersections (Fig. 2). The 4.0 m2 scale was chosen because it is the scale within which most mobile animals in mussel beds can move, thus enabling us to take migration between different habitat patches into consideration. A non-destructive point intercept method was used to estimate the percentage cover of the different habitats (Hawkins and Jones 1992), although at Kidd’s, the algal data were not registered separately as filamentous/foliose or encrusting algae and thus only mussel habitat amount could be investigated at this site. The cover of mussels and algae within the 4.0 m2 area was estimated with 16 non-overlapping quadrats (50 × 50 cm) with the sample in the centre of the total area (Fig. 2). Because estimates of both mussel and algal cover were necessarily made from the same surrounding area, these data can be viewed as non-independent (Underwood 1997). Nevertheless, the importance of the amount of algal habitat was considered as a possible explanation for patterns in the associated macrofauna and mussel recruitment, while recognizing that higher algal cover could possibly reflect lower mussel cover. The size of the mussel patch from which each 10 × 10 cm sample was taken was photographed except at Kini, where this was not possible, and therefore, patch sizes could not be estimated here. The photographs were then used to estimate patch sizes using the software ArcGIS (version 10.1). Sampled mussels were identified, counted and placed into size categories (0.5 mm–0.5 cm, 0.5–1 cm, 1–2 cm, 2–3 cm … 11–12 cm), and associated macrofauna individuals bigger than 0.5 mm were identified and counted. Mussel recruits were estimated directly from our samples, and individuals of 0.5–10 mm were defined as recruits (e.g. Erlandsson and McQuaid 2004). The proportions of P. perna and the non-indigenous mussel M. galloprovincialis were recorded for each sample at all sites. The associated macrofauna were identified to the lowest possible taxonomic level, which in most cases was the species level.

Mussel and algal habitat amount around the 10 × 10 cm sample (black square in the centre) was estimated at the scale 4.0 m2. This was done using a 50 × 50 cm quadrat with crossed strings making 25 intersections (upper right corner), which was placed out 16 times around the sample in the pattern indicated in the figure. By counting number of intersections covering mussels and algae, respectively, one can estimate the percentage in that square
Statistics
Both univariate and multivariate statistical techniques were used to analyse the data. Regression analyses (linear and nonlinear) were used to test the relationships between different variables, and the best-fit relationships were searched for using the models providing the highest R 2 values. To detect significant partial linear regressions, i.e. determining break points in the overall regressions suggesting possible threshold effects, a 3-step procedure was followed for each regression: (1) residual analysis, (2) regression analyses of the different slopes and (3) t tests comparing the different slopes. Step (1) Analysis of patterns among residuals (i.e. estimated differences between observed data points and the fitted regression line) was done to distinguish partial regression lines with different slopes and to determine the level of habitat amount where potential breaks between partial regressions occurred. Starting with the whole range of data points, the maximum positive or negative value (opposite sign to the first point value) of residual data was considered to correspond to a transition between partial regressions (see Kostylev and Erlandsson 2001; Erlandsson and McQuaid 2004). Step (2) Linear regression analysis was conducted for each partial regression and a statistically significant slope suggested that partial regressions should be considered. Step (3) As a last step in the detection of potential threshold effects, slopes of significant partial regressions were tested against each other using t tests in order to eliminate possible redundancy. If slopes of adjacent regressions were significantly different, then the partial regressions were considered valid. Since this procedure of partial regression analyses may include multiple tests, significance can be estimated using the Bonferroni correction.
All p values were corrected with Benjamini–Hochberg’s sequential correction (Benjamini and Hochberg 1995), with the false discovery rate at 5 %, to decrease the risk of making Type I errors. Simpson’s dominance index (1-D), total macrofaunal abundance and species richness were calculated, and multivariate statistical analyses were performed using PRIMER (version 6.1.13). Community data were analysed with nMDS analyses and one-way ANOSIM on Bray–Curtis similarities after square-root transforming the data to balance the relative influence of rare and dominant species. Differences between sites in biodiversity, abundance and species richness were tested using one-way ANOVAs. Prior to analysis, the data were tested for normality using Kolmogorov–Smirnov’s test and for homogeneity of variances using Levene’s test. Appropriate transformations were performed if the assumptions of the tests were violated.
Results
Species diversity and species composition at site level
The overall species richness exceeded 79 at Kidd’s, 81 at Kayser’s and 74 at Kini. These values are conservative, since a number of closely related species were lumped together into higher taxonomic groups (for full species/taxa list see Table 1). The taxonomic groups that were particularly well represented were Bivalvia, Gastropoda, Malacostraca, Maxillopoda and Polychaeta, which are also prominent in other mussel communities (Seed 1996). Despite the high total species richness, the abundances of associated fauna were dominated by only a few species and there were no significant differences in diversity among the three sites. NMDS analyses indicated that species composition at Kini differed from Kidd’s and Kayser’s (Fig. 3). Although, strictly, the stress value of 0.22 exceeded what is considered to be reliable by Kruskal (1964), it is not unusually high for such a large number of samples. ANOSIM further revealed significant overall differences between the sites (Global R = 0.546, p < 0.001) and pair-wise ANOSIM demonstrated that all three sites differed significantly from one another at p < 0.001. Kini differed most from Kidd’s (R = 0.726) and Kayser’s (R = 0.671), whereas Kidd’s and Kayser’s were most similar in species composition (R = 0.262).

NMDS ordination for the sampling sites Kini (upward triangles to the right), Kidd’s (squares) and Kayser’s (downward triangles to the left)
A one-way ANOVA showed that the total abundance of fauna differed significantly between sites (F 2,87 = 11.34, p < 0.001). Pair-wise a posteriori Bonferroni analyses revealed that the average total abundance was significantly higher at Kidd’s (on average 463 individuals per sample) than at Kayser’s (329 individuals, p = 0.029) and at Kini (265 individuals, p < 0.001). The total abundance per sample at Kayser’s and Kini did not differ significantly from each other. The abundance data were fifth root transformed.
Mussel versus algal habitat amount
The total amount of mussel and filamentous/foliose macroalgal cover together at 4.0 m2 ranged from 18 to 82 % at Kini and from 26 to 92 % at Kayser’s, indicating that estimation of both mussels and algae from the same quadrats could lead to some issues of negatively correlated data at these two sites. However, this was specifically checked for by running individual regression analyses between mussel habitat amount on the x-axis and algal habitat amount on the y-axis, and no significant negative relationships were found.
Relationships between mussel (and algal) habitat amount and biodiversity/species richness/abundance
Regression analyses revealed significant negative linear relationships between mussel habitat amount (independent variable) and species richness and total abundance of macrofauna (dependent variables) at Kidd’s (Table 2; Fig. 4). No significant relationship was found between mussel habitat amount and biodiversity.

Relationship between amount of mussel habitat at 4.0 m2 at Kidd’s and species richness in the upper graph (R 2 = 0.154, y = −0.250x + 40.203, F = 5.087, p = 0.032), and total macrofauna abundance in the lower graph (R 2 = 0.165, y = −0.012x + 3.72, F = 5.578, p = 0.025)
One significant positive relationship was found between algal habitat amount and total macrofaunal abundance at Kini. No other significant relationships were found between algal habitat amount and total macrofaunal abundance, neither were there any significant relationships between algal habitat amount and biodiversity or species richness at any of the sites (Table 3).
Relationship between mussel (and algal) habitat amount and P. perna recruitment and size
The number of P. perna recruits in the samples ranged from 0 to 13 per 10 × 10 cm quadrat at Kidd’s, from 5 to 39 at Kayser’s and from 4 to 23 at Kini. No significant positive relationship was found between mussel recruit density and habitat amount at any site, though there was a significant negative relationship at Kayser’s (Table 4; Fig. 5). For relationships between habitat amount and mean mussel size, there was also one significant relationship, a positive power function at Kini (Table 4). When testing recruit density against algal habitat amount, there was one significant relationship, a quadratic one at Kini (Table 3; Fig. 6). This relationship was based on a distinct outlier for which algal habitat was particularly high and recruit density particularly low. If the outlier is ignored, the relationship becomes a positive power function (Fig. 6). In all cases, the R 2 values were very low.

Relationship between amount of mussel habitat at 4.0 m2 and number of P. perna recruits at Kayser’s (R 2 = 0.181, y = −0.02x + 3.22, F = 6.193, p = 0.019)

Relationship between algal habitat amount at the 4.0 m2 scale and the density of mussel recruits at Kini (R 2 = 0.38, y = 1.091 + 0.0938x − 0.00193x 2, F = 8.29, p = 0.002) showed by the dashed line. If the outlier to the right (open circle) is ignored, there is a significantly positive power relationship between the variables (R 2 = 0.249, y = 1,170x 0.195, F = 8.932, p = 0.006) which is shown by the fully drawn line
Relationship between mussel habitat amount and different taxonomic groups
The analysis of the relationships between abundances of separate phyla/classes/species and mussel habitat amount resulted in several significant results (Table 5). Most taxa were investigated for such relationships, leaving out only groups where the total number of individuals per site was less than 50. At Kini 11.6 %, at Kayser’s 9.4 % and at Kidd’s, only 3.7 % of all individuals fell into the category that was not investigated. Positive relationships, either linear or nonlinear, were found for the classes Anthozoa and Malacostraca and the phylum Nemertea and individual species belonging to these taxonomic groups. Negative relationships, also either linear or nonlinear, were found between mussel habitat amount and the classes Bivalvia, Maxillopoda, Ophiuroidea, Polychaeta and Pycnogonida and species within these classes. Taken over all, the class Gastropoda showed a negative relationship with mussel habitat amount, but Fissurella mutabilis showed a positive relationship. A quadratic relationship was found for the cushion star Patiriella exigua and Maxillopoda against mussel habitat amount at Kayser’s.
Relationship between mussel patch size and biodiversity/abundance/recruitment
Actual patch size (the size of the patch from which the sampled quadrat was taken) varied between 0.014 and 1.8 m2 at Kidd’s and between 0.014 and 2.0 m2 at Kayser’s (patch size could not be measured at Kini). No significant relationships were found between patch size and the dependent variables biodiversity, species richness, total abundance or recruit density (p > 0.05 in all cases).
Role of the ratio between P. perna and M. galloprovincialis
At Kini, where M. galloprovincialis was abundant, the influence of the proportion of P. perna on species richness and total abundance of fauna was also examined and significantly positive linear relationships were found. The more P. perna there were in a sample, the higher the species richness (R 2 = 0.201, F = 7.027, p = 0.013) and the higher the total abundance of associated animals (R 2 = 0.161, F = 5.383, p = 0.028), though in both cases R 2 values were low, indicating that there were other important sources of variation.
Discussion
Earlier studies on the relationships between mussels and biodiversity were done mainly at the patch scale, without taking the patch context, e.g. the matrix and surrounding habitat, into account (e.g. Tsuchiya and Nishihira 1985; Dittmann 1990; Peake and Quinn 1993; Hammond and Griffiths 2006; Koivisto et al. 2011; see however Koivisto and Westerbom 2012) and often indicated positive relationships between the two (e.g. Norling and Kautsky 2008; Zaiko et al. 2009). The present study showed several significant relationships, either negative or positive, between habitat amount and various other variables. Critically, however, no thresholds could be detected using residual analysis to suggest break points between partial regressions (Kostylev and Erlandsson 2001) in any of the relationships that were tested, even though some of the significant quadratic relationships suggested nonlinear trends. Thus, at the small scale at which the effects of habitat amount were studied, there was no evidence for abundance or biodiversity dramatically decreasing below a certain level of habitat amount. However, this does not exclude the possibility that this kind of pattern exists at larger or smaller landscape scales.
Habitat loss has been shown to have negative effects on processes that are important for population dynamics and persistence such as population growth, foraging, reproduction, dispersal and predation (Fahrig 2003 and references therein, Kraufvelin et al. 2006a, b; Bulleri et al. 2012). Small and more isolated patches, which are likely to result from habitat loss and fragmentation, may not be as effective as larger patches at sustaining local populations of species, partly because small and often slow moving animals may not be able to migrate through the matrix, which increases in size as habitat is lost (Fahrig 2001). Furthermore, small patches result in increased amounts of edge habitat, and this can cause higher migration rates to the matrix (Fahrig 2003) and simulations indicate that more habitat is required for regional population survival, if the emigration rate becomes higher (Fahrig 2001). Therefore, less matrix and more surrounding habitat should allow more animals to migrate to other mussel patches (or parts of the patches) and thus contribute to a generally higher biodiversity. On the other hand, if habitat fragmentation does not include habitat loss, it may have more positive than negative effects due to habitat complementarity and reduced isolation of patches (Dunning et al. 1992; Fahrig 2003). Thus, some studies indicate that habitat fragmentation affects abundances in a community only if the overall habitat amount is low (Fahrig 1998; Trzcinski et al. 1999). Although not explicitly investigated here, it is thus possible that patterns of fragmentation could explain the absence of any significant relationship between mussel habitat amount and biodiversity and total abundance at Kayser’s and Kini (Table 2).
The observed large differences (in nMDS and in ANOSIM) in community structure between Kini and the two other sites could be due to the relatively large geographical distance separating Kini (e.g. Branch et al. 1999) or by the higher proportion of M. galloprovincialis at Kini than at Kidd’s and Kayser’s, where P. perna was clearly dominant. Iwasaki (1995) showed that different species can dominate the associated faunal community depending on which mussel species dominates the mussel population. On the other hand, no strong differences in the biodiversity of communities associated with P. perna as opposed to M. galloprovincialis have previously been detected (Hammond 2001; Jordaan 2010). Consequently, there was no direct evidence of a strong mussel species effect on associated communities in our system, and in fact earlier work indicates that, instead, the infaunal community is strongly affected by the size structure of mussel populations (O’Connor and Crowe 2007; Cole and McQuaid 2010). This suggests a mechanism for species-specific effects as the two mussel species often differed in size (P. perna being bigger in our samples than M. galloprovincialis), though manipulative experiments using P. perna provided no support for the effects of mussel bed structure (Cole and McQuaid 2011).
Taxonomic groups differed in their relationships with habitat amount, but this was not unexpected (Fahrig 2013), and there may be several explanations for such different patterns. One explanation relates to differences in the life cycles of the various species. For example, larvae settling among mussels may be buried in the large amounts of faeces and pseudofaeces that mussels produce. Species with larval stages that are more tolerant to anoxic conditions thus seem to have higher chances for survival among mussel beds (Commito and Boncavage 1989). Species with a pelagic larval stage are also prone to mussel predation when they settle in the intertidal, since they may be filtered by the mussels themselves (Mileikovsky 1974 and references therein, Porri et al. 2008). This could result in different relationships with mussel cover for species with planktonic larvae as opposed to those that brood their offspring or encapsulate the embryo (Dean 1978; Commito and Boncavage 1989). Species with direct development or species where the larvae are well developed or have already become small juveniles before they enter the adult population also have a higher probability of survival in mussel habitats than other species (Thiel and Ullrich 2002). The link between foraging by mussels and the reproductive strategies of associated fauna could thus help explain the negative nature of some relationships between mussel habitat amount and abundances of different species and taxonomic groups. For example, bivalves and many polychaetes (with pelagic larval stages) showed negative relationships to mussel habitat amount. Amphipods and isopods, on the other hand, are direct developers (Väinölä et al. 2008; Wilson 2008) and most showed positive correlations with mussel habitat amount. In addition to being very abundant at all three sites, the class Malacostraca and a few species within it (e.g. the isopods Jaeropsis spp.) had significant positive relationships with mussel habitat amount. Not having a long larval stage in their life cycle could thus have influenced their abundance among the mussels. Amphipods, and other mobile groups of species, are also more likely to benefit from a larger habitat amount due to their mobility.
The relationship between adult and recruit densities in mussel beds varies with the scale at which it is examined (Reaugh-Flower et al. 2011). This often indicates a strong positive relationship between recruit density and abundance or percent cover of adult mussels at the patch scale (Erlandsson and McQuaid 2004; McQuaid and Lindsay 2007), but in the present study no positive relationships were found between recruitment and the size of surrounding mussel habitat considered. Furthermore, a significantly positive relationship between mussel habitat amount and mean size of the mussels was found at Kini (Table 4). This may be because older, larger mussels create larger mussel beds (Norling and Kautsky 2007, own observations) that have had longer to recruit infauna and undergo succession, but again, it is uncertain that mussel size itself has a direct effect. To our knowledge, no other studies have focused on the relationship between recruit density and the amount of habitat surrounding a patch. The negative relationship between mussel habitat amount and recruit density at Kayser’s (Table 4; Fig. 5) had extremely low predictive power, but was counter to our hypothesis 3 and we have no explanation for it.
Algal habitat amount seemed to have a positive effect on total abundance of associated macrofauna, but only at Kini. Koivisto and Westerbom (2010) found a positive relationship between the presence of algal structures and biodiversity in Mytilus edulis communities and ascribed this pattern to the larger surface area and additional complexity offered by the algae. Some species can also find a hiding place among the algae, while grazers might find more food (e.g. periphytic microalgae) on and among the macroalgae. Algal habitat amount also had a positive effect on mussel recruit density, which could reflect small-scale movement of recruits to nearby mussel habitats. Earlier studies, however, have shown that this is unlikely (Erlandsson et al. 2008, 2011b). Such results should therefore be interpreted with caution, but cannot be ruled out as possible local factors. Additional caution should be applied with regard to the possible non-independence between amounts of mussel and algal habitat (Underwood 1997). These were estimated from the same quadrats, making it impossible to disentangle the relative importance of higher amounts of algae from those of possibly lower amounts of mussels. Nevertheless, this issue was considered as a minor one since there was no significant negative correlation between the two habitat amounts.
The introduction of non-indigenous species has caused changes to many ecosystems (Hooper et al. 2005; Harley et al. 2013; Kraufvelin 2013, 2014; Garbary et al. 2014). In South Africa, M. galloprovincialis was introduced to the west coast more than 30 years ago and has spread along the coast since then (Grant and Cherry 1985; McQuaid and Phillips 2000) and outcompeted the ribbed mussel Aulacomya ater on the west coast (Van Erkom and Griffiths 1990). P. perna and M. galloprovincialis can, however, co-exist on the south coast, partly because of a delicate balance between different competitive and facilitative interactions (Rius and McQuaid 2006, 2009) that results in each dominating a different zone, with overlap and co-existence in the mid mussel zone (Bownes and McQuaid 2006; Erlandsson et al. 2011b). According to earlier studies, relatively few species seem to have been directly negatively affected by this introduction (Hanekom 2008), though these negative effects can be very powerful (e.g. Steffani and Branch 2003, 2005) and it is not clear how the rocky shore ecosystem may be affected by the further spread eastwards of M. galloprovincialis. In the present study, positive relationships were found between the relative amount of P. perna and total abundance of associated animals, although this may have been caused by the response of just a few species. Nevertheless, it could indicate that the possible negative influence of M. galloprovincialis on the biodiversity of mussel-associated macrofauna may be greater than previously reported.
In conclusion, this study showed that there are significant relationships between the amount of mussel habitat surrounding a patch at small landscape scales and numerous response variables in mussel-associated species assemblages. Even though only a few significant relationships were found between mussel habitat amount and species richness and total macrofauna abundance, the mussel habitat amount still seemed to have an effect on the abundance of many specific taxonomic groups and the results suggest that this may be linked to mode of reproduction. No threshold effects in the relationship between habitat amount and abundance/biodiversity were observed at the scale investigated. Our study highlights the important, but complex landscape effects that reflect relationships between habitat amount, species composition and species abundances in the marine environment. The results also show that habitat amount per se is a factor structuring the fauna associated with South African mussel beds, but that the strength of its effects varies between shores and probably with scale, as well as with the studied dependent variable, including species identity.
References
Andrén H (1994) Effects of habitat fragmentation on birds and mammals in landscapes with different proportions of suitable habitat: a review. Oikos 71:355–366
Bayne BL (1964) Primary and secondary settlement in Mytilus edulis (Mollusca). J Anim Ecol 33:513–523
Bell SS, Brooks RA, Robbins BD, Fonseca MS, Hall MO (2001) Faunal response to fragmentation in seagrass habitats: implications for seagrass conservation. Biol Conserv 100:115–123
Benjamini Y, Hochberg Y (1995) Controlling the false discovery rate: a practical and powerful approach to multiple testing. J R Stat Soc 57:289–300
Borthagaray AI, Carranza A (2007) Mussels as ecosystem engineers: their contribution to species richness in a rocky littoral community. Acta Oecol 31:243–250
Bownes SJ, McQuaid CD (2006) Will the invasive mussel Mytilus galloprovincialis Lamarck replace the indigenous Perna perna L. on the south coast of South Africa? J Exp Mar Biol Ecol 338:140–151
Branch GM, Beckley LE, Branch ML, Griffiths CL (1999) Two Oceans: a guide to the marine life of southern Africa, 2nd edn. David Philip, Cape Town
Bulleri F, Benedetti-Cecchi L, Cusson M, Maggi E, Arenas F, Aspden R, Bertocci I, Crowe TP, Davoult D, Eriksson BK, Fraschetti S, Golléty C, Griffin JN, Jenkins SR, Kotta J, Kraufvelin P, Molis M, Sousa Pinto I, Terlizzi A, Valdivia N, Paterson DM (2012) Temporal stability of European rocky shore assemblages: variation across a latitudinal gradient and the role of habitat-formers. Oikos 121:1801–1809
Cain SA (1938) The species–area curve. Am Midl Nat 19:573–581
Caley MJ, Buckley KA, Jones GP (2001) Separating ecological effects of habitat fragmentation, degradation, and loss on coral commensals. Ecology 82:3435–3448
Carpenter SR (2001) Alternate states of ecosystems: evidence and its implications. In: Huntly N, Lewis S (eds) Ecology: achievement and challenge. Blackwell, London, pp 358–383
Cole VJ, McQuaid CD (2010) Bioengineers and their associated fauna respond differently to the effects of biogeography and upwelling. Ecology 91:3549–3562
Cole VJ, McQuaid CD (2011) Broad-scale spatial factors outweigh the influence of habitat structure on the fauna associated with a bioengineer. Mar Ecol Prog Ser 442:101–109
Commito JA, Boncavage EM (1989) Suspension-feeders and coexisting infauna: an enhancement counterexample. J Exp Mar Biol Ecol 125:33–42
Dean D (1978) Migration of the sandworm Nereis virens during winter nights. Mar Biol 45:165–173
Dittmann S (1990) Mussel beds—amensalism or amelioration for intertidal fauna? Helgolander Meeresun 44:335–352
Dunning JB, Danielson BJ, Pulliam HR (1992) Ecological processes that affect populations in complex landscapes. Oikos 65:169–175
Dye AH (1998) Dynamics of rocky intertidal communities: analyses of long time series from South African shores. Estuar Coast Shelf S 46:287–305
Dye AH, Schleyer MH, Lambert G, Lasiak TA (1994) Intertidal and subtidal filter-feeders in Southern Africa. In: Siegfried WR (ed) Ecological studies—rocky shores: exploitation in Chile and South Africa. Springer, Berlin, pp 57–73
Erlandsson J, McQuaid CD (2004) Spatial structure of recruitment in the mussel Perna perna at local scales: effects of adults, algae and recruit size. Mar Ecol Prog Ser 267:173–185
Erlandsson J, Porri F, McQuaid CD (2008) Ontogenetic changes in small-scale movement by recruits of an exploited mussel: implications for the fate of larvae settling on algae. Mar Biol 153:365–373
Erlandsson J, McQuaid CD, Stanczak S (2011a) Recruit/algal interaction prevents recovery of overexploited mussel beds: indirect evidence that post-settlement mortality structures mussel populations. Estuar Coast Shelf S 92:132–139
Erlandsson J, McQuaid CD, Sköld M (2011b) Patchiness and co-existence of indigenous and invasive mussels at small spatial scales: the interaction of facilitation and competition. PLoS ONE 6(11):e26958
Fahrig L (1998) When does fragmentation of breeding habitat affect population survival? Ecol Model 105:273–292
Fahrig L (2001) How much habitat is enough? Biol Conserv 100:65–74
Fahrig L (2003) Effects of habitat fragmentation on biodiversity. Annu Rev Ecol Evol S 34:487–515
Fahrig L (2013) Rethinking patch size and isolation effects: the habitat amount hypothesis. J Biogeogr 40:1649–1663
Flather CH, Bevers M (2002) Patchy reaction-diffusion and population abundance: the relative importance of habitat amount and arrangement. Am Nat 159:40–56
Fuhlendorf S, Smeins F, Grant W (1996) Simulation of a fire-sensitive ecological threshold: case study of Ashe juniper on the Edwards Plateau of Texas, USA. Ecol Model 90:245–255
Garbary DJ, Miller AG, Williams J, Seymour NR (2014) Drastic decline of an extensive eelgrass bed in Nova Scotia due to the activity of the invasive green crab (Carcinus maenas). Mar Biol 161:3–15
Goodsell PJ, Connell SD (2008) Complexity in the relationship between matrix composition and interpatch distance in fragmented habitats. Mar Biol 154:117–125
Grant WS, Cherry MI (1985) Mytilus galloprovincialis Lmk. in southern Africa. J Exp Mar Biol Ecol 90:179–191
Gutiérrez JL, Jones CG, Strayer DL, Iribarne OO (2003) Molluscs as ecosystem engineers: the role of shell production in aquatic habitats. Oikos 101:79–90
Hammond W (2001) Factors affecting the infauna associated with mussel beds. MSc-thesis, Department of Zoology, University of Cape Town, South Africa
Hammond W, Griffiths CL (2006) Biogeographical patterns in the fauna associated with southern African mussel beds. Afr Zool 41:123–130
Hanekom N (2008) Invasion of an indigenous Perna perna mussel bed on the south coast of South Africa by an alien mussel Mytilus galloprovincialis and its effect on the associated fauna. Biol Invasions 10:233–244
Hansen K, Kristensen E (1998) The impact of the polychaete Nereis diversicolor and enrichment with macroalgal (Chaetomorpha linum) detritus on benthic metabolism and nutrient dynamics in organic-poor and organic-rich sediment. J Exp Mar Biol Ecol 231:201–223
Harley CDG, Anderson KM, Lebreton CA-M, MacKay A, Ayala-Diaz M, Chong SL, Pond LM, Amerongen Maddison JH, Hung BHC, Iversen SL, Wong DCM (2013) The introduction of Littorina littorea to British Columbia, Canada: potential impacts and the importance of biotic resistance by native predators. Mar Biol 160:1529–1541
Harris JM, Branch GM, Elliott BL, Currie B, Dye A, McQuaid CD, Tomalin BJ, Velasquez C (1998) Spatial and temporal variability in recruitment of intertidal mussels around the coast of southern Africa. S Afr J Zool 33:1–11
Hawkins SJ, Jones HD (1992) Rocky shores. Marine field course guide. Immel Publishing, London
Hooper DU, Chapin FS, Ewel JJ, Hector A, Inchausti P, Lavorel S, Lawton JH, Lodge DM, Loreau M, Naeem S, Schmid B, Setälä H, Symstad J, Vandermeer J, Wardle DA (2005) Effects of biodiversity on ecosystem functioning: a consensus of current knowledge. Ecol Monogr 75:3–35
Ieno EN, Solan M, Batty P, Pierce GJ (2006) How biodiversity affects ecosystem functioning: roles of infaunal species richness, identity and density in the marine benthos. Mar Ecol Prog Ser 311:263–271
Iwasaki K (1995) Comparison of mussel-bed community between two intertidal mytilids Septifer virgatus and Hormomya mutabilis. Mar Biol 123:109–119
Jordaan TN (2010) The effect of mussel bed structure on the associated infauna in South Africa and the interaction between mussel and epibiotic barnacles. MSc-thesis, Rhodes University
Kiessling W (2005) Long-term relationships between ecological stability and biodiversity in Phanerozoic reefs. Nature 433:410–413
Koivisto ME, Westerbom M (2010) Habitat structure and complexity as determinants of biodiversity in blue mussel beds on sublittoral rocky shores. Mar Biol 157:1463–1474
Koivisto ME, Westerbom M (2012) Invertebrate communities associated with blue mussel beds in a patchy environment: a landscape ecology approach. Mar Ecol Prog Ser 471:101–110
Koivisto ME, Westerbom M, Arnkil A (2011) Quality or quantity: small-scale patch structure affects patterns of biodiversity in a sublittoral blue mussel community. Aquat Biol 12:261–270
Kostylev V (1996) Spatial heterogeneity and habitat complexity affecting marine littoral fauna. PhD-thesis, Göteborg University, Sweden
Kostylev V, Erlandsson J (2001) A fractal approach for detecting spatial hierarchy and structure on mussel beds. Mar Biol 139:497–506
Kostylev VE, Erlandsson J, Yiu Ming M, Williams GA (2005) The relative importance of habitat complexity and surface area in assessing biodiversity: fractal application on rocky shores. Ecol Complex 2:272–286
Kraufvelin P (2013) Pending possible establishment of Littorina littorea in the Pacific: to know the remedy, know thine enemy. Mar Biol 160:1525–1527
Kraufvelin P (2014) Digging for and grabbing the answers: novel whole-ecosystem approach quantifies damage and reveals mechanisms of invader-driven habitat destruction. Mar Biol 161:1–2
Kraufvelin P, Salovius S, Christie H, Moy FE, Karez R, Pedersen MF (2006a) Eutrophication-induced changes in benthic algae affect the behaviour and fitness of the marine amphipod Gammarus locusta. Aquat Bot 84:199–209
Kraufvelin P, Moy FE, Christie H, Bokn TL (2006b) Nutrient addition to experimental rocky shore communities revisited: delayed responses, rapid recovery. Ecosystems 9:1076–1093
Kraufvelin P, Lindholm A, Pedersen MF, Kirkerud LA, Bonsdorff E (2010) Biomass, diversity and production of rocky shore macroalgae at two nutrient enrichment and wave action levels. Mar Biol 157:29–47
Kruskal JB (1964) Multidimensional scaling by optimizing goodness of fit to a nonmetric hypothesis. Psychometrika 9:1–27
Lasiak TA, Barnard TCE (1995) Recruitment of the brown mussel Perna perna onto natural substrata: a refutation of the primary/secondary settlement hypothesis. Mar Ecol Prog Ser 120:147–153
Lasiak TA, Field JG (1995) Community-level attributes of exploited and non-exploited rocky infratidal macrofaunal assemblages in Transkei. J Exp Mar Biol Ecol 185:33–53
Matias MG (2013) Macrofaunal responses to structural complexity are mediated by environmental variability and surrounding habitats. Mar Biol 160:493–502
McQuaid CD, Lindsay JR (2005) Interacting effects of wave exposure, tidal height and substratum on spatial variation in densities of mussel Perna perna plantigrades. Mar Ecol Prog Ser 301:173–184
McQuaid CD, Lindsay TL (2007) Wave exposure effects on population structure and recruitment in the mussel Perna perna suggest regulation primarily through availability of recruits and food, not space. Mar Biol 151:2123–2131
McQuaid CD, Phillips TE (2000) Limited wind-driven dispersal of intertidal mussel larvae: in situ evidence from the plankton and the spread of the invasive species Mytilus galloprovincialis in South Africa. Mar Ecol Prog Ser 201:211–220
McQuaid CD, Lindsay JR, Lindsay TL (2000) Interactive effects of wave exposure and tidal height on population structure of the mussel Perna perna. Mar Biol 137:925–932
Menge BA, Branch GM (2000) Rocky intertidal communities. In: Bertness MD, Gaines SD, Hay M (eds) Marine community ecology. Sinauer Associates, Sunderland, pp 221–251
Mileikovsky SA (1974) On predation of pelagic larvae and early juveniles of marine bottom invertebrates by adult benthic invertebrates and their passing alive through their predators. Mar Biol 26:303–311
Muradian R (2001) Ecological thresholds: a survey. Ecol Econ 38:7–24
Naeem S, Thompson LJ, Lawler SP, Lawton JH, Woodfin RM (1994) Declining biodiversity can alter the performance of ecosystems. Nature 368:734–737
Nicastro KR, Zardi GI, McQuaid CD, Pearson GA, Serrao EA (2012) Love thy neighbour: group properties of gaping behaviour in mussel aggregations. PLoS ONE 7(10):e47382
Norling P, Kautsky N (2007) Structural and functional effects of Mytilus edulis on diversity of associated species and ecosystem functioning. Mar Ecol Prog Ser 351:163–175
Norling P, Kautsky N (2008) Patches of the mussel Mytilus sp. are islands of high biodiversity in subtidal sediment habitats in the Baltic Sea. Aquat Biol 4:75–87
O’Connor NE, Crowe TP (2007) Biodiversity among mussels: separating the influence of sizes of mussels from the ages of patches. J Mar Biol Assoc UK 87:551–557
Peake AJ, Quinn GP (1993) Temporal variation in species-area curves for invertebrates in clumps of an intertidal mussel. Ecography 16:269–277
Pettersson P (2006) Role of Mytilus for biodiversity in sediment habitats of the Skagerrak and Baltic Sea. PhLic-thesis, Stockholm University, Sweden
Porri F, Jordaan T, McQuaid CD (2008) Does cannibalism of larvae by adults affect settlement and connectivity of mussel populations? Estuar Coast Shelf S 79:687–693
Prins TC, Smaal AC, Richard FD (1998) A review of the feedbacks between bivalve grazing and ecosystem processes. Aquat Ecol 31:349–359
Reaugh-Flower K, Branch GM, Harris JM, McQuaid CD, Currie B, Dye A, Robertson B (2011) Scale-dependent patterns and processes of intertidal mussel recruitment around southern Africa. Mar Ecol Prog Ser 434:101–119
Rius M, McQuaid CD (2006) Wave action and competitive interaction between the invasive mussel Mytilus galloprovincialis and the indigenous Perna perna in South Africa. Mar Biol 150:69–78
Rius M, McQuaid CD (2009) Facilitation and competition between invasive and indigenous mussels over a gradient of physical stress. Basic Appl Ecol 10:607–613
Scheffer M, Carpenter S, Foley JA, Folke C, Walker B (2001) Catastrophic shifts in ecosystems. Nature 413:591–596
Seed R (1996) Patterns of biodiversity in the macro-invertebrate fauna associated with mussel patches on rocky shores. J Mar Biol Assoc UK 76:203–210
Siegfried WR, Hockey PAR, Crowe AA (1985) Exploitation and conservation of brown mussel stocks by coastal people of Transkei. Environ Conserv 12:303–307
Sousa R, Gutiérrez JL, Aldridge DC (2009) Non-indigenous invasive bivalves as ecosystem engineers. Biol Invasions 11:2367–2385
Steffani CN, Branch GM (2003) Temporal changes in an interaction between an indigenous limpet Scutellastra argenvillei and an alien mussel Mytilus galloprovincialis: effects of wave exposure. Afr J Mar Sci 25:213–229
Steffani CN, Branch GM (2005) Mechanisms and consequences of competition between an alien mussel, Mytilus galloprovincialis, and an indigenous limpet, Scutellastra argenvillei. J Exp Mar Biol Ecol 317:127–142
Thiel M, Ullrich N (2002) Hard rock versus soft bottom: the fauna associated with intertidal mussel beds on hard bottoms along the coast of Chile, and considerations on the functional role of mussel beds. Helgoland Mar Res 56:21–30
Tilman D (1996) Biodiversity: population versus ecosystem stability. Ecology 77:350–363
Trzcinski MK, Fahrig L, Merriam G (1999) Independent effects of forest cover and fragmentation on the distribution of forest breeding birds. Ecol Appl 9:586–593
Tsuchiya M, Nishihira M (1985) Islands of Mytilus as a habitat for small intertidal animals: effect of island size on community structure. Mar Ecol Prog Ser 25:71–81
Tunley K (2009) State of management of South Africa’s marine protected areas. WWF South Africa Report Series—2009/Marine/001
Underwood AJ (1997) Experiments in ecology: their logical design and interpretation using analysis of variance. Cambridge University Press, Cambridge
Väinölä R, Witt JDS, Grabowski M, Bradbury JH, Jazdzewski K, Sket B (2008) Global diversity of amphipods (Amphipoda; Crustacea) in freshwater. Hydrobiologia 595:241–255
Van Erkom Schurink C, Griffiths CL (1990) Marine mussels of southern Africa—their distribution patterns, standing stocks, exploitation and culture. J Shellfish Res 9:75–85
Wahl M, Link H, Alexandridis N, Thomason J, Cifuentes M, Costello MJ, da Gama BAP, Hillock K, Hobday AJ, Kaufmann MJ, Keller S, Kraufvelin P, Krüger I, Lauterbach L, Antunes BL, Molis M, Nakaoka M, Nyström J, bin Radzi Z, Stockhausen B, Thiel M, Vance T, Weseloh A, Whittle M, Wiesmann L, Wunderer L, Yamakita T, Lenz M (2011) Re-structuring of marine communities exposed to environmental change: a global study on the interactive effects of species and functional richness. PLoS ONE 6(5):e19514
Wilson GDF (2008) Global diversity of Isopod crustaceans (Crustacea; Isopoda) in freshwater. Hydrobiologia 595:231–240
Zaiko A, Daunys D, Olenin S (2009) Habitat engineering by the invasive zebra mussel Dreissena polymorpha (Pallas) in a boreal coastal lagoon: impact on biodiversity. Helgoland Mar Res 63:85–94
Acknowledgments
This work is based upon research supported by the Swedish Research Council and SIDA/Sarec (to J.E.), the South African Research Chairs Initiative of the Department of Science and Technology, the National Research Foundation and the Academy of Finland (126831), the latter to M.W. The kind help from students and staff at Rhodes University, who participated in the field and laboratory work, is gratefully acknowledged.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by M. G. Chapman.
Rights and permissions
About this article

Cite this article
Jungerstam, J., Erlandsson, J., McQuaid, C.D. et al. Is habitat amount important for biodiversity in rocky shore systems? A study of South African mussel assemblages. Mar Biol 161, 1507–1519 (2014). https://doi.org/10.1007/s00227-014-2436-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00227-014-2436-4