
Summary
This chapter deals with some taxonomic and ecological aspects of picocyanobacteria (Pcy) single-cells, microcolonies and other colonial (CPcy), that are common in lakes throughout the world, and abundant across a wide spectrum of trophic conditions. We discussed phenotypic diversity of Pcy in conjunction with a genotypic approach in order to resolve whether a similar morphology also reflects a phylogenetic relationship. Microcolonies of different size (from 5 to 50 cells) constitute a gradient without a net separation from single-celled types and should be considered Pcy, as transition forms from single-cell to colonial morphotypes. The single-celled Pcy populations tend to be predominant in large, deep oligo-mesotrophic lakes, while the CPcy find optimal conditions in warmer, shallower and more nutrient rich lakes. The knowledge of Pcy diversity in pelagic and littoral zone habitats is a key to understand the dominance of certain genotypes in the water column and of their ubiquity. We devoted some paragraphs to analyse the factors (biotic and abiotic) which can influence the dynamics of the different Pcy forms and we have approached the study of their common ecology.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others
Keywords
- Cyanobacterial Bloom
- Oligotrophic Lake
- Dissolve Organic Phosphorus
- Deep Chlorophyll Maximum
- Synechococcus Strain
These keywords were added by machine and not by the authors. This process is experimental and the keywords may be updated as the learning algorithm improves.
1 Introduction
A phylogenomic study of the evolution of cyanobacterial traits shows that the earliest lineages were probably unicellular cells in terrestrial and/or freshwater environments (Sánchez-Baracaldo et al. 2005; Blank and Sánchez-Baracaldo 2010) rather than in the marine habitat as suggested by Honda et al. (1999). This discovery opens new prospects for the study of freshwater Pcy and provides an impetus for phylogenetic and ecological investigations to clarify the many uncertainties in the literature. One of the most striking differences between freshwater and marine Pcy lies in the extraordinary richness of morphotypes and the unresolved phylogeny of Pcy in lakes. However, despite marked phylogenetic differences, Pcy have a similar pattern in their absolute and relative importance in freshwater and marine systems along the trophic gradient (Bell and Kalff 2001).
This chapter discusses freshwater Pcy single-cells together with microcolonies and other colonial forms, that are common in lakes throughout the world (Hawley and Whitton 1991a) and abundant across a wide spectrum of trophic conditions. Though their abundance in meso-eutrophic lakes is often high enough to reduce transparency and cause water discolouration, they seldom create the blooms associated with larger colonial cyanobacteria. Although studies of the ecology of microcolonies and colonial forms are few, there have been sufficient studies within the past 25 year of Pcy in lakes and their role in food webs to warrant synthesis and further review (Stockner et al. 2000; Callieri 2008).
The picocyanobacteria exhibit two common morphologies: single cells (coccoid, rods) and colonies with diverse colonial morphology. We propose here to consider microcolonies as transitional forms from single cells to colonial morphotypes (Fig. 8.1). Under favourable conditions some Pcy can develop mucilage or a sheath and remain near to the mother cell forming a clump. Here we designate as picocyanobacteria (Pcy) the single cells (0.2–2.0 μm) which are the major component of the picophytoplankton community, and as colonial picocyanobacteria (CPcy) all species whose predominant morph is colonial and have single cells ranging from 0.5 to 3.0 μm. Microcolonies of different size (from 5 to 50 cells) constitute a gradient without a clear separation from the single-celled type and should be considered Pcy.

Single cells, microcolonies and larger colonies in Lake Maggiore, a large deep oligo-mesotrophic subalpine lake in northern Italy. PE-rich Pcy under epifluorescence microscopy (1250x) (blue-excitation)
Until the past few decades most research on Pcy focused on the CPcy, because their colonies were easily seen by conventional microscopy, and their ubiquity in meso-eutrophic lakes of Northern Europe attracted the attention of early descriptive taxonomists (Lemmermann 1904; Naumann 1924; Skuja 1948). CPcy also seem to have received more thorough systematic descriptions, along with some comment on habitat preference and distribution (Komárek 1958, 1996; Cronberg 1991; Komárková-Legnerová and Cronberg 1994).
The emergence of Pcy as an important research topic for limnologists and oceanographers provides an opportunity to discuss to what extent this large and diverse group share a common ecology. The current challenge is to understand better the relationship between the diversity and ecology of Pcy, microcolonies and CPcy and their interaction with the environmental factors that allow the proliferation of the most competitive genotypes. The study of genome divergence, lateral gene transfer and genomic islands will provide new opportunities for a better understanding of niche adaptation (Dufresne et al. 2008; Scanlan et al. 2009 and Chap. 20). We conclude this review with a plea for limnologists to pay more attention to these organisms. Their extent and abundance in lakes may provide an important message about the changes likely to occur in pelagic community structure in the warmer world expected in the near future (Mann 1993; Stockner 1998).
2 Taxonomy and Phylogenetic Diversity
2.1 Single Cells and Microcolonies
Even more than for most prokaryotes the morphological features of Pcy are insufficiently distinct to provide a reliable basis for discriminating taxa (Staley 1997; Komárek et al. 2004). The criteria for the definition of genera of single-celled Pcy such as Cyanobium, Synechococcus and Cyanothece dianae/cedrorum-type (Komárek 1996) have been supplanted by molecular methods which focus on clade divergence in the phylogenetic tree rather than on morphological differences. The clade containing Pcy (Synechococcus/Prochlorococcus/Cyanobium sensu Sánchez-Baracaldo et al. 2005) is formed by coccoid and rod-shaped cells with a diameter <3 μm. Analysis of 16S ribosomal DNA (rDNA) of freshwater Synechococcus shows it is a polyphyletic genus and cannot be considered a natural taxon (Urbach et al. 1998; Robertson et al. 2001). In the phylogenetic tree the Antarctic strains represent a unique and highly adapted clade related only peripherally to Synechococcus sp. (Cluster 5.2, Marine Cluster B) (Vincent et al. 2000; Powell et al. 2005).
It has become increasingly clear that phenotypic diversity should be evaluated in conjunction with genotypic analysis in order to resolve whether a similar morphology also reflects a phylogenetic relationship (Wilmotte and Golubić 1991). Even though many genetically distinct Synechococcus strains have been found (Robertson et al. 2001), it is still helpful to classify Pcy into two cell types: the first with yellow autofluorescing phycoerythrin (PE-rich cells), and the second with red autofluorescing phycocyanin (PC-rich cells) as the major light-harvesting pigment (Fig. 8.2) (Wood et al. 1985; Ernst 1991). Phycoerythrin-rich strains have an absorption peak at ∼560 nm, and hence absorb green light effectively. Phycocyanin-rich strains have an absorption peak at ∼625 nm, and absorb orange-red light effectively (Callieri et al. 1996a ; Haverkamp et al. 2008).

Three different views of Pcy: liquid cultures (green: PC-cells, pink: PE-cells); epifluorescence under blue excitation with PC cells (showing in red) and PE cells (showing in yellow); transmission electron microscope photo (showing the different internal structure of PC- and PE- cells)
Phylogenetic studies have mostly been performed using sequence data derived from 16S rDNA which is a conserved gene, but shows high pair-wise similarity in freshwater Pcy (Fig. 8.3, Crosbie et al. 2003a) and cannot resolve the actual genetic variation that accompanies their physiological diversity (Urbach et al. 1998). Less conserved genetic markers can offer a more detailed definition of the diversity of Pcy (Haverkamp et al. 2009). In particular the spacer between the 16S and 23S rDNA (ITS-1) exhibits a great deal of length and sequence variation and can be used to differentiate marine and freshwater Pcy ecotypes using fingerprinting techniques (T-RFLP, DGGE or ARISA) (Rocap et al. 2002; Becker et al. 2002; Ernst et al. 2003). In a glacial Andean lake system (Argentina) the ARISA showed habitat specificity of some Pcy OTU, emphasizing the microdiversity due to geographical barriers (Caravati et al. 2010). Similarly, the study of functional genes as, for example, those encoding for phycocyanin and phycoerythrin (cpcBA and cpeBA) can offer another perspective on the evolution of Pcy, grouping the strains on the basis of pigment composition (Haverkamp et al. 2008; Jasser et al. 2011). Indeed, it has been found that phylogenies based on phycobiliprotein rod gene components are not congruent with the 16S rRNA phylogeny whilst those based on the allophycocyanin core are congruent (Six et al. 2007; Haverkamp et al. 2009). This is explained by the presence of phycobilisome rod components within genomic islands that potentially allow their transfer between Synechococcus lineages (Six et al. 2007). A comparison of marine Synechococcus showed that their adaptation to different ecological niches can be related to the variable number of horizontally acquired genes located in highly variable genomic islands (Dufresne et al. 2008). Similar to what happens in Prochlorococcus (Coleman et al. 2006); it is likely that phages carrying host genes can mediate lateral gene transfer. This discovery opens new perspectives to the understanding of the local adaptation and the definition of species within the Synechococcus group.
The phylogenetic approach combined with RT-qPCR has been used to assess Pcy in both marine (Ahlgren et al. 2006) and freshwater community structure (Becker et al. 2000, 2007). Using small subunit (ssu) rDNA sequences from novel culture isolates together with environmental samples from the Baltic Sea and seven freshwater lakes, Sánchez-Baracaldo et al. (2008) showed that freshwater Pcy communities encompass much greater diversity than is found in marine systems. They hypothesised a more rapid speciation in lakes allowed by geographical barriers and noticed that most of the sequences from the Baltic Sea were related closely to freshwater lineages.

Maximum-likelihood phylogenetic tree of SSU rDNA sequences from unicellular picocyanobacteria, (green circle: PC-rich, pink circle: PE-rich) (For details see Crosbie et al. 2003a, reproduced with permission)
To provide a more realistic phylogenetic tree of cyanobacteria Sánchez-Baracaldo et al. (2005) used a combination of different molecular sequence data instead of individual genes. Flanking a selection of morphological traits into the backbone cyanobacterial tree they showed that the ancestral cyanobacterium were a single cell and that filamentous/colonial forms appeared later in time. The presence of a well-defined sheath, associated with the colonial lifestyle, is a trait which has been lost and attained several times during evolution. In Arctic lakes the Pcy strains isolated appear to be closely related to Microcystis elabens (Vincent et al. 2000) now reclassified as a species of Aphanothece (Komárek and Anagnostidis 1999). Thus, it is tempting to suggest that microcolonies, which are frequently found in freshwater, may be considered transition forms from single cells to true colonial. In this sense the investigations done by Crosbie et al. (2003a) confirm the existence of single cell/single colony strains, with different degrees of aggregation, possibly belonging to the group H and group B subalpine cluster I.
2.2 Colonies
Most of the small pico-cell-sized CPcy belong to the chroococcal cyanobacteria. The cell size ranges between 0.5 and 3 μm in diameter and the cell form is generally spherical, ovoid or rodlike. The cells occur in colonies of different morphology and these colonial morphologies are often species specific, and can be used in discriminating species. The cells inside the CPcy colony can be loosely or densely packed, or can form pseudo-filaments or other net-like structures. In some species the cells are attached to mucilaginous stalks, which are centred in the middle of the colony (e.g. Cyanonephron, Snowella). In lakes where CPcy are common, there is usually a mixture of species each with distinctive colony structure, which are quite readily identifiable. CPcy are found throughout the entire spectrum of lake trophic conditions; however most tend to occur in more productive meso-eutrophic lakes. Some of the most common CPcy in fresh waters are species belonging to the genera Aphanocapsa, Aphanothece, Chroococcus, Coelosphaerium, Cyanobion, Cyanodictium, Merismopedia, Romeria, Snowella and Tetracercus (Table 8.1).
There are few records of the development of CPcy in nature or in culture and accordingly the available literature is sparse. The CPcy are only a small component in water blooms and are thus often overlooked and usually neglected. Due to their minute cell size, unfiltered samples must be taken from lakes, as the CPcy will otherwise pass through the meshes of normal plankton nets (mesh width 10–45 μm) and are lost. The colonies of CPcy are normally counted in sedimentation chambers under 400–1,000× magnification. The settling time of CPcy must be long; for example in a 5 cm long chamber the settling time should be about 4 days and nights and the sample must be well preserved in Lugol’s solution.
Bláha and Marsálek (1999) isolated seven strains of Pcy from water blooms in the Czech Republic and tested their ability to produce toxins. They found that two strains produced the hepatotoxin microcystin. As most all Pcy can readily pass through the filters in drinking water treatment plants, they cannot be neglected and as possible sources of toxins should be carefully monitored.
Several studies of cyanobacteria within Brazil’s drinking water reservoirs have been made and provide an overview of their current status (Domingos et al. 1999; Komárek et al. 2001; Sant’Anna et al. 2004; Furtado et al. 2009). As most of the reservoirs were dominated by cyanobacteria, it was important to assess for cyanobacterial toxicity. Cyanobacteria, including CPcy and Pcy were isolated and tested for algal toxins with HPLC and/or ELISA methods. Several cyanobacteria including CPcy were found to be toxic; for example Aphanocapsa cumulus from Caruaru Reservoirs, where several persons have died after having dialysis with drinking water from these reservoirs (Domingos et al. 1999). In Tabocas reservoir CPcy were also very frequent and strains were isolated, e.g. Aphanocapsa cumulus, Aphanothece stratus, and a new species Romeria carauru was recorded (Komárek et al. 2001). No toxin tests were made on these species, thus it was not confirmed whether these species were responsible for the Carauro (Komárek et al. 2001), but they likely were. The morphology of strains of A. stratus and Cyanobium-like Pcy were studied with TEM, and their morphologies were identical and they evidently belonged to the same species. During the life-cycle A. stratus lived in colonies in the benthos, but as single cells or microcolonies in the plankton (Komárek et al. 2001). Pairwise similarities 16S rDNA sequence of A. stratus, R. carauru and other related cyanobacterial strains were carried out, but further studies are needed to evaluate genotype resemblance.
Furtado et al. (2009) studied cyanobacteria in waste stabilization pond systems in Brazil and recorded several CPcy. Morphological identification and phylogenetic analysis based on 16S rDNA sequence were made on the isolated strains and a phylogenetic tree with related species was obtained. They noted that the strains Synchococcus CENA 108 and Merismopedia CENA 206 were both capable of producing microcystins at large population densities (>1 × 106 cells mL−1, Furtado et al. 2009).
3 A Common Ecology?
3.1 Single Cell and Microcolony Dynamics
Sufficient information is now available to define the different patterns of Pcy, single-cells and microcolonies, in lakes of different size, thermal regime and trophic state (Callieri 2008, 2010). In lakes of temperate regions maxima generally conform to a typical bimodal pattern, with a spring or early summer peak and a second peak during summer-autumn (Stockner et al. 2000). This is the case of most of the subalpine large lakes without ice-cover e.g. Lake Maggiore, Lake Constance, but also for Lake Stechlin, a deep oligo-mesotrophic lake in the Baltic Lake District where ice-cover occurs (Padisák et al. 2003). However looking at the long-term series of Pcy abundance in Lake Maggiore (Fig. 8.4) (Callieri and Piscia 2002), Lake Constance (Gaedke and Weisse 1998), and Lake Stechlin (Padisák et al. 2003) not all the years are clearly bimodal. The great interannual variability of Pcy dynamics is mainly related to differences in weather conditions which cause different spring mixing regimes and timing of water column stabilization (Weisse 1993). Studies in British Columbia’s temperate oligotrophic lakes have shown a clear trend in both magnitude and timing of Pcy seasonal maxima related to levels and duration of seasonal nutrient supplementation (Stockner and Shortreed 1988, 1994).

Long-term dynamics of Pcy abundance in Lake Maggiore, Northern Italy
Large spring peaks are also common in eutrophic, hypereutrophic and dystrophic shallow lakes (Sime-Ngando 1995; Jasser 1997; Mózes et al. 2006). The seasonal patterns found in Danish lakes (Søndergaard 1991), Canadian lakes (Pick and Agbeti 1991), Lake Biwa, Japan (Maeda et al. 1992), English lakes (Hawley and Whitton 1991b; Sánchez-Baracaldo et al. 2008), Lake Mondsee, Austria (Crosbie et al. 2003c), Lakes Bourget and Geneva, France (Personnic et al. 2009a) all lack the spring peak, there being only a summer or autumn maximum. The lack of Pcy spring peak in these was probably due to weak stratification in March-April and hence relatively deep vertical mixing. This interpretation is strengthened by studies on Lake Baikal, where owing to winter ice cover and extended spring isothermal conditions, Pcy reach high abundance only in summer months and lack a spring peak (Belykh et al. 2006).
In Arctic and Antarctic lakes Pcy are widely distributed, despite the fact they are generally present in low abundance in the marine polar environment (Vincent 2000; Chap. 13). In continental Antarctica in meromictic saline Ace Lake Pcy reached concentrations one order of magnitude higher than in temperate lakes in summer – 8 × 106 cells mL−1 (Vincent 2000). In the Antarctic Peninsula in Lake Boeckella (Izaguirre et al. 2003) Pcy abundance was as high as 3.6 × 105 cells mL−1 and represented up to 80% of phytoplankton biomass (Allende and Izaguirre 2003). However very low Pcy concentrations (102–103 cells mL−1) were recorded in a set of shallow lakes and ponds in the Byers peninsula of maritime Antarctica (Toro et al. 2007). In contrast to all these, tropical lakes show high cell numbers (105–106 cell mL−1) throughout the season with higher early-spring peaks (Peštová et al. 2008) or summer peaks (Malinsky-Rushansky et al. 1995), depending upon the interactions of Pcy with other phytoplankton.
In Lake Maggiore the pronounced late summer peak of Pcy is composed by different morphotypes, including microcolonies (Passoni and Callieri 2000) (Fig. 8.5). Microcolonies are generally present throughout the euphotic zone, albeit in low abundance in all oligotrophic lakes, e.g. representing around 25% of the single-cell forms in Lake Maggiore (Passoni and Callieri 2000). The peak abundance of Pcy microcolonies appears in summer or autumn in a variety of freshwater systems (Fahnenstiel and Carrick 1992; Klut and Stockner 1991; Komárková 2002; Szelag-Wasielewska 2003; Crosbie et al. 2003c; Mózes et al. 2006; Ivanikova et al. 2007). Such a variety of morphotypes reflects a genotypic diversity among Pcy communities that accounts for the different Pcy patterns of morphotype composition observed in spring and summer assemblages (Callieri et al. 2007; Callieri et al. 2012: Fig. 8.6). Using RT-qPCR, a type of rapid succession of individual clades of Pcy illustrates the patchy structure of the Pcy community over quite small spatial/temporal scales (Sánchez-Baracaldo et al. 2008). At the same time the co-existence of genetically and physiologically diverse Synechococcus spp. found in the pelagic zone of Lake Constance indicates the possible niche partitioning exploited by the different strains (Postius and Ernst 1999; Ernst et al. 2003). In Lake Maggiore the vertical partitioning of Pcy OTUs down the water column was more evident during summer stratification (Callieri et al. 2012). In marine systems distinct Pcy lineages have also been shown to partition between waters having different environmental characteristics; a feature evident over large spatial scales (Fuller et al. 2006; Zwirglmaier et al. 2007, 2008). In the Sargasso Sea the community composition of Pcy varied during the season with the highest numbers of Synechococcus in spring and Prochlorococcus in summer and autumn (DuRand et al. 2001). We suggest that the new perspective of habitat-related distribution pattern of Synechococcus proposed for Lake Constance (Becker et al. 2007) and Lake Maggiore (Callieri et al. 2012) could be generalized to other aquatic systems as well.

Seasonal changes of single-cell picocyanobacteria, microcolonies and colonial coccoid cyanobacteria (mainly Aphanothece clathrata, A. cf. floccosa and Microcystis sp.) abundance in Lake Maggiore (from Passoni and Callieri 2000, modified)

Changes in relative percentage of the Operational Taxonomic Units (OTU) of Pcy in Lake Maggiore at 3 m (upper panel) and 20 m (lower panel) obtained with ARISA on the ITS-1 (Callieri et al. 2012)
Moreover, there is strong evidence that Pcy of the cyanobacterial evolutionary lineage VI sensu Honda et al. (1999) are not exclusively pelagic, but can also inhabit periphytic biofilm in the euphotic zone of temperate lakes (Becker et al. 2004). We should integrate our knowledge of Pcy diversity in pelagic and littoral zone habitats to better explain the dominance of certain genotypes in the water column, because the adaptability of these microorganisms may well be the key feature that accounts for their ubiquity (Becker et al. 2004).
Studies of the vertical distribution of populations of Pcy have provided important information about their response to changing physical and biological variables within the euphotic zone. Though Pcy cells are small and their settling rate negligible, their abundance and distribution within the water column can change rapidly with different thermal and light regimes, and to the presence of predators or viruses (Pernthaler et al. 1996; Mühling et al. 2005). Water column depth, which is often inversely related to the trophic state of the lake, is an important indicator of the presence of Pcy and/or of its abundance relative to larger species of phytoplankton. Deep, clear oligotrophic lakes typically support Pcy comprising mainly PE-rich cells; conversely in shallow, turbid lakes PC-rich cells prevail (Callieri and Stockner 2002). This disparity in the distribution of PE- and PC-rich cells is due to their characteristic spectral signature (Everroad and Wood 2006), which has been associated with particular underwater light quality (Hauschild et al. 1991; Vörös et al. 1998). In North Patagonian, in which blue light dominate the underwater light climate, Pérez et al. (2002) found that PE-rich cells typically dominate the Pcy forming a deep chlorophyll maxima (DCM) at the base of the euphotic zone (Callieri et al. 2007; Fig. 8.7). In Lake Baikal at offshore stations the Pcy are mainly PE-rich cells, whereas PC-rich cells are found at near-shore stations (Katano et al. 2005, 2008), suggesting the occurrence of water quality differences in various zones of the lake. Similar situations have been described for Lake Balaton where in the eastern basin PE-rich cells dominate, while in the western basin PC-rich cells are dominant (Mózes et al. 2006). It is the establishment of a pronounced thermocline at depth which likely favours the development of DCM, largely made up of Pcy which are suited both to low nutrient and light conditions (Modenutti and Balseiro 2002; Gervais et al. 1997; Camacho et al. 2003; Callieri et al. 2007). In Lake Tahoe Pcy dominated in the nutrient deficient upper water column during the stratified season, in a distinct vertical niche with respect to picoeukaryotes which peaked at the DCM (Winder 2009), similar to patterns found in the Oceans (Vázquez-Domínguez et al. 2008). The dynamics of DCM formed by Pcy is quite unstable and its duration is unpredictable, depending upon the strength of hydrodynamic field and biotic interactions. A good example is provided by Pcy communities in Lake Maggiore where DCM can appear, and also suddenly disappear, in just a few days (Fig. 8.8). Further, it has been shown that the interaction of different biotic and abiotic factors within the water column can affect Pcy vertical distribution patterns, with peaks of abundance in the lower metalimnion and upper hypolimnion of Lakes Huron and Michigan (Fahnenstiel and Carrick 1992); in Lake Stechlin (Padisák et al. 1998, 2003); in the metalimnion, beneath the steepest part of the thermocline, in Lake Constance and Lake Maggiore (Weisse and Schweizer 1991; Callieri and Pinolini 1995); in the metalimnion of Lake Baikal (Nagata et al. 1994); and in the epilimnion of Lake Biwa (Maeda et al. 1992), Lake Kinneret (Malinsky-Rushansky et al. 1995) and Lake Alchichica, Mexico (Peštová et al. 2008).

Vertical distribution of Pcy cells in the water column of six ultra-oligotrophic lakes of northern Patagonia, Argentina (Callieri et al. 2007, modified) PAR: Photosynthetically Active Radiation

Six vertical profiles of Pcy abundance, temperature and PAR in Lake Maggiore, during 30 days of summer stratification (C. Callieri and L. Oboti unpublished data). PAR shown in blue and temperature in red line; Pcy abundance shown with histograms: the black ones indicate the Pcy abundance at the depth of thermocline and the yellow ones the abundance at the other depths
3.2 Colony Dynamics
Although colonial picocyanobacteria (CPcy) are common in meso- to eutrophic lakes, there are few publications about their life-history and ecology. CPcy are “metaphyton” that often appear in the euphotic zone as plankton, but their origins are associated with the littoral and deeper benthos communities. They occur in most cyanobacterial blooms together with larger and more common cyanobacteria like Microcystis, Aphanizomenon and Anabaena. In most cases the CPcy only constitute a small percentage of the total colonial cyanobacteria biomass of the plankton; nevertheless CPcy species alone can also form heavy blooms (Cronberg 1999).
In Sweden monitoring programs for lake water quality started in the 1970s and in some cases continues today with a focus on phytoplankton and water chemistry. A classic case is Lake Trummen, in central south Sweden that was very eutrophic, but was restored to a lower production status in 1970 by dredging and removal of 0.5 m nutrient-rich sediment (Björk et al. 1972). After dredging lake water quality consistently improved and the cyanobacterial blooms diminished both in size and duration. In spring 1975 Lake Trummen was invaded by a large number of small bream and roach from nearby Lake Växjösjön and in consequence the predation of fish on zooplankton was extremely heavy. All cladocerans disappeared and a dense water bloom of the CPcy Cyanodictyon imperfectum (now called Cyanocatena imperfectum, Fig. 8.9), appeared reaching peak densities in September that constituted 95% of the total cyanobacterial bloom. The summer of 1975 was very warm and dry creating optimal conditions that likely contributed to the heavy CPcy bloom (Cronberg and Weibull 1981), some photos of which are shown in Fig. 8.10.

Cyanobacterial bloom of CPcy in Lake Trummen in 1975 caused by arrival of small bream, roach and perch. The fish had reduced all cladocerans, leading to grazing by zooplankton being very reduced. Cyanodictyon imperfectum (= Cyanocatena imperfecta) dominated the bloom

Examples of some colonial forms of Pcy; (a–b) Cyanodictyon imperfectum (= Cyanocatena imperfecta), (c) Aphanothece paralleliformis, (d–f) Cyanodictyon balticum (All micrographs to the same scale: bar = 10 μm)
Lake Ringsjön, situated in central Scania, Sweden, consists of three basins and is one of the largest lakes in the region. The three basins have been monitored since 1982 and CPcy frequently occur in varying population densities (Fig. 8.11). In Lake Ringsjön dense algal blooms have been observed during the summer, and this lake, like Lake Trummen, also had a large population of small bream, roach and perch. During 2005–2009 Lake Ringsjön western basin was biomanipulated and small cyprinid fish were removed, which resulted in a slight reduction of phytoplankton biomass (Bergman et al. 1999). However, the CPcy increased in the beginning of the period suggesting that fish reduction in some way affected CPcy production. The cyanobacterial community was dominated by species of the genera Microcystis, Aphanizomenon, Anabaena and Woronichinia. CPcy consisted mostly of Aphanocapsa delicatissima, Aphanothece clathrata, A. minutissima and Cyanodictyon imperfectum (Cronberg 1999).

Comparison of changes in the biomass of CPcy with those of ‘other cyanobacteria’ and ‘other algae’ in the phytoplankton of Lake Ringsjön western basin during 1998-2008. The lake was biomanipulated during 2005-2008 with 254 t small cyprinid fish being removed
In the late 1990s park ponds in the city of Malmö, southern Sweden, were studied owing to outbreaks of heavy cyanobacterial blooms that sometimes caused large bird kills. These cyanobacterial blooms consisted of species of Microcystis, Aphanizomenon, Anabaena, Anabaenopsis, and sometimes also of CPcy in high densities. In 1995 a toxic bloom of Cyanobion bacillare appeared in the Large Slottspark Pond in Malmö and during July and August about 650 water fowl, mainly mallard ducks, died (Fig. 8.12). In late August water was analyzed for algal toxins and showed high concentrations of microcystin, about 86 μg L−1, and biopsies on several dead birds showed severe damage to the liver and kidney; injuries typical of the toxin microcystin (Cronberg, unpublished data). A few years later, in 2000, the Middle Öresund Pond in Malmö exhibited an algal bloom of another CPcy species, which was a “new” CPcy species to phycological science – Cyanodictyon balticum (Cronberg 2003). C. balticum appeared from June to October with maximal development in September (Fig. 8.13). During the bloom period, the zooplankton population was also monitored, and in August, when the zooplankton declined, the CPcy increased. As long as zooplankton were present C. balticum biomass was kept low, but increased substantially when zooplankton disappeared. Among zooplankters, the rotifers in particular seemed to feed preferentially on the CPcy.

Development of toxic Cyanobion bacillare in Large Slottspark Pond during the summer 1995. The black arrow indicates the periods in which microcystin was found in the lake. During the period July to September 650 water fowl died

Cyanobacterial bloom of CPcy (Cyanodictyon balticum and other cyanobacteria) (a), and development of zooplankton (b), in the Middle Öresund Pond during 2000
C. balticum is a frequent phytoplankter in the Baltic Sea where several different CPcy species are common, and they are also common in lakes in the surrounding countries. They appear in shallow meso-, eutrophic- to hypertrophic lakes and ponds where they live as part of the metaphyton on the lake bottom; but as a result of wind and convective mixing they often become distributed in the pelagic zone and become part of the plankton community (Komárek et al. 2001). Meterological and hydrological conditions doubtless play an important role in the distribution of CPcy. The composition of the surrounding soil and ground water; the presence of anaerobic littoral sediments and attendant nutrient release; even at a slightly elevated salinity, each can either alone or collectively influence the physico-chemical conditions in littoral-benthic water, and create more optimal conditions for CPcy development.
As CPcy are sometimes major components of the pelagic phytoplankton community they are influenced not only by the available nutrients, but also by other planktonic organisms. The grazing by zooplankton and by heterotrophic flagellates may reduce the number of CPcy, and the presence of small fish may also influence the CPcy populations. Furthermore, some CPcy have the ability to produce toxins, so if CPcy colonies are disrupted during the treatment process for drinking water, potentially they can affect human health.
3.3 Single Cells Versus Microcolonies
There is a growing interest in the relationship between the status of Pcy microcolonies in a lake and the trophic state of that lake (Callieri 2010). It has been suggested that the abundance of microcolonies in temperate lakes in mid-summer, when nutrients are likely to be depleted may reflect more efficient nutrient recycling, with the colonies providing a self-sustaining microcosm that offers a competitive advantage over single cells (Klut and Stockner 1991). However, Crosbie et al. (2003c) considered this hypothesis unlikely for Pcy (Crosbie et al. 2003c) in view of results for colony-forming marine alga Phaeocystis, where the boundary layer can strongly limit nutrient diffusion into the colonies (Ploug et al. 1999). At low phosphorus concentrations the colonial forms grow slower than the single-cell forms (Veldhuis and Admiral 1987), presumably due to the lower cell-specific nutrient fluxes in colonies (Ploug et al. 1999). But in microcolonies, formed by 5–10 cells in one plane, the duplication should not be depressed as much as in a large, thick colony, where the diffusion of nutrients is impeded. In this case, exudates adsorbed to the cell surface can act as rich metabolite pools (Klut and Stockner 1991). Therefore, during the initial stage of their formation a microcolony can have a selective advantage in nutrient depleted ecosystems.
Crosbie et al. (2003c) observed an increase of microcolonies in nutrient-poor surface waters in Lake Mondsee and attributed their formation to the production of photosynthate-rich mucilage in Pcy single-cells that were actively photosynthesising organic carbon. As the leakage or excretion of photosynthate has been considered one protection mechanism against photochemical damage (Wood and van Valen 1990), it is likely to also consider the effect of irradiance (PAR and UVR), at near-surface depths, as a stressor promoting clumping of daughter cells during duplication (Fig. 8.14).

Suggested dynamics of microcolony formation, indicating how high irradiance and nutrient deficiency could promote excretion of carbon-rich photosynthate that may in turn promote clump development due to production of daughter cells
To better understand genus-specific microcolony formation one must consider the factors influencing cell aggregation, despite the many differences between microcolonies and aggregates. The results by Koblížek et al. (2000) suggest that in culture Synechococcus elongatus aggregates rapidly if exposed to blue light (30 min, 1,000 μmol photon m−2 s−1) owing to the effect of electron transfer downstream of PSI, with reactive oxygen radicals (ROS) probably triggering the aggregation. PSI therefore may have an important role in the first stages of microcolony formation in lakes. The influence of moderate UVR on microcolony formation was tested on PE- and PC-rich Synechococcus cultures (Callieri et al. 2011). Previous acclimation to low or moderate PAR influences the strain response, mainly due to carotenoid content in the cell. In general microcolony formation appears as a defence strategy of the low acclimated culture (Callieri et al. 2011).
As well as cell metabolism alterations caused by external factors such as light, other important structural changes of Pcy single-cells must be mentioned as a causative factor inducing microcolony formation. Aggregation is an ATP-independent process without any de novo protein synthesis (Koblížek et al. 2000), and this indicates that some structures responsible for the aggregation must be present on the cell surface before irradiation. For example, in selected strains with different genotypes isolated from Lake Constance; Ernst et al. (1996) found that they possess a surface S-layer composed of regularly ordered globular protein layers that would facilitate colony formation. Also, in grazing (by Ochromonas sp.) induced microcolonies of PC-rich Cyanobium sp., rigid tubes from 100 nm to 1 μm long (spinae) have been observed on the cell surface (Jezberová and Komárková 2007). To what extent the formation of microcolonies is due to the presence of specific Synechococcus or Cyanobium genotypes or is the result of a specific survival strategy (Ernst et al. 1999; Passoni and Callieri 2000) needs further clarification. Unicellular Microcystis aeruginosa Kützing, when exposed to Ochromonas sp. grazing, increased the diameter of the colonies and the extracellular polysaccharides (EPS) content (Yang and Kong 2012).
A fascinating hypothesis on microcolony formation is related to the observation by Postius and Böger (1998) that exo-polysaccharides exudated by Pcy as microzones for diazotrophic bacteria growth may affect microcolony formation. This finding opens new perspectives for the study of consortial, synergistic interactions that may be of critical importance to our understanding of colony formation in Pcy.
3.4 Growth Rate and Occurrence Along the Trophic Gradient
The processes of cell growth and division in all the photosynthetic organisms are as tightly coupled as photosynthesis and growth rate, and are light dependent (Kana and Glibert 1987a, b; Chisholm et al. 1986). In Pcy there is little difference between marine and freshwater strains of Synechococcus in both cell division and growth, with cell division reaching a maximum in the afternoon, triggering an increase in cell number that proceeds in the dark (Chisholm et al. 1986; Callieri et al. 1996 b; Jacquet et al. 2001). These light/dark cycles produce rhythmic cell divisions that are related to growth rate and photosynthetic activity and are driven by prevailing light conditions. Experimental laboratory evidence of the influence of light on growth rate is well documented by Fahnenstiel et al. (1991) for freshwater Synechococcus strains and by Campbell and Carpenter (1986a) for marine strains. These investigators have measured growth rates at light intensities up to 75 and 120 μmol photon m−2 s−1 that simulate natural irradiance levels. Kana and Glibert (1987b) have extended the light intensity limit up to 2,000 μmol m−2 s−1, demonstrating that growth rate becomes light saturated at 200 μmol m−2 s−1; however Synechococcus has a mechanism of photo-adaptation which permits cell growth and photosynthetic activity to continue at higher irradiances. Moser et al. (2009) have shown that growth rates of three Pcy strains of different pigment composition (PE-rich and PC-rich strains) and phylogenetic positions differed widely in response to light intensity and photo-acclimation (Fig. 8.15). Their results show that freshwater Pcy possess the ability of photo-adaptation, but the extent of photo-adaptation depends on the duration of photo-acclimation, and is strain-specific.

Growth rates of picocyanobacterial strains at 9 different irradiances after 2 months acclimation to 10 μmol photon m-2 s-1 (left) and 100 μmol photon m-2 s-1 (right). MW4C3 = PE-strain Group B; MW100C3 = PC-strain Group I; BO8801 = PC-strain Group A Cyanobium. The growth response at high light of the two PC-strains acclimated at 100 µmol photon m-2 s-1 shows the importance of ribotype cluster membership (From Moser et al. 2009)
Other environmental conditions such as nutrient concentration and temperature can affect cell specific growth rates. Growth rates of pico-phytoplankton in lakes along a trophic gradient ranged from 0.10 to 2.14 day−1 (Weisse 1993). In the oceans the in situ growth rate of Synechococcus was likely 0.7 day−1 (1 doublings/day) (Furnas and Crosbie 1999). Estimates of Pcy growth rates from Lakes Biwa and Baikal were 0.65 and 0.4 day−1, respectively (Nagata et al. 1994, 1996) and from Lake Kinneret ranged from 0.29 to 0.60 day−1 (Malinsky-Rushansky et al. 1995) all fall within published ranges. The maximum net growth rate of unicellular cyanobacteria in oligotrophic Lake Stechlin was 0.23 day−1 (Padisák et al. 1997) while in Lake Balaton it was 2.27 day−1 (Mastala et al. 1996). Based on studies of 48 lakes in Quebec, Ontario and New York State, Lavallée and Pick (2002) obtained a maximum growth rate of 1.93 day−1. Pcy growth in Lake Maggiore lies between 0.28 and 1.14 day−1 as net growth rate and 0.91–2.36 day−1 as potential growth rate measured using the frequency of dividing cells (FDC) method (Callieri et al. 1996 b).
One of the selective advantages of small cell size in low nutrient environments is that cells are less limited by molecular diffusion of nutrients because of their increase in surface area-to-volume ratio (Raven 1986; Chisholm 1992). The prokaryotic structure of the Pcy cell also gives them the lowest costs for maintenance metabolism, and this has been cited as the primary reason for their success in oligotrophic conditions (Weisse 1993).
The model outlined by Stockner (1991b) of an increase of picophytoplankton abundance and biomass and decrease of its relative importance with the increase of phosphorus concentration in lakes has been widely accepted and confirmed in marine and freshwater systems (Bell and Kalff 2001). Using the extensive freshwater database of Vörös et al. (1998), Callieri and Stockner (2002), and successively Callieri et al. (2007) enriched the dataset at the ultra-oligotrophic extreme, and confirmed a positive correlation between the numbers of Pcy and extant trophic conditions in a wide range of waterbodies. Moreover, the percent contribution of Pcy to the total phytoplankton biomass and production decreased with increasing lake trophic state.
Figure 8.16 provides a comparison of the results of Callieri et al. (2007) with those of Bell and Kalff (2001). The data for Pcy from ultra-oligotrophic pristine lakes in northern Patagonia (Argentina) cluster in the same position as those from marine systems, suggesting a common ecological response in the various environments, despite the phylogenetic differences of the organisms.

4 Factors Affecting Community Dynamics and Composition
4.1 Biotic Versus Abiotic Regulation
Debates about the relative importance of biotic regulation versus abiotic forcing in driving population fluctuations have been recurring over decades in all fields of ecology, and microbial ecology is no exception. In a study on different terrestrial and aquatic communities Houlahan et al. (2007) found that abiotic factors provide the primary forcing that drives temporal variability in species abundance. We know that the complex changeability of community structure is related to a spectrum of environmental variability through the interplay of intrinsic (basin morphometry, thermal stratification, wind mixing) and external factors (e.g. supply of nutrients) (Harris 1980). The exploitation of environmental variability by the Pcy community is the result of evolutionary mechanistic adaptation and the interrelation with other primary producers of larger size and with predators. Adaptation to a changing environment, with phasing of fluctuating events, can subject the community to dominance by the fittest and most adaptive available species. These concepts should be evaluated on the light of the new conceptual framework of community ecology, the meta-community, which considers the communities as shaped at different spatial scales (local and regional) (Wilson 1992; Leibold et al. 2004). Therefore in the debate on biotic versus abiotic regulation of community structure and dynamics we need to consider that local communities are not isolated but are linked by dispersal of multiple, potentially interactive, species (Logue and Lindström 2008).
4.2 Lake Morphometry and Thermal Regime
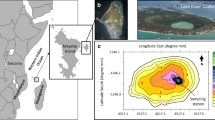
In order to interpret Pcy dynamics in freshwaters it is imperative to take into consideration the morphometric characteristics and thermal regime of a lake. The community composition of the Pcy can strongly depend on lake typology and morphogenesis. In a survey covering 45 lakes and ponds, Camacho et al. (2003) found that picocyanobacterial development was favoured by the stability of the vertical structure of the lake; that is by the inertial resistance to complete mixing owing to vertical density differences and to a long hydrological retention time. In lakes with relatively high water inflow and short retention time, Pcy are scarce. Far from this situation are deep lakes with a complex basin morphometry such as large sub-alpine lakes. In one of these lakes (Lake Maggiore) the Pcy population densities during summer stratification are high but with a pronounced North–south gradient due to a high retention time and peculiar characteristics along the lake axis (Fig. 8.17) (Bertoni et al. 2004). Lake Constance, with a different basin morphometry, has a less pronounced Pcy gradient (Weisse and Kenter 1991).

Map of the spatial heterogeneity of picocyanobacteria (Pcy) abundance in Lake Maggiore during spring. The northern basin of the lake contributes less total nitrogen and total phosphorus than the basin for the rest of the lake (Bertoni et al. 2004, modified)
Pcy composition and abundance vary conspicuously among shallow lakes, with a strong dependence on trophic condition (Stockner 1991a; Søndergaard 1991), altitude (Straškrabová et al. 1999b and cited references), oxidization-reduction conditions (Camacho et al. 2003) and the presence of dissolved humics (Drakare et al. 2003). It is therefore difficult to predict Pcy abundance in shallow lakes without considering the physical and chemical characteristics of the water. In a study of shallow humic lakes of the Boreal Forest Zone, Jasser and Arvola (2003) found Pcy to be light and temperature limited, whereas in humic Swedish lakes dissolved organic carbon (DOC) concentration was the factor most influencing Pcy composition (Drakare et al. 2003).
Lake thermal structure influences Pcy abundance and dynamics of Pcy due to both the effect of temperature per se and mass movements of the water in response to density gradients. In general a temperature increase enhances the potential growth rate of phytoplankton, increasing the reaction rate of RUBISCO (Beardall and Raven 2004). Marine Synechococcus reacts promptly to the temperature increase in laboratory experiments (Fu et al. 2007), and in a 5-year study on Lake Balaton Pcy abundance was positively correlated to water temperature (Vörös et al. 2009). Nevertheless the influence of temperature on the abundance of Pcy in lakes is difficult to separate from the influence of seasonal and geographical location. The widespread assumption that temperature is the driving force for growth and development of microorganisms does not apply so clearly for Pcy in nature. Among diverse marine habitats Li (1998) found a direct relationship between Pcy mean annual abundance and temperatures below 14°C; above 14°C nitrate concentrations were very low and may therefore replace temperature as the most significant. At higher temperatures other factors can also become dominant and control Pcy growth. Weisse (1993) suggested the importance of temperature as triggering the onset of Pcy growth in marine and freshwaters, but not for regulating their population dynamics. In Lake Maggiore the maximum Pcy concentration occurs near the thermocline and at temperatures between 18°C and 20°C (Callieri and Piscia 2002) (Fig. 8.18). This provides an example of where temperature not only has a direct effect on the cell, but also helps to maintain a density gradient which hinders sedimentation (Callieri 2008). Vertical gradients of water density have a profound effect on the distribution and diversity of Pcy in the metalimnion, upper hypolimnion and mixolimnion.

Isotherm map of Lake Maggiore (Northern Italy), 0–50 m layer during 1998. The crosses indicate the highest values of picocyanobacteria abundance. Depths with 10 % of surface solar radiation are also given (thick line). Vertical bars indicate thermocline extension (From Callieri 2008)
4.3 Nutrients, Light Limitation and pH
The main difference between marine and freshwater Pcy is in the differential regulation of primary production by nitrogen, iron and phosphorus. In lakes, primarily phosphorus has been regarded as the limiting nutrient (Schindler 1977, 2006), whereas in the oligotrophic oceans nitrogen and iron are considered the ultimate nutrients limiting primary production (Tyrrell 1999). Nevertheless, in the Mediterranean Sea and in the North Pacific subtropical gyre, a climate-related shift from an N- to P-limited ecosystem over the past several decades has been observed (Moore et al. 2005), due to increased nitrogen fixation (Karl et al. 1997). Conversely, in ultra-oligotrophic lakes nitrogen deficiency, even more than phosphorus, can be the cause of the low productivity (Stockner et al. 2005; Diaz et al. 2007). Furthermore nutrient co-limitation can occur in oligotrophic systems (Mills et al. 2004), where more than one nutrient may effectively co-limit biomass production (Mackey et al. 2009). Thus, past assumptions about whether the N or P is the proximate or ultimate nutrient limiting the productivity of phytoplankton populations in marine and freshwater systems are re-opened to debate.
As regard Pcy we may infer from the Stockner model that as lakes or oceans become more nutrient depleted, i.e. oligotrophic, then the greater the importance and relative contribution of Pcy to total autotrophic biomass (Bell and Kalff 2001). The success of Synechococcus spp. in oligotrophic systems can also be explained by their high affinity for orthophosphate (Moutin et al. 2002) and their maximum cell specific P-uptake rates that are competitively superior to algae and other bacteria under a pulsed supply (Vadstein 2000). Actually it has been demonstrated that growth rates of marine Pcy, under limitation by NH +4 , PO 3−4 , Fe or light, are seldom completely stopped; moreover, cell quotas are low as can be expected for such small cells (Timmermans et al. 2005). Iron’s limited bioavailability makes it limiting despite its abundance. The siderophores are iron-chelating compounds produced by cyanobacteria (Murphy et al. 1976; Hopkinson and Morel 2009). Siderophore production can provide a competitive advantage to cyanobacteria over other algae during iron stress, and can alter the bioavailabity of iron to the aquatic community (Wilhelm 1995). Nevertheless it has been found that in diluted aqueous environment endogenous siderophore uptake is inefficient. In this type of environment, reductive Fe uptake is an effective strategy in the acquisition of organically bound iron (Kranzler et al. 2011). In laboratory studies Prochlorococcus and Synechococcus marine strains, under P-limited conditions, showed high C:P and N:P ratios thus producing new biomass with a bioelemental stoichiometry well in excess of the canonical Redfield ratio (Bertilsson et al. 2003; Heldal et al. 2003). These results allow us to envisage the potentially great importance of Pcy for enhancing carbon sequestration with the ensuing potential to change the structure and complexity of pelagic food webs (Bertilsson et al. 2003).
An alternative explanation for the relative success of Pcy to grow at low inorganic P concentrations is given by the ability of their cells to utilise, in addition to PO 3−4 , organic sources of phosphate. Under orthophosphate limitation, algae hydrolyse ambient organic phosphates using extracellular phosphatases and transport the orthophosphate into their cells (Jansson et al. 1988). Whitton et al. (1991) compared the growth of cyanobacterial strains in the presence of organic or inorganic phosphate, finding a similar growth rate. The extracellular phosphatase activity (APA) in several phytoplankton species has been demonstrated by the enzyme-labelled fluorescence (ELF) technique (Nedoma et al. 2003; Štrojsová et al. 2003). This technique permits both the quantification of the enzyme produced and the microscopic localization of the enzyme. Pcy can produce alkaline phosphatases under conditions of phosphate starvation (Simon 1987) but, up to now, none has been observed to show APA-activity using the ELF technique (A. Štrojsová, 2002 personal communication). The method detects only phosphomonoesterase, not phosphodiesterase, activity; therefore caution is needed in interpreting the significance of surface phosphatase activity (Whitton et al. 2005). A genetic study on marine strains has revealed inter-strain variability in the presence and/or absence of genes governing P-acquisition, scavenging and regulation (Moore et al. 2005). Such genetic variability will clearly influence the different physiological responses to low P concentrations of individual strains, e.g. the production of APA.
There are other alternative ways for Pcy to overcome P limitation. Two pathways are of interest: one has been discovered from the presence of genes necessary for phosphonate utilization in the genome of Pcy (Palenik et al. 2003; Ilikchyan et al. 2009). Phosphonates are refractory high-molecular-weight components of the dissolved organic phosphorus (DOP) pool. Marine and freshwater Pcy have the phnD gene, which is thought to encode a phosphonate-binding protein, and in a study induction of phnD expression in P-deficient media was demonstrated (Ilikchyan et al. 2009). This suggests that in P-limiting conditions Synechococcus is able to survive utilizing this refractory form of DOP, derived also from a common herbicide. The other alternative derives from the ability of marine cyanobacteria, and particularly Prochlorococcus, to substitute sulphate (SO 2−4 ) for PO 3−4 in lipids, thus minimising their phosphorus requirement by using a ‘sulphur-for-phosphorus’ strategy. The strategy of synthesizing a lipid that contains sulphur and sugar instead of phosphate could represent a fundamental biochemical adaptation of Pcy to dominate severely phosphorus-deficient environments (Van Mooy et al. 2006, 2009).
There is evidence that ammonium is the preferred form of nitrogen for Synechococcus in culture (Glibert and Ray 1990), but when ammonium is exhausted Synechococcus can take up nitrate, thanks to a regulatory mechanism that can induce expression of nitrate reductases (Bird and Wyman 2003). Also, under severe N-limitation Pcy can alternatively use the nitrogen reserve that exists in phycobiliproteins as amino acids storage molecules (Grossman et al. 1993). The success of Pcy under low light conditions is tightly coupled with competition for limiting nutrients. In this way, low-light adaptation in Synechococcus is probably of greatest ecological advantage when low-P conditions constrain the growth of all autotrophs (Wehr 1993). Thus the pulsed addition of P can have an interactive effect, because it increases the prevalence of larger algae that can alter the light climate, thereby increasing light limitation which will enhance the growth of Pcy.
Good evidence on the interplay between P, irradiance and primary production of Pcy and how it is mediated in the field is difficult to envisage, but some clues come from the comparison of six ultra-oligotrophic North Patagonian lakes and from the subalpine Lake Maggiore (Fig. 8.19) (Callieri et al. 2007). In the ultra-oligotrophic lakes Pcy production was inversely significantly correlated to PAR but not to P, indicating that in such extremely nutrient deplete ecosystems, low P concentrations were not the limiting resource driving Pcy production. One interpretation of these results is that high irradiance is photo-inhibiting Pcy production and hence is the key driving variable and not phosphorus concentration. Conversely, in the oligo-mesotrophic Lake Maggiore neither P nor light were not correlated to Pcy production. Similar finding are reported by Lavallée and Pick (2002) who found a lack of correlation between pico-phytoplankton growth rates and any form of dissolved phosphorus.

Multiple linear regression analysis of the Pcy primary production (PP) versus irradiance (PAR) and P (total dissolved PO4-P) in: (a) Lake Maggiore; (b) six north Patagonian lakes (Partly from Callieri et al. 2007, modified)
Light is an important factor in niche differentiation for Pcy. The response of Pcy to different light intensity has been studied both in laboratory experiments and in situ, and it has been shown that the optimum growth rate of Synechococcus occurs at low light intensities (Waterbury et al. 1986), notably at a quantum flux of 45 μmol photon m−2 s−1 where their highest growth has been observed (Morris and Glover 1981). These findings agree with field observations where the maximum peak abundance has been found deep in the Atlantic mixed layer (Glover et al. 1985), and in the DCM (deep chlorophyll maximum) of Lake Stechlin (Gervais et al. 1997) and of North Patagonian ultra-oligotrophic Andean lakes (Callieri et al. 2007). In lakes worldwide Pcy have been found at a variety of depths and light irradiance (Fahnenstiel and Carrick 1992; Nagata et al. 1994; Callieri and Pinolini 1995; Callieri and Piscia 2002), confirming the classification of Synechococcus as a euryphotic organism (Kana and Glibert 1987b). One explanation of Pcy tolerance and adaptation to high irradiance is the identification of a process that prevents photo-damage in open ocean Pcy by maintaining oxidized PSII reaction centres, channeling the electrons from PSI to oxygen through a specific oxidase (Mackey et al. 2008). Because of this process Pcy possess an efficient mechanism for dissipating PSII excitation energy, decreasing any potential photo-damage. Nevertheless the relative phylogenetic complexity of the Synechococcus and Cyanobium genera does not presently permit the simple discrimination of high light- and low light-adapted ecotypes, as has been attained for Prochlorococcus (Scanlan and West 2002; Ahlgren and Rocap 2006).
Synechococcus ecotypes exhibit differences in their accessory pigments that affect their adaptation to spectral light quality. It was found that in highly coloured (humic) lakes, non-phycoerythrin cells dominated numerically, while in clearer, oligotrophic hard-water lakes, phycoerythrin-rich cells were the most abundant (Pick 1991). The influence of underwater light quality on the selection of Pcy types having different pigment content has been studied in many aquatic systems, covering a wide spectrum of trophic states and underwater light quality (Vörös et al. 1998; Stomp et al. 2007). Vörös et al. (1998) found that the percentage of PE-rich cells in the total Pcy community increased with increasing values of the KRED/KGREEN ratio, while concurrently the total chlorophyll a concentration decreased and the waters became more transparent and less productive (Fig. 8.20). In laboratory experiments, it has been shown that Pcy grow better when they have a phycobiliprotein whose absorption spectrum is complementary to that of the available light (Callieri 1996) and subsequent experiments showed that PE-rich cells prevail in green light and PC-rich cells in red light but when grown together in white light, can co-exist, absorbing different parts of the light spectrum (Stomp et al. 2004). The importance of red light for phycocyanin and biomass production has been shown in laboratory experiments with a PC-rich Synechococcus strain (Takano et al. 1995), while blue and green wavelengths of light are used more efficiently than red of similar intensity by PE-rich Synechococcus (Glover et al. 1985).

Relative abundance of phycocyanin-rich cells (PC) and phycoerythrin-rich cells (PE) in different aquatic systems in relation to the underwater light climate expressed as the ratio between the extinction coefficient of red and green wavelengths (KRED/KGREEN). When the KRED/KGREEN ratio is >1 the extinction of red light is high and the dominant underwater light is green. Very low values of KRED/KGREEN ratio indicate a red dominant underwater radiation (Modified from Vörös et al. 1998)
The pigment composition of Pcy represents a characteristic spectral signature that can define individual strains, but closely related strains can have different pigment composition (Everroad and Wood 2006). In particular both pigment types have been found in several non-marine Pcy clusters (Crosbie et al. 2003b). A new clade, sister to Cyanobium, was reported from oceanic waters, based upon phylogenetic analysis of concatenated 16S rDNA and rpoC1 data sets (Everroad and Wood 2006). This large clade includes both PE-rich and PC-rich strains. Similarly, marine cluster B (MC-B) also contains PE-rich and PC-rich strains, and this cluster is polyphyletic, consisting of at least two different sub-clusters (Chen et al. 2006). The phylogeny derived from the cpcBA operon of the green PC pigment was better able to separate differently pigmented Pcy than 16S rRNA-ITS phylogeny (Haverkamp et al. 2008). The ecological implication of these findings is that Synechococcus from different lineages can occupy different niches; or alternatively, if the environment offers greater variability and more suitable niches, like in the Baltic Sea (Haverkamp et al. 2009) or in Lake Balaton (Mózes et al. 2006), they can coexist.
Laboratory experiments with freshwater strains from different phylogenetic groups acclimated at low and medium irradiance show that photosynthetic responses are strain-specific and sensitive to photo-acclimation (Callieri et al. 2005; Moser et al. 2009) (Fig. 8.21). PE-rich Pcy from Group B, subalpine cluster I (sensu Crosbie et al. 2003a, b), are more sensitive to photo-acclimation than PC-rich cells from Group I and from Group A, Cyanobium gracile cluster. Therefore ecophysiological differences seem to be more related to the pigment type. Nevertheless the extent of photo-adaptation is strain-specific and depends on the duration of the photo-acclimation (Moser et al. 2009).

Photosynthesis/Irradiance (P/E) curves of three Pcy strains: (a) PE-cells MW4C3 from the Group B, Subalpine cluster; (b) PC-cells MW100C3 from Group I; (c) PC-cells BO8801 from Group A, Cyanobium cluster. Dark symbols = medium light acclimation (100 μmol photon m−2 s−1); light symbols = low light acclimation (10 μmol photon m−2 s−1) (From Moser et al. 2009, modified)
Synechococcus strains are most often grown in a medium at a neutral pH (Stanier et al. 1971). Their preference for neutral or slightly alkaline conditions is also evident in their abundance and distribution patterns in freshwater ecosystems (Stockner 1991b). The trend towards Pcy disappearance with decreasing pH and their replacement by picoeukaryotes has been noted in three dystrophic Canadian lakes (Stockner and Shortreed 1991) and in several low pH, humic Danish lakes (Søndergaard 1991). Also in seven humic lakes situated in Lappland (Sweden) pH was among the abiotic variables most affecting pico-phytoplankton distribution and abundance (Drakare et al. 2003). The effect of lake acidification on the microbial community can be indirect by altering community structure and hence carbon flow to higher trophic levels, or direct by inducing physiological stress. In a study on a Swedish acidified lake before and after liming a non-edible CPcy, Merismopedia tenuissima, was the dominant in the late summer phytoplankton community in the naturally acidic lake, but the population was removed by liming (Bell and Tranvik 1993; Blomqvist 1996). Unfortunately the authors present no data on Pcy abundance in this lake. However, they suggest a likely allelopathic mechanism to explain the population dynamics of Merismopedia tenuissima (Blomqvist 1996; Vrede 1996). In Lake Paione Superiore, an acid sensitive lake above the tree line in the Italian Alps, Pcy populations are very low, and their contribution to microbial food webs appears to be negligible (Callieri and Bertoni 1999). In alpine lakes the effect of low pH and of photo-inhibition has been indicated as the major cause of low Pcy numbers found in those lakes (Straškrabová et al. 1999a). The presence of a shift from Pcy to net plankton has been described in mesocosm experiments (Havens and Heath 1991), and it has been noted that as pH declines the proportion of larger species tends to increase and become dominant (Schindler 1990). Nevertheless, to our knowledge no experimental studies of the influence of pH on Pcy strains have been done, so at this stage it is difficult to discuss ranges of pH tolerance by Pcy or physiological mechanisms of adaptation to low pH in lakes. The only possible generalisation at this stage is that Pcy and many CPcy are not common in lakes with a pH <6.0, and are seldom mentioned or included in studies on acidic lakes because they are probably in low abundance or absent.
4.4 Ultraviolet Radiation
There is considerable evidence that ultraviolet radiation (UVR) has a pronounced influence on aquatic organisms and on their community structure in both marine and freshwaters (Häder et al. 2007). Picoplankton are thought to be particularly vulnerable to UVR because: (1) their small size does not permit the intracellular production of sunscreen compounds (Garcia-Pichel 1994); (2) the small ‘package’ effect leads to higher pigment-specific absorption (Morel and Bricaud 1981) and (3) the distance between the cell surface and the nucleus (DNA) is shortened and the DNA damage induced by UV-B is increased. Thymine dimers like cyclobutane pyrimidine dimer (CPD) are frequently built upon DNA lesions under UV-B radiation and have been recovered in marine phytoplankton (Buma et al. 1995) and Argentinean lakes (Helbling et al. 2006).
Although the higher vulnerability of picoplankton is predictable in theory, contrasting results have been obtained in field studies. Laurion and Vincent (1998) studying size-dependent photosynthesis in a sub-arctic lake have shown that cell size is not a good index of UVR sensitivity. Further, they indicated that Pcy are less sensitive to UVR fluxes and that genetic difference between taxa, more than size, are important in determining the tolerance to UVR; while other authors obtained evidence of an higher vulnerability of smaller algae to UVR (Kasai et al. 2001; Van Donk et al. 2001). A high Pcy sensitivity to UVR radiation in comparison to nano-phytoplankton was observed in the biological weighting functions (BWF) in a high altitude alpine lake by Callieri et al. (2001) (Fig. 8.22). A possible interpretation of these contrasting results is that small cells are likely more susceptible to DNA damage than large cells but they are able to acclimate faster, within hours (Helbling et al. 2001), and are more resistant to photosystem damage (Villafañe et al. 2003).

Biological weighting functions (mW m−2 nm−1)−1 for inhibition of photosynthesis for L. Cadagno picophytoplankton (blue line) and whole assemblage (red line) on 13 September 1999. Broken lines show estimated 95% confidence interval for individual coefficient estimates (From Callieri et al. 2001)
The spectral quality of the UVR exposure, its duration and photon flux density, strongly influences the effect on phytoplankton communities (Harrison and Smith 2009). The damaging power of radiation generally increases from PAR through UV-A into the UV-B wavebands, but this general pattern may still be questioned (Harrison and Smith 2009). There is evidence that many aquatic organisms react promptly to UV-B stress by producing protective substances such as mycosporine-like amino acid compounds (MAAs) (Sinha and Häder 2008), which have absorption maxima ranging from 310 to 359 nm (Carreto et al. 1990; Karentz et al. 1991). In particular cyanobacteria react in response to UV-A radiation by producing an extracellular yellow-brown pigment – scytonemin, that absorbs most strongly in the UV-A spectral region (315–400 nm) (Garcia-Pichel and Castenholz 1991; Dillon et al. 2002). The sunscreen capacities of MAAs and scytonemin are higher if they are present concurrently, and their production is considered an adaptive strategy of photo-protection against UVR irradiance (Garcia-Pichel and Castenholz 1993). Also it is recognized that the UV-B induced production of CPD is counterbalanced by repair mechanisms based on the production of enzymes known as photolyases (Jochem 2000).
Therefore aquatic microorganisms have numerous mechanisms of protection against UVR which influence their global community responses in nature. It has been recognized that many of the effects of solar UVR are caused by wavelengths in the UV-A range, which are not affected by changes in stratospheric ozone (Sommaruga 2009). The higher photo-inhibiting effect of UV-A than UV-B on different size fractions of phytoplankton has been described in several different lakes (Callieri et al. 2001, 2007; Villafañe et al. 1999) and in marine habitat as well (Villafañe et al. 2004; Sommaruga et al. 2005). Callieri et al. (2001) explain the negligible impact of UV-B on in situ phytoplankton production with the lower weighted irradiance brought about by the high Kd at short wavelengths and low incident flux, whereas with UV-A the weighted irradiance is higher due to a greater incident flux and lower Kd.
Mixing is an important factor affecting the degree of exposure to UVR. Vertical mixing transports the cells to depth where active repair takes place and subsequently re-exposes them to higher UVR, upon transport again to near surface depths. Species which form surface blooms, like colonial Microcystis aeruginosa, can also withstand high UVR, synthesizing carotenoids and MAAs (Liu et al. 2004). Toxin biosynthesis by M. aeruginosa may also be influenced by UVR, with the idea that microcystins present inside or outside the cell function as a metal ligands to reduce metal toxicity (Gouvea et al. 2008). The hypothesis that UVR and the bioavailability of trace metals can act as a trigger for microcystin production is fascinating, and would help to explain the selective advantage of cystin production by M. aeruginosa, but it is not yet proven. To explain the resistance of CPcy to UVR it is interesting to note that the colonial morphotype of Microcystis can synthesise substances such as D-galacturonic acid, which is the main component of the slime layer of Microcystis (Sommaruga et al. 2009), and which may hence provide a protective function. To better understand the formation of microcolonies (Callieri et al. 2011) used PE-rich and PC-rich Synechococcus strains of different ribotypes acclimated at moderate (ML) and low (LL) light, and exposed the strains to different levels of UVR under controlled conditions. PE-rich Synechococcus acclimated to LL had a low carotenoid/chlorophyll a (car/chl) ratio but responded faster to UVR treatment, producing the highest percentages of microcolonies (Fig. 8.23) and of cells in microcolonies. Conversely, the same strain acclimated to ML, with a higher car/chl ratio, did not aggregate significantly. These results suggest that microcolony formation by PE-rich Synechococcus is induced by UVR if carotenoid levels are low. PC-rich Synechococcus formed a very low percentage of microcolonies in both acclimations even with low car/chl ratio. It is likely that some Synechococcus strains react to UVR finding a refuge through a morphological adaptation, inclusive of slime layer protection, similar to that noted in Microcystis (Sommaruga et al. 2009). Therefore, in the equilibrium between single cells versus microcolonies or even larger colonial morphologies, the importance of solar radiation (UVR and PAR) should not be underestimated but considered together with other important factors like the nutrient status of the ecosystem and grazing.

Percentage of microcolony number on the sum of single-cell plus microcolony number. PE and PC-rich Synechococcus strains in the treatments (+UVR, +UVR50, -UVR, and control) at two acclimations (LL: 10 μmol m-2 s-1 and ML: 100 μmol m-2 s-1), during experiment times (6 days) (from Callieri et al. 2011, AEM)
4.5 Biotic Interactions
Grazing studies have been stimulated by several new methodologies. Rates of Pcy removal by grazers have been measured using five basic techniques: (1) metabolic inhibitors (Campbell and Carpenter 1986b); (2) diffusion chambers and the dilution technique (Landry and Hassett 1982); (3) fluorescent labelled particles (FLP) (Sherr et al. 1987); (4) direct cell counts (Waterbury et al. 1986) and (5) radioisotope-labelled prey (Iturriaga and Mitchell 1986). Some of these methods have been improved (Landry et al. 1995; Sherr and Sherr 1993) and others developed with the use of modern techniques, e.g. by combining FLP and flow cytometry for cell counting (Vázquez-Domínguez et al. 1999) or using a RNA stable isotope probing technique (Frias-Lopez et al. 2009). In the past various growth inhibitors were tested, including the eukaryote inhibitors colchicine and cycloheximide, which have been used to stop protozoan Pcy grazing activity (Campbell and Carpenter 1986b; Caron et al. 1991). Liu et al. (1995) used kanamycin as an effective growth inhibitor of Synechococcus and Prochlorococcus to estimate growth and grazing rates. Using this approach the mortality of Pcy in marine systems due to grazing has been estimated to range from 43% to 87% of growth rate in marine systems (Liu et al. 1995).
It is not surprising that the existence and continuing development of so many methodologies to measure grazing has produced such diverse and often contradictory results in the literature. For example Sherr et al. (1991) estimated that in Lake Kinneret ciliate carbon requirement could not be obtained only from a Pcy energy source, and they suggest that picoeukaryotic cells must be grazed as well to fulfil growth requirements. However, Šimek et al. (1996) show that some of the most common freshwater ciliate species can survive solely on a diet of Pcy. A tentative annual balance of energy flow in a deep oligotrophic lake estimated that between 83% and 97% of the carbon produced by Pcy is taken up by protozoa and channelled to metazooplankton (Callieri et al. 2002). Nevertheless, there are large losses of organic carbon through respiration during this transfer, along the trophic chain (Sherr et al. 1987). These discrepancies and contrasting results have enhanced the discussion to improve our understanding of the impact of different grazers (protozoans and metazooplankon) on Pcy and on energy transfer, along with this “trophic repackaging”.
Heterotrophic and mixotrophic nanoflagellates and small ciliates have been recognised as the most important grazers of Pcy (Stockner and Antia 1986; Bird and Kalff 1987; Weisse 1990; Christoffersen 1994; Šimek et al. 1995; Sanders et al. 2000). Among the ciliates (Fig. 8.24), oligotrich species and some scuticociliates, which are sometimes at the borderline between nano- and microplankton (<30 μm), can be important picoplanktivores in lakes (Šimek et al. 1995; Callieri et al. 2002). Šimek et al. (1996) have summarised three ecological categories of freshwater ciliates with different feeding strategies and a decreasing importance of pico-size prey in their diet. Among the most efficient suspension feeders there are some very active Pcy grazers, e.g. Vorticella aquadulcis (Fig. 8.25), Halteria grandinella, Cyclidium and Strobilidium hexachinetum. These protozoa are able to graze 560, 210, 80, 76 Pcy ciliate−1 h−1, respectively, with clearance rates highly variable among taxa, 11–3150 nL × cells × h−1 (Šimek et al. 1996). In a warm-monomictic saline lake in Mexico lower uptake rates by vorticellids and mixotrophic Euplotes have been measured, ranging from 16 to 227 Pcy ciliate−1 h−1 (Peštová et al. 2008), but the authors noted the importance of both groups as selective Pcy grazers. Large mixotrophic ciliates, common in ultra-oligotrophic south Andean lakes, are also recognised as preying upon Pcy (Modenutti et al. 2003; Balseiro et al. 2004).

The ciliate Limnostrombidium sp. coloured with DAPI and visualised under UV (above) and blue (below) excitation (epifluorescence microscope 787.5×). In DAPI the nucleus is clearly visible and in blue excitation vacuoles full of yellow Pcy appear

The ciliate Vorticella sp. coloured with DAPI and visualized under UV (above) and blue (below) excitation (epifluorescence microscope 787.5×). In DAPI the nucleus is clearly visible and in blue excitation vacuoles full of yellow Pcy appear
Despite the importance of ciliate grazing on Pcy in some systems, it is generally recognized that among protozoa, both heterotrophic and mixotrophic nanoflagellates are responsible for 90% of the grazing of Pcy and bacteria; whereas ciliates accounted for only 10% (Pernthaler et al. 1996). A study on Lake Maggiore showed that heterotrophic nanoflagellates (HNF) ingested from 0.5 to 3 Pcy h−1, while ciliates ingested from 18 to 80 Pcy h−1 (Callieri et al. 2002). Nevertheless, at the community level, the grazing impact of HNF was one order of magnitude higher than that of ciliates (maxima: 8,000 Pcy mL−1 h−1 and 400 Pcy mL−1 h−1, respectively) (Callieri et al. 2002). In Lake Tanganika similar results were obtained with the higher impact of HNF (av: 8027 Pcy mL-1 h-1) than of ciliates (maxima: 1355 Pcy mL-1 h-1) on Pcy grazing in the dry season (Tarbe et al. 2011). Pernthaler et al. (1996) have emphasised the influence of community composition and taxa-specific clearance rates on the grazing impact on bacteria and Pcy. The size of the prey, its morphological characteristics and nutritional value have been indicated as important factors in the selection carried out by the predators (Šimek and Chrzanowski 1992; Jezberová and Komárková 2007; Shannon et al. 2007). In particular the involvement of the proteinaceous cell surface (S-layer) as grazing protection has also been suggested for freshwater Actinobacteria (Tarao et al. 2009). Morphological characteristics can therefore be considered as group-specific traits and can greatly influence the success of the group in an ecosystem (Tarao et al. 2009). Protozoa grazing and in particular nanoflagellates can influence the characteristics of bacterial and Pcy communities and lead to changes in their structural and taxonomic composition. A laboratory study with 37 Synechococcus strains showed clearly that prey selection discriminates at the strain-specific level (Zwirglmaier et al. 2009).
The selection of food as described for metazooplankton generally takes place during food capture and processing (Porter 1973). According to the theory of “selective digestion” prey selection takes place inside the food vacuoles (Boenigk et al. 2001). The fate of the prey is decided at the moment of digestion, with the possibility of very fast prey-excretion after the uptake. Knowledge of the mechanism of Pcy consumption and excretion/digestion is species-specific both for prey and predator. Boenigk et al. (2001) demonstrated that prey characteristics and predator satiation strongly influence the ingestion and digestion process, e.g. the digestion strategies of Cafeteria, Spumella and Ochromonas are different; with Pcy rapidly excreted while bacteria were directly digested in the food vacuoles. Amoebae can perform food selection in the food vacuole and excrete the toxic or unpalatable prey items similarly to nanoflagellates (Liu et al. 2006; Dillon and Parry 2009).
Ciliates and nanoflagellates can also serve as a trophic link between Pcy production and Daphnia production, thereby upgrading the nutritional value of Pcy as a food source by producing essential lipids such as sterols (Martin-Creuzburg et al. 2005; Martin-Creuzburg and Von Elert 2006; Bec et al. 2006). Among mesozooplankton, Daphnia has the capacity of feeding on a wide particle size range (1–50 μm), filtering Pcy as well (Gophen and Geller 1984; Stockner and Porter 1988) (Fig. 8.26). Together with Daphnia, several cladoceran genera, including Bosmina, Eubosmina and Ceriodaphnia, are able to ingest Pcy (Weisse 1993). Suspension-feeding cladocerans may have a direct grazing effect on Pcy and an indirect effect by regenerating nutrients (Carrillo et al. 1996; Balseiro et al. 1997). The recycling of excreted nutrients moves the nature of algal-bacterial interactions from one of competition to commensalism (Reche et al. 1997).

Cladoceran gut coloured with DAPI and visualised under UV (above) and blue (below) excitation (epifluorescence microscope 787.5×); in blue excitation yellow Pcy appear
An important effect of grazing by Daphnia on Pcy functioning was observed in laboratory experiments (Callieri et al. 2004), where there was an increase in P and C cell-specific uptake by Pcy and in their photosynthetic efficiency. This activity could have been related to the release of P by Daphnia, which has been reported to be 5% of the total P-pool per day (Boersma and Wiltshire 2006). Another possible conjecture is that nutrients are replenished during the passage of Pcy through the digestive tracts of consuming daphnids (Porter 1975; Stockner 1991b). There is evidence that nutrient-limited green algae pass through the gut of Daphnia intact and alive (Van Donk and Hessen 1993) and that during passage they can use some of the P released in the gut (Boersma and Wiltshire 2006).
Among lake studies of Pcy, only a few refer to the impact of copepod grazing, particularly calanoid copepods. It has been shown that copepods have a stronger negative effect on ciliates than do Daphnia (Burns and Schallenberg 1996) and that top-down effects in the short term are stronger in oligotrophic ecosystems than in eutrophic ones (Burns and Schallenberg 2001). The mesocosm experiments of Zöllner et al. (2003) showed the structuring and cascading effects of the cladoceran Daphnia hyalina cf. galeata and copepods (50% Eudiaptomus spp. and 50% copepodite stages of cyclopoid copepods) on microbial food web structure. These investigators found a decrease in Pcy that was probably due to the selective feeding of copepods on intermediate-sized ciliates and a strong increase in the concentrations of HNF, as was previously found for bacteria (Jürgens and Jeppesen 2000). Copepods prey selectively and efficiently on ciliates and algae in the size range 20–40 μm (Yoshida et al. 2001), thereby triggering a trophic cascade, enabling high numbers of HNF and the potential for a greater mortality of Pcy (Zöllner et al. 2003). Sundt-Hansen et al. (2006) have shown that in marine mesocosms, copepods have a profound structuring effect on the pelagic food web, and thus directly and indirectly regulate the abundance of Pcy predators. In this way, the strength of the trophic cascade downward to Pcy depends substantially on the structure of the food web and the inventory of zooplankton species present (Gismervik 2006; Van Gremberghe et al. 2008).
Rotifers can either act directly on Pcy populations by grazing or indirectly by preying on nanoflagellates and small ciliates (Stockner and Shortreed 1989; Arndt 1993) (Fig. 8.27). Many planktonic rotifers (Keratella cochlearis, K. quadrata, Polyarthra dolichoptera) feed on particles in the size-range 0.5–3 μm, interspecific variation in food selection being dependent on differences in the corona sizes of the consuming species (Ronneberger 1998). As Stockner and Antia (1986) asserted, Pcy are within the size range suitable for grazing by nauplii and early copepodite stages of copepods. This possibility has been partially confirmed by the direct estimation of the grazing rate on Pcy and bacteria by a copepod naupliar stage in a marine system (Roff et al. 1995). At present there are few studies, all in marine systems, on the role of bivalve and gastropod larvae as possible picoplanktivores. In one of these, Bell (1991) showed that Crepidula aculeata and Littoraria scabra, two gastropod larvae, can thrive on a Pcy and bacteria diet. Furthermore, the larvae of a bivalve, Mercenaria mercenaria, have been shown to ingest and grow on a Synechococcus strain (Gallager et al. 1994). Thus, there now appears to be some coupling between microbial and littoral benthic food webs in aquatic ecosystems.

The rotifer Keratella cochlearis coloured with DAPI and visualised under UV (above) and blue (below) excitation (epifluorescence microscope 787.5×); in blue excitation vacuoles full of yellow Pcy appear
Zaret and Suffern (1976), who studied the grazing effect of zooplankton on phytoplankton in Gatun Lake, Panama, found that CPcy were not grazed, and they suggested that this was probably due to the fact that cells were embedded in a gelatinous matrix. Blomqvist (1996) and Vrede (1996) have reported Merismopedia tenuissima to be resistant to grazing by the dominant grazer Eudiaptomus in a clear water, oligotrophic lake in central Sweden. Thus, from what little is known at this time, it seems probable that the mucilaginous colonial morphology, as in larger colonial cyanobacteria, is an effective anti-grazing adaptation that, when coupled with the possibility of allelopathic strategies of CPcy colonies, creates the perfect, coupled anti-predation device for these CPcy species. This may very well be the reason for their ubiquity and abundance in lakes.
While predation has been recognised as an important top-down structural and dynamic control of Pcy, little attention has been directed towards the study of other ecological interactions such as symbiosis (Adams 2000; Chap. 23). Marine cyanobacterial symbionts (or ‘cyanobionts’: Taylor 1982) provide an example of proto-cooperation. A symbiotic relationship between oceanic unicellular cyanobacteria and a tintinnid, Codonella sp., was demonstrated by Carpenter and Foster (2002). In an especially interesting study, Foster et al. (2006) used molecular methods to amplify prokaryotic symbiont rRNA sequences from individual marine cells of various marine eukaryotes. Their results showed 53% of the cyanobacterial symbionts to be closely related to Synechococcus sp. and 3% to Prochlorococcus sp. The same symbiont was found capable of forming associations with a variety of organisms, thus opening up the possibility of consortial interconnections. Nevertheless, it must also be recognised that these dinoflagellates, radiolarian and tintinnid symbioses are very low in abundance and generally confined to the upper 50 m of the ocean.
Another approach to the study of biological interactions is to consider the in situ occurrence of groups of species that share similar requirements or even show proto-cooperative interaction. The natural co-occurrence and simultaneous increase or decrease in the numbers of some species may indicate the existence of ‘functional associations’ that help us to interpret and predict their dynamics (Reynolds et al. 2002). The supposition at the base of such associations is that common morphological or physiological properties offer relative, dynamic advantages of component species of the association. A new association was proposed that comprises Synechococcus spp. and potentially mixotrophic flagellates e.g. Rhodomonas lacustris, Ceratium hirundinella, Cryptomonas erosa (Callieri et al. 2006). Co-occurrence of Pcy and Ceratium spp. has been reported from mesotrophic lakes (Kasprzak et al. 2000), from Lake Kinneret (Berman et al. 1992) and Lake Maggiore (Callieri et al. 2006) (Fig. 8.28). In the latter lake, a 3-year study showed a phase of co-existence in which the organisms might each benefit from the association, followed by a phase of predation in which one member of the association prevailed over the other (Fig. 8.29). At low levels of physical and biological disturbance, the cycle can restart with prey recovery driven by nutrient excretion of phagotrophs. The association indicates that assemblages that form a functional group may not only have similar adaptations and requirements, but can exhibit predator–prey interactions, as was shown in a marine lagoon in France where the quasi simultaneous appearance of both Pcy and the dinoflagellate Alexandrium catenella was observed (Collos et al. 2009). These authors hypothesised that Pcy can make up for a particulate nitrogen form during periods of limiting nutrients, thus providing A. catenella an ecological advantage over strictly autotrophic phytoplankton. The co-domination of a desirable prey organism, such as Synechococcus with its potential grazers opens up new perspectives on the interaction between the ecological categories of phytoplankton and the components of the microbial food web. We cannot refrain from conjecturing that these functional associations may be an advanced phase of a symbiotic association of cyanobacteria with eukaryotic plankton hosts, similar to those observed in the ocean (Carpenter and Foster 2002; Foster et al. 2006).

The dinoflagellate Ceratium hirundinella from Lake Maggiore observed with the vacuole full of yellow Pcy. Coloured with DAPI and visualised under UV (left) and blue (right) excitation (epifluorescence microscope 787.5×)

Seasonal dynamics (years: 2002, 2003, 2004) of Synechococcus spp. and Ceratium hirundinella biovolume, in Lake Maggiore (From Callieri et al. 2006, modified)
In our consideration of biological interactions, it is also opportune to refer to viral infections, as it is widely demonstrated that prokaryotic viruses can influence cellular organisms stronger than previously thought (Weinbauer 2004). The occurrence of viruses that infect Synechococcus is widespread and there is agreement that phages exert a significant selection pressure on Synechococcus (Mann 2003). Cyanophages are ubiquitous in aquatic environments, and can occur at abundances in excess of 106 mL−1 (Suttle 2000). Findings indicate that cyanophage infections can exert a major influence on the direction of Pcy succession in the sea (Mühling et al. 2005), and that marine viruses can act as intermediates for exchanging genes (Zeidner et al. 2005). Transduction is the phage-mediated gene transfer between a donor and a host, and has been recognized as is an important factor for bacterial evolution (e.g. Doolittle 1999). In three peri-alpine lakes viral impact on Pcy exceeded predation in autumn, but was highly variable throughout the early season (Personnic et al. 2009b). However, the interplay between viruses and nanoflagellates and their control of prokaryotes is not completely understood, largely due to a lack of knowledge of the direct interactions of viruses on predators and vice-versa (Jacquet et al. 2007; Pradeep Ram and Sime-Ngando 2008; Massana et al. 2007).
The extent of lysogeny in 19 freshwater Synechococcus strains indicated a high level within PC-rich Synechococcus (Dillon and Parry 2008). These authors found that the majority of cyanophages in the eutrophic lake they studied were temperate, that is they exist in a lysogenic association with their hosts. In the majority of the strains cell lysis by the phage was only triggered by an inducing agent used experimentally to assess the level of temperate phage infection in the host population. On the other hand viral DNA might also have a protective role on the host (Bailey et al. 2004). An example is provided by the discovery of a cyanophage encoding polypeptide D1 and D2 of the PSII, inducing repair cycles after photo-damage (Bailey et al. 2004). Nevertheless it is not known at which extent the phage modifies the properties of PSII and therefore manipulates the photosynthetic physiology of the infected cells.
Finally it is interesting to note the changing perceptions of cyanophages and other viruses in recent years; that is they are no longer seen as universally pernicious parasites, but as catalysts of information transfer and sustainers of the microbial web of energy transfer and matter cycling in aquatic ecosystems (Weinbauer 2004; Chap. 21).
5 Conclusions
Important advances in our perception of the significance of picocyanobacteria in freshwaters and oceans have occurred only within the last few decades, and these findings have come largely from an improved understanding of phylogenetic evolution of this major group. We now know that evolution of earliest cyanobacterial lineages were not marine, but likely were of terrestrial or freshwater origin and were unicellular. New paradigms in microbial ecology are now founded on a worldwide appreciation of functionally evolving clades and on genetically definable ecotypes. Taxonomic studies of phenotypic diversity are now coupled with genotypic diagnoses that can confirm whether similar phenotypes are phyletically close or due to convergent evolution.
The genera Synechococcus and Cyanobium are the dominant picocyanobacteria of fresh waters, while Prochloro-coccus is typically marine. The phylogenetic analysis of the 16S rDNA sequences indicates that Synechococcus is polyphyletic. Despite some remaining uncertainties about the phylogenetic evolution of Synechococcus, we know that there are at least seven clusters of non-marine picocyanobacteria that have been found within the picophytoplankton clade, and that PE-rich spectral phenotype does not appear to be a general character that can be used to define a clade. Indeed, it has been found that phylogenies based on phycobiliprotein genes are not easily comparable with 16S rRNA phylogeny and reveal a high diversity of Synechococcus strains. The adaptation of different ecological niches can be related to the highly variable number of horizontally acquired genes located in highly variable genomic regions or islands. This discovery opens new perspectives to the understanding of the local adaptation and the definition of species within the Synechococcus group. Further, it appears that the form-genus Synechococcus likely represents the ancestral morphology from which other types, including colonial forms, evolved.
To what extent the formation of microcolonies is due to the presence of specific Synechococcus genotypes or is the result of survival strategy is under debate. Here we consider microcolonies as transition forms from single-cell to colonial morphotypes. Grazing and UVR have been indicated as important factors which could regulate the equilibrium between single cells versus microcolonies or even larger colonial morphologies. The current challenge is to better understand the relationship between the diversity and ecology of Pcy, microcolonies and colonial Pcy and their interaction with the environmental factors that allow the proliferation of the most competitive genotypes.
Compared to our understanding of the physiology and ecology of Pcy, similar studies of colonial pico-cyanobacteria in lakes are sparse, and most published reports are focused on taxonomy rather than ecology. The picture emerging is that many colonial Pcy are part of the metaphyton community, loosely associated with littoral and benthic sediment and macrophytes, but capable of movement by currents, waves to the pelagic habitat where they experience growth and reproduction as plankton.
The morphometry and trophic state of lakes and ponds strongly influences composition, diversity and abundance of Pcy communities, most notably the relative distribution of Pcy PE-rich versus PC-rich cells. Light is known to be an important factor in niche differentiation of Pcy, and so also is in upper, mixed-layer depth, that can exert a strong influence on nutrient flux and lake trophic state. Lake nutrient status in turn directly affects Pcy presence and abundance relative to larger phytoplankton. The vast majority of lake studies now confirm that of total phytoplankton biomass the percentage contributed by Pcy increases with decreasing trophic state; in this sense there is a common ecological response of freshwater and marine Pcy, despite their phylogenetic differences. The acceptance of the validity of this empirical model can partially be explained by the high affinity of Synechococcus for orthophosphate, and its ability to utilise, at low PO 3−4 concentrations, organic sources of phosphate.
Protozoa grazing can influence the characteristics of Pcy communities and lead to changes in their structural and taxonomic composition. The knowledge of the mechanism of Pcy consumption and excretion/digestion is species-specific both for prey and predator. Ciliates and nanoflagellates can also serve as a trophic link between Pcy production and Daphnia production, thereby upgrading the nutritional value of Pcy as a food source by producing essential lipids such as sterols. The co-domination of a desirable prey organism, such as Synechococcus with its potential grazers opens up new perspectives on the interaction between the ecological categories of phytoplankton and the components of the microbial food web.
Adaptation to a changing environment, with phasing of fluctuating events, can subject the community to dominance by the fittest and most adaptive available species. These concepts should be evaluated in light of the new conceptual framework of community ecology, the meta-community, which considers the communities as shaped at different spatial scales (local and regional). Therefore in the debate on biotic versus abiotic regulation of community structure and dynamics we need to consider that local communities are not isolated but are linked by dispersal of multiple, potentially interactive, species.
Abbreviations
- Pcy:
-
picocyanobacteria
- CPcy:
-
colonial picocyanobacteria
- PE-rich:
-
phycoerythrin containing Pcy
- PC-rich:
-
phycocyanin containing Pcy
- Chl:
-
chlorophyll a
- HNF:
-
heterotrophic nanoflagellates
- ITS-1:
-
internal transcribed spacer region between the 16S rRNA and 23S rRNA genes
- T-RFLP:
-
terminal restriction fragment length polymorphism
- DGGE:
-
denaturating gradient gel electrophoresis
- ARISA:
-
automated ribosomal intergenic spacer analyses
- DCM:
-
deep chlorophyll maximum
- OTU:
-
operational taxonomic unit
- RT-qPCR:
-
real-time quantitative polymerase chain reaction
- FDC:
-
frequency of dividing cells
- RUBISCO:
-
ribulose-1,5-bisphosphate carboxylase oxygenase
- ELF:
-
enzyme labelled fluorescence
- APA:
-
extracellular phosphatase activity
- DOP:
-
dissolved organic phosphorus
- CPD:
-
cyclobutane pyrimidine dimer
- BWF:
-
biological weighting functions
- MAAs:
-
mycosporine-like amino acid compounds
- Kd :
-
extinction coefficient of photosynthetically active radiation
References
Adams DG (2000) Symbiotic interactions. In: Whitton BA, Potts M (eds) The ecology of cyanobacteria: their diversity in time and space. Kluwer Academic Publishers, Dordrecht, pp 523–561, 668 pp
Ahlgren NA, Rocap G (2006) Culture isolation and culture-independent clone libraries reveal new marine Synechococcus ecotypes with distinctive light and N physiologies. Appl Environ Microbiol 72:7193–7204
Ahlgren NA, Rocap G, Chisholm SW (2006) Measurement of Prochlorococcus ecotypes using real-time polymerase chain reaction reveals different abundances of genotypes with similar light physiologies. Environ Microbiol 8:441–454
Allende L, Izaguirre I (2003) The role of physical stability on the establishment of steady states in the phytoplankton community of two Maritime Antarctic lakes. Hydrobiologia 502:211–224
Ålvik G (1934) Plankton-Algen norwegischer Austernpollen I. Systematik und Vorkommen der Arten. Bergens Mus Årb 1934(6):47 pp
Arndt H (1993) Rotifers as predators on components of the microbial web (bacteria, heterotrophic flagellates, ciliates) – a review. Hydrobiologia 255(256):231–246
Bailey S, Clokie MRJ, Millard A, Mann NH (2004) Cyanophage infection and photoinhibition in marine cyanobacteria. Mini-review. Res Microbiol 155:720–725
Balseiro EG, Modenutti BE, Queimaliños CP (1997) Nutrient recycling and shifts in N:P ratio by different zooplankton structures in a South Andes lake. J Plankton Res 19:805–817
Balseiro EG, Queimaliños C, Modenutti BE (2004) Grazing impact on autotrophic picoplankton in two south Andean lakes (Patagonia, Argentina) with different light:nutrient ratios. Rev Chil Hist Nat 77:73–85
Beardall J, Raven JA (2004) Potential effects of global change on microalgal photosynthesis, growth and ecology. Phycologia 43:26–40
Bec A, Martin-Creuzburg D, von Elert E (2006) Trophic upgrading of autotrophic picoplankton by the heterotrophic nanoflagellate Paraphysomonas sp. Limnol Oceanogr 51:1699–1707
Becker S, Böger P, Oehlmann R, Ernst A (2000) PCR bias in ecological analysis: a case study for quantitative Taq nuclease assays in analyses of microbial communities. Appl Environ Microbiol 66:4945–4953
Becker S, Fahrbach M, Böger P, Ernst A (2002) Quantitative tracing, by Taq nuclease assays, of a Synechococcus ecotype in a highly diversified natural population. Appl Environ Microbiol 68:4486–4494
Becker S, Singh AK, Postius C, Böger P, Ernst A (2004) Genetic diversity and distribution of periphytic Synechococcus spp. in biofilms and picoplankton of Lake Constance. FEMS Microbiol Ecol 49:181–190
Becker S, Richl P, Ernst A (2007) Seasonal and habitat-related distribution pattern of Synechococcus genotypes in Lake Constance. FEMS Microbiol Ecol 62:64–67
Bell JL (1991) Patches and picoplankton. Effects on larval life spans on gastropod larvae. Am Zool 31:6A
Bell T, Kalff L (2001) The contribution of picophytoplankton in marine and freshwater systems of different trophic status and depth. Limnol Oceanogr 46:1243–1248
Bell RT, Tranvik L (1993) Impact of acidification and liming on the microbial ecology of lakes. Ambio 22:325–330
Belykh OI, Ekaterina G, Sorokovikova T, Saphonova A, Tikhonova V (2006) Autotrophic picoplankton of Lake Baikal: composition, abundance and structure. Hydrobiologia 568:9–17
Bergman E, Hamrin SF, Romare P (1999) The effect of cyprinid reduction on the fish community. In Hansson LA and Bergman B (eds) Nutrient reduction and biomanipulation as tools to improve water quality. The Lake Ringsjön story. Hydrobiologia 404:65–75
Berman T, Yacobi YZ, Pollingher U (1992) Lake Kinneret phytoplankton: stability and variability during twenty years (1970–1989). Aquat Sci 54:104–127
Bertilsson S, Berglund O, Karl DM, Chisholm SW (2003) Elemental composition of marine Prochlorococcus and Synechococcus: implications for the ecological stoichiometry of the sea. Limnol Oceanogr 48:1721–1731
Bertoni R, Piscia R, Callieri C (2004) Horizontal heterogeneity of seston, organic carbon and picoplankton in the photic zone of Lago Maggiore, Northern Italy. J Limnol 63:244–249
Bird DJ, Kalff J (1987) Algal phagotrophy: regulating factors and importance relative to photosynthesis in Dinobryon (Chrysophyceae). Limnol Oceanogr 32:277–284
Bird C, Wyman M (2003) Nitrate/nitrite assimilation system of the marine picoplanktonic cyanobacterium Synechococcus sp. strain WH8103: effect of nitrogen source and availability on gene expression. Appl Environ Microbiol 69:7009–7018
Björk S et al (1972) Ecosystem studies in connection with the restoration of lakes. Verh Int Ver Limnol 18:379–387
Bláha L, Marsálek B (1999) Microcystin production and toxicity of picocyanobacteria as a risk factor for drinking water treatment plants. Algol Stud 92:95–108
Blank CE, Sánchez-Baracaldo P (2010) Timing of morphological and ecological innovations in the cyanobacteria – a key to understanding the rise in atmospheric oxygen. Geobiology 8:1–23
Blomqvist P (1996) Late summer phytoplankton responses to experimental manipulations of nutrients and grazing in unlimed and limed Lake Njupfatet, central Sweden. Arch Hydrobiol 137:425–455
Boenigk J, Matz C, Jürgens K, Arndt H (2001) The influence of preculture conditions and food quality on the ingestion and digestion process of three species of heterotrophic nanoflagellates. Microb Ecol 42:168–176
Boersma M, Wiltshire K (2006) Gut passage of phosphorus-limited algae through Daphnia: do they take up nutrients in the process? Arch Hydrobiol 167:498–500
Bourrelly P (1985) Les Algues d’Eaux Douce. III. Les Algues Bleues et Rouges, Les Eugléniens, Peridiniens et Cryptomonadines. Société Nouvelle des Éditions Boubée, Paris, 606 pp
Buma AGJ, van Hannen EJ, Veldhuis MJW, Gieskes WWC (1995) Monitoring ultraviolet B-induced DNA damage in individual diatom cells by immuno-fluorescent-thymin dimer detection. J Phycol 31:314–321
Burns CW, Schallenberg M (1996) Relative impacts of copepods, cladocerans and nutrients on the microbial food web of a mesotrophic lake. J Plankton Res 18:683–714
Burns CW, Schallenberg M (2001) Short-term impacts of nutrients, Daphnia, and copepods on microbial food-webs on an oligotrophic and eutrophic lake. N Z J Mar Freshw Res 35:695–710
Butcher RW (1952) Contributions to our knowledge of smaller marine algae. J Mar Biol Assoc UK 31:610–652
Callieri C (1996) Extinction coefficient of red, green and blue light and its influence on Pcy types in lakes at different trophic levels. Mem Ist Ital Idrobiol 54:135–142
Callieri C (2008) Picophytoplankton in freshwater ecosystems: the importance of small-sized phototrophs. Freshw Rev 1:1–28
Callieri C (2010) Single cells and microcolonies of freshwater picocyanobacteria: a common ecology. J Limnol 69:257–277
Callieri C, Bertoni R (1999) Organic carbon and microbial food web assemblages in an oligotrophic alpine lake. In: Straškrabová V, Callieri C, Fott J (eds) Pelagic food web in Mountain Lakes. MOuntain LAkes Research Program. J Limnol 58:136–143
Callieri C, Pinolini ML (1995) Picoplankton in Lake Maggiore, Italy. Int Rev Ges Hydrobiol 80:491–501
Callieri C, Piscia R (2002) Photosynthetic efficiency and seasonality of autotrophic picoplankton in Lago Maggiore after its recovery. Freshw Biol 47:941–956
Callieri C, Stockner JG (2002) Freshwater autotrophic picoplankton: a review. J Limnol 61:1–14
Callieri C, Amicucci E, Bertoni R, Vörös L (1996a) Fluorometric characterization of two picocyanobacteria strains from different underwater light quality. Int Rev Ges Hydrobiol 81:13–23
Callieri C, Bertoni R, Amicucci E, Pinolini ML, Jasser I (1996b) Growth rates of freshwater picocyanobacteria measured by FDC: problems and potentials for the estimation of picoplankton organic carbon synthesis. Arch Hydrobiol Spec Issues Adv Limnol 48:93–103
Callieri C, Lami A, Bertoni (2011) Microcolony formation by single-cell Synechococcus strains as a fast response to UV radiation. Appl Environ Microbiol 77:7533–7540
Callieri C, Morabito G, Huot Y, Neal P, Lichman E (2001) Photosynthetic response of pico- and nanoplanktonic algae to UVB, UVA and PAR in a high mountain lake. Aquat Sci 63:286–293
Callieri C, Karjalainen SM, Passoni S (2002) Grazing by ciliates and heterotrophic nanoflagellates on picocyanobacteria in Lago Maggiore, Italy. J Plankton Res 24:785–796
Callieri C, Balseiro E, Bertoni R, Modenutti B (2004) Picocyanobacterial photosynthetic efficiency under Daphnia grazing pressure. J Plankton Res 26:1471–1477
Callieri C, Moro S, Caravati E, Crosbie ND, Weisse T (2005) Strain specific photosynthetic response of freshwater picocyanobacteria. Verh Int Ver Limnol 29:777–782
Callieri C, Caravati E, Morabito G, Oggioni A (2006) The unicellular freshwater cyanobacterium Synechococcus and mixotrophic flagellates: evidence for a functional association in an oligotrophic, subalpine lake. Freshw Biol 51:263–273
Callieri C, Modenutti B, Queimaliños C, Bertoni R, Balseiro E (2007) Production and biomass of picophytoplankton and larger autotrophs in Andean ultraoligotrophic lakes: differences in light harvesting efficiency in deep layers. Aquat Ecol 80:345–362
Callieri C, Caravati E, Corno G, Bertoni R (2012) Picocyanobacterial community structure and space-time dynamics in the subalpine Lake Maggiore (N. Italy). J Limnol 71:95–103
Camacho A, Picazo A, Miracle MR, Vicente E (2003) Spatial distribution and temporal dynamics of picocyanobacteria in a meromictic karstic lake. Algol Stud 109:171–184
Campbell L, Carpenter EJ (1986a) Diel pattern of cell division in marine Synechococcus spp. (Cyanobacteria): use of frequency of dividing cell technique to measure growth rate. Mar Ecol Prog Ser 32:139–148
Campbell L, Carpenter EJ (1986b) Estimating the grazing pressure of heterotrophic nanoplankton on Synechococcus spp. using the seawater dilution and selective inhibitor techniques. Mar Ecol Prog Ser 33:121–129
Caravati E (2008) Biodiversità e caratteristiche eco-fisiologiche dei picocianobatteri d’acqua dolce. Ph D thesis, University of Parma, Parma, 133 pp
Caravati E, Callieri C, Modenutti B, Corno G, Balseiro E, Bertoni R, Michaud L (2010) Picocyanobacterial assemblages in ultraoligotrophic Andean lakes reveal high regional microdiversity. J Plankton Res 32:357–366
Caron DA, Lim EL, Miceli G, Waterbury JB, Valois FW (1991) Grazing and utilization of chroococcoid cyanobacteria and heterotrophic bacteria by protozoa in laboratory cultures and coastal plankton community. Mar Ecol Prog Ser 76:205–217
Carpenter EJ, Foster RA (2002) Marine symbioses. In: Rai AN, Bergman B, Rasmussen U (eds) Cyanobacteria in symbiosis. Kluwer Academic Publishers, Dordrecht, pp 11–18
Carreto JI, Carignan MO, Daleo G, De Marco SG (1990) Occurrence of mycosporine-like amino acids in the red tide dinoflagellate Alexandrium escavatum: UV-photoprotective compounds? J Plankton Res 12:909–921
Carrillo P, Reche I, Cruz-Pizarro L (1996) Quantification of the phosphorus released by zooplankton in an oligotrophic lake (La Caldera, Spain) – regulating factors and adjustment to theoretical-models. J Plankton Res 18:1567–1586
Chen F, Wang K, Kan JJ, Suzuki MT, Wommack KE (2006) Diverse and unique Pcy in Chesapeake Bay, revealed by 16S-23S rRNA internal transcribed spacer sequences. Appl Environ Microbiol 72:2239–2243
Chisholm SW (1992) Phytoplankton size. In: Falkowski PG, Woodhead AD (eds) Primary productivity and biogeochemical cycles in the sea. Plenum Press, New York, pp 213–237
Chisholm SW, Armbrust EV, Olson RJ (1986) The individual cell in phytoplankton ecology: cell cycle and applications of flow cytometry. Can Bull Fish Aquat Sci 214:343–369
Christoffersen K (1994) Variation of feeding activities of heterotrophic nanoflagellates on picoplankton. Mar Microb Food Web 8:111–123
Coleman ML, Sullivan MB, Martiny AC, Steglich C, Barry K, Delong EF, Chisholm SW (2006) Genomic islands and the ecology and evolution of Prochlorococcus. Science 311:1768–1770
Collos Y, Bec B, Jauzein C, Abadie E, Laugier T, Lautier J, Pastoureaud A, Souchu P, Vaquer A (2009) Oligotrophication and emergence of picocyanobacteria and a toxic dinoflagellate in Thau Lagoon, southern France. J Sea Res 61:68–75
Cronberg G (1988) Cyanodictyon tubiforme, a new chroococcal blue-green alga from Lake Börringesjön, Scania, Sweden. Algol Stud 50–53:191–194
Cronberg G (1991) Cyanothamnos plancticus gen. Et sp. Nov., a new colonial cyanophyte from an eutrophic Scanian lake, Sweden. Algol Stud 64:61–70
Cronberg G (1999) Qualitative and quantitative investigations of phytoplankton in Lake Ringsjön, Scania Sweden. Hydrobiologia 404:27–40
Cronberg G (2003) New and interesting cyanoprokaryotes from temperate, brackish ponds and the Baltic Sea. Algol Stud 109:197–211
Cronberg G, Komárek J (1994) Planktic Cyanoprokaryotes found in South Swedish lakes during the XIIth international symposium on Cyanophyte research, 1992. Algol Stud 75:323–352
Cronberg G, Weibull C (1981) Cyanodictyon imperfectum a new chroococcal blue-green alga from Lake Trummen, Sweden. Algol Stud 27:101–110
Crosbie ND, Pöckl M, Weisse T (2003a) Dispersal and phylogenetic diversity of nonmarine picocyanobacteria, inferred from 16S rRNA gene and cpcBA-intergenic spacer sequence analyses. Appl Environ Microbiol 69:5716–5721
Crosbie ND, Pöckl M, Weisse T (2003b) Rapid establishment of clonal isolates of freshwater autotrophic picoplankton by single-cell and single-colony sorting. J Microbiol Method 55:361–370
Crosbie ND, Teubner K, Weisse T (2003c) Flow-cytometric mapping provides novel insights into the seasonal and vertical distributions of freshwater autotrophic picoplankton. Aquat Microb Ecol 33:53–66
Diaz M, Pedrozo F, Reynolds C(S), Temporetti P (2007) Chemical composition and the nitrogen-regulated trophic state of Patagonian lakes. Limnologica 37:17–27
Dillon A, Parry JD (2008) Characterization of temperate cyanophages active against freshwater phycocyanin-rich Synechococcus spp. Freshw Biol 53:1253–1261
Dillon A, Parry JD (2009) Amoebic grazing of freshwater Synechococcus strains rich in phycocyanin. FEMS Microbiol Ecol 69:106–112
Dillon JG, Tatsumi CM, Tandingan PG, Castenholz RW (2002) Effect of environmental factors on the synthesis of scytonemin, a UV-screening pigment, in a cyanobacterium (Chroococcidiopsis sp.). Arch Microbiol 177:322–331
Domingos P, Rubim TK, Molica RJR, Azevedo SMFO, Carmichael WW (1999) First report of microcystin production by picoplanktic cyanobacteria isolated from a Northeast Brazilian drinking water supply. Environ Toxicol 14:13–35
Doolittle WF (1999) Phylogenetic classification and the universal tree. Science 284:2124–2129
Drakare S, Blomqvist P, Bergström AK, Jansson M (2003) Relationships between picophytoplankton and environmental variables in lakes along a gradient of water colour and nutrient content. Freshw Biol 48:729–740
Drews G, Prauser H, Uhlmann D (1961) Massenvorkommen von Synechococcus plancticus nov. spec., einer solitären, planktischen Cyanophyceae, in einem Abwasserteich. Betrag zur Kenntnis der sogenannten “μ-Algen”. Arch Mikrobiol 39:101–115
Dufresne A, Ostrowski M, Scanlan DJ, Garczarek L, Mazard S, Palenik BP, Paulsen IT, Tandeau de Marsac N, Wincker P, Dossat C, Ferriera S, Johnson J, Post AP, Hess WR, Partensky F (2008) Unraveling the genomic mosaic of a ubiquitous genus of marine cyanobacteria. Genome Biol 9:R90. doi:10.1186/gb-2008-9-5-r90
DuRand MD, Olson RJ, Chisholm SW (2001) Phytoplankton population dynamics at the Bermuda Atlantic Time-series station in the Sargasso Sea. Deep Sea Res Part II 48:1983–2003
Ernst A (1991) Cyanobacterial picoplankton from Lake Constance I. solation by fluorescence characteristics. J Plankton Res 13:1307–1312
Ernst A, Postius C, Böger P (1996) Glycosylated surface proteins reflect genetic diversity among Synechococcus spp. of Lake Constance. Arch Hydrobiol 48:1–6
Ernst A, Becker S, Hennes K, Postius C (1999) Is there a succession in the autotrophic picoplankton of temperate zone lakes? In: Bell CR, Brylinski M, Johnson-Green P (eds) Microbial biosystems: new frontiers. Proceedings of the 8th international symposium on microbial ecology. Atlantic Canada Society for Microbial Ecology, Halifax, Canada, pp 623–629
Ernst A, Becker S, Wollenzien VIA, Postius C (2003) Ecosystem dependent adaptive radiations of picocyanobacteria inferred from 16S rRNA and ITS-1 sequence analysis. Microbiology 149:217–228
Everroad RC, Wood AM (2006) Comparative molecular evolution of newly discovered picocyanobacterial strains reveals a phylogenetically informative variable region of beta-phycoerythrin. J Phycol 42:1300–1311
Fahnenstiel GL, Carrick HJ (1992) Phototrophic picoplankton in lakes Huron and Michigan: abundance, distribution, composition and contribution to biomass and production. Can J Fish Aquat Sci 49:379–388
Fahnenstiel GL, Patton TR, Carrick HJ, McCormick MJ (1991) Diel division cycle and growth rates of Synechococcus in lakes Huron and Michigan. Int Rev Ges Hydrobiol 76:657–664
Foster RA, Collier JL, Carpenter EJ (2006) Reverse transcription PCR amplification of cyanobacterial symbiont 16S rRNA sequences from single non-photosynthetic eukaryotic marine planktonic host cells. J Phycol 42:243–250
Frias-Lopez J, Thompson A, Waldbauer J, Chisholm S (2009) Use of stable isotope-labelled cells to identify active grazers of picocyanobacteria in ocean surface waters. Environ Microbiol 11:512–525
Fu FX, Warner ME, Zhang Y, Feng Y, Hutchins DA (2007) Effects of increased temperature and CO2 on photosynthesis, growth, and elemental ratios in marine Synechococcus and Prochlorococcus (Cyanobacteria). J Phycol 43:485–496
Fuller NJ, Tarran GA, Yallop M, Orcutt KM, Scanlan DJ (2006) Molecular analysis of picocyanobacterial community structure along an Arabian sea transect reveals distinct spatial separation of lineages. Limnol Oceanogr 51:2515–2526
Furnas M, Crosbie ND (1999) In situ growth dynamics of the photosynthetic prokaryotic picoplankters Synechococcus and Prochlorococcus. Bull Inst Oceanogr Monaco N Spec 19:387–417
Furtado ALFF, Calijuri MDC, Lorenzi AS, Honda RY, Genuario DB, Fiore MF (2009) Morphological and molecular characterization of cyanobacteria from a Brazilian facultative wastewater stabilization pond and evaluation of microcystis production. Hydrobiologia 627:195–209
Gaedke U, Weisse T (1998) Seasonal and interannual variability of picocyanobacteria in Lake Costance. Arch Hydrobiol Spec Issues Adv Limnol 53:143–158
Gallager SM, Waterbury JB, Stoecker DK (1994) Efficient grazing and utilization of the marine cyanobacterium Synechococcus by larvae of the bivalve Mercenaria mercenaria. Mar Biol 119:251–259
Garcia-Pichel F (1994) A model for internal self-shading in planktonic organisms and its implications for the usefulness of ultraviolet sunscreens. Limnol Oceanogr 39:1704–1717
Garcia-Pichel F, Castenholz RW (1991) Characterization and biological implications of scytonemin, a cyanobacteria sheath pigment. J Phycol 27:395–409
Garcia-Pichel F, Castenholz RW (1993) Occurrence of UV-absorbing, mycosporine-like compounds among cyanobacterial isolates and an estimate of their screening capacity. Appl Environ Microbiol 59:163–169
Geitler L (1932) Cyanophyceae. In: Rabenhorst L (ed) Kryptogamen-flora, vol 14. Akademische Verlagsgesellschaft, Leipzig, 1069 pp
Gervais F, Padisák J, Koschel R (1997) Do light quality and low nutrient concentration favour picocyanobacteria below the thermocline of the oligotrophic Lake Stechlin? J Plankton Res 19:771
Gismervik I (2006) Top-down impact by copepods on ciliate numbers and persistence depends on copepod and ciliate species composition. J Plankton Res 28:499–507
Glibert PM, Ray RT (1990) Different patterns of growth and nitrogen uptake in two clones of marine Synechococcus spp. Mar Biol 107:273–280
Glover HE, Phinney DA, Yentsch CS (1985) Photosynthetic characteristics of picoplankton compared with those of larger phytoplankton populations in various water masses in the Gulf of Maine. Biol Oceanogr 3:223–248
Gophen M, Geller W (1984) Filter mesh size and food particle uptake by Daphnia. Oecologia 64:408–412
Gouvea AP, Boyer GL, Twiss MR (2008) Influeance of ultraviolet radiation, copper, and zinc on microcystin content in Microcystis aeruginosa (Cyanobacteria). Harmful Algae 7:194–205
Grossman AR, Schaefer MR, Chiang GG, Collier JL (1993) The phycobilisome, a light-harvesting complex responsive to environmental conditions. Microbiol Rev 57:725–749
Häder DP, Kumar HD, Smith RC, Worrest RC (2007) Effects of solar UV radiation on aquatic ecosystems and interactions with climate change. Photochem Photobiol Sci 6:267–285
Harris GP (1980) Temporal and spatial scales in phytoplankton ecology. Mechanisms, methods, models and management. Can J Fish Aquat Sci 37:877–900
Harrison JW, Smith REH (2009) Effects of ultraviolet radiation on the productivity and composition of freshwater phytoplankton communities. Photochem Photobiol Sci 8:1218–1232
Hauschild CA, McMurter HJG, Pick FR (1991) Effect of spectral quality on growth and pigmentation of picocyanobacteria. J Phycol 27:698–702
Havens KE, Heath RT (1991) Increased transparency due to changes in the algal size spectrum during experimental acidification in mesocosms. J Plankton Res 13:673–679
Haverkamp T, Acinas SG, Doeleman M, Stomp M, Huisman J, Stal LJ (2008) Diversity and phylogeny of Baltic Sea picocyanobacteria inferred from their ITS and phycobiliprotein operons. Environ Microbiol 10:174–188
Haverkamp THA, Schouten D, Doeleman M, Wollenzien U, Huisman J, Stal LJ (2009) Colorful microdiversity of Synechococcus strains (picocyanobacteria) isolated from the Baltic Sea. ISME J 3:397–408
Hawley GRW, Whitton BA (1991a) Survey of algal picoplankton from lakes in five continents. Verh Int Ver Limnol 24:1220–1222
Hawley GRW, Whitton BA (1991b) Seasonal changes in chlorophyll-containing picoplankton populations of ten lakes in Northern England. Int Rev Ges Hydrobiol 76:545–554
Helbling EW, Villafañe VE, Barbieri ES (2001) Sensitivity of winter phytoplankton communities from Andean lakes to artificial ultraviolet-B radiation. Rev Chil Hist Nat 74:273–282
Helbling EW, Farías ME, Fernández Zenoff MV, Villafañe VE (2006) In situ responses of phytoplankton from the subtropical Lake La Angostura (Tucumán, Argentina) in relation to solar ultraviolet radiation exposure and mixing conditions. Hydrobiologia 559:123–134
Heldal M, Scanlan DJ, Norlans S, Thingstad F, Mann NH (2003) Elemental composition of single cells of various strains of marine Prochlorococcus and Synechococcus using X-ray microanalysis. Limnol Oceanogr 48:1731–1743
Hickel B (1985) Cyanonephron styloides gen. et sp. nov., a new chroococcal blue-green alga (Cyanophyta) from a brackish lake. Arch Hydrobiol Suppl 71; Algol Stud 38/39:99–104
Hickel B (1991) Two new chroococcal cyanophytes from a brackish environment, (Schlei-Fjord) Germany. Algol Stud 64:97–104
Hindák F (1975) Einige neue und interessante Planktonblaualgen aus der Westslowakei. Arch Hydrobiol Suppl 46(4); Algol Stud 13:330–353
Hindák F (1982) On some planktonic coccoid blue-green algae characteristic by Fe-precipitates. Arch Hydrobiol Suppl 63(3); Algol Stud 32:241–258
Hindák F (1985) The cyanophycean genus Lemmermanniella Geitler 1942. Arch Hydrobiol Suppl 71(3); Algol Stud 40:393–401
Honda D, Yokota A, Sugiyama J (1999) Detection of seven major evolutionary lineages in cyanobacteria based on the 16S rRNA gene sequence analysis with new sequences of five marine Synechococcus strains. J Mol Evol 48:723–739
Hopkinson BM, Morel FMM (2009) The role of siderophores in iron acquisition by photosynthetic marine microorganisms. Biometals 22:656–669
Houlahan JE, Currie DJ, Cottenie K, Cumming GS, Ernest SKM, Findlay CS, Fuhlendorf SD, Gaedke U, Legendre P, Magnuson JJ, McArdle BH, Muldavin EH, Noble D, Russell R, Stevens RD, Willis TJ, Woiwod IP, Wondzell SM (2007) Compensatory dynamics are rare in natural ecological communities. Proc Natl Acad Sci USA 104:3273–3277
Ilikchyan IN, McKay RML, Zehr JP, Dyhrman ST, Bullerjahn GS (2009) Detection and expression of the phosphonate transporter gene PHND in marine and freshwater picocyanobacteria. Environ Microbiol 11:1314–1324
Iturriaga R, Mitchell BG (1986) Chroococcoid cyanobacteria: a significant component in the food web dynamics of the open ocean. Mar Ecol Prog Ser 28:291–297
Ivanikova NV, Popels LC, McKay RML, Bullerjahn GS (2007) Lake Superior supports novel clusters of cyanobacterial picoplankton. Appl Environ Microbiol 73:4055–4065
Izaguirre I, Allende L, Marinone MC (2003) Comparative study of the planktonic communities of three lakes of contrasting trophic status at Hope Bay (Antarctic Peninsula). J Plankton Res 25:1079–1097
Jacquet S, Partensky F, Lennon JF, Vaulot D (2001) Diel patterns of growth and division in marine picoplankton in culture. J Phycol 37:357–369
Jacquet S, Domaizon I, Personnic S, Sime-Ngando T (2007) Do small grazers influence viral-induced bacterial mortality in Lake Bourget? Fund Appl Limnol 170:125–132
Jansson M, Olsson H, Pettersson K (1988) Phosphatases: origin, characteristic and function in lakes. Hydrobiologia 170:157–175
Jasser I (1997) The dynamics and importance of picoplankton in shallow, dystrophic lake in comparison with surface waters of two deep lakes with contrasting trophic status. Hydrobiologia 342–343:87–93
Jasser I, Arvola L (2003) Potential effects of abiotic factors on the abundance of autotrophic picoplankton in four boreal lakes. J Plankton Res 25:873–883
Jasser I, Królicka A, Karnkowska-Ishikawa A (2011) A novel phylogenetic clade of picocyanobacteria from the Mazurian lakes (Poland) reflects the early ontogeny of glacial lakes. FEMS Microbiol Ecol 75:89–98
Jezberová J, Komárková J (2007) Morphological transformation in a freshwater Cyanobium sp. induced by grazers. Environ Microbiol 9:1858–1862
Jochem FJ (2000) Probing the physiological state of phytoplankton at the single-cell level. Sci Mar 64:183–195
Joosten AMT (2006) Flora of the blue-green algae of the Netherlands. KNNV Publishing, Utrecht, p 239
Jürgens K, Jeppesen E (2000) The impact of metazooplankton on the structure of the microbial food web in a shallow, hypertrophic lake. J Plankton Res 22:1047–1070
Kana TM, Glibert PM (1987a) Effect of irradiances up to 2000 μE m−2 s−1 on marine Synechococcus WH7803-I. Growth, pigmentation, and cell composition. Deep Sea Res 34:479–495
Kana TM, Glibert PM (1987b) Effect of irradiances up to 2000 μE m−2 s−1 on marine Synechococcus WH7803 -II. Photosynthetic responses and mechanisms. Deep Sea Res 34:497–516
Karentz D, McEuen FS, Land MC, Dunlap WC (1991) Survey of mycosporine-like amino acid compounds in Antarctic marine organisms: potential protection from ultraviolet exposure. Mar Biol 108:157–166
Karl DM, Letelier R, Tupas L, Dore JE, Christian J, Hebel DV (1997) The role of nitrogen fixation in biogeochemical cycling in the subtropical North Pacific Ocean. Nature 388:533–538
Kasai F, Waiser MJ, Robarts RD, Arts MT (2001) Size dependent UVR sensitivity in Redberry lake phytoplankton communities. Ver Int Ver Limnol 27:2018–2023
Kasprzak P, Gervais F, Adrian R, Weiler W, Radke R, Jaeger I, Riest S, Siedel U, Schneider V, Boehme M, Eckmann R, Walz N (2000) Trophic characterization, pelagic food web structure and comparison of two mesotrophic lakes in Brandenburg (Germany). Int Rev Ges Hydrobiol 85:167–189
Katano T, Nakano S, Ueno H, Mitamura U, Anbutsu K, Kihira M, Satoh Y, Drucker V, Sugiyama M (2005) Abundance, growth and grazing loss rates of picophytoplankton in Barguzin Bay, Lake Baikal. Aquat Ecol 39:431–438
Katano T, Nakano S, Mitamura O, Yoshida H, Azumi H, Matsuura Y, Tanaka Y, Maezono H, Satoh Y, Satoh T, Sugiyama Y, Watanabe Y, Mimura T, Akagashi Y, Machida H, Drucker V, Tikhonova I, Belykh O, Fialkov VA, Han MS, Kang SH, Sugiyama M (2008) Abundance and pigment type composition of picocyanobacteria in Barguzin Bay, Lake Baikal. Limnology 9:105–114
Klut EM, Stockner JG (1991) Picoplankton associations in an ultra-oligotrophic lake on Vancouver Island, British Columbia. Can J Fish Aquat Sci 48:1092–1099
Koblížek M, Komenda J, Masojídek J, Pechar L (2000) Cell aggregation of the cyanobacterium Synechococcus elongatus: role of the electron transport chain. J Phycol 36:662–668
Komárek J (1958) Die taxonomische Revision der planktischen Blaualgen der Tschechoslowakei. In: Komárek J, Ettl H (eds) Algologische Studien. Naklad, ČSAV, Prague, pp 10–106
Komárek J (1976) Taxonomic review of the genera Synechocystis SAUV. 1892, Synechococcus Näg. 1849, and Cyanothece gen. nov. (Cyanophyceae). Arch Protistenk 118:119–179
Komárek J (1996) Towards a combined approach for the taxonomic and species delimitation of picoplanktic cyanoprokaryotes. Algol Stud 83:377–401
Komárek J, Anagnostidis K (1998) Cyanoprokaryota 1. Teil Chroococcales. Süsswasserflora von Mitteleuropas. Gustav Fischer, Stuttgart, 548 pp
Komárek J, Anagnostidis K (1999) Cyanoprokaryota 1. Teil Chroococcales. Süsswasserflora von Mitteleuropa 19/1. Gustav Fischer, Jena, 548 p
Komárek J, Kling H (1991) Variation in six planktonic cyanophyte genera in Lake Victoria (East Africa). Algol Stud 61:21–45
Komárek J, Komárková-Legnerová J (1992) Variability of some planktic gomphosphaeriod cyanoprocaryotes in northern lakes. Nord J Bot Sect Phycol 12:513–524
Komárek J, Azevedo SMFO, Domingos P, Komárková J, Tichý M (2001) Background of the Caruaru tragedy; a case taxonomic study of toxic cyanobacteria. Algol Stud 103:9–29
Komárek J, Cepák V, Kaštovský J, Sulek J (2004) What are the cyanobacterial genera Cyanothece and Cyanobacterium? Contribution to the combined molecular and phenotype taxonomic evaluation of cyanobacterial diversity. Algol Stud 113:1–36
Komárková J (2002) Cyanobacterial picoplankton and its colonial formations in two eutrophic canyon reservoirs (Czech Republic). Arch Hydrobiol 154:605–623
Komárková J, Cronberg G (1985) Lemmermanniella pallida (LEMM.) GEITL. from South-Swedish lakes. Arch Hydrobiol Suppl 71(3); Algol Stud 40:403–413
Komárková-Legnerová J, Cronberg G (1994) Planktic blue-green algae from lakes in South Scania Sweden Part I: Chroococcales. Algol Stud 72:13–51
Kranzler C, Lis H, Shaked Y, Keren N (2011) The role of reduction in iron uptake processes in a unicellular, planktonic cyanobacterium. Environ Microbiol 13:2990–2999
Lagerheim G (1883) Bidrag till Sveriges algflora. Öfv Kgl Vetensk-Akad Förhandl 40(2):37–39
Landry MR, Hassett RP (1982) Estimating the grazing impact of marine microzooplankton. Mar Biol 67:283–288
Landry MR, Kirshtein J, Constantinou J (1995) A refined dilution technique for measuring the community grazing impact of microzooplankton with experimental tests in the Central Equatorial Pacific. Mar Ecol Prog Ser 120:53–63
Laurion I, Vincent WF (1998) Cell size vs. taxonomic composition as determinants of UV sensitivity in natural phytoplankton communities. Limnol Oceanogr 43:1774–1779
Lavallée BF, Pick FR (2002) Picocyanobacteria abundance in relation to growth and loss rates in oligotrophic to mesotrophic lakes. Aquat Microb Ecol 27:37–46
Leibold MA, Holyoak M, Mouquet N, Amarasekare P, Chase JM, Hopes MF, Holt RD, Shurin JB, Law R, Tilman D, Loreau M, Gonzales A (2004) The metacommunity concept: a framework for multi-scale community ecology. Ecol Lett 7:601–613
Lemmermann E (1904) Das Plankton schwedischer Gewasser. Ark Bot 2:1–209
Li WKW (1998) Annual average abundance of heterotrophic bacteria and Synechococcus in surface ocean waters. Limnol Oceanogr 43:1743–1753
Liu H, Campbell L, Landry MR (1995) Growth and mortality rates of Prochlorococcus and Synechococcus measured with a selective inhibitor technique. Mar Ecol Prog Ser 116:277–287
Liu Z, Häder DP, Sommaruga R (2004) Occurrence of mycosporine-like aminoacids (MAAs) in the bloom-forming cyanobacterium Microcyctis aeruginosa. J Plankton Res 26:963–966
Liu X, Shi M, Liao Y, Gao Y, Zhang Z, Wen D, Wu W, An C (2006) Feeding characteristics of an amoeba (Lobosea: Naegleria) grazing upon cyanobacteria: food selection, ingestion and digestion process. Microb Ecol 51:315–325
Logue JB, Lindström ES (2008) Biogeography of bacterioplankton in inland waters. Freshw Rev 1:99–114
Mackey KRM, Paytan A, Grossman AR, Bailey S (2008) A photosynthetic strategy for coping in a high-light, low-nutrient environment. Limnol Oceanogr 53:900–913
Mackey KRM, Rivlin T, Grossman AR, Post AF, Paytan A (2009) Picophytoplankton responses to changing nutrient and light regimes during a bloom. Mar Biol 156:1535–1546
Maeda H, Kawai A, Tilzer MM (1992) The water bloom of cyanobacterial picoplankton in Lake Biwa, Japan. Hydrobiologia 248:93–103
Malinsky-Rushansky N, Berman T, Dubinsky Z (1995) Seasonal dynamics of picophytoplankton in Lake Kinneret, Israel. Freshw Biol 34:241–254
Mann KH (1993) Physical oceanography, food chains, and fish stocks: a review. ICES J Mar Sci 50:105–119
Mann NH (2003) Phages of marine cyanobacterial picophytoplankton. FEMS Microbiol Rev 27:17–34
Martin-Creuzburg D, Von Elert E (2006) Trophic upgrading of autotrophic picoplankton by the heterotrophic nanoflagellate Paraphysomonas sp. Limnol Oceanogr 51:1699–1707
Martin-Creuzburg D, Bec A, Von Elert E (2005) Trophic upgrading of picocyanobacterial carbon by ciliates for nutrition of Daphnia magna. Aquat Microb Ecol 41:271–280
Massana R, del Campo J, Dinter C, Sommaruga R (2007) Crash of a population of the marine heterotrophic flagellate Cafeteria roenbergensis by viral infection. Environ Microbiol 9:2660–2669
Mastala Z, Herodek S, V-Balogh K, Borbély G, Shafik HM, Vörös L (1996) Nutrient requirement and growth of a Synechococcus species isolated from Lake Balaton. Int Rev Ges Hydrobiol 81:503–512
Meyer B (1994) A new species of Cyanodictyon (Cyanophyceae, Chroococcales) planktic in eutrophic lakes. Algol Stud 75:183–188
Mills MM, Ridame C, Davey M, La Roche J (2004) Iron and phosphorus co-limit nitrogen fixation in the eastern tropical North Atlantic. Nature 429:292–294
Modenutti BE, Balseiro EG (2002) Mixotrophic ciliates in an Andean lake: dependence on light and prey of an Ophrydium naumanni population. Freshw Biol 47:121–128
Modenutti BE, Queimaliños C, Balseiro EG, Reissig M (2003) Impact of different zooplankton structures on the microbial food web of a South Andean oligotrophic lake. Acta Oecol 24:S289–S298
Moore LR, Ostrowski M, Scanlan DJ, Feren K, Sweetsir T (2005) Ecotypic variation in phosphorus acquisition mechanisms within marine picocyanobacteria. Aquat Microb Ecol 39:257–269
Morel A, Bricaud A (1981) Theoretical results concerning light absorption in a discrete medium, an application to specific absorption of phytoplankton. Deep Sea Res 28:1375–1393
Morris I, Glover HE (1981) Physiology of photosynthesis by marine coccoid cyanobacteria some ecological implications. Limnol Oceanogr 26:957–961
Moser M, Callieri C, Weisse T (2009) Photosynthetic and growth response of freshwater picocyanobacteria are strain-specific and sensitive to photoacclimation. J Plankton Res 31:349–357
Moutin T, Thingstad TR, Van Wambeke F, Marie D, Slawyk G, Raimbault P, Claustre H (2002) Does competition for nanomolar phosphate supply explain the predominance of the cyanobacterium Synechococcus ? Limnol Oceanogr 47:1562–1567
Mózes A, Présing M, Vörös L (2006) Seasonal dynamics of picocyanobacteria and picoeukaryotes in a large shallow lake (Lake Balaton, Hungary). Int Rev Ges Hydrobiol 91:38–50
Mühling M, Fuller NJ, Millard A, Somerfield PJ, Marie D, Wilson WH, Scanlan DJ, Post AF, Joint I, Mann NH (2005) Genetic diversity of marine Synechococcus and co-occurring cyanophage community: evidence for viral control of phytoplankton. Environ Microbiol 7:499–508
Murphy TP, Lean DRS, Nalewajko C (1976) Blue-green algae: their excretion of iron-selective chelators enables them to dominate other algae. Science 192:900–902
Nagata T, Takai K, Kawanobe K, Kim D, Nakazato R, Guselnikova N, Bondarenko N, Mologawaya O, Kostrnova T, Drucker V, Satoh Y, Watanabe Y (1994) Autotrophic picoplankton in southern Lake Baikal: abundance growth and grazing mortality during summer. J Plankton Res 16:945–959
Nagata T, Takay K, Kawabata K, Nakanishi M, Urabe J (1996) The trophic transfer via a picoplankton-flagellate-copepod food chain during a picocyanobacterial bloom in Lake Biwa. Arch Hydrobiol 137:145–160
Naumann E (1924) Sotvattnets plankton Stockholm, Sweden 267 pp
Nedoma J, Štrojsová A, Vrba J, Komárková J, Šimek K (2003) Extracellular phosphatase activity of natural plankton studied with ELF97 phosphate: fluorescence quantification and labelling kinetics. Environ Microbiol 5:462–472
Padisák J, Krienitz L, Koschel R, Nedoma J (1997) Deep-layer autotrophic picoplankton maximum in the oligotrophic Lake Stechlin, Germany: origin, activity, development and erosion. Eur J Phycol 32:403–416
Padisák J, Krienitz L, Scheffler W, Koschel R, Kristiansen J, Grigorszky I (1998) Phytoplankton succession in the oligotrophic Lake Stechlin (Germany) in 1994 and 1995. Hydrobiologia 369/370:179–197
Padisák J, Barbosa FAR, Koschel R, Krienitz L (2003) Deep layer cyanoprokaryota maxima are constitutional features of lakes: examples from temperate and tropical regions. Arch Hydrobiol Spec Issues Adv Limnol 58:175–199
Palenik B, Brahamsha B, Larimer FW, Land M, Hauser L, Chain P, Lamerdin J, Regala W, Allen EE, McCarren J, Paulsen I, Dufresne A, Partensky F, Webb EA, Waterbury J (2003) The genome of a motile marine Synechococcus. Nature 424:1037–1042
Passoni S, Callieri C (2000) Picocyanobacteria single forms, aggregates and microcolonies: survival strategy or species succession? Ver Int Ver Limnol 27:1879–1883
Pérez G, Quemaliños C, Modenutti B (2002) Light climate and plankton in the deep chlorophyll maxima in North Patagonian Andean lakes. J Plankton Res 24:591–599
Pernthaler J, Šimek K, Sattler B, Schwarzenbacher A, Bobkova J, Psenner R (1996) Short-term changes of protozoan control on autotrophic picoplankton in an oligo-mesotrophic lake. J Plankton Res 18:443–462
Personnic S, Domaizon I, Dorigo U, Berdjeb L, Jacquet S (2009a) Seasonal and spatial variability of virio-, bacterio-, and picophytoplanktonic abundances in three peri-alpine lakes. Hydrobiologia 627:99–116
Personnic S, Domaizon I, Sime-Ngando T, Jacquet S (2009b) Seasonal variations of microbial abundances and virus-versus flagellate-induced mortality of picoplankton in three peri-alpine lakes. J Plankton Res 31:1161–1177
Peštová D, Macek M, Martínez Pérez ME (2008) Ciliates and their picophytoplankton-feeding activity in a high-altitude warm-monomictic saline lake. Eur J Protistol 44:13–25
Pick FR (1991) The abundance and composition of freshwater picocyanobacteria in relation to light penetration. Limnol Oceanogr 36:1457–1462
Pick FR, Agbeti DM (1991) The seasonal dynamic and composition of photosynthetic picoplankton communities in temperate lakes in Ontario, Canada. Int Rev Ges Hydrobiol 76:565–580
Ploug H, Stolte W, Jørgensen BB (1999) Diffusive boundary layers of the colony-forming plankton alga Phaeocystis sp. – implications for nutrient uptake and cellular growth. Limnol Oceanogr 44:1959–1967
Porter KG (1973) Selective grazing and differential digestion of algae by zooplankton. Nature 244:179–180
Porter KG (1975) Viable gut passage of gelatinous green algae ingested by Daphnia. Ver Int Ver Limnol 19:2840–2850
Postius C, Böger P (1998) Different interactions of phycoerythrin- and phycocyanin-rich Synechococcus spp. with diazotrophic bacteria from the picoplankton of Lake Constance. Arch Hydrobiol 141:181–194
Postius C, Ernst A (1999) Mechanisms of dominance: coexistence of picocyanobacterial genotypes in a freshwater ecosystem. Arch Microbiol 172:69–75
Powell LM, Bowman JP, Skerratt JH, Franzmann PD, Burton HR (2005) Ecology of a novel Synechococcus clade occurring in dense populations in saline Antarctic lakes. Mar Ecol Prog Ser 291:65–80
Pradeep Ram AS, Sime-Ngando T (2008) Functional responses of prokaryotes and viruses to grazer effects and nutrient additions in freshwater microcosms. ISME J 2:498–509
Raven JA (1986) Physiological consequences of extremely small size for autotrophic organisms in the sea. In: Platt T, Li WKW (eds) Photosynthetic picoplankton. Department of Fisheries and Oceans, Ottawa. Can Bull Fish Aquat Sci 214:1–70
Reche I, Carrillo P, Cruz-Pizarro L (1997) Influence of metazooplankton on interactions of bacteria and phytoplankton in an oligotrophic lake. J Plankton Res 19:631–646
Reynolds CS, Huszar V, Kruk C, Naselli-Flores L, Melo S (2002) Towards a functional classification of the freshwater phytoplankton. J Plankton Res 24:417–428
Robertson BR, Tezuka N, Watanabe MM (2001) Phylogenetic analyses of Synechococcus strains (Cyanobacteria) using sequences of 16S rDNA and part of the phycocyanin operon reveal multiple evolutionary lines and reflect phycobilin content. Int J Syst Evol Microbiol 51:861–871
Rocap G, Distel DL, Waterbury JB, Chisholm SW (2002) Resolution of Prochlorococcus and Synechococcus ecotypes by using 16S-23S ribosomal DNA internal transcribed spacer sequences. Appl Environ Microbiol 68:1180–1191
Roff JC, Turner JT, Webber MK, Hopcroft RR (1995) Bacterivory by tropical copepod nauplii extent and possible significance. Aquat Microb Ecol 9:165–175
Ronneberger D (1998) Uptake of latex beads as size-model for food of planktonic rotifers. Hydrobiologia 387(388):445–449
Sánchez-Baracaldo P, Hayes PK, Blank CE (2005) Morphological and habitat evolution in the cyanobacteria using a compartmentalization approach. Geobiology 3:145–165
Sánchez-Baracaldo P, Handley BA, Hayes PK (2008) Picocyanobacterial community structure of freshwater lakes and the Baltic Sea revealed by phylogenetic analyses and clade-specific quantitative PCR. Microbiology 154:3347–3357
Sanders RW, Berninger UG, Lim EL, Kemp PF, Caron DA (2000) Heterotrophic and mixotrophic nanoplankton predation on picoplankton in the Sargasso Sea and Georges Bank. Mar Ecol Prog Ser 192:103–118
Sant’Anna CL, Azevedo MTP, Senna PAC, Komárková J, Komárková J (2004) Planktic cyanobacteria from Sao Paulo State, Brazil: Chroococcales. Rev Bras Bot 27(2):213–227
Scanlan DJ, West NJ (2002) Molecular ecology of the marine cyanobacterial genera Prochlorococcus and Synechococcus. FEMS Microbiol Ecol 40:1–12
Scanlan DJ, Ostrowski M, Mazard S, Dufresne A, Garczarek L, Hess WR, Post AF, Hagemann M, Paulsen I, Partensky R (2009) Ecological genomics of marine picocyanobacteria. Microbiol Mol Biol Rev 73:249–299
Schindler DW (1977) Evolution of phosphorus limitation in lakes: natural mechanisms compensate for deficiencies of nitrogen and carbon in eutrophied lakes. Science 195:260–262
Schindler DW (1990) Experimental perturbations of the whole lakes as tests of hypotheses concerning ecosystem structure and function. Oikos 57:25–41
Schindler DW (2006) Recent advantages in understanding and management of eutrophication. Limnol Oceanogr 51:351–355
Shannon S, Chrzanowski T, Grover J (2007) Prey food quality affects flagellate ingestion rates. Microb Ecol 53:66–73
Sherr EB, Sherr BF (1993) Protistan grazing rates via uptake of fluorescently labeled prey. In: Kemp P, Sherr B, Sherr E, Cole J (eds) Handbook of methods in aquatic microbial ecology. Lewis Publisher, Boca Raton, pp 695–701
Sherr BF, Sherr EB, Albright LJ (1987) Bacteria: link or sink? Science 235:88–89
Sherr EB, Sherr BF, Berman T, Hadas O (1991) High abundance of picoplankton-ingesting ciliates during late fall in Lake Kinneret Israel. J Plankton Res 13:789–799
Šimek K, Chrzanowski TH (1992) Direct and indirect evidence of size-selective grazing on pelagic bacteria by freshwater nanoflagellates. Appl Environ Microbiol 58:3715–3720
Šimek K, Bobkova J, Macek M, Nedoma J, Psenner R (1995) Ciliate grazing on picoplankton in a eutrophic reservoir during the summer phytoplankton maximum: a study at the species and community level. Limnol Oceanogr 40:1077–1090
Šimek K, Macek M, Pernthaler J, Straskrabová V, Psenner R (1996) Can freshwater planktonic ciliates survive on a diet of picoplankton? J Plankton Res 18:597–613
Sime-Ngando T (1995) Population dynamics of autotrophic picoplankton in relation to environmental factors in a productive lake. Aquat Sci 57:91–105
Simon RD (1987) Inclusion bodies in the cyanobacteria: cyanophycin, polyphospate, polyhedral bodies. In: Fay P, Van Baalen C (eds) The cyanobacteria. Elsevier, Amsterdam/New York/Oxford, pp 199–225, 543 pp
Sinha RP, Häder DP (2008) UV-protectants in cyanobacteria. Plant Sci 174:278–289
Six C, Thomas JC, Garczarek L, Ostrowski M, Dufresne A, Blot N, Scanlan DJ, Partensky F (2007) Diversity and evolution of phycobilisomes in marine Synechococcus spp.: a comparative genomics study. Genome Biol 8:R259
Skuja H (1932) Vorarbeiten zu einer Algenflora von Lettland. Bibl Phycol 26:1–302
Skuja H (1948) Taxonomie des Phytoplanktons einiger Seen in Uppland, Schweden. Symb Bot Upsal 9:1–399
Skuja H (1964) Grundzüge der Algenflora und Algenvegetation der Fjeldgegenden um Abisko in schwedisch-Lappland. Nova Acta Reg Soc Sci Upsal Ser 4 18(3):1–465
Sommaruga R (2009) Perspectives and research on environmental effects of ultraviolet radiation. Photochem Photobiol Sci 8:1217
Sommaruga R, Hofer JS, Alonso-Sáez L, Gasol JM (2005) Differential sunlight sensitivity of picophytoplankton from surface Mediterranean coastal waters. Appl Environ Microbiol 71:2154–2157
Sommaruga R, Chen Y, Liu Z (2009) Multiple strategies of bloom-forming Microcyctis to minimize damage by solar ultraviolet radiation in surface waters. Microb Ecol 57:667–674
Søndergaard M (1991) Phototrophic picoplankton in temperate lakes: seasonal abundance and importance along a trophic gradient. Int Rev Ges Hydrobiol 76:505–522
Staley JT (1997) Biodiversity: are microbial species threatened? Curr Opin Biotechnol 8:340–345
Stanier RY, Kuniswawa R, Mandel R, Cohen-Bazire G (1971) Purification and properties of unicellular blue-green algae (Order Chroococcales). Bacteriol Rev 35:171–205
Stockner JG (ed) (1991a) Autotrophic picoplankton in freshwater ecosystems. Int Rev Ges Hydrobiol 76:664 pp
Stockner JG (1991b) Autotrophic picoplankton in freshwater ecosystems: the view from the summit. Int Rev Ges Hydrobiol 76:483–492
Stockner JG (1998) Global warming, picocyanobacteria and fisheries decline: is there a connection? In: Atti del 12° Congresso AIOL, Vol.II, Genova, pp 29–37
Stockner JG, Antia NJ (1986) Algal picoplankton from marine and freshwater ecosystems: a multidisciplinary perspective. Can J Fish Aquat Sci 43:2472–2503
Stockner JG, Porter KG (1988) Microbial food webs in fresh-water planktonic ecosystems. In: Carpenter SR (ed) Complex interactions in lake communities. Springer, New York, pp 69–83, 283 pp
Stockner JG, Shortreed KS (1988) Response of Anabaena and Synechococcus to manipulation of nitrogen:phosphorus ratios in a lake fertilization experiment. Limnol Oceanogr 33(1348):1361
Stockner JG, Shortreed KS (1989) Algal picoplankton production and contribution to food webs in oligotrophic British Columbia lakes. Hydrobiologia 173:151–166
Stockner JG, Shortreed KS (1991) Phototrophic picoplankton: community composition abundance and distribution across a gradient of oligotrophic British Columbia and Yukon Territory lakes. Int Rev Ges Hydrobiol 76:581–601
Stockner JG, Shortreed KS (1994) Autotrophic picoplankton community dynamics in a pre-alpine lake in British Columbia, Canada. Hydrobiologia 274:133–142
Stockner J, Callieri C, Cronberg G (2000) Picoplankton and other non-bloom forming cyanobacteria in lakes. In: Whitton BA, Potts M (eds) The ecology of cyanobacteria: their diversity in time and space. Kluwer Academic Publishers, Dordrecht, pp 195–238, 688 pp
Stockner JG, Langston A, Sebastian D, Wilson G (2005) The limnology of Williston Reservoir: British Columbia’s largest lacustrine ecosystem. Water Qual Res J Can 40:28–50
Stomp M, Huisman J, de Jongh F, Veraart AJ, Gerla D, Rijkeboer M, Ibelings BW, Wollenzien UIA, Stal LJ (2004) Adaptive divergence in pigment composition promotes phytoplankton biodiversity. Nature 432:104–107
Stomp M, Huisman J, Vörös L, Pick FR, Laamanen M, Haverkamp T, Stal LJ (2007) Colorful coexistence of red and green picocyanobacteria in lakes and seas. Ecol Lett 10:290–298
Straškrabová V, Callieri C, Carrillo P, Cruz-Pizarro L, Fott J, Hartman P, Macek M, Medina-Sánchez JM, Nedoma J, Šimek K (1999a) Investigation on pelagic food web in mountain lakes – aims and methods. In: Straškrabová V, Callieri C, Fott J (eds) Pelagic food web in Mountain Lakes. MOuntain LAkes Research Program. J Limnol 58:77–87
Straškrabová V, Callieri C, Fott J (eds) (1999b) Pelagic food web in mountain lakes (Mountain Lakes Research Program). J Limnol 58:222 pp
Štrojsová A, Vrba J, Nedoma J, Komarková J, Znachor P (2003) Seasonal study of extracellular phosphatase expression in the phytoplankton of a eutrophic reservoir. Eur J Phycol 38:295–306
Sundt-Hansen LE, Olsen Y, Stibor H, Heldal M, Vadstein O (2006) Trophic cascades mediated by copepods, not nutrient supply rate, determine the development of picocyanobacteria. Aquat Microb Ecol 45:207–218
Suttle C (2000) Cyanophages and their role in the ecology of cyanobacteria. In: Whitton BA, Potts M (eds) The ecology of cyanobacteria: their diversity in time and space. Kluwer Academic Publishers, Dordrecht, pp 563–589, 668 pp
Szelag-Wasielewska E (2003) Phytoplankton community structure in non-stratified lakes of Pomerania (NW Poland). Hydrobiologia 506–509:229–236
Takano H, Arai T, Hirano M, Matsunaga T (1995) Effects of intensity and quality of light on phycocyanin production by a marine cyanobacterium Synechococcus sp. NKBG 042902. Appl Microbiol Biotechnol 43:1014–1018
Tarao M, Jezbera J, Hahn M (2009) Involvement of cell surface structures in size-independent grazing resistance of freshwater Actinobacteria. Appl Environ Microbiol 75:4720–4726
Tarbe AL, Unrein F, Stenuite S, Pirlot S, Sarmento H, Sinyinza D, Jean-Descy JP (2011) Protist herbivory: a key pathway in the pelagic food web of Lake Tanganyika. Microb Ecol 62:314–323
Taylor GT (1982) The role of pelagic heterotrophic protozoa in nutrient cycling: a review. Ann Inst Oceanogr 58:227–241
Timmermans KR, van der Wagt B, Veldhuis MJW, Maatman A, de Baar HJW (2005) Physiological responses of three species of marine pico-phytoplankton to ammonium, phosphate, iron and light limitation. J Sea Res 53:109–120
Toro M, Camacho A, Rochera C, Rico E, Bañón M, Fernandez-Valiente E, Marco E, Justel A, Avendañn MC, Ariosa Y, Vincent WF, Quesada A (2007) Limnological characteristics of freshwater ecosystems of Byers peninsula, Livingstone Island, in maritime Antarctica. Polar Biol 30:635–649
Tyrrell T (1999) The relative influences of nitrogen and phosphorus on oceanic primary production. Nature 400:525–531
Urbach E, Scanlan DJ, Distel DL, Waterbury JB, Chisholm SW (1998) Rapid diversification of marine picophytoplankton with dissimilar light-harvesting structure inferred from sequences of Prochlorococcus and Synechococcus (Cyanobacteria). J Mol Evol 46:188–201
Vadstein O (2000) Heterotrophic planktonic bacteria and cycling of phosphorus: phosphorus requirements, competitive ability, and food web interactions. In: Schink B (ed) Advances in microbial ecology, vol 16. Kluwer Academic Publisher, New York, pp 115–167
Van Donk E, Hessen DO (1993) Grazing resistance in nutrient-stressed phytoplankton. Oecologia 93:508–511
Van Donk E, Faafeng BA, De Lange HJ, Hessen DO (2001) Differential sensitivity to natural ultraviolet radiation among phytoplankton species in Arctic lakes (Spitsbergen, Norway). Plant Ecol 154:247–259
Van Gremberghe I, Van Wichelen J, Van der Gucht K, Vanormelingen P, D’hondt S, Boutte C, Wilmotte A, Vyverman W (2008) Covariation between zooplankton community composition and cyanobacterial community dynamics in Lake Blaarmeersen (Belgium). FEMS Microbiol Ecol 63:222–237
Van Mooy BAS, Rocap G, Fredericks HF, Evans CT, Devol AH (2006) Sulfolipids dramatically decrease phosphorus demand by picocyanobacteria in oligotrophic marine environments. Proc Natl Acad Sci USA 103:8607–8612
Van Mooy BAS, Fredericks HF, Pedler BE, Dyhrman ST, Karl DM, Koblížek M, Lomas MW, Mincer TJ, Moore LR, Moutin T, Rappé MR, Webb EA (2009) Phytoplankton in the ocean use non-phosphorus lipids in response to phosphorus scarsity. Nature 458:69–72
Vázquez-Domínguez E, Peters F, Gasol JM, Vaqué D (1999) Measuring the grazing losses of picoplankton: methodological improvements in the use of fluorescently labeled tracers combined with flow cytometry. Aquat Microb Ecol 20:119–128
Vázquez-Domínguez E, Duarte CM, Agustí S, Jürgens K, Vaqué D, Gasol JM (2008) Microbial plankton abundance and heterotrophic activity across the Central Atlantic Ocean. Prog Oceanogr 79:83–94
Veldhuis MJW, Admiral W (1987) Influence of phosphate depletion on the growth and colony formation of Phaeocystis pouchetii. Mar Biol 95:47–54
Villafañe VE, Andrade M, Lairana V, Zaratti F, Helbling EW (1999) Inhibition of phytoplankton photosynthesis by solar ultraviolet radiation: studies in Lake Titicaca, Bolivia. Freshw Biol 42:215–224
Villafañe VE, Sundbäck K, Figueroa FL, Helbling EW (2003) Photosynthesis in the aquatic environment as affected by UVR. In: Helbling EW, Zagarese H (eds) UV effects in aquatic organisms and ecosystems. Comprehensive series in photochemistry and photobiology. The Royal Society of Chemistry, Cambridge, pp 357–398
Villafañe VE, Marcoval MA, Helbling EW (2004) Photosynthesis versus irradiance characteristics in phytoplankton assemblages off Patagonia (Argentina): temporal variability and solar UVR effects. Mar Ecol Prog Ser 284:23–34
Vincent WF (2000) Cyanobacteria dominance in the polar region. In: Whitton BA, Potts M (eds) The ecology of cyanobacteria: their diversity in time and space. Kluwer Academic Publishers, Dordrecht, pp 321–340, 668 pp
Vincent WF, Bowman JP, Rankin LM, McMeekin TA (2000) Phylogenetic diversity of picocyanobacteria in Arctic and Antarctic ecosystems. In: Bell CR, Brylinsky M, Johnson-Green P (eds) Microbial biosystems: new frontiers. Atlantic Canada Society for Microbial Ecology, Halifax, pp 317–322
Vörös L, Callieri C, Balogh KV, Bertoni R (1998) Freshwater picocyanobacteria along trophic gradient and light quality range. Hydrobiologia 369(370):117–125
Vörös L, Mózes A, Somogyi B (2009) A five-year study of autotrophic winter picoplankton in Lake Balaton, Hungary. Aquat Ecol 43:727–734
Vrede K (1996) Regulation of bacterioplankton production and biomass in an oligotrophic clearwater lake – the importance of the phytoplankton community. J Plankton Res 18:1009–1032
Waterbury JB, Watson SW, Valois FW, Franks DG (1986) Biological and ecological characterisation of the marine unicellular cyanobacterium Synechococcus. Can Bull Fish Aquat Sci 214:17–120
Wehr JD (1993) Effects of experimental manipulations of light phosphorus supply on competition among picoplankton and nanoplankton in an oligotrophic lake. Can J Fish Aquat Sci 50:936–945
Weinbauer MG (2004) Ecology of prokaryotic viruses. FEMS Microbiol Rev 28:127–181
Weisse T (1990) Trophic interactions among heterotrophic microplankton, nanoplankton, and bacteria in Lake Constance (FRG). Hydrobiologia 191:111–122
Weisse T (1993) Dynamics of autotrophic picoplankton in marine and freshwater ecosystems. In: Jones JG (ed) Advances in microbial ecology, vol 13. Plenum Press, New York, pp 327–370
Weisse T, Kenter U (1991) Ecological characteristics of autotrophic picoplankton in a prealpine lake. Int Rev Ges Hydrobiol 76:493–504
Weisse T, Schweizer A (1991) Seasonal and interannual variation of autotrophic picoplankton in a large prealpine lake (Lake Constance). Verh Int Ver Limnol 24:821–825
West W, West GS (1894) On some algae from the West Indies. J Linn Soc 30(208):264–280
Whitton BA, Grainger SLJ, Hawley GRW, Simon JW (1991) Cell-bound and extracellular phosphatase activities of cyanobacterial isolates. Microb Ecol 21:85–98
Whitton BA, Al-Shehri AH, Ellwood NTW, Turner BL (2005) Ecological aspects of phosphatase activity in cyanobacteria, eukaryotic algae and bryophytes. In: Turner BL, Frossard E, Baldwin DS (eds) Organic phosphorus in the environment. Commonwealth Agricultural Bureau, Wallingford, pp 205–241, 399 pp
Wilhelm SW (1995) Ecology of iron-limited cyanobacteria: a review of physiological responses and implications for aquatic systems. Aquat Microb Ecol 9:295–303
Wilmotte A, Golubić S (1991) Morphological and genetic criteria in the taxonomy of Cyanophyta/Cyanobacteria. Algol Stud 64:1–24
Wilson DS (1992) Complex interactions in metacommunities, with implications for biodiversity and higher levels of selection. Ecology 73:1984–2000
Winder M (2009) Photosynthetic picoplankton dynamics in Lake Tahoe: temporal and spatial niche partitioning among prokaryotic and eukaryotic cells. J Plankton Res 31:1307–1320
Wood AM, Van Valen LM (1990) Paradox lost? On the release of energy-rich compounds by phytoplankton. Mar Microb Food Web 4:103–116
Wood AM, Horan PK, Muirhead K, Phinney DA, Yentsch CM, Waterbury JB (1985) Discrimination between types of pigments in marine Synechococcus spp. by scanning spectroscopy, epifluorescence microscopy and flow cytometry. Limnol Oceanogr 30:1303–1315
Yang Z, Kong F (2012) Formation of large colonies: a defense mechanism of Microcystis aeruginosa under continuous grazing pressure by flagellate Ochromonas sp. J Limnol 71:61–66
Yoshida T, Gurung TB, Kagami M, Urabe J (2001) Contrasting effects of cladoceran (Daphnia galeata) and calanoid copepod (Eodiaptomus japonicus) on algal and microbial plankton in a Japanese lake, Lake Biwa. Oecologia 129:602–610
Zaret M, Suffern KL (1976) Vertical migration in zooplankton as a predator avoidance mechanism. Limnol Oceanogr 21:804–813
Zeidner G, Bielawski JP, Shmoish M, Scanlan DJ, Sabehi G, Beja O (2005) Potential photosynthesis gene recombination between Prochlorococcus and Synechococcus via viral intermediates. Environ Microbiol 7:1505–1513
Zöllner E, Santer B, Boersma M, Hoppe HG, Jürgens K (2003) Cascading predation effects of Daphnia and copepods on microbial food web components. Freshw Biol 48:2174–2193
Zwirglmaier K, Heywood JL, Chamberlain K, Malcolm E, Woodward S, Zubkov MV, Scanlan DJ (2007) Basin-scale distribution patterns of picocyanobacterial lineages in the Atlantic Ocean. Environ Microbiol 9:1278–1290
Zwirglmaier K, Jardillier L, Ostrowski M, Mazard S, Garczarek L, Vaulot D, Not F, Massana R, Ulloa O, Scanlan DJ (2008) Global phylogeography of marine Synechococcus and Prochlorococcus reveals a distinct partitioning of lineages among oceanic biomes. Environ Microbiol 10:147–161
Zwirglmaier K, Spence E, Zybkov MV, Scanlan DJ, Mann NH (2009) Differential grazing of two heterotrophic nanoflagellates on marine Synechococcus strains. Environ Microbiol 11:1767–1776
Acknowledgements
The authors thank colleagues of the Microbial Ecology group of CNR-ISE Verbania, Italy, for their collaboration and strong support, and equally the Department of Fisheries and Oceans in British Columbia for supporting food chain research in lakes over the past three decades, including the special efforts of staff of Plankton Ecology Laboratory, West Vancouver, notably Ken Shortreed, Erland MacIsaac and Bruce Nidle. We also thank David Scanlan for valuable comments during preparation of the chapter, Roberto Bertoni for providing laboratory facilities in the microbial ecology laboratory at Verbania and Mario Contesini for technical assistance and field work on Lago Maggiore. A special thank to Brian Whitton for his untiring help to improve the manuscript.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2012 Springer Science+Business Media B.V.
About this chapter
Cite this chapter
Callieri, C., Cronberg, G., Stockner, J.G. (2012). Freshwater Picocyanobacteria: Single Cells, Microcolonies and Colonial Forms. In: Whitton, B. (eds) Ecology of Cyanobacteria II. Springer, Dordrecht. https://doi.org/10.1007/978-94-007-3855-3_8
Download citation
DOI: https://doi.org/10.1007/978-94-007-3855-3_8
Published:
Publisher Name: Springer, Dordrecht
Print ISBN: 978-94-007-3854-6
Online ISBN: 978-94-007-3855-3
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)