
Abstract
Phyto-microbiome are the microorganisms (fungi and bacteria) associated with all major plant components such as flowers, stems, roots, leaves, and fruits. They form symbiotic association with the plant, inhabit the intra- and intercellular positions without harming the host and frequently profit the host plant. They indicate the complex connections within the host plants involving the symbiotic, mutualistic relationship, and rarely the parasitism relationship. They are omnipresent and are known to improve the nutrient enrichment and growth of the plant. They not only produce root exudates but also release signal molecules which regulate various biochemical and genetic activities. They provide the immunity to plants from pests and insects and enhance the ability of plants to tolerate the impacts of abiotic and biotic stress and also produce bioactive compounds and phytohormones of biotechnological interest. In this book chapter, we will review the advent role of microbiome in plant growth and development. Efforts have been made to summarize the utilization of various hormones to mitigate the effects of various environmental stresses on cultivated plant communities. The final sections of the book chapter describe the applications of phyto-microbiome in twenty-first century and the clear out cut to commercialize of a phyto-microbiome-based technology.
Simranjeet Singh, Daljeet Singh Dhanjal, Satyender Singh, Vijay Kumar—Equally contributed.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others

Keywords
14.1 Introduction
Soil is considered to be the reservoir for various microbes and organisms like ants, moles, and nematode (Dhanjal et al. 2017). Recent advances in sequencing techniques and elevation in number of microbial libraries have expanded the aura of the tree of life, as it is dominated by microbes (Anand et al. 2019). The horizontal gene transfer and mutation events have evolved wide array of variation among the microbial community. This significantly increases the diversity within species and endows them their functional characteristic (Bengtsson-Palme et al. 2017a). Here, soil plays the major role in recycling the nutrients (phosphorus and nitrogen) and imparting protection against abiotic and biotic stress (Dhanjal et al. 2018). Although the agricultural activities have increased the yield of crop, simultaneously it has also deteriorated the biological and physical properties of the soil (Gomiero 2017). Even usage of fertilizer aids in maintaining the soil fertility, but on tillage microbial communities get disrupted (Dong et al. 2012). The degradation of soil due to anthropogenic activities has emerged as a global concern and sustainable agriculture has become the need of the time for sustaining the life of humans on this earth. Thus, to sustain the life, plant–microbiome plays significant role in improving soil quality and plant growth, and providing resistance from stress (Rashid et al. 2016; Kumar et al. 2019a).
Plant–microbiome is essential as they contain distinct properties like production of secondary metabolites as well as phytohormone and nitrogen fixation and many more. Therefore, it represents the microbial community which is directly or indirectly associated with plants. Hence, they have been generally characterized into epiphytic, endophytic, and rhizosphere microbiome (Igiehon and Babalola 2018a; Kour et al. 2019; Kumar et al. 2019b; Rana et al. 2019a; Rana et al. 2019b). The soil contains both types of microbes, i.e., pathogenic and nonpathogenic in nature (Finkel et al. 2017). Nonpathogenic involves the symbiotic and neutral microbes which plays variety of roles in diverse fields like biodegradation, biofuel production, biocontrol, biotransformation, seed production, phytoremediation, and many more (Dwibedi and Saxena 2019). These potentials of microbes prompt us to understand the hyper-diversity of these unexplored plant microbial communities, not only for sustaining the ecosystem but also to preserve these biodiverse microbial communities beneficial for mankind (Braga et al. 2016). Hence, untapping and deep understanding of these plant microbes as whole have become important to explore the positive interactions for sustainable agriculture, especially during microbiome-dependent cropping approach (Busby et al. 2017). In this chapter, we will discuss the challenges and efforts put forward to advance our knowledge about different properties of microbes and how these properties affect plants. Further, we will also discuss about the soil microbiome improving the crop production.
14.2 Composition and Function of Plant Microbiome
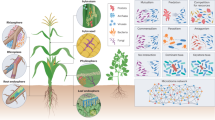
Plants are surrounded by a variety of microbes and vary according to their location like anthosphere, carposphere, phyllosphere, rhizosphere, and spermosphere (Shade et al. 2017) (Fig. 14.1). Here, we categorized the bacteria into three broad categories as follows.

Compartmentalization of plant microbiome
14.3 Plant Microflora Below the Ground
Root microflora generally get horizontally transferred as they are predominantly present in soil (Lareen et al. 2016). Most dominating microbes belong to the family of Acidobacteria, Actinobacteria, Bacteroidetes, Planctomycetes, Proteobacteria, and Verrucomicrobia (Mendes et al. 2013; Kour et al. 2019; Suman et al. 2016; Verma et al. 2017b). There is a possibility of vertical transmission through seeds. Seeds also serve as reservoir for microbes, which allows them to multiply in the roots during plant development (Shahzad et al. 2018). Root system of plant provides distinctive niche to soil microbes residing in the rhizosphere (roots and certain portion above the ground) (Raaijmakers et al. 2009). Recently, Donn with his colleagues reported about the changes in bacterial community surrounding the roots of wheat and found 10-fold increase in population of actinobacteria, copiotrophs, oligotrophs, and pseudomonads at rhizosphere. Moreover, they also reported that over time, variation takes place in rhizoplane and rhizosphere community but there is no variation in soil microbial population (Donn et al. 2015). Similar results were also seen for Brachypodium distachyon rhizosphere which reported about the predominance of species of Burkholderiales, Sphingobacteriales, and Xanthomonadales family (Kawasaki et al. 2016). There have been reports which claimed that root exudates like amino acids, fatty acids, organic acids, plant growth regulators, phenolics, putrescine, sterols, sugars, and vitamins also affect microbes present in the rhizosphere, proclaimed as rhizosphere effect (Hunter et al. 2014). For example, secondary metabolite benzoxazinoids (BXs) synthesized by the roots of maize amends the composition of root-associated microbes, out of which members of Actinobacteria and Proteobacteria family were highly affected (Neal et al. 2012; Kudjordjie et al. 2019). Moreover, other researchers are investigating the mechanisms responsible for the assembly of microbial community and effects of substrate and root exudation on microbial community. Hence, this confirms that composition of rhizosphere microbes gets influenced by plants species and root exudates (Jacoby et al. 2017; Yadav et al. 2015a; Yadav et al. 2015b).
Various bacterial endophytes have colonized the roots of plants internally (Santoyo et al. 2016). These bacterial endophytes enter the root tissues via passive processes or root abrasion and active mechanisms (Santos et al. 2018). The transmission and colonization of these bacterial endophytes inside the plant depend on various factors like ability of endophytes to colonize plant and distribution of plant resources (Kandel et al. 2017). Various microbes have gained access to root tissues, for instance, Acidobacteria, Actinobacteria, Bacteroidetes, Chloroflexi, Firmicutes, Gemmatimonadetes, Planctomycetes, Proteobacteria, and Verrucomicrobia are the bacterial taxa which have gained access to grapevine roots (Morgan et al. 2017; Verma et al. 2016; Yadav et al. 2016). On the other hand, members of Bradyrhizobiaceae, Comamonadaceae, Rhizobiaceae, and Streptomycetaceae family are the one predominantly found in rice roots (Edwards et al. 2015).
14.4 Plant Microflora Above the Ground
Floral parts, leaves, and vegetative parts present above the ground provides distinctive environment for epiphyte and endophyte microbes (Frank et al. 2017). The endophytes predominantly translocate themselves through xylem to different parts of plants, which can either be fruit, leaves, or stem (Hardoim et al. 2015). With respect to location on the plant, aboveground microbes distribute themselves with other members of endophytic community (Nair and Padmavathy 2014). It has been observed that phyllosphere microbes obtained from the soil get influenced by external stimuli and later shows the profound effect (Carvalho and Castillo 2018). Subsequently, various microbes have been found in the phyllosphere and endosphere up to species level (Schlaeppi and Bulgarelli 2015). For example, on analyzing the carposphere and phyllosphere microbes of grapevine uncovered the presence of Acinetobacter, Bacillus, Citrobacter, Curtobacterium, Enterobacter, Erwinia, Frigoribacterium, Methylobacterium, Pantoea, and Pseudomonas species (Kecskeméti et al. 2016; Verma et al. 2017a; Verma et al. 2017b; Yadav et al. 2019; Yadav et al. 2018a; Yadav et al. 2018b; Yadav et al. 2018c). On the other hand, endophyte analysis of grape berries uncovered that Bacillus, Burkholderia, Dyella, Mesorhizobium, Propionibacterium, Pseudomonas, and Ralstonia species are the dominant (Campisano et al. 2014).
There is a study conducted on 300 different varieties of maize to analyze the leaf microbiome that revealed the predominance of methylobacteria and sphingomonads taxa (Wagner et al. 2019), whereas in apple flowers there is predominance of Enterobacteriaceae and Pseudomonas species (Steven et al. 2018). Various other studies conducted on almond, apple, grapefruit, pumpkin flower, and pumpkin also revealed the abundance of Pseudomonas species (Aleklett et al. 2014). Lately, seed-associated microbes have highlighted the abundance of Actinobacteria, Bacteroidetes, Firmicutes, and Proteobacteria (Qiao et al. 2017). Usually, microbes found on aboveground have originated from air, seed, and soil and adapted themselves to sustain their life on or inside the tissue of plant, where various factors like environmental condition, soil, and management influence microbial composition (Raaijmakers et al. 2009). Compartment-specific assembly with the host illustrates the strong relationship among the aboveground microbes and plant on the basis of functionality, but still there is a need to understand this association deeply (Garcia and Kao-Kniffin 2018). Aboveground microbes and endophytes are recognized for their ability to alleviate stress tolerance, increase disease resistance, and promote plant growth (Kumar and Verma 2018).
14.5 Satellite and Core Microflora
Core microflora or core plant–microbiome (CPM) refers to those microbes that closely linked with particular type of plant and are independent of soil and environmental conditions (Lakshmanan et al. 2014). Bradyrhizobium, Microvirga, and Sphingobium were found to be the core microbiome of Solanum tuberosum (potato) (Pfeiffer et al. 2016). Another study revealed that Hyphomicrobiaceae, Micrococcaceae, and Pseudomonadaceae sps. are the core microbes found in grapevine (Zarraonaindia et al. 2015). The CPM contains those microbial taxa that are essential for plant and have established themselves in plant during the course of evolution (Jacoby et al. 2017). The enhancement of microbial taxa through evolutionary selection process contains the genes responsible for survival and holobiont of the plant (Rosenberg and Zilber-Rosenberg 2018).
Distinctively, the microbial taxa found in less affluence and fewer locations are referred to as satellite taxa. They are described based on their habitat, geography of habitat, and their regional population. These taxa are perceived as the regulators of important mechanisms in an ecosystem (Banerjee et al. 2018). Few researchers found that microbial taxa having the scarce population play a crucial role in obstructing the entrance of undesirable microbial taxa in soil. Parallelly, the scarce bacterial population produces different antifungal chemicals which guard the plants against soil-induced infectious organisms (Ciancio et al. 2016). Various researches demonstrate that forfeiture of scarce microbial population compromises with plants’ efficiency (Garcia and Kao-Kniffin 2018). Researchers also found that microbial taxa regulate vital mechanisms that can be unequal to their population number (Pineda et al. 2017). Numerous ecological factors should be taken into account to comprehend mechanisms of plant–microbiome taken in response to certain environmental tensions (Braga et al. 2016).
14.6 Factors Affecting Plant Microbiota
The microbial organization in plant’s tissues is linked with various organic and inorganic constituents. These constituents can be type, pH, salinity, structure, moisture, organic matter, and exudates of soil, respectively (Jacoby et al. 2017; Yadav et al. 2015c; Yadav and Yadav 2018). These constituents above are important for parts of the plants which are present beneath the ground level. Other constituents such as infectious agents, environmental conditions, and human activities affect the microbial community of above- and belowground plant parts (Mendes et al. 2013). Using high-throughput approaches like a shotgun and 16S rRNA sequencing, Bulgarelli with his colleagues examined the root microbial community of various barley species and concluded that root exudates and innate immune system determine the organization of root microbiota (Bulgarelli et al. 2015). Various host-influence factors such as growth stage, age, and wellness of plants affect organization of plant microbiota by the formulation of root exudates and plant signaling processes (like induced and acquired systemic resistance, respectively) (Ortíz-Castro et al. 2009).
14.7 Function and Role of Plant Microbiome
Plant–microbiome consists of all neutral, beneficial as well as pathogenic microbes. There are few plant growth-promoting (PGP) bacteria which promote the growth of plant by synthesizing different phytohormones such as gibberellin, cytokinin, and auxin (Egamberdieva et al. 2017; Yadav 2017a, b; Yadav 2019; Yadav et al. 2017a). However, few PGPB produce ACC deaminase (1-aminocyclopropane-1-carboxylate deaminase) which regulate the level of ethylene (stress hormone) in plant (Glick 2014). Literature survey revealed that Arthrobacter spp., Bacillus spp., and Pseudomonas spp. are the ACC deaminase producer, which enables them to enhance the growth of plant (Souza et al. 2015). On analyzing the root microflora of soybean and wheat, it showed the dominance of Pantoea spp., Paraburkholderia spp., and Pseudomonas spp. These species promote the plant growth because of the properties like phytohormones (ACC deaminase, indole acetic acid), nitrogen fixation, and phosphate solubilization mechanism to enhance the stress tolerance and improve nutrient uptake (Rascovan et al. 2016). Schematic representation of the function performed by microbiome is illustrated in Fig. 14.2.

Schematic overview of function and impact of plant–microbiome on the host plant
On the other hand, there are few bacteria which show disease symptoms because of the production of phytohormones and phytotoxic compounds (Olanrewaju et al. 2017). For instance, Pseudomonas syringae is a plant pathogen which affects various plants like green bean, olive, tobacco, and tomato (Hirano and Upper 2000). Erwinia amylovora, another pathogenic bacterium, causes fire blight disease in ornamental plants. Ralstonia solanacearum, Xylella fastidiosa, and Xanthomonas sps. are the bacterial species that are associated with various diseases of banana and potato (Vrancken et al. 2013). There are certain factors which determines the severity as well as outcome of disease like biotic factors (like plant microflora), favorable environmental factor, host susceptibility, and population size of pathogens (Lareen et al. 2016). Both aboveground plant microflora and belowground microflora have been found to increase the resistance in host via commensal interactions of pathogen or due to modification in plant defense system (Igiehon and Babalola 2018b).
The plant microbiota uses diverse biocontrol mechanism like antibiotic production, siderophore production, lytic enzyme production, and pathogen-inhibiting volatile compound production to prevent the disease and pathogenic invasion (Compant et al. 2005). Few plants modulate the hormone level and induce resistance in plant to protect them from plant pathogens. Therefore, agricultural lands containing disease-suppressing microbes build pressure on pathogens and make it the disease-suppressive soil (Gómez Expósito et al. 2017). Particularly, Bacillus, Burkholderia, Enterobacter, Paenibacillus, Pantoea, Paraburkholderia, Pseudomonas, and Streptomyces have been found to play the main role in pathogen suppression (Yadav et al. 2017b). Carrión and his colleagues revealed that Paraburkholderia graminis PHS1 produces cysteine desulfurase and dimethyl sulfoxide reductase (sulfurous volatile compounds), which provide suppressive ability against fungal root pathogens (Carrión et al. 2018). On the other hand, Durán and his colleagues highlighted the role of endospheric bacteria belonging to Enterobacter and Serratia family, which have the biocontrol activity against Gaeumannomyces graminis (Durán et al. 2017).
14.8 Advancements in Plant–Microbiome Research (PMR)
Prediction about the advances in plant–microbiome research is quite difficult (Thijs et al. 2016). But following are the three points which help us in shaping the future of both applied and basic studies of soil microbiome.
14.8.1 Upgradation of Culturing Techniques
The advent of molecular techniques has rapidly increased the genomic data of uncultured microbial taxa persisting in the soil, for which closely related strains are currently unknown (Warinner et al. 2017). There is an exponential increase in data because of culture-independent approach like single-cell genomics or by the assembling of individual genome obtained via metagenome analysis (Blainey 2013). Although we have the genomic information of all the microbes present in the soil, still we lack to understanding about their functionality. Therefore, categorizing them in ecological taxa considering their genomic data is risky (Peršoh 2015). Additionally, the slow-growing microbe is another challenge. Therefore, approaches which use genomic data to intercept the information about ideal cultivation and isolation of uncultured microbes will unquestionably be helpful for the field to me in forward direction (Streit and Schmitz 2004).
14.8.2 Role of Viruses in Soil Microbiome
Approximately, 107–109 virus particles are found in one gram of soil, and this number is less than the number of viruses found in the aquatic ecosystem (Jacquet et al. 2010). Apparently, these viruses found in soil are highly diverse and remained unexplored (Williamson et al. 2017). Marine studies have revealed that phages play chief role in nutrient dynamics, as they maintain the nutrient dynamics by killing the 20–40% microbial population present in the water column (Sime-Ngando 2014). Various researches have demonstrated soil to be the reservoir for phages which specifically targets bacteria like Rhizobium spp. and Xanthomonas spp., still the effect of these viruses on activity and composition of soil remains comprehended (Buttimer et al. 2017). According to statistics, > 90% of viruses found in soil mainly get absorbed by clay as well as other soil surfaces, which leaves very low number of viruses in soil. Additionally, it is also uncertain that these viruses are capable enough to infect the plant–microbiome (Andika et al. 2016). Due to advancement in viral metagenomic field and development of new techniques, there is an enumeration in viral population, and it has prompted us to explore new viral community as well as study the effect of these viruses on microbial processes and populations (Jacquet et al. 2010). Basically, it clear to develop a holistic understanding of how these microbes (directly or indirectly) interact with each other instead of studying the individual microbial group (Tshikantwa et al. 2018).
14.8.3 Importance of Horizontal Gene Transfer
There are three main mechanisms of horizontal gene transfer, i.e., conjugation, transduction, and transformation, by which microbes their gene from one generation to another (Von Wintersdorff et al. 2016). Genes contains the sequence of various different traits like arsenic detoxification, antibiotic resistance, and xenobiotic degradation. Therefore, mobile genes may cause evolution of new phenotypic characteristic and may generate close relation with dissimilar genomes (Janssen et al. 2005). However, horizontal gene transfer can impose dilemma during the attempt of linking particular gene to targeted phylogenetic lineages. This definite control of horizontal gene transfer, its frequency in soil microbial communities, and their effect on the soil population are the topics which nurtures this expedition and motivate us to work in this direction (Andrews et al. 2018).
14.9 Use of Plant–Microbiome as a Biocontrol Agent
Worldwide, plant diseases have been held responsible for farmers’ losses. FAO stated that 25% of crop loss occurs due to plant diseases and pests. Therefore, there is need for control plant disease (Savary et al. 2012). To achieve this, specific plants which are resistant to various diseases are bred along with different cultivation techniques like use of pathogen-free seeds, crop rotation, or by using biological and chemical agents (Cheng and Cheng 2015). All these plant protection methods influence the microbiome of plant; hence, information gathered from plant protection strategies should be taken into consideration during cultivation (Berg et al. 2014). In various cases, the onset of disease takes place due to imbalance of plant–microbiome and obstructs the desired objective of whole microbiome. Hence, analyzing metabolic pathway and plant–microbiome of the host plant unlock new opportunities to advance biocontrol approaches (Thijs et al. 2016).
Overall, better understanding unfolds the information about the diverse microbes which aid in plant growth and produce antagonistic chemicals against phytopathogens (Bargaz et al. 2018). Mosses are known to have unique microbial diversity and harbor microbes with exceptional antagonist potential because of their ecology (Bragina et al. 2012). Moreover, medicinal plants are also to be known to nurture diverse microbes as their secondary metabolites determine the composition of microbiome (Berg et al. 2015). Because of large diversity among microbes, it is believed that endophytes, especially seed endophytes, can emerge as the unique biocontrol agent (Ryan et al. 2008). Till now, largely bacteria as well as fungi have been used for biocontrol purposes (Whipps 2001). Recently, Archaea have been exploited to become the member of plant–microbiome but its biocontrol potential is still under evaluation (Müller et al. 2015).
As stated above, microbes linked with plant develops a network which gets influence during pathogen invasion. Therefore, these networks will enable us to develop new model and enhance disease management of the plant–soil microbes (Hassani et al. 2018). Even researchers have developed a framework to decipher these microbiome networks and showed the benefit of these frameworks to test their hypothesis for targeted microbes responsible for particular plant disease. They proposed four types of network analysis: (a) first type involves the general analysis of network to discover the aspiring taxa to maintain the existing microbial community; (b) second type involves the analysis majorly focusing on host as well as plant response; (c) third type involves the identification of pathogenic taxa which are directly or indirectly associated with prior known taxa of pathogens; and (d) last type involves the identification of those taxa which are associated with disease (Poudel et al. 2016).
Now, most of the biocontrol agents are screened through in vitro antagonisms effective against particular pathogen. As this screening allows the identification of potent biocontrol agent, still intense debate is going on about this screening approach (Larran et al. 2016). The chief reason is that biocontrol and plant growth-promoting (PGPR) microbes during in vivo evaluation show no antagonisms, whereas in the case of in vitro studies it shows modest result (Beneduzi et al. 2012). Different high-throughput techniques have been developed for plant assays but were found to be ineffective as includes artificial characters (Rasheed et al. 2017). For instance, rapeseed treated with Paenibacillus showed plant growth in natural soil, whereas no negative effect was observed in sterile soil under gnotobiotic conditions. Therefore, there is need for testing the potential of biocontrol and plant growth-promoting strains in soil as well as evaluation of their effect on soil type and plant species also be done (Bashan et al. 2014). Another research study showed the presence of similar genera of Pseudomonas in lettuce rhizosphere in three different soil samples from the field. This confirms that soil types do not play any role in biocontrol activity and rhizo-competence (Schreiter et al. 2018).
Additionally, targeted enhanced diversity is also being exploited as biomarker for screening purpose by researchers. These applicability and development in biological control is attaining attention globally (Atanasov et al. 2015), whereas previously only one microbe was used in biocontrol strategies, which explains about inconsistency. But recent advancements have enabled us to develop microbiome-dependent biocontrol strategy (Ab Rahman et al. 2018). In 1999, Emmert and Handelsman stated Gram-positive bacteria to be potential candidate in biocontrol practices. On assessing the registered products for biocontrol and along with those that are in pipeline, almost all the products are bacillus based (Emmert and Handelsman 1999). There is technical reason for using Bacillus sps. as the base product in contrast to other Gram-negative bacteria, as form spores under unfavorable condition for survival. This spore formation increases their shelf-life (Berg et al. 2017). On examining the plant-associated microbes which harbor large number of bacterial species, it make us realize that we are seeing the very small portion of taxonomic diversity for biocontrol purpose (Chowdhury et al. 2017).
Moreover, the researchers have reported that 2% mean population is occupied by Bacillus species from the total bacterial community in the soil (Li et al. 2017). As an exception, another researcher published a paper in which they stated that 37% of Bacillus and Paenibacillus species are found in arid soil (Köberl et al. 2013). Still, the debate is going on comprehending the role of Bacillus on the plant and against its pathogen, or does it trigger the plant growth hormones or induce resistance as they are involved in plant–microbe interaction. It is already known that Bacillus and Paenibacillus strains synthesize various antibiotics and secondary metabolites (Mhlongo et al. 2018). As per studies, strains and spores of both Bacillus and Paenibacillus are predominantly found in the environment and this might be reason for reducing number of plant-associated microbial community. Hence, more emphasis is given on increasing the microbial diversity within products during biocontrol activity. This prompts to develop new strategies to explore the antagonistic potential of microbes associated with plants (Yadav et al. 2017c).
Nowadays, antibiotic resistance is becoming the global concern and making our treatment ineffective as a result people is suffering from disability, prolonged illness, and at last death (Laxminarayan et al. 2013). Regrettably, strategies used in agriculture like use of antibiotics in livestock are increasing the resistance level. These livestock aid in transferring the resistance in plant production system, which causes the enrichment of resistant bacteria in the rhizosphere (Founou et al. 2016), even though many microbes have developed resistance and are highly prevalent in the environment. Therefore, risk management studies to suppress the emergence of resistant microbes have become our first priority (Larsson et al. 2018). Though the different biocontrol products have shown lower persistence on soil and plant, usage of spore-forming bacteria will change the whole scenario (Paul et al. 2019). Additionally, soil microbes are found to be bacterial antagonists in nature. In particular, Paenibacillus have been found to contain not only the antibiotic synthesis gene but also contain antibiotic gene, justifying about their antagonistic ability within the microbiome (Cycoń et al. 2019). Therefore, further investigation is required to understand the mobilization of resistant gene through different ways of horizontal gene transfer (Bengtsson-Palme et al. 2017b).
14.10 Studies Related to Plant Microbiomes
Even though plant–microbiome is a broad topic, microbial group associated with plant is an active research area. The major reason is the exponential increase in publications in recent year on this topic as well as research targeting the niche of specific plant and how interaction takes place among particular microbial community (Naylor and Coleman-Derr 2018). Additionally, it has been observed that microbial assembly varies according to environmental conditions and plant species. This creates curiosity to learn more about how microbes are acquired by plant, either the microbes are subsequently passed to next generation or recruited from the environment (Yu and Hochholdinger 2018).
In spite of the role of the microbes in promoting the plant growth and nutrient cycle, we still require to gain more insight about microbial interaction which makes up the plant microbiome. This will allow us to comprehend the functioning of plant microbiome to changing agricultural practices and environmental conditions (Garcia and Kao-Kniffin 2018).
Various studies are being conducted to extend the link the shift in environmental condition with microbiome response (Purahong et al. 2018). For instance, the researcher conducted on the grape vineyard using culture-independent method revealed the presence of dynamic microbial community on different stages of plant lifecycle, enlightening the effect of agrochemicals on the plant (Morgan et al. 2017). Numerous studies have associated microbiome with physiology of plant, as microbes found in plant–microbiome in desert aid in providing the resistance to plant to survive in unfavorable conditions (Vurukonda et al. 2016). In 2012, Marasco and his colleague proposed that plant microbiomes are essential to develop resistance against drought stresses (Marasco et al. 2012). Another effective interaction was reported by another research group, where the described role of microbiome determines the composition of plant root exudates. These examples highlight the role of soil microbiome not only in plants but also in their evolutionary mechanism, regulated by host–microbe interaction (Lareen et al. 2016).
14.11 Conclusion and Future Prospects
The microbial communities possessing beneficial traits act as a powerful tool to elevate the sustainable agriculture by reducing the usage of fertilizes and combatting plant diseases to enhance the crop yield. The potential of microbiome is to elicit the plant growth, generate stress resistance, and improve plant health. Various microbial inoculants have been developed to achieve field success by either designing smart microbial consortia or engineering the microbiota with beneficial characteristics. In order to upsurge the crop yield and support the enhanced technologies, new plant breeding practices and suitable formulations are required. To achieve these goals, there is need to gather insight about the plant–microbiome interaction, microbial community of soil, and their stability during environmental changes. As single microbes are the key regulators within microbial community, therefore comprehensive investigation on these microbes along with microbial community of soil can assist us in expanding the horizon of this field. Moreover, the knowledge gained will enable us to completely comprehend the impact of these microbes on disease resistance, nutrient cycles, and yield of the crop. Furthermore, it will prompt us to explore novel strategies for microbiome engineering to move toward the sustainable agriculture.
References
Ab Rahman SF, Singh E, Pieterse CM, Schenk PM (2018) Emerging microbial biocontrol strategies for plant pathogens. Plant Sci 267:102–111
Aleklett K, Hart M, Shade A (2014) The microbial ecology of flowers: an emerging frontier in phyllosphere research. Botany 92(4):253–266
Anand P, Chopra RS, Dhanjal DS, Chopra C (2019) Isolation and characterization of microbial diversity of soil of Dhanbad coal mines using molecular approach. Res J Pharm Technol 12(3):1137–1140
Andika IB, Kondo H, Sun L (2016) Interplays between soil-borne plant viruses and RNA silencing-mediated antiviral defense in roots. Front Microbiol 7:1458
Andrews M, De Meyer S, James E, Stępkowski T, Hodge S, Simon M, Young J (2018) Horizontal transfer of symbiosis genes within and between rhizobial genera: occurrence and importance. Genes 9(7):321
Atanasov AG, Waltenberger B, Pferschy-Wenzig EM, Linder T, Wawrosch C, Uhrin P et al (2015) Discovery and resupply of pharmacologically active plant-derived natural products: a review. Biotechnol Adv 33(8):1582–1614
Banerjee S, Schlaeppi K, van der Heijden MG (2018) Keystone taxa as drivers of microbiome structure and functioning. Nat Rev Microbiol 16(9):567
Bargaz A, Lyamlouli K, Chtouki M, Zeroual Y, Dhiba D (2018) Soil microbial resources for improving fertilizers efficiency in an integrated plant nutrient management system. Front Microbiol 9:1606
Bashan Y, Bashan LE, Prabhu SR, Hernandez JP (2014) Advances in plant growth-promoting bacterial inoculant technology: formulations and practical perspectives (1998–2013). Plant Soil 378(1–2):1–33
Beneduzi A, Ambrosini A, Passaglia LM (2012) Plant growth-promoting rhizobacteria (PGPR): their potential as antagonists and biocontrol agents. Genet Mol Biol 35(4):1044–1051
Bengtsson-Palme J, Kristiansson E, Larsson DJ (2017a) Environmental factors influencing the development and spread of antibiotic resistance. FEMS Microbiol Rev 42(1):fux053
Bengtsson-Palme J, Kristiansson E, Larsson DJ (2017b) Environmental factors influencing the development and spread of antibiotic resistance. FEMS Microbiol Rev 42(1):fux053
Berg G, Grube M, Schloter M, Smalla K (2014) The plant microbiome and its importance for plant and human health. Front Microbiol 5:1
Berg G, Köberl M, Rybakova D, Müller H, Grosch R, Smalla K (2017) Plant microbial diversity is suggested as the key to future biocontrol and health trends. FEMS Microbiol Ecol 93(5):fix050
Berg G, Rybakova D, Grube M, Köberl M (2015) The plant microbiome explored: implications for experimental botany. J Exp Bot 67(4):995–1002
Blainey PC (2013) The future is now: single-cell genomics of bacteria and archaea. FEMS Microbiol Rev 37(3):407–427
Braga RM, Dourado MN, Araújo WL (2016) Microbial interactions: ecology in a molecular perspective. Braz J Microbiol 47:86–98
Bragina A, Berg C, Cardinale M, Shcherbakov A, Chebotar V, Berg G (2012) Sphagnum mosses harbour highly specific bacterial diversity during their whole lifecycle. ISME J 6(4):802–813
Bulgarelli D, Garrido-Oter R, Münch PC, Weiman A, Dröge J, Pan Y et al (2015) Structure and function of the bacterial root microbiota in wild and domesticated barley. Cell host Microb 17(3):392–403
Busby PE, Soman C, Wagner MR, Friesen ML, Kremer J, Bennett A et al (2017) Research priorities for harnessing plant microbiomes in sustainable agriculture. PLoS Biol 15(3):e2001793
Buttimer C, McAuliffe O, Ross RP, Hill C, O’Mahony J, Coffey A (2017) Bacteriophages and bacterial plant diseases. Front Microbiol 20(8):34
Campisano A, Antonielli L, Pancher M, Yousaf S, Pindo M, Pertot I (2014) Bacterial endophytic communities in the grapevine depend on pest management. PLoS ONE 9(11):e112763
Carrión VJ, Cordovez V, Tyc O, Etalo DW, de Bruijn I, de Jager VC (2018) Involvement of Burkholderiaceae and sulfurous volatiles in disease-suppressive soils. The ISME J 12(9):2307
Carvalho SD, Castillo JA (2018) Influence of light on plant-phyllosphere interaction. Front Plant Sci 9:1482
Cheng F, Cheng Z (2015) Research progress on the use of plant allelopathy in agriculture and the physiological and ecological mechanisms of allelopathy. Front Plant Sci 6:1020
Chowdhury ME, Jeon J, Rim SO, Park YH, Lee SK, Bae H (2017) Composition, diversity and bioactivity of culturable bacterial endophytes in mountain-cultivated ginseng in Korea. Sci Rep 7(1):10098
Ciancio A, Pieterse CM, Mercado-Blanco J (2016) Harnessing useful rhizosphere microorganisms for pathogen and pest biocontrol. Front Microbiol 7:1620
Compant S, Duffy B, Nowak J, Clément C, Barka EA (2005) Use of plant growth-promoting bacteria for biocontrol of plant diseases: principles, mechanisms of action, and future prospects. Appl Environ Microbiol 71(9):4951–4959
Cycoń M, Mrozik A, Piotrowska-Seget Z (2019) Antibiotics in the soil environment-degradation and their impact on microbial activity and diversity. Front Microbiol 10:338
Dhanjal DS, Chopra C, Anand P, Chopra RS (2017) Accessing the microbial diversity of sugarcane fields from Gujjarwal village, Ludhiana and their molecular identification. Res J Pharm Technol 10(10):3439–3442
Dhanjal DS, Sharma D (2018) Microbial metagenomics for industrial and environmental bioprospecting: the unknown envoy. Microbial bioprospecting for sustainable development. Springer, Singapore, pp 327–352
Dong W, Zhang X, Wang H, Dai X, Sun X, Qiu W, Yang F (2012) Effect of different fertilizer application on the soil fertility of paddy soils in red soil region of southern China. PLoS ONE 7(9):e44504
Donn S, Kirkegaard JA, Perera G, Richardson AE, Watt M (2015) Evolution of bacterial communities in the wheat crop rhizosphere. Environ Microbiol 17(3):610–621
Durán P, Jorquera M, Viscardi S, Carrion VJ, Mora MD, Pozo MJ (2017) Screening and characterization of potentially suppressive soils against Gaeumannomyces graminis under extensive wheat cropping by Chilean indigenous communities. Front Microbiol 8:1552
Dwibedi V, Saxena S (2019) Diversity and phylogeny of resveratrol-producing culturable endophytic fungi from Vitis species in India. 3 Biotech 9(5):182
Edwards J, Johnson C, Santos-Medellín C, Lurie E, Podishetty NK, Bhatnagar S (2015) Structure, variation, and assembly of the root-associated microbiomes of rice. Proc Natl Acad Sci 112(8):E911–920
Egamberdieva D, Wirth SJ, Alqarawi AA, Abd_Allah EF, Hashem A (2017) Phytohormones and beneficial microbes: essential components for plants to balance stress and fitness. Front Microbiol 8:2104
Emmert EA, Handelsman J (1999) Biocontrol of plant disease: a (Gram-) positive perspective. FEMS Microbiol Lett 171(1):1–9
Finkel OM, Castrillo G, Paredes SH, González IS, Dangl JL (2017) Understanding and exploiting plant beneficial microbes. Curr Opin Plant Biol 38:155–163
Founou LL, Founou RC, Essack SY (2016) Antibiotic resistance in the food chain: a developing country-perspective. Front Micro 23(7):1881
Frank A, Saldierna Guzmán J, Shay J (2017) Transm Bact Endophytes Microorg 5(4):70
Garcia J, Kao-Kniffin J (2018) Microbial group dynamics in plant rhizospheres and their implications on nutrient cycling. Front Micro 11(9):1516
Glick BR (2014) Bacteria with ACC deaminase can promote plant growth and help to feed the world. Microbiol Res 169(1):30–39
Gómez Expósito R, de Bruijn I, Postma J, Raaijmakers JM (2017) Current insights into the role of rhizosphere bacteria in disease suppressive soils. Front Micro 8:2529
Gomiero T (2017) Soil degradation, land scarcity and food security: Reviewing a complex challenge. Sustainability 8(3):281
Hardoim PR, Van Overbeek LS, Berg G, Pirttilä AM, Compant S, Campisano A (2015) The hidden world within plants: ecological and evolutionary considerations for defining functioning of microbial endophytes. Microbiol Mol Biol Rev 79(3):293–320
Hassani MA, Durán P, Hacquard S (2018) Microbial interactions within the plant holobiont. Microbiome 6(1):58
Hirano SS, Upper CD (2000) Bacteria in the leaf ecosystem with emphasis on Pseudomonas syringae—a pathogen, ice nucleus, and epiphyte. Microbiol Mol Biol Rev 64(3):624–653
Hunter PJ, Teakle G, Bending GD (2014) Root traits and microbial community interactions in relation to phosphorus availability and acquisition, with particular reference to Brassica. Front Plant Sci 5:27
Igiehon N, Babalola O (2018a) Rhizosphere microbiome modulators: contributions of nitrogen fixing bacteria towards sustainable agriculture. Int J Environ Res Public Health 15(4):574
Igiehon NO, Babalola OO (2018b) Below-ground-above-ground plant-microbial interactions: focusing on soybean, rhizobacteria and mycorrhizal fungi. Open Microbiol J 12:261
Jacoby R, Peukert M, Succurro A, Koprivova A, Kopriva S (2017) The role of soil microorganisms in plant mineral nutrition—current knowledge and future directions. Front Plant Sci 8:1617
Jacquet S, Miki T, Noble R, Peduzzi P, Wilhelm S (2010) Viruses in aquatic ecosystems: important advancements of the last 20 years and prospects for the future in the field of microbial oceanography and limnology. Adv Ocean Limnol 1(1):97–141
Janssen DB, Dinkla IJ, Poelarends GJ, Terpstra P (2005) Bacterial degradation of xenobiotic compounds: evolution and distribution of novel enzyme activities. Environ Microbiol 7(12):1868–1882
Kandel S, Joubert P, Doty S (2017) Bacterial endophyte colonization and distribution within plants. Microorganisms 5(4):77
Kawasaki A, Donn S, Ryan PR, Mathesius U, Devilla R, Jones A et al (2016) Microbiome and exudates of the root and rhizosphere of Brachypodium distachyon, a model for wheat. PLoS ONE 11(10):e0164533
Kecskeméti E, Berkelmann-Löhnertz B, Reineke A (2016) Are epiphytic microbial communities in the carposphere of ripening grape clusters (Vitis vinifera L.) different between conventional, organic, and biodynamic grapes? PloS One 11(8):e0160852
Köberl M, Schmidt R, Ramadan EM, Bauer R, Berg G (2013) The microbiome of medicinal plants: diversity and importance for plant growth, quality and health. Front Micro 20(4):400
Kour D, Rana KL, Yadav N, Yadav AN, Singh J, Rastegari AA, et al (2019) Agriculturally and industrially important fungi: current developments and potential biotechnological applications. In: Yadav AN, Singh S, Mishra S, Gupta A (eds) Recent advancement in white biotechnology through fungi, volume 2: perspective for value-added products and environments. Springer International Publishing, Cham, pp 1–64. https://doi.org/10.1007/978-3-030-14846-1_1
Kudjordjie EN, Sapkota R, Steffensen SK, Fomsgaard IS, Nicolaisen M (2019) Maize synthesized benzoxazinoids affect the host associated microbiome. Microbiome 7(1):59
Kumar A, Chaturvedi AK, Yadav K, Arunkumar KP, Malyan SK, Raja P, Kumar R, Khan SA, Yadav KK, Rana KL, Kour D, Yadav N, Yadav AN (2019) Fungal phytoremediation of heavy metal-contaminated resources: current scenario and future prospects. In: Yadav AN, Singh S, Mishra S, Gupta A (eds) Recent advancement in white biotechnology through fungi: volume 3: perspective for sustainable environments. Springer International Publishing, Cham, pp 437–461. https://doi.org/10.1007/978-3-030-25506-0_18
Kumar A, Verma JP (2018) Does plant—microbe interaction confer stress tolerance in plants: a review? Microbiol Res 207:41–52
Kumar M, Saxena R, Rai PK, Tomar RS, Yadav N, Rana KL, Kour D, Yadav AN (2019) Genetic diversity of methylotrophic yeast and their impact on environments. In: Yadav AN, Singh S, Mishra S, Gupta A (eds) Recent advancement in white biotechnology through fungi: volume 3: perspective for sustainable environments. Springer International Publishing, Cham, pp 53–71. https://doi.org/10.1007/978-3-030-25506-0_3
Lakshmanan V, Selvaraj G, Bais HP (2014) Functional soil microbiome: belowground solutions to an aboveground problem. Plant Physiol 166(2):689–700
Lareen A, Burton F, Schäfer P (2016) Plant root-microbe communication in shaping root microbiomes. Plant Mol Biol 90(6):575–587
Larran S, Simon MR, Moreno MV, Siurana MS, Perelló A (2016) Endophytes from wheat as biocontrol agents against tan spot disease. Biol Control 92:17–23
Larsson DJ, Andremont A, Bengtsson-Palme J, Brandt KK, de Roda Husman AM, Fagerstedt P (2018) Critical knowledge gaps and research needs related to the environmental dimensions of antibiotic resistance. Environ Int 117:132–138
Laxminarayan R, Duse A, Wattal C, Zaidi AK, Wertheim HF, Sumpradit N, Greko C (2013) Antibiotic resistance—the need for global solutions. Lancet Infect Dis 13(12):1057–1098
Li T, Liu T, Zheng C, Kang C, Yang Z, Yao X (2017) Changes in soil bacterial community structure as a result of incorporation of Brassica plants compared with continuous planting eggplant and chemical disinfection in greenhouses. PLoS ONE 12(3):e0173923
Marasco R, Rolli E, Ettoumi B, Vigani G, Mapelli F, Borin S, Abou-Hadid AF (2012) A drought resistance-promoting microbiome is selected by root system under desert farming. PLoS ONE 7(10):e48479
Mendes R, Garbeva P, Raaijmakers JM (2013) The rhizosphere microbiome: significance of plant beneficial, plant pathogenic, and human pathogenic microorganisms. FEMS Microbiol Rev 37(5):634–663
Mhlongo MI, Piater LA, Madala NE, Labuschagne N, Dubery IA (2018) The chemistry of plant–microbe interactions in the rhizosphere and the potential for metabolomics to reveal signaling related to defense priming and induced systemic resistance. Front Plant Sci 9:112
Morgan HH, Du Toit M, Setati ME (2017) The grapevine and wine microbiome: insights from high-throughput amplicon sequencing. Front Micro 11(8):820
Nair DN, Padmavathy S (2014) Impact of endophytic microorganisms on plants, environment and humans. Sci World J. https://doi.org/10.1155/2014/250693
Naylor D, Coleman-Derr D (2018) Drought stress and root-associated bacterial communities. Front Plant Sci 8:2223
Neal AL, Ahmad S, Gordon-Weeks R, Ton J (2012) Benzoxazinoids in root exudates of maize attract Pseudomonas putida to the rhizosphere. PLoS ONE 7(4):e35498
Olanrewaju OS, Glick BR, Babalola OO (2017) Mechanisms of action of plant growth promoting bacteria. World J Microbiol Biotechnol 33(11):197
Ortíz-Castro R, Contreras-Cornejo HA, Macías-Rodríguez L, López-Bucio J (2009) The role of microbial signals in plant growth and development. Plant Signal Behav 4(8):701–712
Paul C, Filippidou S, Jamil I, Kooli W, House GL, Estoppey A (2019) Bacterial spores, from ecology to biotechnology. Adv Appl Microbiol 106:79–111
Peršoh D (2015) Plant-associated fungal communities in the light of meta’omics. Fungal Divers 75(1):1–25
Pfeiffer S, Mitter B, Oswald A, Schloter-Hai B, Schloter M, Declerck S, Sessitsch A (2016) Rhizosphere microbiomes of potato cultivated in the High Andes show stable and dynamic core microbiomes with different responses to plant development. FEMS Microbiol Ecol 93(2):fiw242
Pineda A, Kaplan I, Bezemer TM (2017) Steering soil microbiomes to suppress aboveground insect pests. Trends Plant Sci 22(9):770–778
Poudel R, Jumpponen A, Schlatter DC, Paulitz TC, Gardener BM, Kinkel LL, Garrett KA (2016) Microbiome networks: a systems framework for identifying candidate microbial assemblages for disease management. Phytopathology 106(10):1083–1096
Purahong W, Orrù L, Donati I, Perpetuini G, Cellini A, Lamontanara A, (2018) Plant microbiome and its link to plant health: tree species, organs and Pseudomonas syringae pv. actinidiae infection shaping bacterial phyllosphere communities of kiwifruit plants. Front Plant Sci 9:1563
Qiao Q, Wang F, Zhang J, Chen Y, Zhang C, Liu G (2017) The variation in the rhizosphere microbiome of cotton with soil type, genotype and developmental stage. Sci Rep 7(1):3940
Raaijmakers JM, Paulitz TC, Steinberg C, Alabouvette C, Moënne-Loccoz Y (2009) The rhizosphere: a playground and battlefield for soilborne pathogens and beneficial microorganisms. Plant Soil 321(1–2):341–361
Rana KL, Kour D, Sheikh I, Dhiman A, Yadav N, Yadav AN, Rastegari AA, Singh K, Saxena AK (2019a) Endophytic fungi: biodiversity, ecological significance and potential industrial applications. In: Yadav AN, Mishra S, Singh S, Gupta A (eds) Recent advancement in white Biotechnology through fungi, vol 1. Diversity and Enzymes Perspectives. Springer, Switzerland, pp 1–62
Rana KL, Kour D, Sheikh I, Yadav N, Yadav AN, Kumar V, Singh BP, Dhaliwal HS, Saxena AK (2019b) Biodiversity of endophytic fungi from diverse niches and their biotechnological applications. In: Singh BP (ed) Advances in Endophytic Fungal Research: Present Status and Future Challenges. Springer International Publishing, Cham, pp 105–144. https://doi.org/10.1007/978-3-030-03589-1_6
Rascovan N, Carbonetto B, Perrig D, Díaz M, Canciani W, Abalo M, Alloati et al (2016) Integrated analysis of root microbiomes of soybean and wheat from agricultural fields. Sci Rep 6:28084
Rasheed A, Hao Y, Xia X, Khan A, Xu Y, Varshney RK, He Z (2017) Crop breeding chips and genotyping platforms: progress, challenges, and perspectives. Mol Plant 10(8):1047–1064
Rashid MI, Mujawar LH, Shahzad T, Almeelbi T, Ismail IM, Oves M (2016) Bacteria and fungi can contribute to nutrients bioavailability and aggregate formation in degraded soils. Microbiol Res 183:26–41
Rosenberg E, Zilber-Rosenberg I (2018) The hologenome concept of evolution after 10 years. Microbiome 6(1):78
Ryan RP, Germaine K, Franks A, Ryan DJ, Dowling DN (2008) Bacterial endophytes: recent developments and applications. FEMS Microbiol Lett 278(1):1–9
Santos ML, Berlitz DL, Wiest SL, Schünemann R, Knaak N, Fiuza LM (2018) Benefits associated with the interaction of endophytic bacteria and plants. Braz Arch Biol Technol 61:e18160431
Santoyo G, Moreno-Hagelsieb G, del Orozco-Mosqueda M C, Glick BR (2016) Plant growth-promoting bacterial endophytes. Microbiol Res 183:92–99
Savary S, Ficke A, Aubertot JN, Hollier C (2012) Crop losses due to diseases and their implications for global food production losses and food security Food Security 4(4):519–537
Schlaeppi K, Bulgarelli D (2015) The plant microbiome at work. Mol Plant-Microbe Interact 28(3):212–217
Schreiter S, Babin D, Smalla K, Grosch R (2018) Rhizosphere competence and biocontrol effect of Pseudomonas sp. RU47 independent from plant species and soil type at the field scale. Front Micro 9:97
Shade A, Jacques MA, Barret M (2017) Ecological patterns of seed microbiome diversity, transmission, and assembly. Curr Opin Microbiol 37:15–22
Shahzad R, Khan AL, Bilal S, Asaf S, Lee IJ (2018) What is there in seeds? Vertically transmitted endophytic resources for sustainable improvement in plant growth. Front Plant Sci 9:24
Sime-Ngando T (2014) Environmental bacteriophages: viruses of microbes in aquatic ecosystems. Front Micro 5:355
Souza RD, Ambrosini A, Passaglia LM (2015) Plant growth-promoting bacteria as inoculants in agricultural soils. Genet Mol Biol 38(4):401–419
Steven B, Huntley RB, Zeng Q (2018) The influence of flower anatomy and apple cultivar on the apple flower phytobiome. Phytobiomes 2(3):171–179
Streit WR, Schmitz RA (2004) Metagenomics–the key to the uncultured microbes. Curr Opin Microbiol 7(5):492–498
Suman A, Yadav AN, Verma P (2016) Endophytic Microbes in Crops: Diversity and Beneficial impact for Sustainable Agriculture. In: Singh D, Abhilash P, Prabha R (eds) Microbial inoculants in sustainable agricultural productivity, research perspectives. Springer-Verlag, India, pp 117–143. https://doi.org/10.1007/978-81-322-2647-5_7
Thijs S, Sillen W, Rineau F, Weyens N, Vangronsveld J (2016) Towards an enhanced understanding of plant–microbiome interactions to improve phytoremediation: engineering the metaorganism. Front Micro 16(7):341
Tshikantwa TS, Ullah MW, He F, Yang G (2018) Current trends and potential applications of microbial interactions for human welfare. Front Micro 9:1156
Verma P, Yadav AN, Khannam KS, Kumar S, Saxena AK, Suman A (2016) Molecular diversity and multifarious plant growth promoting attributes of Bacilli associated with wheat (Triticum aestivum L.) rhizosphere from six diverse agro-ecological zones of India. J Basic Microbiol 56:44–58
Verma P, Yadav AN, Khannam KS, Saxena AK, Suman A (2017a) Potassium-solubilizing microbes: diversity, distribution, and role in plant growth promotion. In: Panpatte DG, Jhala YK, Vyas RV, Shelat HN (eds) Microorganisms for green revolution: volume 1: microbes for sustainable crop production. Springer Singapore, Singapore, pp 125–149. https://doi.org/10.1007/978-981-10-6241-4_7
Verma P, Yadav AN, Kumar V, Singh DP, Saxena AK (2017b) Beneficial plant-microbes interactions: biodiversity of microbes from diverse extreme environments and its impact for crop improvement. In: Singh DP, Singh HB, Prabha R (eds) Plant-microbe interactions in agro-ecological perspectives: volume 2: microbial interactions and agro-ecological impacts. Springer Singapore, Singapore, pp 543–580. https://doi.org/10.1007/978-981-10-6593-4_22
Von Wintersdorff CJ, Penders J, van Niekerk JM, Mills ND, Majumder S, van Alphen LB (2016) Dissemination of antimicrobial resistance in microbial ecosystems through horizontal gene transfer. Front Micro 7:173
Vrancken K, Holtappels M, Schoofs H, Deckers T, Valcke R (2013) Pathogenicity and infection strategies of the fire blight pathogen Erwinia amylovora in Rosaceae: state of the art. Microbiology 159(5):823–832
Vurukonda SS, Vardharajula S, Shrivastava M, SkZ A (2016) Enhancement of drought stress tolerance in crops by plant growth promoting rhizobacteria. Microbiol Res 184:13–24
Wagner MR, Busby PE, Balint-Kurti P (2019) Breeding for broad-spectrum disease resistance alters the maize leaf microbiome. bioRxiv 1:647446
Warinner C, Herbig A, Mann A, Fellows Yates JA, Weiß CL, Burbano HA (2017) A robust framework for microbial archaeology. Annu Rev Genomics Hum Genet 31(18):321–356
Whipps JM (2001) Microbial interactions and biocontrol in the rhizosphere. J Exp Bot 1:487–511
Williamson KE, Fuhrmann JJ, Wommack KE, Radosevich M (2017) Viruses in soil ecosystems: an unknown quantity within an unexplored territory. Annu Rev Virol 4:201–219
Yadav AN (2017a) Agriculturally important microbiomes: biodiversity and multifarious PGP attributes for amelioration of diverse abiotic stresses in crops for sustainable agriculture. Biomed J Sci Tech Res 1:1–4
Yadav AN (2017b) Beneficial role of extremophilic microbes for plant health and soil fertility. J Agric Sci 1:1–4
Yadav AN (2019) Microbiomes of Wheat (Triticum aestivum L.) Endowed with Multifunctional Plant Growth Promoting Attributes. EC Microbiol 15:1–6
Yadav AN, Kour D, Sharma S, Sachan SG, Singh B, Chauhan VS, Sayyed RZ, Kaushik R, Saxena AK (2019) Psychrotrophic microbes: biodiversity, mechanisms of adaptation, and biotechnological implications in alleviation of cold stress in plants. In: Sayyed RZ, Arora NK, Reddy MS (eds) Plant growth promoting rhizobacteria for sustainable stress management: volume 1: rhizobacteria in abiotic stress management. Springer Singapore, Singapore, pp 219–253. https://doi.org/10.1007/978-981-13-6536-2_12
Yadav AN, Kumar V, Prasad R, Saxena AK, Dhaliwal HS (2018a) Microbiome in crops: diversity, distribution and potential role in crops improvements. In: Prasad R, Gill SS, Tuteja N (eds) Crop improvement through microbial biotechnology. Elsevier, USA, pp 305–332
Yadav AN, Sachan SG, Verma P, Saxena AK (2015a) Prospecting cold deserts of north western Himalayas for microbial diversity and plant growth promoting attributes. J Biosci Bioeng 119:683–693
Yadav AN, Sachan SG, Verma P, Saxena AK (2016) Bioprospecting of plant growth promoting psychrotrophic Bacilli from cold desert of north western Indian Himalayas. Indian J Exp Biol 54:142–150
Yadav AN, Sachan SG, Verma P, Tyagi SP, Kaushik R, Saxena AK (2015b) Culturable diversity and functional annotation of psychrotrophic bacteria from cold desert of Leh Ladakh (India). World J Microbiol Biotechnol 31:95–108
Yadav AN, Verma P, Kaushik R, Dhaliwal HS, Saxena AK (2017a) Archaea endowed with plant growth promoting attributes. EC Microbiol 8:294–298
Yadav AN, Verma P, Kour D, Rana KL, Kumar V, Singh B, Chauahan VS, Sugitha T, Saxena AK, Dhaliwal HS (2017b) Plant microbiomes and its beneficial multifunctional plant growth promoting attributes. Int J Environ Sci Nat Resour 3(1):1–8
Yadav AN, Verma P, Kumar M, Pal KK, Dey R, Gupta A, Padaria JC, Gujar GT, Kumar S, Suman A, Prasanna R, Saxena AK (2015c) Diversity and phylogenetic profiling of niche-specific Bacilli from extreme environments of India. Ann Microbiol 65:611–629
Yadav AN, Verma P, Kumar S, Kumar V, Kumar M, Singh BP, Saxena AK, Dhaliwal HS (2018b) Actinobacteria from Rhizosphere: molecular diversity, distributions and potential biotechnological applications. In: Singh B, Gupta V, Passari A (eds) New and future developments in microbial biotechnology and bioengineering. USA, pp 13–41. https://doi.org/10.1016/b978-0-444-63994-3.00002-3
Yadav AN, Verma P, Sachan SG, Kaushik R, Saxena AK (2018c) Psychrotrophic Microbiomes: Molecular Diversity and Beneficial Role in Plant Growth Promotion and Soil Health. In: Panpatte DG, Jhala YK, Shelat HN, Vyas RV (eds) Microorganisms for green revolution-volume 2: microbes for sustainable agro-ecosystem. Springer, Singapore, pp 197-240. https://doi.org/10.1007/978-981-10-7146-1_11
Yadav AN, Verma P, Singh B, Chauhan VS, Suman A, Saxena AK (2017c) Plant growth promoting bacteria: biodiversity and multifunctional attributes for sustainable agriculture. Adv Biotechnol Microbiol 5(5):1–6
Yadav AN, Yadav N (2018) Stress-adaptive microbes for plant growth promotion and alleviation of drought stress in plants. Acta Sci Agric 2:85–88
Yu P, Hochholdinger F (2018) The role of host genetic signatures on root-microbe interactions in the rhizosphere and endosphere. Front Plant Sci 9:1896
Zarraonaindia I, Owens SM, Weisenhorn P, West K, Hampton-Marcell J, Lax S (2015) The soil microbiome influences grapevine-associated microbiota. MBio 6(2):e02527–14
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2020 Springer Nature Switzerland AG
About this chapter

Cite this chapter
Singh, S., Kumar, V., Singh, S., Dhanjal, D.S., Datta, S., Singh, J. (2020). Global Scenario of Plant–Microbiome for Sustainable Agriculture: Current Advancements and Future Challenges. In: Yadav, A., Singh, J., Rastegari, A., Yadav, N. (eds) Plant Microbiomes for Sustainable Agriculture. Sustainable Development and Biodiversity, vol 25. Springer, Cham. https://doi.org/10.1007/978-3-030-38453-1_14
Download citation
DOI: https://doi.org/10.1007/978-3-030-38453-1_14
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-030-38452-4
Online ISBN: 978-3-030-38453-1
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)