
Abstract
Soil salinization negatively impacts plant growth and soil structure, which leads to environmental stress and agricultural/economic losses. Improved plant growth during salt-induced ionic and osmotic plant stress is the key to successful phytoremediation of salt-impacted sites. Using plant growth-promoting rhizobacteria (PGPR) in PGPR-Enhanced Phytoremediation Systems (PEPS), positive effects of PGPR on plant biomass and health have been observed in greenhouse and field experiments. Revegetation is arguably the most important aspect of salt phytoremediation and substantial biomass increases occur in PGPR-treated plants in both sodic and saline soils. PGPR protect against inhibition of photosynthesis and plant membrane damage, which suggests that they confer tolerance to plants under salt stress. Using PEPS, decreases in soil salinity are observed due to uptake of sodium and chloride from the soil into foliar plant tissue. Although rates of uptake do not change due to PGPR inoculation, higher plant biomass due to PGPR enhancement of plant performance leads to greater salt uptake on a per area basis relative to that of untreated plants. Significant improvements in plant growth and commensurate sodium chloride uptake, and the results of mass balance studies used to assess the direct impact of ion uptake on actual observed changes in soil salinity, provide evidence that phytoremediation of salt-impacted soil is feasible within acceptable time frames using PEPS.
Access provided by CONRICYT-eBooks. Download chapter PDF
Similar content being viewed by others


Keywords
- Field trials
- NaCl
- PGPR-Enhanced Phytoremediation System(s) (PEPS)
- Polyamines
- Reactive oxygen species (ROS)
- Revegetation
- Salt remediation
ABA abscisic acid.
ACC 1-aminocyclopropane-1-carboxylic acid.
ACCD ACC deaminase.
Ca2+ calcium ion.
Chl a chlorophyll a.
Cl− chloride ion.
CT composite tailings.
ECe electrical conductivity of a soil-saturated paste extract.
F 0 minimal fluorescence.
F m maximal fluorescence.
F m′ maximal fluorescence in light-adapted tissue.
F s steady-state fluorescence.
F v/F m maximum quantum yield.
IAA indole acetic acid.
K+ potassium ion.
Mg2+ magnesium ion.
NaCl sodium chloride.
PAH polycyclic aromatic hydrocarbon.
PAM pulse amplitude modulated.
PEPS PGPR-Enhanced Phytoremediation Systems.
PGPR plant growth-promoting rhizobacteria.
PHC petroleum hydrocarbon(s).
PSII photosystem II.
qN non-photochemical quenching of fluorescence.
qP photochemical quenching of fluorescence.
ROS reactive oxygen species.
SAR sodium adsorption ratio.
SOS salt overly sensitive.
1 Introduction
1.1 Overview of Phytoremediation
Phytoremediation is a strategy whereby plants are used to extract, immobilize, contain and/or degrade soil contaminants. Although the term “phytoremediation” was not coined until the 1980s, the strategy has been employed for removing soil contaminants for at least 300 years [1, 2]. Rapid expansion occurred in this field in the 1990s, and phytoremediation has now become a useful strategy for on site and/or in situ removal of many contaminants, including petroleum hydrocarbons (PHC) , metals, radionucleotides, munitions waste (e.g., trinitrotoluene) and salt [1, 3,4,5]. Microbe-assisted phytoremediation, especially when used in conjunction with contaminant-tolerant plant species and high-level agronomic practices, can be a particularly effective green strategy for remediation and revegetation of impacted soils [6,7,8,9,10,11,12,13].
Plants have extensive rooting systems that can explore large volumes of soil to allow for effective remediation of various contaminants within different soil types. Typically, four types of phytoremediation processes for impacted soils are discussed in the literature [1, 4, 8]. During phytoremediation, contaminants can be broken down in the soil (e.g., rhizodegradation of PHC, also referred to as rhizoremediation ) or taken up by the roots and stored in plant tissue (typically in the foliage, as in phytoextraction of metals and salt). Some small molecules can be taken up by the roots, and the unmodified or modified forms are then transported via the transpiration stream to leaves, where they are released to the atmosphere via transpiration (e.g., phytovolatilization of trichloroethylene). Various contaminants can be bound within the rhizosphere (area immediately surrounding plant roots), making them less bioavailable (phytostabilization), and therefore less harmful to biota. In addition to these four main processes, plant roots can also alter soil chemistry via pH changes, which can further aid phytoremediation (e.g., breakdown of calcium carbonate in sodic soils provides calcium ions that can replace sodium ions at binding sites in the soil and allows for leaching or uptake of sodium) [14].
1.2 Prevalence and Sources of Salt-Impacted Soils
Soil salts can occur naturally (e.g., weathering of geologic formations, encroachment of seawater) or they can be released into the environment as a result of anthropogenic activities (e.g., irrigation, upstream oil and gas exploration/production, application of road salts) [15, 16]. Various environmental impacts associated with excess salt in soil include degradation of chemical and physical properties of the soil, diminished groundwater quality, and impaired plant growth. This results in substantial global agricultural and economic losses , sustenance issues for subsistence farmers, and ecosystem imbalances [17].
A significant buildup of salt often occurs in soils due to crop irrigation , and this has been suggested as “the first man-made environmental problem” [18, 19]. Irrigation waters tend to have high concentrations of calcium, magnesium, and sodium ions [16]. Use of this brackish water, particularly without adequate drainage management, results in the accumulation of salts in the rooting zone of plants due to evapotranspiration [17]. Calcium and magnesium tend to precipitate into carbonates, leaving sodium as the most prevalent ion in the soil, and this negatively impacts both plant growth and soil structure. Soil salinization affects 20% of irrigated land worldwide, which equates to an area approximately the size of France (62 million ha) [20, 21]. The resulting annual crop value losses have been estimated to be $27 billion (US) [17, 21]. Salinization, which occurs in virtually all geographic regions, has been a problem for millenia and continues to be a global concern of paramount importance: Soil salinity due to irrigation is thought to be a contributing factor to the downfall of the Sumerian civilization more than 4000 years ago, and irrigated land continues to be degraded by salt at a rate of 2000 ha/day [16, 22, 23].
Elevated salt levels in soils are as much of a problem for the upstream oil and gas industry as petroleum-impacted soils [24,25,26]. Most petroleum was formed from the remains of marine life that existed in ancient shallow seas. Consequently, oil deposits often occur in reservoirs that contain water with dissolved salts (brine), and the brine (which usually contains sulfates, bicarbonates, and chlorides of sodium, calcium, and magnesium) is frequently co-extracted with the oil [24, 27]. Any leakage into, or on, soils around the oil well will result in not only petroleum impacts, but also salt impacts. Furthermore, salt may be used during oil extraction . For example, sodium is often introduced during the extraction of bitumen from oil sands ore, and then winds up in the tailings. This is a major concern in the Athabasca oil sands region of Alberta, Canada where large volumes of fluid fine tailings are produced and stored in tailings ponds [25]. It was estimated that by the end of current upstream heavy oil operations, more than one billion cubic meters of fine tailings will be stored in these ponds. To reduce the stored volume, the composite tailings (CT) process is used, which involves the addition of gypsum or alum as a coagulant [28]. During this process, water containing high levels of salt is released from the CT, and this saline CT water makes reclamation of the CT deposit areas difficult. Efforts to revegetate the CT are hindered if salt from the CT water accumulates in the rooting zone.
Application of road salts (sodium chloride [NaCl], calcium chloride, potassium chloride and magnesium chloride), particularly in large urban areas, also leads to elevated soil salt levels in ecosystems adjacent to roads, snow removal dump sites, and some salt storage facilities [29, 30]. An average of 5 × 106 tonnes of road salts (primarily NaCl) are applied annually to Canadian roadways [29, 30]. This negatively impacts physical and chemical properties of surrounding soils, which consequently impacts associated biota. This problem was deemed critically important in the Canadian Environmental Protection Act, 1999, which categorized road salts as toxicants [30], and implemented new guidelines for their use (Code of Practice for Environmental Management of Road Salts) [29].
1.3 Soil Salt Chemistry
Based on a system developed by the US Salinity Laboratory [31], salt-impacted soils can be broadly classified as either saline (high concentration of soluble salts), sodic (high concentration of sodium), or saline-sodic (high concentrations of both soluble salts and sodium). More recently, the USDA Natural Resources Conservation Service classified salt-impacted soils into seven types that incorporate soil characteristics that are observable in the field as well as chemical analyses [32]. In this chapter, the US Salinity Laboratory classifications will be used when discussing soil salt impacts. Because NaCl is the most prevalent salt contaminant in the environment [33], the term “salt” refers to NaCl in subsequent sections of this chapter, unless specified otherwise.
One of the most common ways to measure total soluble soil salt concentration is electrical conductivity of a saturated soil-water paste extract (ECe, measured as dS/m) [31]. Soil sodicity can be calculated using the sodium adsorption ratio (SAR) . It is based on the ratio of sodium ions (Na+) to calcium ions (Ca2+) and magnesium ions (Mg2+) in the soil, and takes into account the difference in adsorption strengths of the ions to clay particles :
where the ionic concentrations are expressed in milliequivalents per liter in soil extract solution in equilibrium [31, 32]. Saline soils have an ECe > 4 dS/m and SAR < 13 in their saturation extract. Sodic soils have an ECe < 4 dS/m and SAR > 13. Saline-sodic soils have an ECe > 4 dS/m and SAR > 13.
Saline soils tend to have white crusts formed from crystallized salts that have precipitated at the soil surface. Sodic soils tend to have poor physical structure, low permeability (i.e., restricted movement of water and air through the soil), and high pH (7.8–8.5), all of which are detrimental to plant growth [32]. Poor structure is, in part, because Na+ displaces Ca2+ and Mg2+, which are important for holding clay lattices/particles together [34, 35]. In weakly aggregated soils, dispersion of soil particles can fill soil pores and impermeable surface crusts can form after repeated wet/dry cycles, which inhibit root penetration and growth. High pH limits the availability of some key plant nutrients and micronutrients such as phosphates, cobalt, copper, iron, manganese, and zinc, which are all more bioavailable at pH < 7. For more detailed descriptions of characteristics and chemistry of sodic and saline-sodic soils, please refer to Qadir et al. [36].
1.4 Responses of Plants to Salt Impacts
Plants are perhaps the most vulnerable sector of the biosphere to salt. Rapidly manifesting drought stress symptoms can occur in salt-impacted soils despite the presence of adequate water, because the resulting increase in osmotic pressure diminishes water uptake by plants [37, 38]. Over time, uptake of salt ions can lead to toxicity in plant tissues (particularly accumulation of Na+; as well, chloride ions [Cl−] can reach toxic levels in some sensitive species), and the presence of excess ions in the soil can interfere with nutrient availability (e.g., high concentrations of Na+ in the rhizosphere interfere with K+ uptake, due to the similar chemical nature of the ions, and this leads to K+ deficiency and growth inhibition in plants) [33, 37,38,39,40]. Salt stress negatively impacts germination, plant growth, and reproduction by affecting physiological processes such as photosynthesis, respiration, transpiration, and enzyme function; membrane properties are affected, upregulation of the stress-responsive hormones abscisic acid (ABA) and ethylene occurs, and reactive oxygen species (ROS) are generated [41,42,43,44,45].
1.4.1 Uptake and Transport of Na+, K+, and Cl−
During phytoextraction of salt, ions are taken up from the soil into plant tissues. Na+ and Cl− are taken up by plants primarily through passive symplastic pathways driven by concentration gradients and transpiration fluxes [46, 47]. Ions are transported from the root cells to the leaves via the transpirational stream of the xylem (Fig. 2.1) [33]. These ions are typically stored in the leaves, and little ion flow occurs via the phloem down to the roots (Fig. 2.1) [38, 47, 48]. Ion homeostasis, involving primarily Na+, K+, Ca2+ and Cl− is extremely complex, both at the cellular and whole plant level [38, 40, 48, 49]. Various ion channels and pumps in plant cells, many of them tissue-specific, regulate the flow of ions from the soil into roots, translocation from roots to foliar tissue, and storage within the cells or excretion from them [40, 50,51,52,53,54] (Fig. 2.2a). Phytoextracted salt can be removed from a site by harvesting the foliar tissues with accumulated salt ions.

Phytoextraction of salt. Uptake of ions from the soil to root epidermal cells occurs first. Ions are translocated via the root symplast to the xylem. Na+, Cl−, and other ions extracted from the soil are transported through the xylem to leaf tissue, and are stored in vacuoles. There is minimal flow of Na+ and Cl− back down the phloem

Ion pumps, channels, and signaling in plant salt stress and adaptation . (a) Numerous ion pumps and channels involved in salt stress and tolerance are shown. Not all of them are found in all species, or in all cells, and the specifics of ion conductance depend on a variety of conditions. ABA abscisic acid, ACC 1-aminocyclopropane-1-carboxylic acid, ANN annexin-formed channel, CAX cation/H+ exchanger, CCC cation-chloride-cotransporter, DA-NSCC depolarization-activated non-selective cation channel (NSCC), DAO diamine oxidase, FV fast vacuolar channel, H 2 O 2 hydrogen peroxide, HACC hyperpolarization-activated Ca2+ influx channel, HKT1 low-affinity Na+ histidine kinase transporter, HKT2 Na+/K+ histidine kinase symporter, KIRC K+ inward-rectifying, KORC K+ outward rectifying channel, ROSIC non-selective voltage-independent conductance, NHX Na+/H+ antiporter, NORC, •OH hydroxyl radical, PA polyamine, PAO polyamine oxidase, PEROX peroxiporin, Rboh respiratory burst oxidase homolog (an NADPH oxidase), ROS-NSCC ROS activated non-selective cation channel, SOS1 Na+/H+ antiporter, SOS2 protein kinase, SOS3 Ca2+ sensor, SV slow vacuolar channel, V-ATPase vacuolar H+-ATPase, VI-NSCC voltage-independent NSCC, VK K+-selective channel, VP1 vacuolar H+ pyrophosphatase. Hyperpolarization of the plasma membrane activates KIRC (more influx of K+ than N+). At the onset of salinity stress, KORC is activated by membrane depolarization, allowing the influx of Na+ and efflux of K+. Details for other ion channels and pumps can be found in Sects. 2.1.4.1 and 2.1.4.3, and references therein. (b) Some of the signaling pathways involved in salt stress and adaptation are outlined, with emphasis on PA/ROS-related pathways. Dotted lines with arrows indicate some relevant sources of the ionic and molecular pools, solid lines with arrows indicate positive regulatory actions, and lines with bars indicate negative regulatory actions. Salt stress leads to a Ca2+ burst that activates the SOS pathway. Increased Ca2+ is perceived by SOS3, which interacts with the kinase SOS2. This complex phosphorylates the SOS1 antiporter at the plasma membrane, which leads to diminished accumulation of Na+ in the cytosol. It also leads to increased activity of NHX at the tonoplast, which results in sequestration of excess Na+ in vacuoles. Further details for signaling pathways can be found in Sects. 2.1.4.2 and 2.1.4.3, and references therein
1.4.2 Salt Stress and ROS Damage
Salt stress (both osmotic and ionic) frequently results in an increase in ROS, including hydrogen peroxide, superoxide anion radicals, hydroxyl radicals, and singlet oxygen [55,56,57]. Formation of ROS occurs primarily in chloroplasts; however, it also occurs in mitochondria and peroxisomes [55, 56, 58]. During salt stress, cytosolic polyamines are exported to the apoplast, where they are oxidized to ROS [59]. Excessive formation of ROS leads to oxidative damage of many cellular components, including proteins, DNA, and lipids (e.g., membrane lipid peroxidation), ultimately leading to growth inhibition or capitulation of plants [15, 60].
1.4.3 Salt Stress and Acclimation Signaling Pathways
Although excessive salt-induced generation of ROS can impair metabolic processes, leading to oxidative stress and cellular damage, ROS can also signal responses to mitigate salt stress damage [56, 61,62,63]. Increases in antioxidant enzyme activities have been correlated with salt tolerance [57]. For example, the antioxidant enzymes catalase, superoxide dismutase, glutathione reductase, and glutathione peroxidase are activated in response to salinity stress in the European olive [64]. The ROS signaling pathways that result in acclimation to salt stress are integrated with numerous other signaling pathways related to salt tolerance. These include calcium, hormone and protein phosphorylation pathways, as well as complex interactions with polyamine pathways [56, 63].
Accumulation of polyamines is a key factor in achieving plant tolerance to salt stress [63]. Polyamines such as spermine, spermidine, and putrescine play a significant dual role in ROS homeostasis by acting both as ROS scavengers, and as substrates for amine oxidases in the apoplast that catalyze formation of ROS involved in stress response signaling [59, 63, 65]. Some of the ways by which polyamines influence ion transport during salt stress via complex signaling pathways are shown in Fig. 2.2b. For example, polyamines exported from the cytosol to the apoplast can block non-specific cation channels in the plasma membrane to limit Na+ influx and K+ efflux, and cytoplasmic polyamines can inhibit cation channels in the tonoplast to limit Na+ efflux to the cytoplasm, thereby helping to maintain proper cellular K+/Na+ ratios: this may be crucial for achieving salt stress tolerance [49, 63].
One well-researched signaling pathway for Na+ exclusion from cells was discovered using the salt overly sensitive (SOS) line in Arabidopsis [16, 48, 66, 67] (see Fig. 2.2). Following exposure to salt, an unidentified salt sensor in the root plasma membrane perceives the stress and a Ca2+ spike is generated in the cytoplasm. This activates a signal transduction cascade involving the SOS proteins: SOS3, a calcium binding protein, activates SOS2, a kinase that phosphorylates the plasma membrane antiporter, SOS1. Cytoplasmic Na+ is then transported out of root cells, either from the cytosol to the apoplast (epidermal cells), or from the cytosol to the xylem (parenchyma cells) [68]. In leaves, a similar pathway exists, but SOS3 is replaced by SCaBP8) [67]. SOS3 and SOS2 have been shown to play regulatory roles in salt tolerance [69].
Salt stress can result in increased levels of the ethylene precursor 1-aminocyclopropane-1-carboxylic acid (ACC) , resulting in stress ethylene production [39, 70]. This response is mediated by ABA, and ultimately leads to leaf abscission, ridding the plant of tissue that contains toxic levels of Na+ [56]. Upregulation of ABA also promotes stomatal closure to avoid water loss during osmotic stress, but may cause a shortage of CO2 for carbon fixation, which leads to a decline in photosynthesis [44, 55,56,57].
Signaling pathways involved in salt stress and subsequent acclimation are very complex (some of the signaling pathways are illustrated in Fig. 2.2b). The linear SOS pathway is the best understood, but it is not the only signaling pathway for adaptation to salt stress [67, 69]. There is good evidence that crosstalk between SOS and ABA signaling pathways occurs, and a complex signaling network with crosstalk between polyamine, ROS, and ABA pathways has also been shown [63, 67]. Plant growth-promoting rhizobacteria (PGPR) have been shown to positively influence many of these pathways and processes to mitigate salinity stress (see Sect. 2.1.5.3). Detailed descriptions of the numerous interconnected signaling pathways, and the salt ion channels, pumps and molecules involved in toxicity and tolerance are beyond the scope of this review. For further details, please see Blumwald [71], Gao et al. [72], Kronzucker and Britto [49], Kumar et al. [73], Kurusu et al. [74], Miller et al. [56], Pottosin and Shabala [65], Saha et al. [63], Uozumi and Schroeder [75], and Zhu [33].
1.4.4 Physiology of Salt Tolerance in Halophytes and Glycophytes
There are many different parameters that have been used to define halophytes in the literature [15, 40]. Generally, they can be defined as plants that grow well in salt-impacted soils. Plants that are not halophytes are frequently classified as glycophytes . A more realistic view is not a division into two broad categories of plants, but rather a continuum of salt tolerance ranging from extremely tolerant to extremely sensitive plants [76]. Many major agricultural crops are sensitive to salt stress [57]. Salt tolerance can be assessed in terms of survival (more meaningful for perennials than for annuals) and/or biomass production [77].
Halophytes can be obligate (absolute requirement for elevated salt habitats), facultative (can grow in salt-impacted soils, but optimum growth and health occurs in soils with low or no salt) or habitat-indifferent (can flourish in soil with or without salt) [15]. Glycophytes have varying sensitivities to salt, ranging from tolerant to completely intolerant. Depending on the circumstances, a given plant might be described as a facultative halophyte or a salt-tolerant glycophyte.
Halophytes have evolved different mechanisms that allow them to survive and thrive in salt-impacted soils: salt exclusion (minimizing uptake), salt accumulation, and salt excretion [78]. Some of these strategies are also employed by salt-tolerant glycophytes. Salt exclusion mechanisms are varied and complex; however, the main contributing factors are low permeability of root epidermal cell membranes, low net uptake of Na+ by root cortex cells, and tight control of xylem loading via the pericycle [15, 40, 51]. In salt accumulators, Na+ is taken up, transferred to leaf tissue and sequestered in vacuoles to minimize damage to cytoplasmic components (Figs. 2.1 and 2.2). Small organic osmolytes (compatible solutes), such as proline, betaine, and mannitol, accumulate in the cytoplasm to maintain osmotic balance within the cells, and some act as osmoprotectants to scavenge/quench ROS and prevent damage to membrane structure, enzymes, and proteins [33, 38, 55]. Salt excretion, prevalent in halophytes, is accomplished using leaf epidermal salt glands and hairs that remove salt from mesophyll cells via secretion at the leaf surface [14, 79].
1.5 Remediation and Phytoremediation of Salt
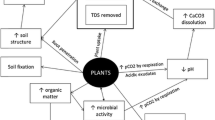
Remediation of salt-impacted soils has proven difficult and costly due to the absence of a versatile in situ technology [24]. Often the impacted soil must be removed to landfill and replaced with clean soil. In addition to the physical challenges encountered with ex situ remediation (soil excavation and soil replacement), these methods are also costly and unsightly. Various in situ treatments have also been employed to remediate salt-impacted soils. Three widely used methods are leaching, chemical amendments followed by leaching, and organic amendments [14, 24, 36]. For leaching, excess water is applied to the soil to move soluble salts from the surface soil to lower horizons. This can lower ECe values in surface and rooting zone soils, but not SAR, and is therefore effective primarily for saline soils [14]. Leaching results in valuable water resources being wasted, and diminishes soil stability and quality [14]. For sodic soils, numerous chemicals can be applied to the soil to promote ion exchange, often replacing Na+ with Ca2+ at the cation binding sites on clay particles. The amendments can be very costly, however, and leaching is required afterward to remove the Na+ to lower soil horizons [14]. Organic amendments can be used to increase dissolution of soil calcite and improve soil structure, however these amendments are also costly and dissolution is a slow process [14].
Several mechanisms are involved in salt remediation when using plants (phytoremediation). Uptake of salt ions into plant tissue results in a decrease in soil ECe and, possibly, SAR. Lowering soil pH via root exudates can increase the dissolution of soil calcium carbonate (calcite), thereby providing Ca2+ to displace the adsorbed Na+ in the soil. Displaced Na+ leads to improved soil structure with the resultant uptake and removal of Na+ from soil by plants. Root growth and the associated organic matter additions to the rhizosphere within impacted areas will increase hydraulic conductivity of the soil, which increases the potential for natural leaching of salt from upper to lower soil horizons [14, 36, 80, 81].
1.5.1 Advantages of Phytoremediation of Salt
Clearly, technologies are needed that can remediate salt-impacted soils in an environmentally responsible and cost-effective way. Phytoremediation has numerous advantages over conventional techniques for salt remediation. Some of the advantages are greater environmental stewardship (e.g., soil is treated and reused, not hauled to landfill for disposal), ease of application, and lower cost. Using plants, co-contaminants such as salt, PHC, and metals can be remediated simultaneously [82]. As an added benefit, some crops that are grown for phytoremediation can be sold for bioenergy sources, cellulose production, or livestock feed [14, 81].
1.5.2 Choosing Plants for Phytoremediation of Salt
Numerous plant species have been shown to effectively decrease ECe and SAR in salt-impacted soils [14, 81, 83, 84]. Plants chosen for phytoremediation must be sufficiently salt-tolerant to survive and grow in impacted soils. Some of the most salt-tolerant halophytes are very slow growing, and consequently these plants do not attain sufficient biomass to achieve phytoremediation in an acceptable time frame. Many halophytes excrete salt ions through specialized leaf glands, and others drop older organs that have accumulated toxic levels of Na+ and other ions: neither of these tolerance mechanisms leads to phytoremediation because the salt essentially returns to the soil near the plant. Similarly, plants that exclude Na+ and Cl− from roots are not efficient remediators of NaCl because the salt remains in the soil, although it might be more easily leached due to plant-related improvements in soil hydraulic conductivity.
A salt tolerance mechanism that is desirable for phytoremediation is uptake and storage of Na+ and Cl− into above-ground tissues. Maintaining low concentrations of cytoplasmic Na+ is a key factor in salt tolerance. As noted in Sect. 2.1.4.4, many halophytes and salt-tolerant glycophytes sequester Na+ to leaf vacuoles to achieve this [85, 86]. This prevents damage to cytosolic enzymes, and also counteracts the low extracellular osmotic potential resulting from salt stress [33]. Ideally, if soils are highly sodic, the chosen plant will have high Na+ uptake, but lower uptake of Ca2+ and Mg2+, which will lower SAR values [14]. Also, for effective phytoremediation, the plants chosen should be suitable for repeated harvesting of the foliar tissues containing phytoextracted salt.
1.5.3 PGPR-Enhanced Phytoremediation
One criterion that is essential for successful phytoremediation is substantial plant biomass production. Unfortunately, as discussed in Sect. 1.4, plants growing in salt-impacted soils are prone to the combined detrimental effects of water stress, ion toxicity, and nutritional deficiencies, which result in substandard plant growth. Traditional plant breeding programs and genetic engineering have been employed in attempts to improve salt tolerance in plants; however, the suite of genes and multiple pathways involved in salt tolerance, as well as the time involved to successfully breed or engineer salt tolerant plants, make this a daunting task [55, 87, 88].
One strategy that has been utilized to overcome the challenges of abiotic stressors is to employ plant growth-promoting rhizobacteria (PGPR) ; these soil microbes can promote growth and health in plants during stress conditions [8, 10, 89,90,91,92]. PGPR accelerate plant growth under stress conditions by increasing plant tolerance to elevated salt, PHC and/or trace metal levels, as well as other environmental stressors such as saturated soil or drought conditions. This leads to rapid growth of plants, including their roots. The vigorous plant growth that ensues leads to greater proliferation of naturally existing microbes in the soil, resulting in a very active rhizosphere that is typical of soils with normal plant growth. The substantial root biomass that accumulates in the soil provides a sink which allows for rapid partitioning of salt ions out of the soil, and their subsequent accumulation in the foliar tissues of some plants.
PGPR have been shown to confer salt tolerance in a variety of plants, by ameliorating both the osmotic and ion toxicity effects of salt stress [6, 58, 89, 93,94,95]. Some PGPR confer salt tolerance via tissue-specific regulation of HKT1, a plasma membrane Na+ uniporter [95] (see Fig. 2.2a). When plant growth inhibition is the result of stress ethylene production, PGPR with ACC deaminase (ACCD) can be employed [70]. ACCD metabolizes ACC, a precursor of ethylene in the biosynthetic pathway, thereby limiting the amount of stress ethylene that can be produced [6, 90]. Polyamines produced by PGPR have also been shown to lower stress ethylene levels and mitigate osmotic stress [96, 97]. PGPR have been shown to promote synthesis of antioxidants (including polyamines), and indole acetic acid (an auxin) which can promote root growth [6, 58, 89]. Recently, PGPR were shown to regulate a ROS-triggered caspase-like activity in rice; there was a concomitant decrease in programmed cell death, a phenomenon previously linked to caspase-like activity and salt-induced oxidative stress [58, 98]. Other mechanisms linking PGPR to salt tolerance in plants include altered mineral uptake, which results in a beneficial increase in the cellular K+/Na+ ratio; and elevated production of quorum sensing molecules, which can lead to alterations in the rhizosphere [92, 99,100,101].
1.5.4 Successful Remediation of Salt-Impacted Soils
Numerous studies have been conducted to assess phytoremediation of salt-impacted soils. For example, beet and millet were grown for 70 days in the greenhouse, in saline calcareous soil from Southern Ghor in Jordan [81]. Substantial amounts of Na+, K+, and Cl− were taken up into above-ground tissues, decreases in EC1:1 of 54–69% occurred, and better soil hydraulic conductivity was observed. Purslane has been shown to remove considerable amounts of NaCl from saline soils, and was recommended as an intercrop for salt removal in salt-sensitive fruit orchards based on pot experiments [102]. Hue et al. [82] used material dredged from Pearl Harbor, Hawaii. This material was amended with a soil from Oahu, Hawaii that was high in calcium and magnesium, to achieve a final ECe of ~18 dS/m. After growing a combination of two salt-tolerant grasses and a legume for 3 months in a greenhouse, soil ECe decreased by ~50%. This was attributed primarily to Na+ uptake by the legume and one of the grasses. Atriplex halimus plants were grown for 90 days in pot experiments using saline and saline-sodic soils from the Ninavah province of Iraq [103]. Decreases in EC were observed in both saline and saline-sodic soils (21 and 32%, respectively). Decreases in SAR were also observed for both saline and saline-sodic soils (29 and 50%, respectively).
Field experiments were performed in the Khorezm Region of Uzbekistan [104], where Chenopodium album and Apocynum lancifolium were grown in soil with ECe values of ~10.5 and 13, respectively (top 15 cm). Uptake of Cl− and Na+ in C. album (105 and 34 mg/g dry weight, respectively) was substantially higher than that of A. lancifolium (49 and 12 mg/g dry weight, respectively). Despite a root depth that was less than half that of A. lancifolium, C. album had much greater salt uptake from the soil (570 kg/ha, compared to 130 kg/ha for A. lancifolium). Thus, C. album was deemed a good choice for remediation of salt-impacted soils, and was recommended for integration into crop rotation programs, whereas A. lancifolium was deemed a salt-tolerant species not suitable for salt remediation because, relative to C. album, the rate of salt removal was deemed too low. For more results of phytoremediation experiments in the greenhouse and field, please see the following reviews of the literature [8, 14, 81, 85].
1.5.5 Obstacles Affecting Phytoremediation of Salt-Impacted Soils in the Field
Efforts to translate phytoremediation research from successful laboratory and greenhouse experiments to the field have proven challenging [9]. Although there have been many successful trials, there have also been numerous inconclusive and unsuccessful attempts at phytoremediation of salt in the field. A few general problems have emerged: different experimental conditions between the laboratory and the field, difficulty in accurately assessing salt remediation, and length of time required for salt remediation.
Numerous biotic and abiotic plant stress factors not present in laboratory and greenhouse studies can result in significant problems in field applications of phytoremediation. These include, but are not limited to, variations in temperature, nutrients, and precipitation; herbivory (insects and/or animals); plant pathogens; and competition by weed species that are native to the area [105]. Further, in the greenhouse, soils are generally homogeneous; in the field, contaminant concentrations vary across any given site, resulting in “hot spots”. Factors such as root structure, soil structure, organic composition of the soil, soil pH, moisture content, and microbial activity also exhibit spatial variability at a given site, and can change over time [105, 106].
For salt, conventional means of assessing phytoremediation (e.g., decrease in ECe in soil over time) may not be adequate to show that salt impacts are actually decreasing, although in many cases active remediation may be occurring. Salt readily migrates from lower soil horizons into the rooting zone of plants (i.e., the area where phytoremediation takes place) due to evaporation and transpiration [107]. Thus, it can be difficult to assess remediation exclusively by measuring soil salt levels in upper horizons. Assessing ion uptake into plant tissues and calculating estimates of total salt uptake at a given site can provide an estimate of actual salt removal and remediation over time.
Another challenge to phytoremediation of salt in the field is the length of time required to fully remediate the impacted soils [36]. Although this cannot be considered a failure of the technology, it is a disadvantage compared with traditional methods such as excavation and soil removal . It has also been suggested that salt remediation rates decrease over time, because in terms of mass balance, fewer salt ions are removed from the soil when salt gets diluted in leaching water (natural or applied) [14]. If salt uptake is the predominant removal mechanism, this should not be a factor. In fact, the reverse should be true: as soil quality improves with each successive growing season, plant root and shoot biomass should increase, providing a greater sink for salt ions.
1.5.6 Revegetation as a Measure of Successful Phytoremediation of Salt
As noted in Sect. 2.1.5.5, salt readily migrates from lower horizons to upper horizons in the soil, and moves with water flow in general. When plants grow in soil, this upward migration of water and salt is enhanced. Thus, it can be problematic to accurately assess remediation based on soil salt levels. Unlike heavy metals, which can be highly toxic to humans and other animals at levels found in soils, NaCl is generally not considered hazardous. Therefore, in the case of salt, the essential goal of phytoremediation is to overcome plant salt stress. Ideally, plants that grow rapidly with high rates of salt uptake and accumulation (e.g., kallar grass and oats) can be used to achieve both revegetation and salt removal from the soil [80]; however, generic regulatory criteria that depend solely on diminishing soil ECe levels and SAR in impacted soils may be too stringent and unnecessary in some cases. Because soil salts (including NaCl) are generally not hazardous to humans and other animals, and plants are the most sensitive part of the biosphere, we propose that, rather than achieving mandated levels of ECe and SAR, sustained revegetation of an impacted site should be the goal of salt phytoremediation. In this case, achieving 75% sustainable plant productivity compared to reference sites should qualify as successful phytoremediation of salt. This is in accordance with the reclamation objectives of some Canadian and American regulatory bodies that seek to ensure a self-sustaining ecosystem devoid of long-term toxicity, and to establish equivalent land capability that existed prior to industrial activities [26, 108].
If revegetation is the goal, selection of plant species is important. Many halophytes that accumulate large quantities of salt on a per mass basis grow too slowly to provide sufficient biomass for revegetation within an acceptable timeframe. Also, they may not be native to the site being remediated. Plants that exclude salt by limiting uptake into the root, or plants that excrete the salt from aerial tissues, cannot effectively remove salt from the soil. However, if the goal is only revegetation, these species could be considered for use. In general, for revegetation, the goal should be to achieve aggressive plant growth with species native to the impacted site.
2 PGPR-Enhanced Phytoremediation Systems (PEPS)
To fill the need for a versatile, green, in situ technology for remediation of contaminated soils, PGPR-Enhanced Phytoremediation Systems (PEPS) have been developed [9, 10, 83, 84, 91, 109,110,111,112,113,114]. To achieve successful PGPR-enhanced phytoremediation, a skill set beyond being able to plant seeds is required. A fundamental understanding of soil science, contaminant chemistry, plant biology, soil microbiology, agriculture, forestry, and regulatory guidelines is crucial for application of this green technology . The key to successful remediation within an acceptable time frame is to achieve vigorous plant growth because large amounts of biomass are necessary for phytoremediation; however, this is generally difficult due to suboptimal soil conditions (e.g., low organic content and poor soil structure) at impacted sites. Phytoremediation is therefore facilitated by preparing high-quality seed beds and utilizing other agronomic practices. After plant growth is established, contaminant chemistry, including degradation and/or uptake of the contaminants from soil, must be monitored. The standard PEPS protocol includes inoculation of seeds with PGPR to accelerate plant growth under stress conditions (see Sect. 1.5.3), soil pre-treatment (tilling soils to achieve homogeneity, as well as fertilizing and adding other required amendments to the rooting zone of plants), and adequate monitoring of the site (including contaminant assessments).
Mixtures of grass species, including cereals, are most commonly used in PEPS. Most Poaceae species (grass family) are facultative halophytes (also described in the literature as salt-tolerant glycophytes): they can grow on salt-impacted soils, but plant growth and health will be negatively impacted relative to growth in soils with low or normal salt levels [15]. Specific grass species have been used because they have been shown to be salt-tolerant, they accumulate salt in foliar tissue which can be removed easily from impacted sites, and they produce substantial amounts of root biomass. Using more than one plant species (co-cropping) can enhance overall microbe-assisted phytoremediation because the unique characteristics and properties of each plant species may support different microbial communities in the rhizosphere, differentially penetrate the soil matrix, and have different temperature and moisture optima, which increase the overall odds of success in the field; co-cropping has also been shown to limit weed proliferation and herbivory [115].
The PGPR used in PEPS are non-pathogenic, non-genetically modified soil bacteria (usually pseudomonads) that are present in the soils under remediation [9, 112]. These strains are naturally occurring, and express ACCD. They also synthesize indoleacetic acid (IAA) , which promotes root cell growth of host plants [116]. They are sensitive to common antibiotics, do not grow at 37 °C (i.e., they cannot proliferate in the human body), and are all classified as Biosafety Level 1 (the safest possible designation). They are ubiquitous in nature, common to soils around the world, and pose no threat to humans, wildlife, or the environment. With PEPS, the PGPR are used only via a seed treatment, whereby the seeds are treated in a controlled environment. The plant roots of the treated seeds are thus inoculated with PGPR as they pass through the seed coat during germination. Notably, PGPR, including those used in PEPS, increase the number of root hairs in grass seedlings under stress, relative to plants without PGPR ([100], Greenberg et al. unpublished data). Root hairs contribute substantially to the surface area of roots, and most of the ion uptake (including Na+) occurs across the plasma membrane of the root hair epidermal cells [51]. Thus, PGPR-treated PEPS plants have a greater capacity for Na+ uptake from the soil than untreated plants, which generally corresponds to decreases in both soil salinity and sodicity.
A mixture of PGPR can be used if the right combination of mixed microbial strains can be found (e.g., [10, 58]). The rationale is that taxonomically different PGPR have different optimum pH, temperature, and moisture requirements for colonizing rhizospheres/roots; and different PGPR may have different modes of action for promoting plant growth that could be additive or synergistic in a microbial mix, further increasing the odds of successful phytoremediation in the field. Sometimes, however, it is preferable to use a single strain of PGPR to avoid antagonistic effects. Greenhouse experiments have been performed to ascertain whether or not different PGPR strains should be used independently (e.g., CMH3) or in combination (e.g., UW3 + UW4) [83, 117].
When possible, PEPS plant species are chosen that are native to the area in which phytoremediation is being undertaken. This eliminates the ecological risk associated with introducing a non-native species to an ecosystem and facilitates native habitat reconstruction/reclamation following remediation. Native PGPR are also used, whenever possible. For instance, PGPR that have been isolated from the site being remediated can be used. This provides multiple benefits: PGPR isolated from salt-contaminated soils are salt-tolerant, acclimated to the soil conditions in that area, and may be more competitive in situ than non-native bacteria [118].
2.1 Development, Proof, and Full-Scale Application of PEPS
In the initial stages of PEPS development, remediation of PHC (including large recalcitrant polycyclic aromatic hydrocarbons [PAHs] ), heavy metals (lead, copper, and cadmium) and a pesticide (DDT) were the focus of the research [91, 113, 114, 119, 120]. The original process involved proven agronomic techniques, and plant growth with PGPR [91, 113, 114], with both laboratory and small-scale field trials [9, 112]. A variety of monocot and dicot species were used in the initial plant growth and phytoremediation experiments. Seeds were treated with various naturally occurring, non-pathogenic Pseudomonas strains, both individually and in microbial mixes. Although phytoremediation was observed in the absence of PGPR treatments, enhanced remediation rates were observed with PGPR seed treatments. A summary of the development, proof, and full-scale application of PEPS for PHC remediation was published recently [10].
2.2 Adapting PEPS for Salt Remediation
As discussed in Sect. 2.1.2, soil salinization is as much of an environmental issue as soil contaminated with compounds such as PHC (including PAHs) and metals [24,25,26]. For this reason, PEPS research was expanded to include phytoremediation of salt. Laboratory, greenhouse, and field experiments were conducted, resulting in the adaptation of PEPS for salt remediation [83, 84, 109,110,111, 117, 121, 122].
2.2.1 Lab/Greenhouse Experiments
The effects of salt stress on plant growth, photosynthesis, and membrane integrity were assessed in a series of greenhouse and laboratory experiments [109, 117, 121, 122]. Soils with a range of salinity (ECe) and sodicity (SAR) values were obtained from sites in Saskatchewan, Canada. Three strains of PGPR, Pseudomonas sp. UW3 (GenBank Accession Number KF145175), Pseudomonas sp. UW4 (GenBank Accession Number CP003880), and Pseudomonas corrugata CMH3 (GenBank Accession Number KF041156), were used for seed treatment prior to phytoremediation to promote plant growth and increase tolerance to salt [83, 123, 124]. All these strains are naturally occurring, produce IAA and express ACCD, the enzyme that consumes the precursor to ethylene, a plant stress hormone. UW3 and UW4 were isolated from unimpacted Ontario soils during a previous research project. CMH3 was isolated from the rhizosphere of grasses grown on a highly saline soil (ECe value of 20–50 dS/m) at an upstream petroleum site in Saskatchewan. Details of PGPR isolation, analysis, and identification; ACCD and IAA assays; bacterial inoculation of seeds; greenhouse trials; fluorescence assays; and electrolyte leakage assays can be found in Chang et al. [83] and Greenberg et al. [109].
2.2.1.1 Effects of Salinity and PGPR on Plant Growth
Previously, a decrease in biomass for wheatgrass grown without PGPR for 90 days in saline soils (ECe = 30 dS/m) was reported [109]. Biomass decreases for barley and oats grown for 45 days in saline soils (ECe = 9 dS/m) were also reported previously [83]. Data for oats grown on saline (ECe = 14 dS/m) and sodic (SAR = 24) soils are provided here as other examples. A decrease in oat biomass due to salt stress was observed in the absence of PGPR seed treatments. Oats without PGPR had 40% lower shoot biomass (Fig. 2.3) and 50% lower root biomass [117] than control plants grown on unimpacted soils (ProMix™). PGPR (UW3 + UW4) completely alleviated the root and shoot growth inhibition caused by salinity.

Effects of PGPR treatment on plant growth in salt-impacted soil. (a) A representative photograph of oat growth after 20 days in sodic field soil from a site in Saskatchewan, Canada with low ECe (3.2 dS/m) and high SAR [24]. “Control” shows the baseline normal plant growth in ProMix™ (ECe < 2 dS/m) growth medium, “-PGPR” shows plants grown in saline soil without PGPR treatment, “UW3” shows plants that were grown from oat seeds treated with UW3 PGPR, “UW4” were treated with UW4 PGPR and “UW3 + 4” were treated with a mix of UW3 and UW4 PGPR. (b) A representative photograph of oat growth after 45 days in saline field soil with moderate ECe (14 dS/m) and SAR [11]. (c) Fresh weight (g) of oat shoot biomass after 45 days growth in moderately saline field soil (ECe = 14 dS/m, SAR = 11). The results are expressed as means ± SEM of four independent replicates (n = 4). Data were analyzed by one-way analysis of variance (ANOVA) and Dunnett’s test. * indicates statistical differences (P < 0.01) in biomass relative to untreated oats grown in saline soil
In fact, the shoot biomass of plants treated with PGPR exceeded that of the controls. UW3 + UW4 improved the fresh weight of oat shoots (Fig. 2.3) and roots [117] by ~100%, relative to untreated (−PGPR) plants after 20 days in sodic soil (ECe, 3.2 dS/m; SAR, 24) and 45 days in saline soil (ECe, 14 dS/m; SAR, 11). Under the more saline conditions in wheatgrass experiments, PGPR (UW3 + UW4, CMH3) ameliorated salt stress, but did not bring biomass levels back to those of control plants grown under non-saline conditions [109]. Notably, the growth promotion effect was much greater using a mix of UW3 and UW4 than using either UW3 or UW4 independently (Fig. 2.3a). When the kinetics of oat growth is examined, with and without PGPR, it can be seen that PGPR protected the seedlings, especially during emergence and early growth phases (Fig. 2.4). This allows the plants to become established in impacted soils. It has been suggested that protection of young leaves is crucial for salt tolerance, due to the dearth of vacuoles available for Na+ sequestration in these leaves, and the detrimental effects of Na+ on protein synthesis and other processes crucial to plant growth [32, 79].

Effects of PGPR on kinetics of plant growth in salt-impacted soil. Shoot biomass (FW) of oats ± PGPR in sodic field soil (ECe, 3.2 dS/m; SAR, 24) at 10, 20, and 30 days
The studies described in this section, and those described in Sect. 1.5.3, show that PGPR can improve plant growth on salt-impacted soils. This indicates that crops and other plants not considered salt-tolerant can grow on saline soils with PGPR inoculation. This also supports the concept that equivalent land use can be achieved with PEPS and that phytoremediation of salt-impacted land via revegetation is feasible.
2.2.1.2 Alleviation of Salt Inhibition of Photosynthesis in PGPR-Treated Plants
Numerous abiotic environmental stresses, including salinization, result in deleterious effects on photosynthesis in plants [16, 57, 91, 109, 125,126,127]. Inhibition of photosynthesis is a good measure of the physiological state of the plant; therefore, measurement of various photosynthetic parameters can be used as an indication of the extent to which plants are salt-stressed. Indeed, negative impacts on plant growth due to salt stress are often associated with a decrease in photosynthetic rate, possibly the result of a decrease in stomatal conductance and the ensuing decrease in CO2 uptake [44, 128,129,130,131]. Osmotic stress , which occurs rapidly following plant exposure to salt, results in a decrease in chloroplast volume and an increase in Na+ concentration in the cytosol and chloroplasts. This can lead to inhibition of the photosynthetic electron transport chain [57, 125].
Chlorophyll a (Chl a) fluorescence is a useful technique for assessing photosynthetic activity [125, 132]. Chl a fluorescence parameters obtained using pulse amplitude-modulated (PAM) fluorometry (e.g., F v/F m, yield, qP, and qN) can be used to assess the efficiency of photochemistry in plants and to study the effect of salinity on photosynthetic electron transport [133]. F m (maximal fluorescence of dark-adapted tissue) and F 0 (minimal fluorescence [background fluorescence]) can be used to calculate F v/F m ([F m−F 0]/F m) which indicates the maximum quantum yield of photosystem II (PSII) [133]. Optimal F v/F m values range from 0.79 to 0.83 for most plant species [134, 135]. Lower values indicate damage to the photosynthetic apparatus, and resultant plant stress. Yield of steady-state photosynthesis [(F m′−F s)/F m′] can be calculated from the maximal fluorescence in light-adapted tissue (F m′) and steady-state fluorescence (F s). Yield is a measurement of continuous photosynthesis (i.e., the amount of light absorbed by PSII chlorophyll that gets used in photochemical reactions) [136]. The parameter qP ([F m′−F s]/[F m′−F 0′]) is a measure of photochemical quenching, which indicates the proportion of open (or functional) PSII reaction centers [137,138,139,140]. Non-photochemical quenching of fluorescence, qN (1−[F m′−F 0]/[F m−F 0]), is related to the dissipation of energy as heat and indicates the extent of photoinhibition [133, 139, 141].
An example of salt effects on photosynthesis is given in Table 2.1. Oats, with and without PGPR treatment , planted on salt-impacted soil (ECe = 30 dS/m) and control soil (ProMix, ECe < 2 dS/m), were grown for 20 days [122]. Various photosynthetic parameters were measured using a PAM fluorometer using methods published previously [109, 142]. For plants without PGPR, all Chl a fluorescence parameters (F v/F m, yield, qP and qN) showed significant negative impacts due to growth on saline soil (note: qN rises under stress conditions, while the other parameters fall during stress) (Table 2.1). This suggests that the photosynthetic apparatus was damaged and photosynthesis was impaired [136, 143, 144]. These data are in agreement with our previously published results, and with those of numerous other researchers using oats and other plant species [61, 91, 109, 145, 146].
The negative impacts of salinity on overall photosynthesis were largely alleviated by PGPR treatment of plants (both UW3 + UW4 and CMH3): most Chl a fluorescence parameters (F v/F m, yield and qP) of the PGPR-treated plants had values that were similar to plants grown in control soil (Table 2.1). These results are consistent with the improvements in plant growth on salt-impacted soils that were observed in greenhouse experiments (Figs. 2.3 and 2.4). Similar relationships between growth promotion and photosynthetic capacity were reported in Brassica [130] and lettuce [44].
Oats are considered to have low tolerance to salt [147]. Despite the sensitivity of this species, treatment with PGPR alleviated photosynthetic stress. Thus, PGPR seed treatment can result in salt-sensitive species becoming more tolerant, thereby making them candidates for phytoremediation of salt and/or revegetation of salt-impacted soil.
2.2.1.3 Effects of Salinity and PGPR on Cell Membrane Integrity
Plant cell membranes play an important role in the maintenance of the micro-environment and metabolism of plant cells, and are often the first targets of abiotic plant stressors [41]. ROS-mediated membrane damage is a major cause of the cellular toxicity induced by salt stress in a variety of plants (see Sect. 2.1.4.2). Salt-induced ROS lead to damage to plant cell membranes and increase their permeability, allowing electrolytes that are contained within the membrane to leak into surrounding tissues [148]. Therefore, maintaining cell membrane stability and integrity is important for salt tolerance. The degree of damage to cell membranes can be estimated by measuring electrolyte leakage from cells, by comparing the electrical conductivity of the leaked contents (into water) from salt-stressed plant tissues to that of control plant tissues [109, 148, 149].
As an example, data on electrolyte leakage in oat leaves is shown in Fig. 2.5. The objective of the experiment was to assess cell membrane integrity following salt stress, and to determine whether PGPR could ameliorate the damage. Oat seeds, with and without PGPR treatment, were planted on moderately and highly impacted saline soils (ECe = 12 and 18 dS/m, respectively) and control soil (ProMix™, ECe < 2 dS/m) [122]. Shoots were removed from plants for electrolyte leakage analysis after 12 days of growth. Electrolyte leakage was measured as electrical conductivity (dS/m) of solutions containing ions that escaped from oat cells, presumably via damaged plasma membranes [109] (Fig. 2.5). The higher the EC (dS/m) value of the receiving water, the greater the damage to plant membranes.

Effects of salinity and PGPR on membrane damage in oats. Electrolyte leakage assays were performed on excised oat leaves from plants grown for 12 days on ProMix™ (ECe < 2 dS/m), moderately impacted saline soil (medium salt, ECe = 12 dS/m), or highly impacted saline soil (high salt, ECe = 18 dS/m) ± PGPR (UW3 + UW4). Results are expressed as means ± SEM of six independent replicates (n = 6). Data were analyzed by one-way analysis of variance (ANOVA) and post-hoc Bonferroni tests. * (P < 0.05) indicates a significant difference between values for “No PGPR” relative to PGPR-treated plants
Electrolyte leakage from plant tissues increased as soil salinity increased (Fig. 2.5), indicating that plant membrane damage increased with salinity level. The amount of electrolyte leakage was greatly diminished in PGPR-treated oats, indicating less damage to plasma membranes (Fig. 2.5). Similarly, Kang et al. [42] observed a protective effect of PGPR following induction of high salt stress: leaves of PGPR-treated cucumber had 21% less electrolyte leakage than control plants. The electrolyte leakage results indicate membrane damage due to salt stress, and are consistent with the photosynthesis results in the previous section (Alleviation of Salt Inhibition of Photosynthesis in PGPR-treated Plants). For instance, the lower yield and higher qN values in the absence of PGPR inoculation indicate loss of thylakoid membrane integrity relative to that in PGPR-treated plants.
2.2.2 Field Trials
The effects of salinity and PGPR on plant growth and salt uptake in field experiments were reported previously [83, 84, 110, 111, 117, 122]. Field trials were performed at upstream oil and gas sites with poor quality soils of varying soil salinities (ECe, 2–40 dS/m) and sodicities (SAR, 1–45) in Manitoba, Saskatchewan, Alberta, and the Northwest Territories, Canada. Three strains of PGPR (UW3 + UW4, CMH3) were used to treat various grass species (tall fescue, tall wheatgrass, ryegrass, barley, oats) prior to phytoremediation to promote plant growth and increase tolerance to salt. Field trials were conducted over a period of two or three consecutive growing seasons. Details of bacterial inoculation of seeds, field trials, and analyses for Na+ and Cl− in plant tissues can be found in previously published work [83, 84].
2.2.2.1 Effects of Salinity and PGPR on Plant Growth
Results for the effects of salinity and PGPR on plant growth in the field have been published previously [83, 84]. Effects on plant growth were similar to those observed in the greenhouse: increases in soil salinity led to decreased plant biomass production in the absence of PGPR treatment. Treatment of seeds with PGPR alleviated the plant stress such that root and shoot biomass and ground cover were comparable to control plants grown in non-saline/sodic soils. In general, on saline soils, shoot biomass increases of 100–200% were observed in PGPR-treated plants, relative to untreated plants.
2.2.2.2 Uptake of NaCl from Soil
The NaCl concentrations in above-ground tissue of barley and oats from nine upstream oil and gas sites in Saskatchewan were measured following a single growing season [83, 84]. On a per mass basis, above-ground plant NaCl concentrations ranged from 22 to 97 g/kg (DW). Generally, on a per mass basis, about 2–3 times more Cl− than Na+ was stored in above-ground plant tissues. Notably, NaCl accumulation in plant foliage was accompanied by decreases in soil salinity (10–20%) at the field sites. Results from the field were in agreement with results previously obtained in greenhouse experiments [83, 109]. Salt removal (kg/ha) by barley and oats was comparable to literature values for various glycophytes, and Na+ uptake was comparable to that of millet, another grass species [81].
Data from the Saskatchewan sites where PEPS was applied were compiled to obtain average values for typical salt remediation using this remedial strategy [84]. Standard PEPS experimental protocols were similar to those detailed in Chang et al. [83]. NaCl uptake into foliage averaged 29 g/kg (DW), with three times more Cl− than Na+ accumulation by weight in the foliage. An average of 150 kg/ha of NaCl was removed from the sites per harvest of above-ground biomass. A concomitant average annual decrease in ECe of 15% was observed when the salt was only in the top 30 cm of soil (i.e., the rooting zone).
PGPR treatment did not result in increased NaCl uptake on a plant biomass basis [83]. That is, the concentrations of salt in the foliage were similar with and without PGPR treatment. However, the increases in plant biomass due to PGPR treatment were substantial (generally 100–200%). This is in agreement with our results from greenhouse experiments [109] and the findings of other researchers. For instance, Jesus et al. [14] indicated that a search of the literature showed biomass increases as a result of PGPR inoculation, but there was not a reference that showed an increase in salt phytoextraction on a biomass basis in any plant as a result of PGPR treatment. Similarly, treatment of a perennial rhizome grass with a plant growth-promoting mycorrhizal fungus did not increase uptake of Na+ and Cl− from salt-impacted soils [150]. Because of the increases in biomass due to PGPR or fungal treatments, the net effect was a much higher rate of NaCl removal from the soil for inoculated plants than that of untreated plants [14, 150]. These results are in contrast with those of Ozawa et al. [86]. They found that inoculation of a glasswort (a halophyte from the Chenopodiaceae family that sequesters Na+ in valcuoles) with Pseudomonas pseudoalcaligenes did not increase fresh or dry weight of the glasswort shoots, but did increase Na+ accumulation relative to uninoculated plants. This difference may be due to dissimilar plant growth conditions in general, the plant species used (a succulent marine halophyte), or the PGPR (an endophytic nitrogen-fixing bacteria).
Little research has been done to determine the connection of ion uptake by plants to actual observed changes in soil salinity in full-scale phytoremediation trials of salt-impacted soils. This was investigated when PEPS was employed on a salt-impacted (ECe = 5.97 dS/m) upstream petroleum site in Saskatchewan, Canada [121]. Data from this field trial were used to conduct mass balance studies, to determine the efficacy of PEPS on saline soils. Plant tissue collected over two successive growing seasons was assayed for ionic content and these data were compared to measured changes in soil salinity (ECe) for each field season. Based on the amount of five predominant ions (Ca2+, Mg2+, K+, Na+, and Cl−) in the plant tissue samples, removal of these ions from soil was measured, and the expected change in soil EC was calculated. These values were used to determine how much of the observed change in soil salinity could be attributed to ion uptake by PEPS plants during a given field season.
Soil ECe decreased by 0.96 and 0.45 dS/m in the first and second year, respectively. The mass of salt ion uptake into plant biomass and total annual biomass were compared to the measured changes in soil salinity over the two field seasons. Taking into account the effect of each salt ion on the ionic strength of the soil solution, uptake of soil salt ions into foliar plant tissue accounted for 60.5 and 76.8% of the change in salinity in the first and second year, respectively. Notably, only five salt ions were included in the mass balance calculations; therefore, the change in soil salinity that was attributed directly to phytoremediation using PEPS was likely underestimated. This research provided evidence that, for PEPS field trials, the uptake of ions from the soil into plant biomass plays a predominant role in soil salinity decreases , and is not the result of water flux through the soil and movement of ions into deeper soil horizons.
In general, phytoremediation research in the greenhouse and the field has shown that salt concentrations in the foliage tend to be fairly similar on a per mass basis, independent of PGPR or fungal treatment, plant type, soil ECe, and SAR. However, the increases in plant growth due to PGPR or fungal treatment tend to be large (average shoot biomass increase of 150%), particularly in poor soils and those with moderate to severe salt impacts. The extra biomass due to PGPR treatment will translate to greater salt removal from the soil. We suggest that the key to salt phytoremediation is to maximize growth with PGPR treatment or other means. Greater plant biomass should result in higher rates of salt remediation each growing season when PEPS are employed.
2.3 Feasibility of Salt Phytoremediation Using PEPS
For salt remediation, PEPS are effective for several reasons: (1) The PGPR alleviate plant stress and promote growth by conferring salt tolerance to the plants, as well as conferring tolerance to potential co-contaminants such as PHC and metals. (2) The PGPR protect plants against other potential abiotic stressors (e.g., cold) that result in the production of stress ethylene and decreased rates of plant growth. (3) The large amount of root biomass produced in the soil allows for effective partitioning of NaCl out of the soil into the biosphere. (4) Foliar tissues of PEPS plants can be harvested, thereby removing accumulated salt from the site. Harvested vegetation will not have sufficiently high levels of salt ions to be considered high-salt waste. (5) PEPS are adapted to site-specific conditions (i.e., from the site in question), which increases the chance of successful remediation.
Since 2009, PEPS has been deployed for full-scale remediation of several salt-impacted sites in Manitoba, Saskatchewan, Alberta, and the Northwest Territories, Canada [111]. Remediation goals were met at eight of these sites, either by lowering soil salinity (ECe) levels to generic regulatory criteria, or by restoring plant growth and productivity to equivalent land use (i.e., equivalent growth and productivity to areas surrounding the site). We have observed that the ECe drops at a rate of approximately 15% per year when the salt is present only in the rooting zone [84]. The average amount of NaCl taken up into the leaves of PEPS grass plants is 29 g/kg (DW). An average of 150 kg/ha NaCl is removed from a field per crop harvest. Thus, soils with an ECe of 10–15 dS/m, spread to a depth of 0.5 m (approximate rooting zone of grasses used in PEPS) can be remediated in about 5 years [111]. We note that as remediation proceeds, and soil salt levels drop and the soils improve, the plants will grow better, which should lead to accelerated rates of revegetation and remediation. More biomass will be produced per growing season, and the levels of NaCl taken up by plants does not drop as the ECe decreases (see the section entitled “Uptake of NaCl from Soil”). Given that research has shown that we can successfully establish plant growth using PEPS on salt-impacted sites before soil salt levels drop to generic regulatory criteria, phytoremediation based on revegetation and equivalent land use will occur sooner than the 5 year estimate based on salt uptake and biomass calculations.
3 Conclusions
Research described in this chapter indicates that salt phytoremediation is feasible using PEPS and other systems. Rapid plant growth leads to revegetation of salt-impacted sites, typically in less than 5 years. The calculations for the estimated time required to remove NaCl from salt-impacted soils suggest that salt ions can be phytoextracted from soil at an acceptable rate, which will lead to unimpacted soil in the long term. Revegetation and removal of salt from impacted soils should accelerate in successive years of PEPS treatment as the soil improves, because this will facilitate increased plant growth over time, which in turn will provide a larger sink for soil salts. Finally, revegetation may be the most important aspect of salt phytoremediation, and in many cases can be considered a key measure of successful salt phytoremediation.
References
Kaushal J, Bhasin SK, Bhardwaj P (2015) Phytoremediation: a review focusing on phytoremediation mechanisms. Int J Res Chem Environ 5:1–9
Willey N. (2007) Preface. In: Willey N (ed) Methods in biotechnology. Phytoremediation: methods and reviews, vol 23. Humana Press, Totowa, NJ, p ν
Meagher RB (2000) Phytoremediation of toxic elemental and organic pollutants. Curr Opin Plant Biol 3:153–162
Pilon-Smits E (2005) Phytoremediation. Annu Rev Plant Biol 56:15–39
Salt DE, Smith RD, Raskin I (1998) Phytoremediation. Annu Rev Plant Phys Plant Mol Biol 49:643–668
Arora NK, Tewari S, Singh S, Lal N, Maheshwari DK (2012) PGPR for protection of plant health under saline conditions. In: Maheshwari DK (ed) Bacteria in agrobiology: stress management. Springer, Berlin, pp 239–258
Chaudhry Q, Blom-Zandstra M, Gupta S, Joner EJ (2005) Utilizing the synergy between plants and rhizosphere microorganisms to enhance breakdown of organic pollutants in the environment. Environ Sci Pollut Res 12:34–48
de Bashan LE, Hernandez J-P, Bashan Y (2012) The potential contribution of plant growth-promoting bacteria to reduce environmental degradation—a comprehensive evaluation. Appl Soil Ecol 61:171–189
Gerhardt KE, Huang X-D, Glick BR, Greenberg BM (2009) Phytoremediation of organic soil contaminants: potential and challenges. Plant Sci 176:20–30
Gerhardt KE, Gerwing PD, Huang X-D, Greenberg BM (2015) Microbe-assisted phytoremediation of petroleum impacted soil: a scientifically proven green technology. In: Fingas M (ed) Handbook of oil spill science and technology. Wiley, Hoboken, NJ, pp 407–427
Kuiper I, Lagendijk EL, Bloemberg GV, Lugtenberg BJJ (2004) Rhizoremediation: a beneficial plant-microbe interaction. Mol Plant Microbe Interact 17:6–15
Singer AC, Thompson IP, Bailey MJ (2004) The tritrophic trinity: a source of pollutant-degradaing enzymes and its implications for phytoremediation. Curr Opin Microbiol 7:239–244
Wenzel W (2009) Rhizosphere processes and management in plant-assisted bioremediation (phytoremediation) of soils. Plant Soil 321:385–408
Jesus JM, Danko AS, Fiúza A, Borges M-T (2015) Phytoremediation of salt-affected soils: a review of processes, applicability, and the impact of climate change. Environ Sci Pollut Res 22:6511–6525. doi:10.1007/s11356-015-4205-4
Hasanuzzaman M, Nahar K, Alam M, Bhowmik PC, Hossain A, Rahman MM et al (2014) Potential use of halophytes to remediate saline soils. BioMed Res Int 2014:589341. doi:10.1155/2014/589341
Carillo P, Annunziata MG, Pontecorvo G, Fuggi A, Woodrow P (2011) Salinity stress and salt tolerance. In: Shanker A (ed) Abiotic stress in plants—echanisms and adaptations. InTech Europe, Rijeka, Croatia, pp 21–38
Qadir M, Quillérou E, Nangia V, Murtaza G, Singh M, Thomas RJ et al (2014) Economics of salt-induced land degradation and restoration. Nat Resour Forum 38:282–295
Qadir M, Oster JD (2002) Vegetative bioremediation of calcareous sodic soils: history, mechanisms and evaluation. Irrig Sci 21:91–101
Singh KN, Chatrath R (2001) Salinity tolerance. In: Reynolds MP, Monasterio JIO, McNab A (eds) Application of physiology in wheat breeding. CIMMYT, Mexico, DF, pp 101–110
Ghassemi F, Jakeman AJ, Nix HA (1995) Salinisation of land and water resources. CAB International, Canberra
United Nations University Institute for Water, Environment and Health (UNU-INWEH) (2014) World losing 2,000 hectares of farm soil daily to salt-induced degradation. http://inweh.unu.edu/world-losing-farm-soil-daily-salt-induced-degradation/. Accessed 11 Nov 2015
Jacobsen T, Adams RM (1958) Salt and silt in ancient Mesopotamian agriculture. Science 128:1251–1258
US Geological Survey (USGS) (2015) Where do the salts go?: a lesson from an ancient civilization. http://pubs.usgs.gov/fs/fs-170-98/. Accessed 11 Nov 2015
Alberta Environment, Environmental Sciences Division (2001) Salt Contamination Assessment & Remediation Guidelines, Pub. No. T/606 [Internet]. Alberta Environment, Edmonton, AB. environment.gov.ab.ca/info/library/6144.pdf. Accessed 16 Nov 2015
Renault S, MacKinnon M, Qualizza C (2003) Barley, a potential species for early reclamation of saline oil sands composite tailings. J Environ Qual 32:2245–2253
Saskatchewan Petroleum Industry/Government Environmental Committee (SPIGEC) (2009) Guideline No. 4, Saskatchewan upstream petroleum sites remediation guidelines [Internet]. Regina, SK. http://www.environment.gov.sk.ca/Default.aspx?DN=676d3b44-b996-478b-846b-e87406b67880&c=all&q=spigec. Accessed 16 Nov 2015
US Environmental Protection Agency (2015) Class II Wells—oil and gas related injection wells (Class II). http://water.epa.gov/type/groundwater/uic/class2/. Accessed 10 Nov 2015
Matthews JG, Shaw WH, MacKinnon MD, Cuddy RG (2002) Development of composite tailings technology at Syncrude. Int J Surf Mining Reclam Environ 16:24–39
Environment Canada (2013) Code of practice: the environmental management of road salts [Internet]. Environment Canada, Ottawa. http://www.ec.gc.ca/sels-salts/default.asp?lang=En&n=F37B47CE-1. Accessed 16 Nov 2015
Environment Canada/Health Canada. Canadian Environmental Protection Act (1999) Priority substances list assessment report—road salts [Internet]. Environment Canada/Health Canada, Ottawa. http://www.hc-sc.gc.ca/ewh-semt/pubs/contaminants/psl2-lsp2/road_salt_sels_voirie/index-eng.php. Accessed 16 Nov 2015
US Salinity Laboratory Staff (1954) Saline and alkali soils: diagnosis and improvement, Handbook No. 60 [Internet]. USDA, Washington, DC. http://www.ars.usda.gov/News/docs.htm?docid=10158&page=2. Accessed 16 Nov 2015
Seelig BD (2000) Salinity and sodicity in North Dakota soils—EB 57 [Internet]. NDSU Extension Service, North Dakota State University of Agriculture and Applied Science/U.S. Department of Agriculture, Fargo, ND. https://www.ndsu.edu/soilhealth/wp-content/uploads/2014/07/ND-saline-sodic-soils_20001.pdf. Accessed 16 Nov 2015
Zhu J-K (2007) Plant salt stress. In: Encyclopedia of life sciences (eLS). Wiley, Chichester. http://www.els.net. doi:10.1002/9780470015902.a0001300.pub2. Accessed 11 Nov 2015
Hu F, Li H, Liu X, Li S, Ding W, Xu C et al (2015) Quantitative characterization of non-classic polarization of cations on clay aggregate stability. PLoS One 10:e0122460. doi:10.1371/journal.pone.0122460
Levy GJ, Van Der Watt HVH (1990) Effect of exchangeable potassium on the hydraulic conductivity and infiltration rate of some South African soils. Soil Sci 149:69–77
Qadir M, Oster JD, Schubert S, Noble AD, Sahrawatk KL (2007) Phytoremediation of sodic and saline-sodic soils. Adv Agron 96:197–247. doi:10.1016/S0065-2113(07)96006-X
Qadir M, Steffens D, Yan F, Schubert S (2003) Sodium removal from a calcareous saline–sodic soil through leaching and plant uptake during phytoremediation. Land Degrad Dev 14:301–307
Munns R, Tester M (2008) Mechanisms of salinity tolerance. Annu Rev Plant Biol 59:651–681
Arbona V, Flors V, Jacas J, García-Agustín P, Gómez-Cadenas A (2003) Enzymatic and non-enzymatic antioxidant responses of Carrizo citrange, a salt-sensitive citrus rootstock, to different levels of salinity. Plant Cell Physiol 44:388–394
Flowers TJ, Colmer TD (2008) Salinity tolerance in halophytes. New Phytol 179:945–963
Barkla BJ, Pantoja O (2011) Plasma membrane and abiotic stress. In: Angus SM, Peer W, Schulz B (eds) The plant plasma membrane—plant cell monographs, vol 19. Springer, Berlin, pp 457–470
Kang S-M, Khan AL, Waqas M, You Y-H, Kim J-H, Kim J-G et al (2014) Plant growth-promoting rhizobacteria reduce adverse effects of salinity and osmotic stress by regulating phytohormones and antioxidants in Cucumis sativus. J Plant Interact 9:673–682. doi:10.1080/17429145.2014.894587
Nawaz K, Hussain K, Majeed A, Khan F, Afghan S, Ali K (2010) Fatality of salt stress to plants: morphological, physiological and biochemical aspects. Afr J Biotechnol 9:5475–5480
Neocleous D, Koukounaras A, Siomos AS, Vasilakakis M (2014) Changes in photosynthesis, yield, and quality of baby lettuce under salinity stress. J Agric Sci Technol 16:1335–1343
Shibli RA, Kushad M, Yousef GG, Lila MA (2007) Physiological and biochemical responses of tomato microshoots to induced salinity stress with associated ethylene accumulation. J Plant Growth Regul 51:159–169
Hedrich R (2012) Ion channels in plants. Physiol Rev 92:1777–1811. doi:10.1152/physrev.00038.2011
White PJ, Broadley MR (2001) Chloride in soils and its uptake and movement within the plant: a review. Ann Bot 88:967–988
Hasegawa PM (2013) Sodium (Na+) homeostasis and salt tolerance of plants. Environ Exp Bot 92:19–31. doi:10.1016/j.envexpbot.2013.03.001
Kronzucker HJ, Britto DT (2011) Sodium transport in plants: a critical review. New Phytol 189:54–81
Almeida P, Katschnig D, de Boer AH (2013) HKT transporters—state of the art. Int J Mol Sci 14:20359–20385. doi:10.3390/ijms141020359
Apse MP, Blumwald E (2007) Na+ transport in plants. FEBS Lett 581:2247–2254. doi:10.1016/j.febslet.2007.04.014
Morth JP, Pedersen BP, Buch-Pedersen MJ, Andersen JP, Vilsen B, Palmgren MG et al (2011) A structural overview of the plasma membrane Na+,K+-ATPase and H+-ATPase ion pumps. Nat Rev Mol Cell Biol 12:60–70
Waters S, Gilliham M, Hrmova M (2013) Plant high-affinity potassium (HKT) transporters involved in salinity tolerance: structural insights to probe differences in ion selectivity. Int J Mol Sci 14:7660–7680. doi:10.3390/ijms14047660
Zepeda-Jazo I, Shabala S, Chen Z, Pottosin II (2008) Na+-K+ transport in roots under salt stress: article addendum. Plant Signal Behav 3:401–403
Bartels D, Sunkar R (2005) Drought and salt tolerance in plants. Crit Rev Plant Sci 24:23–58. doi:10.1080/07352680590910410
Miller G, Suzuki N, Ciftci-Yilmaz S, Mittler R (2010) Reactive oxygen species homeostasis and signalling during drought and salinity stresses. Plant Cell Environ 33:453–467
Yadav S, Irfan M, Ahmad A, Hayat S (2011) Causes of salinity and plant manifestations to salt stress: a review. J Environ Biol 32:667–685
Jha Y, Subramanian RB (2014) PGPR regulate caspase-like activity, programmed cell death, and antioxidant enzyme activity in paddy under salinity. Physiol Mol Biol Plant 20:201–207
Pottosin I, Velarde-Buendía AM, Bose J, Zepeda-Jazo I, Shabala S, Dobrovinskaya O (2014) Cross-talk between reactive oxygen species and polyamines in regulation of ion transport across the plasma membrane: implications for plant adaptive responses. J Exp Bot 65:1271–1283. doi:10.1093/jxb/ert423
Sharma P, Jha AB, Dubey RS, Pessarakli M (2012) Reactive oxygen species, oxidative damage, and antioxidative defense mechanism in plants under stressful conditions. J Bot 2012:217037. doi:10.1155/2012/217037
Oukarroum A, Bussotti F, Goltsev V, Kalaji HM (2015) Correlation between reactive oxygen species production and photochemistry of photosystems I and II in Lemna gibba L. plants under salt stress. Environ Exp Bot 109:80–88
Singh S, Khatri N, Katiyar A, Mudgil Y (2015) Molecular approaches in deciphering abiotic stress signaling mechanisms in plants. In: Pandey GK (ed) Elucidation of abiotic stress signaling in plants: functional genomics perspectives, vol 1. Springer, New York, pp 41–73
Saha J, Brauer EK, Sengupta A, Popescu SC, Gupta K, Gupta B (2015) Polyamines as redox homeostasis regulators during salt stress in plants. Front Environ Sci 3:21. doi:10.3389/fenvs.2015.00021
Valderrama R, Corpas FJ, Carreras A, Gómez-Rodríguez MV, Khaki M, Pedrajas JR et al (2006) The dehydrogenase-mediated recycling of NADPH is a key antioxidant system against salt-induced oxidative stress in olive plants. Plant Cell Environ 29:1449–1459
Pottosin I, Shabala S (2014) Polyamines control of cation transport across plant membranes: implications for ion homeostasis and abiotic stress signaling. Front Plant Sci 5:154. doi:10.3389/fpls.2014.00154
Chinnusamy V, Jagendorf A, Zhu J-K (2005) Understanding and improving salt tolerance in plants. Crop Sci 45:437–448
Ji H, Pardo JM, Batelli G, Van Oosten MJ, Bressan RA, Li X (2013) The salt overly sensitive (SOS) pathway: established and emerging roles. Mol Plant 6:275–286
Shi H, Quintero FJ, Pardo JM, Zhu J-K (2002) The putative plasma membrane Na+/H+ antiporter SOS1 controls long-distance Na+ transport in plants. Plant Cell 14:465–477
Zhu J, Fu X, Koo YD, Zhu J-K, Jenney FE Jr, Adams MWW et al (2007) An enhancer mutant of Arabidopsis salt overly sensitive 3 mediates both ion homeostasis and the oxidative stress response. Mol Cell Biol 27:5214–5224
Saleem M, Arshad M, Hussain S, Bhatti AS (2007) Perspective of plant growth promoting rhizobacteria (PGPR) containing ACC deaminase in stress agriculture. J Ind Microbiol Biotechnol 34:635–648. doi:10.1007/s10295-007-0240-6
Blumwald E (2000) Sodium transport and salt tolerance in plants. Curr Opin Cell Biol 12:431–434
Gao H-J, Yang H-Y, Bai J-P, Liang X-Y, Lou Y, Zhang J-L et al (2015) Ultrastructural and physiological responses of potato (Solanum tuberosum L.) plantlets to gradient saline stress. Front Plant Sci 5:787. doi:10.3389/fpls.2014.00787
Kumar K, Kumar M, Kim S-R, Ryu H, Cho Y-G (2013) Insights into genomics of salt stress response in rice. Rice 6:27. doi:10.1186/1939-8433-6-27
Kurusu T, Kuchitsu K, Tada Y (2015) Plant signaling networks involving Ca2+ and Rboh/Nox-mediated ROS production under salinity stress. Front Plant Sci 6:427. doi:10.3389/fpls.2015.00427
Uozumi N, Schroeder JI (2010) Ion channels and plant stress: past, present and future. In: Demidchik V, Maathuis F (eds) Ion channels and plant stress responses. Springer, Berlin, pp 1–22
Gorham J, Wyn Jones RG, McDonnell E (1985) Some mechanisms of salt tolerance in crop plants. Plant Soil 89:15–40
Munns R (2002) Comparative physiology of salt and water stress. Plant Cell Environ 25:239–250
Walter H (1961) Salinity problems in the acid zones. The adaptations of plants to saline soils. Arid Zone Res 14:65–68
Tester M, Davenport R (2003) Na+ tolerance and Na+ transport in higher plants. Ann Bot 91:503–527
Akhter J, Murray R, Mahmood K, Malik KA, Ahmed S (2004) Improvement of degraded physical properties of a saline-sodic soil by reclamation with kallar grass (Leptochloa fusca). Plant Soil 258:207–216
Ammari TG, Al-Hiary S, Al-Dabbas M (2013) Reclamation of saline calcareous soils using vegetative bioremediation as a potential approach. Arch Agron Soil Sci 59:367–375. doi:10.1080/03650340.2011.629813
Hue NV, Campbell S, Li QX, Lee CR, Fong J (2002) Reducing salinity and organic contaminants in the Pearl Harbor dredged material using soil amendments and plants. Remed J 12:45–63
Chang P, Gerhardt KE, Huang X-D, Yu X-M, Glick BR, Gerwing PD et al (2014) Plant growth-promoting bacteria facilitate the growth of barley and oats in salt-impacted soil: implications for phytoremediation of saline soils. Int J Phytoremediat 16:1133–1147
Greenberg BM, Gerwing P, Huang X-D (2011) A novel phytoremediation technology used to successfully treat salt impacted soils in situ. Can Reclam 11:8–12
Manousaki E, Kalogerakis N (2011) Halophytes present new opportunities in phytoremediation of heavy metals and saline soils. Ind Eng Chem Res 50:656–660. doi:10.1021/ie100270x
Ozawa T, Wu J, Fujii S (2007) Effect of inoculation with a strain of Pseudomonas pseudoalcaligenes isolated from the endorhizosphere of Salicornia europea on salt tolerance of the glasswort. Soil Sci Plant Nutr 53:12–16
Bennett TH, Flowers TJ, Bromham L (2013) Repeated evolution of salt-tolerance in grasses. Biol Lett 9:20130029. doi:10.1098/rsbl.2013.0029
Flowers TJ (2004) Improving crop salt-tolerance. J Exp Biol 55:307–319
Dodd IC, Pérez-Alfocea F (2012) Microbial amelioration of crop salinity stress. J Exp Bot 63:3415–3428. doi:10.1093/jxb/ers033
Gerhardt KE, Greenberg BM, Glick BR (2006) The role of ACC deaminase in facilitating the phytoremediation of organics, metals and salt. Curr Trends Microbiol 2:61–72
Huang X-D, El-Alawi YS, Penrose D, Glick BR, Greenberg BM (2004) A multi-process phytoremediation system for removal of polycyclic aromatic hydrocarbons from contaminated soils. Environ Pollut 130:465–476
Upadhyay SK, Singh DP (2015) Effect of salt-tolerant plant growth-promoting rhizobacteria on wheat plants and soil health in a saline environment. Plant Biol 17:288–293. doi:10.1111/plb.12173
Kohler J, Caravaca F, Roldán A (2010) An AM fungus and a PGPR intensify the adverse effects of salinity on the stability of rhizosphere soil aggregates of Lactuca sativa. Soil Biol Biochem 42:429–434
Saravanakumar D, Samiyappan R (2007) ACC deaminase from Pseudomonas fluorescens mediated saline resistance in groundnut (Arachis hypogea) plants. J Appl Microbiol 102:1283–1292
Zhang H, Kim M-S, Sun Y, Dowd SE, Shi H, Paré W (2008) Soil bacteria confer plant salt tolerance by tissue-specific regulation of the sodium transporter HKT1. Mol Plant-Microb Interact 21:737–744
Cassán F, Maiale S, Masciarelli O, Vidal A, Luna V, Ruiz O (2009) Cadaverine production by Azospirillum brasilense and its possible role in plant growth promotion and osmotic stress mitigation. Eur J Soil Biol 45:12–19
Xie S-S, Wu H-J, Zang H-Y, Wu L-M, Zhu Q-Q, Gao X-W (2014) Plant growth promotion by spermidine-producing Bacillus subtilis OKB105. Mol Plant Microb Interact 27:655–663. doi:10.1094/MPMI-01-14-0010-R
Joseph B, Jini D (2010) Salinity induced programmed cell death in plants: challenges and opportunities for salt-tolerant plants. J Plant Sci 5:376–390
Barriuso J, Ramos Solano B, Gutierrez Manero FJ (2008) Protection against pathogen and salt stress by four plant growth-promoting rhizobacteria isolated from Pinus sp. on Arabidopsis thaliana. Phytopathology 98:666–672
Vacheron J, Desbrosses G, Bouffaud M-L, Touraine B, Moënne-Loccoz Y, Muller D et al (2013) Plant growth-promoting rhizobacteria and root system functioning. Front Plant Sci 4:356. doi:10.3389/fpls.2013.00356
Yildrim E, Donmez MF, Turan M (2008) Use of bioinoculants in ameliorative effects on radish plants under salinity stress. J Plant Nutr 31:2059–2074
Kiliç CC, Kukul YS, Ana D (2008) Performance of purslane (Portulaca oleracea L.) as a salt-removing crop. Agric Water Manage 95:854–858
Nazzal KE, Abdul-Kareem AW (2013) Phytoremediation of salt-affected soils at Al-Jazeera northern irrigation project/Ninavah/Iraq. Mesop J Agric 41:294–298
Hamidov A, Beltrao J, Neves A, Khaydarova V, Khamidov M (2007) Apocynum lancifolium and Chenopodium album—potential species to remediate saline soils. 3(7):123–128. http://www.worldses.org/journals/environment/environment-july2007.htm
Nedunuri KV, Govindaraju RS, Banks MK, Schwab AP, Chen Z (2000) Evaluation of phytoremediation for field-scale degradation of total petroleum hydrocarbons. J Environ Eng 126:483–490
Alberta Environment, Air, Land and Waste Policy Branch (CAN) (2010) Alberta Tier 2 Soil and Groundwater Remediation Guidelines. [Internet]. Alberta Environment, Edmonton. http://environment.gov.ab.ca/info/library/7752.pdf. Accessed 12 Nov 2015
Rozena J, Flowers T (2008) Crops for a salinized world. Science 322:1478
North Dakota State Government (USA) (2006) A guide for remediation of salt/hydrocarbon impacted soil. [Internet]. North Dakota Industrial Commission, Department of Mineral Resources, Bismark. https://www.dmr.nd.gov/downloads/soilRemediationGuideL.pdf . Accessed 27 Nov 2015
Greenberg BM, Huang X-D, Gerwing P, Yu X-M, Chang P-C, Wu SS, et al (2008) Phytoremediation of salt impacted soils: greenhouse and field trials using plant growth promoting rhizobacteria (PGPR) to improve plant growth and salt phytoaccumulation. In: Proceedings of the 31st AMOP Technical Seminar on Environmental Contamination and Response, Ottawa, ON, Canada. Environment Canada, Ottawa, pp 627–637, 3–5 June 2008
Greenberg, BM, X-D Huang, K Gerhardt, P Mosley, X-M Yu, S Liddycoat, et al (2012) Phytoremediation of petroleum and salt impacted soils: a scientifically-based innovative remediation process. In: Proceedings of the 35th AMOP Technical Seminar on Environmental Contamination and Response, Vancouver, BC, Canada. Environment Canada, Ottawa, pp 420–433, 5–7 June 2012
Greenberg, B, X-D Huang, P Mosley, K Gerhardt, X-M Yu, S Liddycoat, et al (2014) Phytoremediation of PHC and salt impacted soil in terrestrial and riparian environments—Remediation to generic Tier 1 guideline values and soil toxicity. In: Proceedings of the 37th AMOP Technical Seminar on Environmental Contamination and Response, Canmore, AB, Canada. Environment Canada, Ottawa, pp 468–481, 3–5 June 2014
Gurska J, Wang W, Gerhardt KE, Khalid AM, Isherwood DM, Huang X-D et al (2009) Three year field test of a plant growth promoting rhizobacteria enhanced phytoremediation system at a land farm for treatment of hydrocarbon waste. Environ Sci Technol 43:4472–4479
Huang X-D, El-Alawi YS, Penrose D, Glick BR, Greenberg BM (2004) Responses of three grass species to creosote during phytoremediation. Environ Pollut 130:453–463
Huang X-D, El-Alawi YS, Gurska J, Glick BR, Greenberg BM (2005) A multi-process phytoremediation system for decontamination of persistent total petroleum hydrocarbons (TPHs) from soils. Microchem J 81:139–147
Gliessman SR, Altieri MA (1982) Polyculture cropping has advantages. Calif Agric 36:14–16
Patten CL, Glick BR (2002) The role of bacterial indoleacetic acid in the development of the host plant root system. Appl Environ Microbiol 68:3795–3801
Chang P-C (2007) The use of plant growth-promoting rhizobacteria (PGPR) and an arbuscular mycorrhizal fungus (AMF) to improve plant growth in saline soils for phytoremediation. MSc thesis. University of Waterloo, Waterloo, ON
Bhattarai T, Hess D (1993) Yield responses of Nepalese spring wheat (Triticum aestivum L) cultivars to inoculation with Azospirillum spp of Nepalese origin. Plant Soil 151:67–76
Burd GI, Dixon DG, Glick BR (2000) Plant growth-promoting bacteria that decrease heavy metal toxicity in plants. Can J Microbiol 46:237–245
Greenberg BM, X-D Huang, Y Gurska, KE Gerhardt, W Wang, MA Lampi, et al (2006) Successful field tests of a multi-process phytoremediation system for decontamination of persistent petroleum and organic contaminants. In: Proceedings of the 29th Arctic and Marine Oilspill Program (AMOP) Technical Seminar, vol 1, Vancouver, BC, Canada. Environment Canada, Ottawa, pp 389–400, 6–8 June 2008
MacNeill G (2011) Plant-growth promoting rhizobacteria enhanced phytoremediation of saline soils and salt uptake into plant biomass. MSc thesis. University of Waterloo, Waterloo, ON
Wu SS (2009) Enhanced phytoremediation of salt-impacted soils using plant growth-promoting rhizobacteria (PGPR). MSc thesis. University of Waterloo, Waterloo, ON
Glick BR, Karaturovic DM, Newell PC (1995) A novel procedure for rapid isolation of plant growth promoting pseudomonads. Can J Microbiol 41:533–536
Duan J, Jiang W, Cheng Z, Heikkila JJ, Glick BR (2013) The complete genome sequence of the plant growth-promoting bacterium Pseudomonas sp. UW4. PLoS One. doi:10.1371/journal.pone.0058640
Asheraf M, Harris PJC (2013) Photosynthesis under stressful environments: an overview. Photosynthetica 51:163–190
Gururani MA, Venkatesh J, Tran L-SP (2015) Regulation of photosynthesis during abiotic stress-induced photoinhibition. Mol Plant 8:1304–1320
Jiang Q, Roche D, Monaco TA, Durham S (2006) Gas exchange, chlorophyll fluorescence parameters and carbon isotope discrimination of 14 barley genetic lines in response to salinity. Field Crops Res 96:269–278
Brugnoli E, Björkman O (1992) Growth of cotton under continuous salinity stress-influence on allocation pattern, stomatal and nonstomatal components of photosynthesis and dissipation of excess light energy. Planta 187:335–347
Meloni DA, Oliva CA (2003) Photosynthesis and activity of superoxide dismutase, peroxidase and glutathione reductase in cotton under salt stress. Environ Exp Bot 49:69–76
Nazir N, Ashraf M, Ejaz R (2001) Genomic relationships in oilseed Brassicas with repect to salt tolerance-photosynthetic capacity and ion relations. Pak J Bot 33:483–501
Raza SH, Athar HR, Ashraf M (2006) Influence of exogenously applied glycine betaine on photosynthetic capacity of two differently adapted wheat cultivars under salt stress. Pak J Bot 38:341–351
Baker NR (2008) Chlorophyll fluorescence: a probe of photosynthesis in vivo. Annu Rev Plant Biol 59:89–113
Maxwell K, Johnson GN (2000) Chlorophyll fluorescence—a practical guide. J Exp Bot 51:659–668
Björkman O, Demmig B (1987) Photon yield of O2 evolution and chlorophyll fluorescence at 77 K among vascular plants of diverse origins. Planta 170:489–504
Johnson GN, Young AJ, Scholes JD, Horton P (1993) The dissipation of excess excitation energy in British plant species. Plant Cell Environ 16:673–679
Genty B, Briantais J-M, Baker NR (1989) The relationship between quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. Biochim Biophys Acta 990:87–92
Dietz KJ, Schreiber U, Heber U (1985) The relationship between the redox state of QA and photosynthesis in leaves at various carbon-dioxide, oxygen and light regimes. Planta 166:219–226
Schreiber U, Bilger W, Neubauer C (1994) Chlorophyll fluorescence as a nonintrusive indicator for rapid assessment of in vivo photosynthesis. In: Schulze E-D, Caldwell MM (eds) Ecophysiology of photosynthesis. Springer, Berlin, pp 49–70
Schreiber U, Schliwa U, Bilger W (1986) Continuous recording of photochemical and non-photochemical chlorophyll fluorescence quenching with a new type of modulation fluorometer. Photosyn Res 10:51–62
Weis E, Berry JA (1987) Quantum efficiency of photosystem II in relation to energy-dependent quenching of chlorophyll fluorescence. Biochim Biophys Acta 894:198–208
Rosenqvist E, Kooten OV (2003) Chlorophyll fluorescence: a general description and nomenclature. In: DeEll JR, Toivonen PMA (eds) Practical applications of chlorophyll fluorescence in plant biology. Kluwer Academic, New York, pp 32–71
Babu TS, Marder JB, Tripuranthakam S, Dixon DG, Greenberg BM (2001) Synergistic effects of a photooxidized polycyclic aromatic hydrocarbon and copper on photosynthesis and plant growth: evidence that in vivo formation of reactive oxygen species is a mechanism of copper toxicity. Environ Toxicol Chem 20:1351–1358
Naidoo G, Somaru R, Achar P (2008) Morphological and physiological responses of the halophyte, Odyssea paucinervis (Staph) (Poaceae), to salinity. Flora 203:437–447
DeEll JR, Toivonen PMA (eds) (2003) Practical applications of chlorophyll fluorescence in plant biology. Kluwer Academic, New York
Netondo GW, Onyango JC, Beck E (2004) Sorghum and salinity: II. Gas exchange and chlorophyll fluorescence of sorghum under salt stress. Crop Sci 44:806–811
Zhao GQ, Ma BL, Ren CZ (2007) Growth, gas exchange, chlorophyll fluorescence, and ion content of naked oat in response to salinity. Crop Sci 47:123–131
Ogle D, St. John L (2010) Plants for saline to sodic soil conditions, TN Plant Materials No. 9A. Boise, Idaho—Salt Lake City. USDA-Natural Resources Conservation Service, Utah, Feb 2010 Revision. http://www.nrcs.usda.gov/Internet/FSE_PLANTMATERIALS/publications/idpmstn9328.pdf
Campos PS, Quartin V, Ramalho JC, Nunes MA (2003) Electrolyte leakage and lipid degradation account for cold sensitivity in leaves of Coffea sp. plants. J Plant Physiol 160:283–292
Bajji M, Kinet JM, Lutts S (2002) The use of the electrolyte leakage method for assessing cell membrane stability as a water stress tolerance test in durum wheat. J Plant Growth Regul 36:61–70
Zhang YF, Wang P, Yang YF, Bi Q, Tian SY, Shi XW (2011) Arbuscular mycorrhizal fungi improve reestablishment of Leymus chinensis in bare saline-alkaline soil: implication on vegetation restoration of extremely degraded land. J Arid Environ 75:773–778
Acknowledgements
The work performed in the lab of B. M. G. was supported by grants from the Natural Sciences and Engineering Research Council of Canada.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2017 Springer International Publishing AG
About this chapter
Cite this chapter
Gerhardt, K.E., MacNeill, G.J., Gerwing, P.D., Greenberg, B.M. (2017). Phytoremediation of Salt-Impacted Soils and Use of Plant Growth-Promoting Rhizobacteria (PGPR) to Enhance Phytoremediation. In: Ansari, A., Gill, S., Gill, R., R. Lanza, G., Newman, L. (eds) Phytoremediation. Springer, Cham. https://doi.org/10.1007/978-3-319-52381-1_2
Download citation
DOI: https://doi.org/10.1007/978-3-319-52381-1_2
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-52379-8
Online ISBN: 978-3-319-52381-1
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)