
Abstract
Agricultural crop yield is often reduced by continuous variations of environmental conditions including both abiotic and biotic stresses. The use of nanoparticles (NPs) has the potential to offset the ill effects of these stresses and increased the plant productivity. The application of synthetic chemicals including fungicides, herbicides, and insecticides to maintain optimum crop yield is not only targeting specific flora and fauna in the fields but also kills other beneficial species which disturbs the balance of ecosystem. On the contrary, the use of NPs (1–100 nm size) having enlarged surface area and good associations with the plant systems is the best alternative route of crop improvement. These can be applied at any developmental stage of plants. NPs can be made up of either inorganic or organic molecules or combination of both. Usually, NPs are used in agriculture with the aim of protecting crop plants from oxidative stresses by activating the machinery of antioxidative enzymes in plant cells. The main mechanism by which NPs interact and alter particular protein function in plants system involves chemical processes such as reaction with sulfhydryl and carboxyl groups. When plants faced the environmental disturbances, cytosolic calcium ions (Ca2+) and calcium-binding proteins (CaBPs) in plant cells play a pivotal role in activating defense mechanisms. NPs can take the place of Ca2+ ions and nitric oxide as messenger molecules inside the cells. By using the right concentration of NPs, expression of the stress-responsive genes as well as the activation of the plant’s defense system can be regulated in a controlled manner. Here, we have discussed thoroughly how judicious implication of nano-particles would be the best option in near future to enhance the crop management practices in a very precise manner.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others

Keywords
- Abiotic stresses
- Biotic stresses
- Calcium ions
- Calcium-binding proteins
- Nanoparticles
- Reactive oxygen species
1 Introduction
Plants are the most vulnerable living entities as they are continually exposed to a variety of environmental changes and many stress factors, either individually or in combination, throughout their life cycles. Since the last decade, crop output has been deteriorated globally due to lack of arable land, water scarcity, ever-changing climate, and low pesticide usage efficiency. Drought, salt, freezing, flooding, UV radiation, and other abiotic stresses on plants are becoming more widely recognized as important agricultural concerns (Wani et al. 2016). All these stresses in combination cause more than 50% yield loss from important agricultural crops (Hirayama and Shinozaki 2010) throughout the globe. As the enhancement of food production in this scenario is becoming a difficult task, and progress in production of abiotic stress-tolerant crop varieties is slow, more research is needed to understand the negative impact of these stresses on plant developmental growth, as well as possible reclaiming strategies using minimal agricultural inputs are required to overcome the effects of such stresses on plants (Wani et al. 2016; Elsakhawy et al. 2018; Zhao et al. 2020). Plants can acclimate to such pressures by introducing molecular, cellular, and physiological changes (Yadav et al. 2020). It has been discovered that numerous enzymatic and non-enzymatic antioxidants that scavenge the damaging reactive oxygen species (ROS) are present at a satisfactory amount inside the plant system. Catalase (CAT), glutathione reductase, superoxide dismutase (SOD), and ascorbate peroxidase (APX) constitute enzymatic antioxidants while glutathione and ascorbate are the non-enzymatic antioxidants that work continuously to destroy harmful ROS through shikimate-phenylpropanoid or ascorbate biosynthesis pathways (Khan et al. 2017; Zhao et al. 2020). The ROS molecules such as hydroxyl radical, superoxides, hydrogen peroxides, etc., are basically produced in mitochondria, chloroplasts, and peroxisomes at the time of photosynthesis and cellular respiration under stressful environment (Khan et al. 2017; Tripathy and Oelmüller 2012). Excessive ROS build-up caused by oxidative stress destroys membrane lipids, proteins, and nucleic acids, resulting in abnormal development (Khan et al. 2017). NPs imitate the activity of antioxidative enzymes in plants, making them effective ROS scavengers (Wei and Wang 2013; Khan et al. 2017). Engineered NPs with dimensions of 1–100 nm (Khan and Upadhyaya 2019) possess a very high reactivity and enlarged surface area which have ambiguous devoir in minimizing the harmful and negative effects of diverse environmental stresses on plant system (Elsakhawy et al. 2018; Mittal et al. 2020).
Eric Drexler has gained huge appreciation for popularizing the term “nanotechnology” in the realm of producing tools and technology for precision agriculture (Khan and Upadhyaya 2019). Nanotechnology is the science of using NPs which are molecular aggregates, synthesized at the nanoscale through either physical, chemical, and biological methods (Singh et al. 2016). They interact with plants to improve their developmental growth, increase productivity, and regulate the expression of various genes to overcome unpredictable abiotic stresses (Tan et al. 2017; Khan and Upadhyaya 2019). Due to their tiny size, they have a variety of physicochemical features, including increased reactivity rate in plant cells and tissues (Dubchak et al. 2010). However, plant response to NPs is debatable since it differs depending on plant type and NPs used (Lin and Xing 2007). Besides the positive effects, so many studies have shown several toxicological effects of NPs (Husen and Siddiqi 2014). Excessive use of NPs to plants under stressed condition can alter the process of ROS production resulting in oxidative damage (Chichiriccò and Poma 2015). NPs used in food and agriculture sector may be made up of inorganic, organic, and combined in nature. Inorganic NPs include metal and metal oxides, organic NPs include mainly natural products while combined NPs include clay. NPs can be of different morphological structures like rod shape, pyramid, micro-flower, etc., depending on the purpose of use like food-packaging, nanomedicine, agriculture biotechnology, preparation of biosensors, etc. Silver NPs are mostly commercially used because of their promising antimicrobial activity; while, gold NPs are used as sensor or detector. There is an urgent need for understanding the interaction between plants and NPs for the sake of crop production, nutrient utilization, and crop-health management. The general applications of NPs in the field of agriculture science have been shown in Fig. 6.1.

Applications of NPs in agricultural practices
2 Commonly Used NPs and Their Synthesis
NPs are biocompatible, stable, and may be directed to a specific location inside the plant body. Their ability to boost crops and control various stresses is solely determined by their shape, size, and chemical behaviors. Copper (Cu), silver (Ag), gold (Au), and other noble metals having a large absorption range (Marmiroli et al. 2015) are often used as NPs and work on the basis of localized surface plasmon resonance (LSPR). They are synthesized by either top-down or bottom-up processes (Omar et al. 2019; Kumar et al. 2020). In bottom-up approach (also known as sol-gel approach), atoms and molecules are assembled together to form NPs of desired size and shape. In top-down approach, atoms and molecules are separated from the bulk material to get the desired NPs. Nano-sized materials can be prepared by different physical, chemical, biological methods either singly or in combination. NPs that are synthesized using the plant extracts possess a functionalized surface having proteins, ligands, polysaccharides, which are otherwise absent when synthesized by either physical or chemical methods. These components increase the stability of particles and help in the attachment of functional molecules like antibodies, DNA to the NPs. Preparation of metallic NPs through the green synthesis method using plant extract is economic, eco-friendly, and less time-consuming as compared to the chemical synthesis method. Incorporation of the desired metal NPs in starch films reduces the problem of biodegradability and food containment. Nano-fertilizers, nano-sized nutrients, nano-sensors, carbon-based nano-materials, and nano-pesticides have been reported beneficial to agricultural practices by easy environmental detection, pathogen detection, sensing, degrading persistent chemicals, and remediation (Iavicoli et al. 2017).
Magnetic NPs are usually made from iron (Fe), cobalt (Co), magnesium (Mg), and manganese (Mn) either by microemulsions or co-precipitation thermal decomposition preparatory methods (Ashfaq et al. 2013). Graphene, carbon nanotubes (CNTs), carbon nanofibrils (CNFs), fullerenes, and other carbon-based nanostructures have recently been created for agricultural enhancement, crop protection, and environmental clean-up (Omar et al. 2019). CNTs and CNFs have the capacity to translocate from root to shoot inside plants, making them useful in agriculture (Ashfaq et al. 2017). Hybrid NPs on the other hand are synthesized as the conjugates of organic and inorganic nanostructures having lots of utility values in several biological applications. They are efficient in stress management in plant system than the other commonly used NPs as they enhance the biocompatibility of NPs (Qi et al. 2013).
3 Absorption, Translocation, and Accumulation of NPs into Plant System
Absorption of NPs by plant system and genuine interactions with them prominently depend on several factors including plant physiology and physiochemical properties of the used NPs such as their size, charge, shape, functionalization, and stability (Pérez-de-Luque 2017). Above ground plant parts such as stomata, cuticles, trichome, and hydathodes generally act as the sites through which NPs enter into the plant tissues; even the wound regions play the same role (Verma et al. 2019). Enormous scientific reports have mentioned that the uptake of NPs in plant system occurs through root absorption and transported by specialized plant tissues with some modifications, like biotransformation, bioaccumulation, and dissolution through crystal phases (Ali et al. 2021). The size of NPs is considered as the most important parameter as it enables their entrance either through cell wall pores or stomatal openings and even their subsequent transport processes intercellularly inside the plant system. The attachment and absorption of NPs in plant system is believed to rely directly on the existing negative charges on the cell wall surface. In addition, the hydrophobicity present on the plant surfaces has been found to play important role in absorption and translocation of NPs throughout the plant body (Kaphle et al. 2018).
NPs can enter the plant body via lateral roots with the aid of capillary forces and osmotic pressure, or they can enter directly through root epidermal cells (Lin and Xing 2008; Du et al. 2011) and finally translocated to xylem vessels via cortex and pericycle tissues (Dietz and Herth 2011; Miralles et al. 2012). NPs of sizes below 5 nm, under foliar application, are able to penetrate the leaf cuticle and reach the cell cytoplasm (Jeyasubramanian et al. 2016), while the NPs larger than the size of 10 nm enter through leaf stomata, with apoplastic and symplastic pathways for cellular transport (Ali et al. 2021). Through plasmodesmata, NPs can travel from one cell to another cell and are later on internalized in the cytoplasm (Wang et al. 2012). NPs of size 50–100 nm are translocated via apoplastic routes between the cells. Internalized NPs are conveyed through the vascular transport system via phloem sieve, allowing them to flow bidirectionally and accumulate in varying concentrations in leaves, stems, roots, fruits, and/or other tissues (Ruttkay-Nedecky et al. 2017).
In cytoplasm, the NPs are thought to bind with different cellular organelles and take part in various cellular metabolisms (Zhang and Monteiro-Riviere 2009). The overall mechanism of NP interaction with plant system is based mostly on chemical reactions involving generation of ROS, ionic movement through cell membrane, oxidative damage, and lipid peroxidation. When NPs penetrate plant cells, they react with sulfhydryl and carboxyl groups, causing change in protein function (Das and Das 2019). In order to find out the movement and localization of NPs to various organelles within the plant cell, we need to monitor and track NPs very precisely.
4 Mode of Action of NPs to Counter Harmful Effects of Environmental Stresses
Plants can show minimum effort to withstand acute adverse environment conditions (drought, salinity, chilling, radiation, and metallic stress), but they need a precise perception and transduction of stress stimulus for immediate activation of appropriate defense units against that stress factor before it becomes severe and damage plant’s cellular machinery. Activation of defense regiment in plant system primarily involves a temporary elevation of cytoplasmic Ca2+, production of intracellular secondary metabolites such as inositol and polyphosphate, generation of ROS, raise in cellular plant hormones concentration especially of abscisic acid (ABA), and higher increase in mitogen-activated protein kinase (MAPK)-dependent pathways which ultimately trigger the molecular machinery against the particular stress (Baxter et al. 2014; Jalil and Ansari 2019). Calcium ions (Ca2+) act as secondary messengers and play important role in signaling responses under various stresses. Ca2+-binding proteins (CaBPs) are activated by cytosolic Ca2+ and stimulate downstream pathways that result in gene expression modifications and results in plants’ adaptation to stresses (Khan et al. 2014; Khan et al. 2017).
When plant cells are exposed to various stresses, nitric oxide (NO) has been shown to cause an increase in calcium levels (Khan et al. 2014), implying that calcium stimulates NO generation. NPs act as an efficient Ca2+ substitution in CaBPs, resulting in the development of a stress-responsive gene cascade (Khan et al. 2017). NPs show duality in plant system under abiotic stress by enhancing production of ROS and scavenging of ROS simultaneously (Simon et al. 2013; Wei and Wang 2013). If any disturbance occurs between these two processes, it might result in oxidative stress and failure in the signal transduction process. The equilibrium of ROS inside the plant cells is managed by continuous synthesis and scavenging of ROS (Khan et al. 2017). To understand critically the mechanism of stress-signaling and significant response by NPs, in-depth investigations are needed with proteomic and genomic approaches. Furthermore, the application of NPs influences gene expression, cell division, cell elongation, and overall plant growth.
5 Biotic Stress: A Major Problem in Agricultural Crops
Diminution of agricultural productivity mainly depends on the exposure of biotic and abiotic factors. The growth of agricultural crops and plants, seed quality, and crop yield are affected by various biotic stresses. Biotic stresses involve damages primarily caused by living micro-organisms such as viruses, bacteria, fungi, insects, nematodes, etc. Wang et al. (2011) studied a proteomic-based technology in rice crop under biotic stress and found several biomarkers. Potential biomarkers like proteins that are common and unique to each stress can be used for generating the next generation crop plants using molecular breeding and genetic engineering to enhance crop yield and seed quality. Dresselhaus and Hückelhoven (2018) studied the role of metabolites during biotic and abiotic stresses in plants and how these two stresses are inter-related to each other for causing huge losses in the productivity of crops. Several agricultural crops like rice, wheat, maize, cotton, barley, chickpea, and other pulses are damaged every year due to biotic stresses and cause huge losses. Winter crop like wheat is affected by several diseases like powdery mildew, leaf blight, loose smut, rusts, etc.
Among these, a disease known as fusarium head blight caused by the fungi Fusarium graminearum and Fusarium poae causes huge losses to the wheat crop. Fungal diseases in wheat crop results in a reduction in kernel weight, vigor of seeds, and overall plant yield (Martínez et al. 2020; Dandve et al. 2019). Fusarium oxysporum damage different parts of wheat like leaves of seedlings, roots, etc., and results in qualitative and quantitative losses (Banerjee and Mittra 2018). Bocquet et al. (2018) conducted a study on Septoria tritici blotch disease of wheat, which is caused by fungus Zymoseptoria tritici and concluded that this fungus can cause extensive damage to the wheat crop. Ceresini et al. (2018) reviewed the past and present status of wheat blast disease caused by Pyricularia graminis tritici and also studied about its physiology, genetics, and its remedial measures.
5.1 Role of NPs to Deal with Biotic Stress in Crops
Plant pathologists and researchers are looking forward to safeguard food and agricultural products against biotic stress caused by bacteria, fungi, and viruses. Pathogen can be detected using this new technology which is nanoscale in size, have biological sensors with nano-based miniature detection devices. The use of various NPs in the crop plants during biotic stresses has been shown in Table 6.1. Abd-Elsalam (2013) has made efforts to develop a harmless method to manage fungal pathogens using nanoplatforms and nanomaterials. Rajeshkumar (2019) studied the antifungal impact of NPs against various fungal plant pathogens. According to him, various nanomaterials can be used to serve the purpose of fighting against fungal pathogens like Rhizoctonia crocorum and Cercospora canescens. Green methods are used to synthesize nanoparticles, which have antimicrobial activity. For example, gold NPs are synthesized using marine brown seaweed Turbinaria conoides and are used as an antibacterial agent against Bacillus subtilis, Klebsiella pneumoniae, and Streptococcus sp., while titanium oxide (TiO)-NPs are synthesized using a bacterial strain of Planomicrobium sp. and are used as antibacterial and antifungal agents against gram-positive bacteria like B. subtilis, gram-negative bacteria Klebsiella planticola, and against fungal cultures such as Aspergillus niger. Engineered NPs also had several impacts like plant growth, cell structure, physiological and biochemical functions on plants along with applications (Siddiqui et al. 2015). These engineered NPs also had an effect on photosystems, mechanisms of reactive oxygen species, and several enzymes in crops, which ultimately help in fighting against the biotic stresses.
Another example of nanoparticle used in agriculture is the use of nano-ZnO to inhibit the activity of Fusarium head blight pathogen in wheat crop. Therefore, it is used as a novel seed treatment method as it prevents seed disease, increases seedling growth, and increases activities of antioxidant enzymes such as SOD and CAT (Liu et al. 2020). Rice is often damaged by one of the severe biotic factors, like Xanthomonas oryzae, which causes bacterial leaf blight (BLB) disease in rice. Specific use of biogenic silver NPs (AgNPs) has proved beneficial rather than the use of chemical methods to control this disease. These AgNPs were prepared using a naturally isolated Bacillus cereus strain SZT1 of bacteria. Aside from combating pathogens, these NPs aim to boost plant biomass, reduce cellular concentrations of ROS, and enhance antioxidant enzyme activity (Ahmed et al. 2020). Alvarez-Carvajal et al. (2020) showed that AgNPs coated with chitosan function effectively in restricting the mycelial growth of Fusarium oxysporum, which is responsible for causing wilt by up to more than 70% and also reduce the severity of disease in tomato. Silver NPs can also be synthesized by biological method using endophytic Pseudomonas poae strain CO of bacteria. This NP works as an excellent fungicide against Fusarium graminearum pathogen. Application of biosynthesized AgNPs also showed inhibition in spore germination, hyphal proliferation, germ tube elongation, and mycotoxin production of various pathogens. Ibrahim et al. (2020) also approved that AgNPs have great potential in defending wheat from fungal attack.
Another example of engineered nanoparticles used during biotic stress is titanium dioxide nanoparticles (TIO2-NPs), which plays significant role in inactivating various pathogenic bacteria’s. TIO2-NPs are synthesized by hydrothermal treatment using different temperatures ranging from 250 °C or 310 °C. It has photocatalytic activity and inactivates various plant pathogens like Xanthomonas arboricola, Pseudomonas syringae, and Erwinia amylovora (Kőrösi et al. 2020). Therefore, this engineered nanoparticle is very promising with its antimicrobial activity. The innovative use of silicon as nanoparticle in agriculture can not only meet the rising demand of food and agriculture but also help in developing sustainable environment by using agricultural wastes for the production of silicon NPs (Si-NPs) leading to waste management through recycling. Since Si-NPs are mesoporous in nature, it acts as a specific carrier for pesticides and fertilizers also known as nano-pesticides and nano-fertilizers respectively. It also involves in site-targeted delivery of nucleic acid, nutrients and increases crop protection, as well as plant growth and development. It reduces the damages caused by biotic and abiotic stresses in plants (Roychoudhury 2020). Since commercial fungicides are harmful to use due to the rise of microbial resistance nowadays, biological NPs are an alternative management tool used more preferably. Zinc NPs have an antimycotic activity against dark spore-forming pathogens like Bipolaris sorokiniana, Alternaria brassicicola, etc. (Kriti et al. 2020). It inhibits the spore formation, mycelial growth of fungus.
The interaction between NPs and protein (present in microbe) results in the inflammatory signal because microorganisms consider NPs as foreign material and results in microbial inhibition and reduced mycelial growth, when zinc-based NPs were used (Kriti et al. 2020). Luksiene et al. (2020) reported the importance of ZnO-NP as it has antifungal activity against pathogen Botrytis cinerea, infecting strawberry. However, ZnO-NP worked more efficiently in the presence of sunlight as it gets activated by ultraviolet (UV) and visible light, resulting in increased harvest. It therefore can replace conventional fungicide fenhexamid. Chitosan copper NPs (Ch-CuNPs) are known to possess antifungal properties (Rhizoctonia solani and Pythium aphanidermatum) against several fungal pathogens and can also be synthesized. In addition, NPs promote development in chili seeds by overcoming the low germination rate of sensitive seeds (Vanti et al. 2020). An endophytic fungus isolated from Solanum nigrum is used to make myco-NPs. Several plant pathogenic fungi, including Fusarium graminearum, Fusarium udum, Rhizoctonia solani, and Aspergillus niger, displayed antifungal activity and reduced pathogenic pressure by limiting their radial growth when treated with mycosilver NPs (Akther and Hemalatha 2019).
To combat dangerous phytopathogens, green production of AgNPs utilizing Azadirachta indica leaf extract is a preferable option to chemical agents (Haroon et al. 2019). The use of this NP is eco-friendly, safe, and inexpensive.
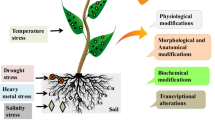
6 Advances in Abiotic Stress Mitigation by NPs
Fluctuation in the environmental conditions due to irregular rainfall, forest fire, melting of glaciers, faulty agricultural practices, and adverse effects of global warming due to industrial revolution constantly disturbs the metabolic equilibrium and developmental growth of plants (Jalil and Ansari 2019). Consequently, farmers as well as researchers become clueless to maintain sustainability in agriculture by mitigating all those stress factors. Advances in nanotechnology prestigiously offer huge scopes to check the detrimental impacts of various abiotic stress factors including drought, salinity, chilling, heat, metallic, and to some extent radiation on plant system. The use of NPs to mitigate the abiotic stresses in diverse crop plants has been put on view in Table 6.2.
6.1 Drought Stress
Drought is one of the serious abiotic stress factors that occurs regularly in dry locations and reduces agricultural productivity (Tripathi et al. 2017). A number of studies have recently demonstrated that applying silica NPs (SiNPs) at various concentrations increases drought tolerance in sorghums (Sorghum bicolor L.) by reducing their shoot to root ratio (Hattori et al. 2005). Varied concentrations of SiNPs cause physiological and biochemical reactions in hawthorns (Crataegus sp.) at different levels of drought stress (Ashkavand et al. 2015). Under water stress, SiNPs have also been shown to boost the activities of catalase (CAT) and peroxidase (POD) in the leaves of faba bean (Kalteh et al. 2018) and tomato (Siddiqui and Al-Whaibi 2014). Furthermore, SiNPs have been shown to have an influence on xylem humidity, water translocation, and turgor pressure, resulting in an increase in leaf relative water content and water usage efficiency in plants. Application of sodium silicate at low concentration (1.0 mM) has been found in mitigation of the harmful ramifications of moisture stress as well as membrane lipid peroxidation in wheat (Pei et al. 2010). In addition, foliar application (0.02%) of TiO2-NPs have been reported to give promising results on certain agronomic traits like plant height, seed number, ear weight, ear number, thousand-seed weight, starch and gluten content in grains, yield, biomass, and harvest index of various existing wheat cultivars under limited water condition (Jaberzadeh et al. 2013).
Similarly, applications of TiO2-NPs were studied in spinach (Lei et al. 2007) and Canola (Mahmoodzadeh et al. 2013) plants to mitigate the effect of drought stress by activating the antioxidant enzymes (SOD, CAT) machinery. Exogenous application of TiO2-NPs at lower dose has been observed to increase photosynthetic rate and nitrogen metabolism in spinach (Yang et al. 2006), linseed (Aghdam et al. 2016), and sugar beet (Borišev et al. 2016). Shallan et al. (2016) looked into the impacts of nano-TiO2 and nano-SiO2 on cotton production in drought-stricken conditions. They discovered that pre-treating cotton plants with nano-TiO2 and nano-SiO2 increased the content of plant pigments, total soluble sugars, proline content, total soluble proteins, total antioxidant capacity, and antioxidant enzyme activities under prolonged drought stress. Sedghi et al. (2013) showed that ZnONPs, which have the ability to boost seed germination percentage and rate, successfully lowered the fresh and dry weight of seeds, promoting drought tolerance in soybeans (Glycine max).
Taran et al. (2017) reported enhancement of ability to withstand drought by increasing enzymatic activities of SOD and CAT due to nano-zinc application in wheat (Triticum aestivum). Davar et al. (2014) exhibited how foliar application of iron NPs (FeNPs) promoted tolerance against drought stress in safflower cultivars. Advances in the application of AgNPs to decrease the deleterious effects of drought stress on lentils were also praised (Lens culinaris Medic). Hojjat and Ganjali (2016) found that judicious application of AgNPs can help lentil seeds germination, root length, and dry weight by alleviating drought stress, moderating loss of plant growth, and enhancing germination rate, root length, and dry weight.
6.2 Salinity Stress
Around 20% of arable land throughout the world has perished of salinity. Salinity or salt stress is the most critical environmental issue for the plant species belonging to the lycophyte category (Negrão et al. 2017; Khan and Upadhyaya 2019). In the arid, semiarid, and coastal environments, salinity stress may be triggered by the build-up of chloride (Cl−), sulfate (SO42−), sodium (Na+), calcium (Ca2+), and magnesium (Mg2+) ions (Singh and Husen 2020). It reduces agricultural output by interfering with a variety of physiological and biochemical processes involved in plant growth. The salinity stress caused by improper use of agrochemicals like pesticides, insecticides, etc., affects the soil’s fertility, leading to imbalance of osmotic potential and nutritional components of soil, which become toxic to plants growth and development (Khan and Upadhyaya 2019). Furthermore, synthetic chemicals used in agricultural practices badly impact photosynthesis, protein synthesis, lipid metabolism, and other vital processes (Parida and Das 2005). In this context, the use of nano-fertilizers may be a viable option for mitigating these challenges and reclaiming environmental sustainability.
Kalteh et al. (2014) found that applying SiNPs to basil (Ocimum basilicum) increased its chlorophyll content and proline levels significantly. Many studies also revealed that application of SiO2 NPs had the potential to increase the amount of chlorophyll, proline accumulation, and the activities of antioxidant enzyme in plant system under salt stress (Gao et al. 2006; Haghighi et al. 2012; Kalteh et al. 2018). SiO2 NPs encourage seed germination and improve yield-related features in lentil when salinity stress is prevalent. Furthermore, SiNPs are thought to promote osmotic potential and plant development by reducing Na+ toxicity under severe salt stress (Khan and Upadhyaya 2019). By altering the characteristics of the salt-stressed root plasma membrane, multi-walled carbon nanotubes exposure on broccoli under salinity stress has been shown to boost net CO2 assimilation and aquaporin transduction (Martínez-Ballesta et al. 2016).
Rossi et al. (2016) have shown that the application of CeO2 NPs significantly mitigated detrimental impacts on biomass production and activities of chloroplast in Brassica napus L. when subjected to grow under saline water. The limited solubility of CuO-NPs in alkaline soil may explain why CuO and ZnO-NPs boosted wheat growth and yield (Fathi et al. 2017). Hussein and Abou-Baker (2018) carried out a pot experiment to understand the effects of nano-zinc fertilizer on cotton growth, yield, and mineral status under salt stress. They revealed that irrigation with seawater reduced leaf dry weight and boll production, but foliar application of nano-zinc fertilizer at the optimal dose alleviated the negative effects of salinity and enhanced the yield parameters. To investigate the expression rate of salt stress genes, Almutairi (2016) implemented semiquantitative RT-PCR. He discovered that Ag-NPs caused up-regulation of four genes, including AREB, MAPK2, P5CS, and CRK1, as well as down-regulation of three other genes, including TAS14, DDF2, and ZFHD1. Such a pattern of expression revealed the involvement of Ag-NPs and their response to stress conditions, suggesting that they have the ability to improve plant tolerance to salt stress.
6.3 Heavy Metal Stress
Heavy metal (HM) pollution is becoming so crucial for retarded plant growth and severe yield loss throughout the globe (Chibuike and Obiora 2014; Khan et al. 2017). Under HM stress, plants lose their potential to withstand in a challenging environment. Severe metallic stress influences rapid ROS production, eventually promotes oxidative damage by disturbing cellular structure as well as membrane integrity along with degradation of cellular proteins (Sharma et al. 2012). Plants try to fight back the negative effects of heavy metal shock by developing their defense system at the cellular level. Plants undergo shock when heavy metals breach the cell wall and enter the cell. Infected plant parts respond quickly to HMs by accumulating biomolecules and nutrients, which activate the antioxidant enzyme machinery. Nanotechnology in this scenario has been recognized as one of the promising approaches for decreasing phytotoxicity caused by heavy metals (Tripathi et al. 2015; Gunjan and Zaidi 2014). Exposure to TiO2 NPs limited cadmium toxicity and increases photosynthetic rate in soybean (Singh and Lee 2016). Foliar treatment of ZnO-NPs to Leucaena leucocephala has been reported to enhance total soluble protein and photosynthetic pigment while decreasing membrane lipid peroxidation (Venkatachalam et al. 2017). NPs are becoming more important in the cost-effective rehabilitation of heavy metal-contaminated soils. NPs alleviated metal-induced toxicities during phytoremediation, according to Martínez-Fernández et al. (2017). Many studies have shown, however, that using NPs for crop improvement and phytoremediation has both good and negative effects on the agri-ecosystem.
6.4 UV-Irradiation
Plants exposed to radiation such as UV-B (frequency range: 280–315 nm) acquire an elevated amount of ROS in their cells, which directly or indirectly affects cell functions such as DNA damage, damage to chloroplast structure, disturbs cell functions, etc. (Chen et al. 2011; Wang et al. 2012; Hideg et al. 2013). Plants have evolved to accumulate phenolic chemicals, such as flavonoids and flavones, which absorb harmful UV rays, as well as enzymatic and non-enzymatic antioxidant defense mechanisms (Shen et al. 2010). Inclusion of NPs in the growth medium, on the other hand, may exacerbate detrimental effects of UV exposure.
For example, Regier et al. (2015) found that while CuO-NPs applied alone had no detrimental impacts when combined with UV light, they had a substantial negative influence on many physiological and morphological characteristics of Elodea nuttallii. Similarly, in wheat, subjected to a combination of cadmium telluride quantum dots (CdTe-QDs) and UV radiation, Chen et al. (2014) found reduced chlorophyll content, antioxidant enzyme activity, and enhanced DNA damage. When exposed to UV light, Lei et al. (2008) found that nano-anatase TiO2-treated spinach increased photosynthesis, electron transfer, photoreduction activity of photosystem II, oxygen evolution, and photophosphorylation (PSP) in chloroplast. Under UV-B exposure, nano-anatase exposure resulted in a large drop in superoxide radicals (O2−), H2O2, and MDA build-up, as well as a significant rise in antioxidant enzymes levels, indicating a prominent reduction in oxidative stress in plant cells (Lei et al. 2008).
6.5 Chilling Stress
Chilling or low-temperature stress is generally characterized by the occurrence of frost injuries in plant cells due to the accumulation of ice crystals within the tissues (Hasanuzzaman et al. 2013) under the exposure of very low temperature (0–15 °C). It negatively impacts on plant health by triggering loss of fluid and electrolyte leakage from the cellular membranes and consequently poor seed germination, stunted growth as well as reduced productivity (Singh and Husen 2020). Although vulnerability to cold stress depends on the degree of tolerance in various species (Heidarvand et al. 2011), plants suffering from low-temperature condition show disrupted photosynthesis by lowering down the chlorophyll content, transpiration rate, and degradation of Rubisco enzyme and its activity (Liu et al. 2012). In this context, many research studies showed that the use of NPs significantly increased production of Rubisco (Khan and Upadhyaya 2019), light immersion by chloroplast organelle (Ze et al. 2011), and inhibition in the generation of ROS (Giraldo et al. 2014). After application of TiO2 NPs, the activities of antioxidant enzymes have increased in plants (Mohammadi et al. 2016). The use of nano-TiO2 as a viable technique to reducing the detrimental effects of chilling stress has been recognized significantly. Application of NPs thus, causing suppression of lipid peroxidation, chlorophyll degradation, and synthesis of H2O2 under chilling stress, has been well-popularized in managing plant growth and health under various stresses (Kohan-Baghkheirati and Geisler-Lee 2015; Khan and Upadhyaya 2019; Singh and Husen 2020).
6.6 Heat Stress
Tending to temperature rise above the threshold limit for a long period of time often results in irreclaimable damage to growth and biological yield of crop plants (Wahid 2007). High-temperature stress increases production of ROS which leads to oxidative stress, and ultimately membrane lipid degradation as well as membrane ion leakage (Savicka and Škute 2010; Karuppanapandian et al. 2011; Aref et al. 2016). It also influences the reduction in photosynthetic rate as well as chlorophyll content (Khan and Upadhyaya 2019). In this regard, heat shock proteins (HSPs) have been identified as the key molecular chaperons to function in thermo-tolerance by plants (Singh and Husen 2020). Khodakovskaya et al. (2011) reported a significant contribution of multiwall carbon nanotubes (MWCNTs) in the upregulation of stress-responsive genes like HSP90 in both leaves and roots of tomato. Additionally, when NPs such as CeO2NP were exposed to maize, it resulted in the generation of hydrogen peroxide and also upregulated HSP70 gene (Zhao et al. 2012). Treatment with TiO2NPs also found to improve the mechanism of stomatal opening in tomato leaves under heat stress (Qi et al. 2013). Application of SeNPs at the nanoscale has been observed to mitigate the detrimental effect of heat stress in tomato (Haghighi et al. 2014) as well as sorghum (Djanaguiraman et al. 2018) by increasing the level of hydration ability, chlorophyll content, and various antioxidative features.
7 Conclusion
Nanotechnology is undoubtedly an essential element of several fields of agriculture. NPs reveal extremely low levels and have a kind and dose-dependent effect on plants. The scientific community is concerned about how to combat the loss of agricultural output caused by biotic and abiotic stresses. Several NPs are being investigated to determine their involvement in protecting plants from various environmental challenges, as well as their roles in promoting plant growth, crop production, and modifying plant processes. NPs have been demonstrated to be a more efficient and effective option for the production of nano-fertilizers than conventional fertilizers. Smaller NPs provide for easier penetration and control of water channels, which aid seed germination and plant growth; moreover, increased surface area allows for greater adsorption and controlled delivery of chemicals.
In contrast, the production of ROS is also described for NPs. The increased ROS level by NPs could be related to a stress signal amplification that can activate the defensive mechanism of the plant more efficiently. The uses of NPs approach have high potential to minimize the use of toxic chemicals as well as pesticides, fungicides, and insecticides on plants to combat environmental stresses and thus can lower the level of pollution of contaminants in air, groundwater, and in soil. Further research at multiple levels, including plant molecular and cellular levels, is needed for the function of nanomaterial to alleviate the damage caused by various environmental stresses.
References
K.A. Abd-Elsalam, Nanoplatforms for plant pathogenic fungi management. Fungal Genom. Biol. 2, e107 (2013)
I.O. Adisa, S. Rawat, V.L. Pullagurala, C.O. Dimkpa, W.H. Elmer, J.C. White, J.A. Hernandez-Viezcas, J.R. Peralta-Videa, J.L. Gardea-Torresdey, Nutritional status of tomato (Solanum lycopersicum) fruit grown in Fusarium-infested soil: Impact of cerium oxide nanoparticles. J. Agric. Food Chem. 68(7), 1986–1997 (2020)
M.T. Aghdam, H. Mohammadi, M. Ghorbanpour, Effects of nanoparticulate anatase titanium dioxide on physiological and biochemical performance of Linum usitatissimum (Linaceae) under well-watered and drought stress conditions. Rev. Bras. Bot. 39(1), 139–146 (2016)
T. Ahmed, M. Shahid, M. Noman, M.B. Niazi, F. Mahmood, I. Manzoor, Y. Zhang, B. Li, Y. Yang, C. Yan, J. Chen, Silver nanoparticles synthesized by using Bacillus cereus SZT1 ameliorated the damage of bacterial leaf blight pathogen in rice. Pathogens 9(3), 160 (2020)
T. Akther, S. Hemalatha, Mycosilver nanoparticles: Synthesis, characterization and its efficacy against plant pathogenic fungi. BioNanoScience 9(2), 296–301 (2019)
S. Ali, A. Mehmood, N. Khan, Uptake, translocation, and consequences of nanomaterials on plant growth and stress adaptation. J. Nanomater. 2021, 1–17 (2021)
Z.M. Almutairi, Effect of nano-silicon application on the expression of salt tolerance genes in germinating tomato (‘Solanum lycopersicum’ L.) seedlings under salt stress. Plant Omics 9(1), 106–114 (2016)
F. Alvarez-Carvajal, T. Gonzalez-Soto, A.D. Armenta-Calderón, R. Méndez Ibarra, E. Esquer-Miranda, J. Juarez, D. Encinas-Basurto, Silver nanoparticles coated with chitosan against Fusarium oxysporum causing the tomato wilt. Biotecnia 22(3), 73–80 (2020)
J. An, P. Hu, F. Li, H. Wu, Y. Shen, J.C. White, X. Tian, Z. Li, J.P. Giraldo, Emerging investigator series: Molecular mechanisms of plant salinity stress tolerance improvement by seed priming with cerium oxide nanoparticles. Environ. Sci. Nano 7(8), 2214–2228 (2020)
I.M. Aref, P.R. Khan, S. Khan, H. El-Atta, A.I. Ahmed, M. Iqbal, Modulation of antioxidant enzymes in Juniperus procera needles in relation to habitat environment and dieback incidence. Trees 30(5), 1669–1681 (2016)
M. Ashfaq, S. Singh, A. Sharma, N. Verma, Cytotoxic evaluation of the hierarchical web of carbon micronanofibers. Ind. Eng. Chem. Res. 52(12), 4672–4682 (2013)
M. Ashfaq, N. Verma, S. Khan, Carbon nanofibers as a micronutrient carrier in plants: Efficient translocation and controlled release of Cu nanoparticles. Environ. Sci. Nano 4(1), 138–148 (2017)
P. Ashkavand, M. Tabari, M. Zarafshar, I. Tomásková, D. Struve, Effect of SiO2 nanoparticles on drought resistance in hawthorn seedlings. Lesne Prace Badawcze 76(4), 350–359 (2015)
S. Avestan, M. Ghasemnezhad, M. Esfahani, C.S. Byrt, Application of nano-silicon dioxide improves salt stress tolerance in strawberry plants. Agronomy 9(5), 246 (2019)
A. Banerjee, B. Mittra, Morphological modification in wheat seedlings infected by Fusarium oxysporum. Eur. J. Plant Pathol. 152(2), 521–524 (2018)
A. Baxter, R. Mittler, N. Suzuki, ROS as key players in plant stress signalling. J. Exp. Bot. 65(5), 1229–1240 (2014)
F. Behboudi, Z. Tahmasebi Sarvestani, M.Z. Kassaee, S.A. Modares Sanavi, A. Sorooshzadeh, S.B. Ahmadi, Evaluation of chitosan nanoparticles effects on yield and yield components of barley (Hordeum vulgare L.) under late season drought stress. J. Water Environ. Nanotechnol. 3(1), 22–39 (2018)
L. Bocquet, C. Rivière, C. Dermont, J. Samaillie, J.L. Hilbert, P. Halama, A. Siah, S. Sahpaz, Antifungal activity of hop extracts and compounds against the wheat pathogen Zymoseptoria tritici. Ind. Crop. Prod. 122, 290–297 (2018)
J. Borgatta, C. Ma, N. Hudson-Smith, W. Elmer, C.D. Plaza Perez, R. De La Torre-Roche, N. Zuverza-Mena, C.L. Haynes, J.C. White, R.J. Hamers, Copper based nanomaterials suppress root fungal disease in watermelon (Citrullus lanatus): Role of particle morphology, composition and dissolution behavior. ACS Sustain. Chem. Eng. 6(11), 14847–14856 (2018)
M. Borišev, I. Borišev, M. Župunski, D. Arsenov, S. Pajević, Ž. Ćurčić, J. Vasin, A. Djordjevic, Drought impact is alleviated in sugar beets (Beta vulgaris L.) by foliar application of fullerenol nanoparticles. PLoS One 11(11), e0166248 (2016)
J.T. Buchman, W.H. Elmer, C. Ma, K.M. Landy, J.C. White, C.L. Haynes, Chitosan-coated mesoporous silica nanoparticle treatment of Citrullus lanatus (watermelon): Enhanced fungal disease suppression and modulated expression of stress-related genes. ACS Sustain. Chem. Eng. 7(24), 19649–19659 (2019)
P.C. Ceresini, V.L. Castroagudín, F.Á. Rodrigues, J.A. Rios, C. Eduardo Aucique-Pérez, S.I. Moreira, E. Alves, D. Croll, J.L. Maciel, Wheat blast: Past, present, and future. Annu. Rev. Phytopathol. 56, 427–456 (2018)
H. Chen, J. Zhai, R. Han, Influence of enhanced UV-B radiation on F-actin in wheat division cells. Plant Diver. Resour. 33(3), 306–310 (2011)
H. Chen, Y. Gong, R. Han, Cadmium telluride quantum dots (CdTe-QDs) and enhanced ultraviolet-B (UV-B) radiation trigger antioxidant enzyme metabolism and programmed cell death in wheat seedlings. PLoS One 9(10), e110400 (2014)
G.U. Chibuike, S.C. Obiora, Heavy metal polluted soils: Effect on plants and bioremediation methods. Appl. Environ. Soil Sci. 2014, 1–12 (2014)
G. Chichiriccò, A. Poma, Penetration and toxicity of nanomaterials in higher plants. Nano 5(2), 851–873 (2015)
S.C. Chun, M. Chandrasekaran, Chitosan and chitosan nanoparticles induced expression of pathogenesis-related proteins genes enhances biotic stress tolerance in tomato. Int. J. Biol. Macromol. 125, 948–954 (2019)
M.S. Dandve, S.G. Wagh, P.R. Bhagat, K. Pawar, S.A. Timake, A.A. Daspute, M.B. Pohare, Bacterial and fungal pathogen synergetics after co-infection in the wheat (Triticum aestivum L.). Biotechnol. J. Int. 11, 1–9 (2019)
M. Danish, M. Altaf, M.I. Robab, M. Shahid, S. Manoharadas, S.A. Hussain, H. Shaikh, Green synthesized silver nanoparticles mitigate biotic stress induced by Meloidogyne incognita in Trachyspermum ammi (L.) by improving growth, biochemical, and antioxidant enzyme activities. ACS Omega 6(17), 11389–11403 (2021)
Das, A. and Das, B., 2019. Nanotechnology a potential tool to mitigate abiotic stress in crop plants. In De Oliveira A. (eds), Abiotic and Biotic Stress in Plants, IntechOpen.
Z.F. Davar, A. Roozbahani, A. Hosnamidi, Evaluation the effect of water stress and foliar application of Fe nanoparticles on yield, yield components and oil percentage of safflower (Carthamus tinctorious L.). Int. J. Adv. Biol. Biomed. Res. 2(4), 1150–1159 (2014)
K.J. Dietz, S. Herth, Plant nanotoxicology. Trends Plant Sci. 16(11), 582–589 (2011)
C.O. Dimkpa, J.E. McLean, D.W. Britt, A.J. Anderson, Antifungal activity of ZnO nanoparticles and their interactive effect with a biocontrol bacterium on growth antagonism of the plant pathogen Fusarium graminearum. Biometals 26(6), 913–924 (2013)
C.O. Dimkpa, U. Singh, P.S. Bindraban, W.H. Elmer, J.L. Gardea-Torresdey, J.C. White, Zinc oxide nanoparticles alleviate drought-induced alterations in sorghum performance, nutrient acquisition, and grain fortification. Sci. Total Environ. 688, 926–934 (2019)
M. Djanaguiraman, N. Belliraj, S.H. Bossmann, P.V. Prasad, High-temperature stress alleviation by selenium nanoparticle treatment in grain sorghum. ACS Omega 3(3), 2479–2491 (2018)
T. Dresselhaus, R. Hückelhoven, Biotic and abiotic stress responses in crop plants. Agronomy 8(11), 267 (2018)
W. Du, Y. Sun, R. Ji, J. Zhu, J. Wu, H. Guo, TiO2 and ZnO nanoparticles negatively affect wheat growth and soil enzyme activities in agricultural soil. J. Environ. Monit. 13(4), 822–828 (2011)
S. Dubchak, A. Ogar, J.W. Mietelski, K. Turnau, Influence of silver and titanium nanoparticles on arbuscular mycorrhiza colonization and accumulation of radiocaesium in Helianthus annuus. Span. J. Agric. Res. 1, 103–108 (2010)
R.M. El-Ashry, M.T. El-Saadony, A.E. El-Sobki, A.M. El-Tahan, S. Al-Otaibi, A.M. El-Shehawi, A.M. Saad, N. Elshaer, Biological silicon nanoparticles maximize the efficiency of nematicides against biotic stress induced by Meloidogyne incognita in eggplant. Saudi J. Biol. Sci. (2021). https://doi.org/10.1016/j.sjbs.2021.10.013
T. Elsakhawy, A.E. Omara, T. Alshaal, H. El-Ramady, Nanomaterials and plant abiotic stress in agroecosystems. Environ. Biodiver. Soil Secur. 2(2018), 73–94 (2018)
J. Faraji, A. Sepehri, Exogenous nitric oxide improves the protective effects of TiO2 nanoparticles on growth, antioxidant system, and photosynthetic performance of wheat seedlings under drought stress. J. Soil Sci. Plant Nutr. 20(2), 703–714 (2020)
A. Fathi, M. Zahedi, S. Torabian, A. Khoshgoftar, Response of wheat genotypes to foliar spray of ZnO and Fe2O3 nanoparticles under salt stress. J. Plant Nutr. 40(10), 1376–1385 (2017)
X. Gao, C. Zou, L. Wang, F. Zhang, Silicon decreases transpiration rate and conductance from stomata of maize plants. J. Plant Nutr. 29(9), 1637–1647 (2006)
J.P. Giraldo, M.P. Landry, S.M. Faltermeier, T.P. McNicholas, N.M. Iverson, A.A. Boghossian, N.F. Reuel, A.J. Hilmer, F. Sen, J.A. Brew, M.S. Strano, Plant nanobionics approach to augment photosynthesis and biochemical sensing. Nat. Mater. 13(4), 400–408 (2014)
B. Gunjan, M.G. Zaidi, Impact of gold nanoparticles on physiological and biochemical characteristics of Brassica juncea. J. Plant Biochem. Physiol. (2014). https://doi.org/10.4172/2329-9029.1000133
M. Haghighi, Z. Afifipour, M. Mozafarian, The effect of N-Si on tomato seed germination under salinity levels. J. Biol. Environ. Sci. 6(16), 87–90 (2012)
M. Haghighi, R. Abolghasemi, J.A. da Silva, Low and high temperature stress affect the growth characteristics of tomato in hydroponic culture with Se and nano-Se amendment. Sci. Hortic. 178, 231–240 (2014)
M. Haroon, A. Zaidi, B. Ahmed, A. Rizvi, M.S. Khan, J. Musarrat, Effective inhibition of phytopathogenic microbes by eco-friendly leaf extract mediated silver nanoparticles (AgNPs). Indian J. Microbiol. 59(3), 273–287 (2019)
M. Hasanuzzaman, K. Nahar, and M. Fujita, Extreme temperature responses, oxidative stress and antioxidant defense in plants. In Vahadati K. and Leslie C. (eds), Abiotic stress-plant responses and applications in agriculture, Intech, 13, pp.169–205 (2013).
T. Hattori, S. Inanaga, H. Araki, P. An, S. Morita, M. Luxová, A. Lux, Application of silicon enhanced drought tolerance in Sorghum bicolour. Physiol. Plant. 123(4), 459–466 (2005)
L. Heidarvand, R.M. Amiri, M.R. Naghavi, Y. Farayedi, B. Sadeghzadeh, K. Alizadeh, Physiological and morphological characteristics of chickpea accessions under low temperature stress. Russ. J. Plant Physiol. 58(1), 157–163 (2011)
É. Hideg, M.A. Jansen, A. Strid, UV-B exposure, ROS, and stress: Inseparable companions or loosely linked associates? Trends Plant Sci. 18(2), 107–115 (2013)
T. Hirayama, K. Shinozaki, Research on plant abiotic stress responses in the post-genome era: Past, present and future. Plant J. 61(6), 1041–1052 (2010)
S.S. Hojjat, A. Ganjali, The effect of silver nanoparticle on lentil seed germination under drought stress. Int. J. Farm Allied Sci. 5(3), 208–212 (2016)
A. Husen, K.S. Siddiqi, Phytosynthesis of nanoparticles: Concept, controversy and application. Nanoscale Res. Lett. 9(1), 1–24 (2014)
M.M. Hussein, N.H. Abou-Baker, The contribution of nano-zinc to alleviate salinity stress on cotton plants. R. Soc. Open Sci. 5(8), 171809 (2018)
I. Iavicoli, V. Leso, D.H. Beezhold, A.A. Shvedova, Nanotechnology in agriculture: Opportunities, toxicological implications, and occupational risks. Toxicol. Appl. Pharmacol. 329, 96–111 (2017)
E. Ibrahim, M. Zhang, Y. Zhang, A. Hossain, W. Qiu, Y. Chen, Y. Wang, W. Wu, G. Sun, B. Li, Green-synthesization of silver nanoparticles using endophytic bacteria isolated from garlic and its antifungal activity against wheat Fusarium head blight pathogen Fusarium graminearum. Nano 10(2), 219 (2020)
K. Imada, S. Sakai, H. Kajihara, S. Tanaka, S. Ito, Magnesium oxide nanoparticles induce systemic resistance in tomato against bacterial wilt disease. Plant Pathol. 65(4), 551–560 (2016)
M. Iqbal, N.I. Raja, Z.U. Mashwani, M. Hussain, M. Ejaz, F. Yasmeen, Effect of silver nanoparticles on growth of wheat under heat stress. Iran. J. Sci. Technol. Trans. A Sci. 43(2), 387–395 (2019)
A. Jaberzadeh, P. Moaveni, H.R. Moghadam, H. Zahedi, Influence of bulk and nanoparticles titanium foliar application on some agronomic traits, seed gluten and starch contents of wheat subjected to water deficit stress. Notulae Botanicae Horti Agrobotanici Cluj-Napoca 41(1), 201–207 (2013)
S.U. Jalil, M.I. Ansari, Nanoparticles and abiotic stress tolerance in plants: Synthesis, action, and signaling mechanisms. Plant Signal. Mol. 1, 549–561 (2019)
K. Jeyasubramanian, U.U. Thoppey, G.S. Hikku, N. Selvakumar, A. Subramania, K. Krishnamoorthy, Enhancement in growth rate and productivity of spinach grown in hydroponics with iron oxide nanoparticles. RSC Adv. 6(19), 15451–15459 (2016)
M. Kalteh, Z.T. Alipour, S. Ashraf, M. Marashi Aliabadi, and A. Falah Nosratabadi, Effect of silica nanoparticles on basil (Ocimum basilicum) under salinity stress. Journal of Chemical Health Risks, 4(3), pp.49–55 (2014).
M. Kalteh, Z.T. Alipour, S. Ashraf, M. Marashi Aliabadi, and A. Falah Nosratabadi, Effect of silica nanoparticles on basil (Ocimum basilicum) under salinity stress. Journal of Chemical Health Risks, 4(3), pp.49–55 (2018).
A. Kaphle, P.N. Navya, A. Umapathi, H.K. Daima, Nanomaterials for agriculture, food and environment: Applications, toxicity and regulation. Environ. Chem. Lett. 16(1), 43–58 (2018)
T. Karuppanapandian, H.W. Wang, N. Prabakaran, K. Jeyalakshmi, M. Kwon, K. Manoharan, W. Kim, 2, 4-dichlorophenoxyacetic acid-induced leaf senescence in mung bean (Vigna radiata L. Wilczek) and senescence inhibition by co-treatment with silver nanoparticles. Plant Physiol. Biochem. 49(2), 168–177 (2011)
Z. Khan, and H. Upadhyaya, Impact of nanoparticles on abiotic stress responses in plants: an overview. In Tripathi D.K., Ahmad P., Sharma S., Chauhan D.K., Dubey N.K. (eds), Nanomaterials in plants, algae and microorganisms, Academic press, Cambridge, USA, pp.305–322 (2019).
M.N. Khan, F. Mohammad, M. Mobin, M.A. Saqib, Tolerance of plants to abiotic stress: A role of nitric oxide and calcium, in Nitric Oxide in Plants: Metabolism and Role in Stress Physiology, (Springer, Cham, 2014), pp. 225–242
M.N. Khan, M. Mobin, Z.K. Abbas, K.A. AlMutairi, Z.H. Siddiqui, Role of nanomaterials in plants under challenging environments. Plant Physiol. Biochem. 110, 194–209 (2017)
M.V. Khodakovskaya, K. de Silva, D.A. Nedosekin, E. Dervishi, A.S. Biris, E.V. Shashkov, E.I. Galanzha, V.P. Zharov, Complex genetic, photothermal, and photoacoustic analysis of nanoparticle-plant interactions. Proc. Natl. Acad. Sci. 108(3), 1028–1033 (2011)
E. Kohan-Baghkheirati, J. Geisler-Lee, Gene expression, protein function and pathways of Arabidopsis thaliana responding to silver nanoparticles in comparison to silver ions, cold, salt, drought, and heat. Nano 5(2), 436–467 (2015)
L. Kőrösi, B. Pertics, G. Schneider, B. Bognár, J. Kovács, V. Meynen, A. Scarpellini, L. Pasquale, M. Prato, Photocatalytic inactivation of plant pathogenic bacteria using TiO2 nanoparticles prepared hydrothermally. Nano 10(9), 1730 (2020)
A. Kriti, A. Ghatak, N. Mandal, Antimycotic efficacy of Zinc nanoparticle on dark-spore forming Phytopathogenic fungi. J. Pharmacogn. Phytochem. 9(2), 750–754 (2020)
P. Kumar, P. Mahajan, R. Kaur, S. Gautam, Nanotechnology and its challenges in the food sector: A review. Mater. Today Chem. 17, 100332 (2020)
Z. Lei, S. Mingyu, L. Chao, C. Liang, H. Hao, W. Xiao, L. Xiaoqing, Y. Fan, G. Fengqing, H. Fashui, Effects of nanoanatase TiO2 on photosynthesis of spinach chloroplasts under different light illumination. Biol. Trace Elem. Res. 119(1), 68–76 (2007)
Z. Lei, S. Mingyu, W. Xiao, L. Chao, Q. Chunxiang, C. Liang, H. Hao, L. Xiaoqing, H. Fashui, Antioxidant stress is promoted by nano-anatase in spinach chloroplasts under UV-B radiation. Biol. Trace Elem. Res. 121(1), 69–79 (2008)
D. Lin, B. Xing, Phytotoxicity of nanoparticles: Inhibition of seed germination and root growth. Environ. Pollut. 150(2), 243–250 (2007)
D. Lin, B. Xing, Root uptake and phytotoxicity of ZnO nanoparticles. Environ. Sci. Technol. 42(15), 5580–5585 (2008)
Y.F. Liu, M.F. Qi, T.L. Li, Photosynthesis, photoinhibition, and antioxidant system in tomato leaves stressed by low night temperature and their subsequent recovery. Plant Sci. 196, 8–17 (2012)
X. Liu, Y. Ren, Z. Zhou, Y. Wu, C. Zhang, J. Jiang, B. Liu, Effect of nano-ZnO on activity of wheat fusarium head blight pathogen. Pak. J. Bot. 52(5), 1815–1821 (2020)
Z. Luksiene, N. Rasiukeviciute, B. Zudyte, N. Uselis, Innovative approach to sunlight activated biofungicides for strawberry crop protection: ZnO nanoparticles. J. Photochem. Photobiol. B Biol. 203, 111656 (2020)
H. Mahmoodzadeh, M. Nabavi, H. Kashefi, Effect of nanoscale titanium dioxide particles on the germination and growth of canola (Brassica napus). J. Ornam. Hortic. Plants 3(1), 25–32 (2013)
M. Marmiroli, D. Imperiale, L. Pagano, M. Villani, A. Zappettini, N. Marmiroli, The proteomic response of Arabidopsis thaliana to cadmium sulfide quantum dots, and its correlation with the transcriptomic response. Front. Plant Sci. 6, 1104 (2015)
M. Martínez, L. Ramírez Albuquerque, A.F. Arata, F. Biganzoli, V. Fernández Pinto, S.A. Stenglein, Effects of Fusarium graminearum and Fusarium poae on disease parameters, grain quality and mycotoxins contamination in bread wheat (Part I). J. Sci. Food Agric. 100(2), 863–873 (2020)
M.C. Martínez-Ballesta, L. Zapata, N. Chalbi, M. Carvajal, Multiwalled carbon nanotubes enter broccoli cells enhancing growth and water uptake of plants exposed to salinity. J. Nanobiotechnol. 14(1), 1–4 (2016)
D. Martínez-Fernández, M. Vítková, Z. Michálková, M. Komárek, Engineered nanomaterials for phytoremediation of metal/metalloid-contaminated soils: Implications for plant physiology, in Phytoremediation, (Springer, Cham, 2017), pp. 369–403
P. Miralles, T.L. Church, A.T. Harris, Toxicity, uptake, and translocation of engineered nanomaterials in vascular plants. Environ. Sci. Technol. 46(17), 9224–9239 (2012)
S. Mishra, B.R. Singh, A. Singh, C. Keswani, A.H. Naqvi, H.B. Singh, Biofabricated silver nanoparticles act as a strong fungicide against Bipolaris sorokiniana causing spot blotch disease in wheat. PLoS One 9(5), e97881 (2014)
D. Mittal, G. Kaur, P. Singh, K. Yadav, S.A. Ali, Nanoparticle-based sustainable agriculture and food science: Recent advances and future outlook. Front. Nanotechnol. 2, 10 (2020)
A.K. Mohamed, M.F. Qayyum, A.M. Abdel-Hadi, R.A. Rehman, S. Ali, M. Rizwan, Interactive effect of salinity and silver nanoparticles on photosynthetic and biochemical parameters of wheat. Arch. Agron. Soil Sci. 63(12), 1736–1747 (2017)
E.A. Mohamed, S.F. Elsharabasy, D. Abdulsamad, Evaluation of in vitro nematicidal efficiency of copper nanoparticles against root-knot nematode Meloidogyne incognita. South Asian J. Parasitol. 14, 1–6 (2019)
R. Mohammadi, R. Maali-Amiri, N.L. Mantri, Effect of TiO2 nanoparticles on oxidative damage and antioxidant defense systems in chickpea seedlings during cold stress. Russ. J. Plant Physiol. 61(6), 768–775 (2014)
H. Mohammadi, M. Esmailpour, A. Gheranpaye, Effects of TiO2 nanoparticles and water-deficit stress on morpho-physiological characteristics of dragonhead (Dracocephalum moldavica L.) plants. Acta agriculturae Slovenica 107(2), 385–396 (2016)
G. Mustafa, S. Komatsu, Insights into the response of soybean mitochondrial proteins to various sizes of aluminum oxide nanoparticles under flooding stress. J. Proteome Res. 15(12), 4464–4475 (2016)
S. Negrão, S.M. Schmöckel, M. Tester, Evaluating physiological responses of plants to salinity stress. Ann. Bot. 119(1), 1–11 (2017)
I. Ocsoy, M.L. Paret, M.A. Ocsoy, S. Kunwar, T. Chen, M. You, W. Tan, Nanotechnology in plant disease management: DNA-directed silver nanoparticles on graphene oxide as an antibacterial against Xanthomonas perforans. ACS Nano 7(10), 8972–8980 (2013)
R.A. Omar, S. Afreen, N. Talreja, D. Chauhan, M. Ashfaq, Impact of nanomaterials in plant systems, in Plant Nanobionics, (Springer, Cham, 2019), pp. 117–140
N.M. Palmqvist, G.A. Seisenbaeva, P. Svedlindh, V.G. Kessler, Maghemite nanoparticles acts as nanozymes, improving growth and abiotic stress tolerance in Brassica napus. Nanoscale Res. Lett. 12(1), 1–9 (2017)
K. Pandey, M. Anas, V.K. Hicks, M.J. Green, M.V. Khodakovskaya, Improvement of commercially valuable traits of industrial crops by application of carbon-based nanomaterials. Sci. Rep. 9(1), 1–4 (2019)
M.L. Paret, G.E. Vallad, D.R. Averett, J.B. Jones, S.M. Olson, Photocatalysis: Effect of light-activated nanoscale formulations of TiO2 on Xanthomonas perforans and control of bacterial spot of tomato. Phytopathology 103(3), 228–236 (2013)
A.K. Parida, A.B. Das, Salt tolerance and salinity effects on plants: A review. Ecotoxicol. Environ. Saf. 60(3), 324–349 (2005)
S. Pei, J. Zhao, J. Du, W. Ren, H.M. Cheng, Direct reduction of graphene oxide films into highly conductive and flexible graphene films by hydrohalic acids. Carbon 48(15), 4466–4474 (2010)
A. Pérez-de-Luque, Interaction of nanomaterials with plants: What do we need for real applications in agriculture? Front. Environ. Sci. 10(5), 12 (2017)
M. Qi, Y. Liu, T. Li, Nano-TiO2 improve the photosynthesis of tomato leaves under mild heat stress. Biol. Trace Elem. Res. 156(1), 323–328 (2013)
S. Rajeshkumar, Antifungal impact of nanoparticles against different plant pathogenic fungi. In Nanomaterials in Plants, Algae and Microorganisms, Elsevier (pp. 197–217). (2019) Academic Press.
N. Regier, C. Cosio, N. von Moos, V.I. Slaveykova, Effects of copper-oxide nanoparticles, dissolved copper and ultraviolet radiation on copper bioaccumulation, photosynthesis and oxidative stress in the aquatic macrophyte Elodea nuttallii. Chemosphere 128, 56–61 (2015)
L. Rossi, W. Zhang, L. Lombardini, X. Ma, The impact of cerium oxide nanoparticles on the salt stress responses of Brassica napus L. Environ. Pollut. 219, 28–36 (2016)
A. Roychoudhury, Silicon-nanoparticles in crop improvement and agriculture. International Journal on Recent Advancement in Biotechnology & Nanotechnology [ISSN: 2582–1571 (online)], 3(1), pp.54–65 (2020).
B. Ruttkay-Nedecky, O. Krystofova, L. Nejdl, V. Adam, Nanoparticles based on essential metals and their phytotoxicity. J. Nanobiotechnol. 15(1), 1–9 (2017)
V. Saharan, G. Sharma, M. Yadav, M.K. Choudhary, S.S. Sharma, A. Pal, R. Raliya, P. Biswas, Synthesis and in vitro antifungal efficacy of Cu–chitosan nanoparticles against pathogenic fungi of tomato. Int. J. Biol. Macromol. 75, 346–353 (2015)
M. Savicka, N. Škute, Effects of high temperature on malondialdehyde content, superoxide production and growth changes in wheat seedlings (Triticum aestivum L.). Ekologija 56(1), 26–33 (2010)
M. Sedghi, M. Hadi, S.G. Toluie, Effect of nano zinc oxide on the germination parameters of soybean seeds under drought stress. Ann. West Univ. Timis. Ser. Biol. 16(2), 73 (2013)
M.A. Shallan, H.M. Hassan, A.A. Namich, A.A. Ibrahim, Biochemical and physiological effects of TiO2 and SiO2 nanoparticles on cotton plant under drought stress. Res. J. Pharma. Biol. Chem. Sci. 7(4), 1540–1551 (2016)
P. Sharma, A.B. Jha, R.S. Dubey, M. Pessarakli, Reactive oxygen species, oxidative damage, and antioxidative defense mechanism in plants under stressful conditions. J. Bot. 2012, 1–27 (2012)
X. Shen, Y. Zhou, L. Duan, Z. Li, A.E. Eneji, J. Li, Silicon effects on photosynthesis and antioxidant parameters of soybean seedlings under drought and ultraviolet-B radiation. J. Plant Physiol. 167(15), 1248–1252 (2010)
M.H. Siddiqui, M.H. Al-Whaibi, Role of nano-SiO2 in germination of tomato (Lycopersicum esculentum seeds Mill.). Saudi J. Biol. Sci. 21(1), 13–17 (2014)
M.H. Siddiqui, M.H. Al-Whaibi, F. Mohammad, Nanotechnology and Plant Sciences, vol 10 (Springer International Publishing Switzerland, 2015), pp. 978–973
D.F. Simon, R.F. Domingos, C. Hauser, C.M. Hutchins, W. Zerges, K.J. Wilkinson, Transcriptome sequencing (RNA-seq) analysis of the effects of metal nanoparticle exposure on the transcriptome of Chlamydomonas reinhardtii. Appl. Environ. Microbiol. 79(16), 4774–4785 (2013)
S. Singh, A. Husen, Behavior of agricultural crops in relation to nanomaterials under adverse environmental conditions, in Nanomaterials for Agriculture and Forestry Applications, (Elsevier, 2020), pp. 219–256
J. Singh, B.K. Lee, Influence of nano-TiO2 particles on the bioaccumulation of Cd in soybean plants (Glycine max): A possible mechanism for the removal of Cd from the contaminated soil. J. Environ. Manag. 170, 88–96 (2016)
S. Singh, D.K. Tripathi, N.K. Dubey, D.K. Chauhan, Effects of nano-materials on seed germination and seedling growth: Striking the slight balance between the concepts and controversies. Mater. Focus 5(3), 195–201 (2016)
W. Tan, W. Du, A.C. Barrios, R. Armendariz Jr., N. Zuverza-Mena, Z. Ji, C.H. Chang, J.I. Zink, J.A. Hernandez-Viezcas, J.R. Peralta-Videa, J.L. Gardea-Torresdey, Surface coating changes the physiological and biochemical impacts of nano-TiO2 in basil (Ocimum basilicum) plants. Environ. Pollut. 222, 64–72 (2017)
N. Taran, V. Storozhenko, N. Svietlova, L. Batsmanova, V. Shvartau, M. Kovalenko, Effect of zinc and copper nanoparticles on drought resistance of wheat seedlings. Nanoscale Res. Lett. 12(1), 1–6 (2017)
Torabian, S., Zahedi, M. And Khoshgoftar, A. H. (2017) ‘Effects of foliar spray of nano-particles of FeSO4 on the growth and ion content of sunflower under saline condition’, J. Plant Nutr., 40(5):615–623
D.K. Tripathi, V.P. Singh, S.M. Prasad, D.K. Chauhan, N.K. Dubey, Silicon nanoparticles (SiNp) alleviate chromium (VI) phytotoxicity in Pisum sativum (L.) seedlings. Plant Physiol. Biochem. 96, 189–198 (2015)
D.K. Tripathi, S. Singh, V.P. Singh, S.M. Prasad, N.K. Dubey, D.K. Chauhan, Silicon nanoparticles more effectively alleviated UV-B stress than silicon in wheat (Triticum aestivum) seedlings. Plant Physiol. Biochem. 110, 70–81 (2017)
B.C. Tripathy, R. Oelmüller, Reactive oxygen species generation and signaling in plants. Plant Signal. Behav. 7(12), 1621–1633 (2012)
D. Van Nguyen, H.M. Nguyen, N.T. Le, K.H. Nguyen, H.T. Nguyen, H.M. Le, A.T. Nguyen, N.T. Dinh, S.A. Hoang, C. Van Ha, Copper nanoparticle application enhances plant growth and grain yield in maize under drought stress conditions. J. Plant Growth Regul. 2, 1–2 (2021)
G.L. Vanti, S. Masaphy, M. Kurjogi, S. Chakrasali, V.B. Nargund, Synthesis and application of chitosan-copper nanoparticles on damping off causing plant pathogenic fungi. Int. J. Biol. Macromol. 156, 1387–1395 (2020)
P. Venkatachalam, M. Jayaraj, R. Manikandan, N. Geetha, E.R. Rene, N.C. Sharma, S.V. Sahi, Zinc oxide nanoparticles (ZnO NPs) alleviate heavy metal-induced toxicity in Leucaena leucocephala seedlings: A physiochemical analysis. Plant Physiol. Biochem. 110, 59–69 (2017)
M.L. Verma, P. Kumar, D. Sharma, A.D. Verma, A.K. Jana, Advances in nanobiotechnology with special reference to plant systems, in Plant Nanobionics, (Springer, Cham, 2019), pp. 371–387
A. Wahid, Physiological implications of metabolite biosynthesis for net assimilation and heat-stress tolerance of sugarcane (Saccharum officinarum) sprouts. J. Plant Res. 120(2), 219–228 (2007)
Y. Wang, S.G. Kim, S.T. Kim, G.K. Agrawal, R. Rakwal, K.Y. Kang, Biotic stress-responsive rice proteome: An overview. J. Plant Biol. 54(4), 219–226 (2011)
G. Wang, S. Deng, C. Li, Y. Liu, L. Chen, C. Hu, Damage to DNA caused by UV-B radiation in the desert cyanobacterium Scytonema javanicum and the effects of exogenous chemicals on the process. Chemosphere 88(4), 413–417 (2012)
S.H. Wani, V. Kumar, V. Shriram, S.K. Sah, Phytohormones and their metabolic engineering for abiotic stress tolerance in crop plants. Crop J. 4(3), 162–176 (2016)
H. Wei, E. Wang, Nanomaterials with enzyme-like characteristics (nanozymes): Next-generation artificial enzymes. Chem. Soc. Rev. 42(14), 6060–6093 (2013)
S. Yadav, P. Modi, A. Dave, A. Vijapura, D. Patel, and M. Patel, Effect of abiotic stress on crops. In Hasanuzzaman M., Filho M.C.M.T., Nogueira T.A.R. (eds), Sustainable Crop Production, IntechOpen, London, UK (2020).
F. Yang, F. Hong, W. You, C. Liu, F. Gao, C. Wu, P. Yang, Influence of nano-anatase TiO2 on the nitrogen metabolism of growing spinach. Biol. Trace Elem. Res. 110(2), 179–190 (2006)
Y. Ze, C. Liu, L. Wang, M. Hong, F. Hong, The regulation of TiO2 nanoparticles on the expression of light-harvesting complex II and photosynthesis of chloroplasts of Arabidopsis thaliana. Biol. Trace Elem. Res. 143(2), 1131–1141 (2011)
L.W. Zhang, N.A. Monteiro-Riviere, Mechanisms of quantum dot nanoparticle cellular uptake. Toxicol. Sci. 110(1), 138–155 (2009)
L. Zhao, B. Peng, J.A. Hernandez-Viezcas, C. Rico, Y. Sun, J.R. Peralta-Videa, X. Tang, G. Niu, L. Jin, A. Varela-Ramirez, J.Y. Zhang, Stress response and tolerance of Zea mays to CeO2 nanoparticles: Cross talk among H2O2, heat shock protein, and lipid peroxidation. ACS Nano 6(11), 9615–9622 (2012)
L. Zhao, L. Lu, A. Wang, H. Zhang, M. Huang, H. Wu, B. Xing, Z. Wang, R. Ji, Nano-biotechnology in agriculture: Use of nanomaterials to promote plant growth and stress tolerance. J. Agric. Food Chem. 68(7), 1935–1947 (2020)
Das, A. and Das, B., 2019. Nanotechnology a potential tool to mitigate abiotic stress in crop plants. In De Oliveira A. (eds), Abiotic and Biotic Stress in Plants, IntechOpen. Hasanuzzaman, M., Nahar, K. and Fujita, M., 2013.
Extreme temperature responses, oxidative stress and antioxidant defense in plants. In Vahadati K. and Leslie C. (eds), Abiotic stress-plant responses and applications in agriculture, Intech, 13, pp.169-205. Khan, Z. and Upadhyaya, H., 2019.
Impact of nanoparticles on abiotic stress responses in plants: an overview. In Tripathi D.K., Ahmad P., Sharma S., Chauhan D.K., Dubey N.K. (eds), Nanomaterials in plants, algae and microorganisms, Academic press, Cambridge, USA, pp.305-322.
Yadav, S., Modi, P., Dave, A., Vijapura, A., Patel, D. and Patel, M., 2020. Effect of abiotic stress on crops. In Hasanuzzaman M., Filho M.C.M.T., Nogueira T.A.R. (eds), Sustainable Crop Production, IntechOpen, London, UK.
Author information
Authors and Affiliations
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2022 The Author(s), under exclusive license to Springer Nature Switzerland AG
About this chapter

Cite this chapter
Chandel, S.S., Shree, B., Mondal, K., Mali, S. (2022). Mechanism of Nanoparticles-Mediated Alleviating Biotic and Abiotic Stresses in Agricultural Crops: Recent Advances and Future Perspectives. In: Rajput, V.D., Verma, K.K., Sharma, N., Minkina, T. (eds) The Role of Nanoparticles in Plant Nutrition under Soil Pollution. Sustainable Plant Nutrition in a Changing World. Springer, Cham. https://doi.org/10.1007/978-3-030-97389-6_6
Download citation
DOI: https://doi.org/10.1007/978-3-030-97389-6_6
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-030-97388-9
Online ISBN: 978-3-030-97389-6
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)