
Abstract
Ushuaia, a coastal city located in Patagonia, Argentina, amid unique natural habitats such as sub-antarctic Nothofagus forests and Sphagnum peatlands, preserves green areas of natural ecosystems within the urban matrix. Forests and peatlands have persisted as natural islands of different sizes, conservation status and anthropic disturbances. These urban green areas are underappreciated and administratively abandoned, and several threats could lead to their irreversible transformation (e.g., replacement by new neighborhoods, urban waste deposits). However, natural green urban areas can serve as biodiversity islands that make substantial contributions to conserve local biota and also to house new species assemblages (different from those in pristine areas), bringing native forest and peatland species closer to the local residents and visitors. These green urban islands can also help to preserve traditional cultural values, enhance education and generate local touristic attractions. In this chapter, we assessed the assemblages of plants and birds in urban native forests, and plants and insects in urban peatlands of Ushuaia city considering two levels of urbanization surrounding these green areas, and analyzed their variations compared with assemblages in nearby similar non-urban (low level of disturbance) ecosystems. Despite some differences in richness, abundance, biodiversity indices, and species composition of vascular plants, birds and insects, natural urban and peri-urban forests and peatlands still conserve several species and characteristics similar to those of unmodified ecosystems. However, the presence of new species, mainly plants, introduced into these urban patches has modified the original communities, establishing new assemblages that may contribute positively to urban biodiversity islands.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others

Keywords
1 Introduction
Cities located in mountainous regions and coastal areas are one of the most attractive destinations for tourism. These areas are still perceived as natural treasures, offering striking landscapes and a wide variety of highly attractive plant and animal species. In Patagonia, one of the main attractions are the pristine landscapes surrounding the cities and the possibility to be in close contact with wildlife through the broad spectrum of existing opportunities for outdoor activities. In addition, natural areas within the urban matrix (i.e., parks or wild areas containing original vegetation) are of great value to residents and visitors, as urban green areas provide a number of ecosystem services to the community. These ecosystem services include regulating (e.g., noise reduction, modulation of temperature, removal of air pollution, protection of water quality), supporting (e.g., increased biodiversity, habitat, soil formation and storage and cycling of nutrients), cultural (recreation, enhancement of property value, community cohesion, source of knowledge), and provisioning (e.g., food, water, fuel) services (Millennium Ecosystem Assessment 2005; Gómez-Baggethun and Barton 2013). To benefit from ecological services provided by urban green areas it is necessary to incorporate these overlooked areas in the urban planning of our cities. The places where people live and work need to be designed so that they can serve to promote contact with the natural world (Miller 2005). Even those areas whose original biological composition has been profoundly modified are still important for people. The benefits provided by green urban areas, even those that are somewhat altered, are invaluable (Thomas and Geller 2013).
1.1 Biodiversity in Urban Green Areas
From the perspective of island biogeography (MacArthur and Wilson 1967), urban green areas can be considered as a type of ecological island more or less isolated from the other green habitats by the surrounding urban landscape (McGregor-Fors et al. 2011). Urban green areas, even with some levels of disturbance, are considered to function as biodiversity islands providing potential habitat refuge for different organisms within the urban matrix (Matthies et al. 2017; Montagnini et al. 2022). They are also important in maintaining key ecosystem services (e.g., water purification and regulation) which are necessary for human well-being and public health in cities (Gómez-Baggethun and Barton 2013). Biodiversity is essential for maintaining ecosystem function (Maestre et al. 2012; Reich et al. 2012) therefore it is important to maintain urban green areas as undisturbed as possible.
Plants play a significant role in animal diversity because they constitute the first trophic level, i.e. they are the primary producers of energy that is used by other members of the food chain. Poorly managed urban green areas have lower biodiversity compared to undisturbed ones (Burghardt et al. 2010; Tallamy et al. 2010). Similarly, poorly managed landscapes can promote the introduction of non-native plants (e.g., Rozzi et al. 2003; Marzluff et al. 2008; Ceplová et al. 2017; Lencinas et al. 2017) which can have negative effects on native plant diversity and microhabitat conditions (Ceplová et al. 2017). Non-native plants sustain significantly fewer insects (e.g., caterpillars) than native-plants (Tallamy et al. 2010). Insects play a significant role in supporting further biodiversity because they are eaten by other animals (e.g., birds) and represent the main energy source in critical stages of the life cycle of many species (e.g., many birds feed their hatchlings with insects). Furthermore, insects are involved in soil formation and organic matter decomposition, which affect both the diversity and structure of the vegetation (Hartley and Jones 2008).
The integration of green urban areas for biodiversity conservation in city planning is among the most difficult problems to resolve by public and local policies (Murphy 1999; Nilon et al. 2017). Even the effort to keep the native biota intact in urban green areas may represent an unlikely goal to achieve for local administrations. This is the case of young and developing cities which suffer from intense and regular disturbances (e.g., constant building, new streets) which significantly alter local biota in green areas (Lososová et al. 2011). In contrast, green areas in less urbanized habitats (e.g., those that suffer irregular and less strong disturbances) support rich species diversity and native biota (Zerbe et al. 2003; Ceplová et al. 2017).
In order to identify conservation priorities and/or prevent future environmental problems in cities it is necessary to understand how urbanization influences different biological groups. It is important to assess the potential role of biodiversity islands to conserve local biota and ultimately avoid its contamination, transformation and total loss inside an urban matrix.
1.2 Case Study: Ushuaia City and its Urban Green Areas
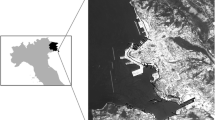
Ushuaia, a coastal city founded in 1884, is one of the southermost urban settlements in the world, and the most populated city (56,900 inhabitants, INDEC 2010) at such latitude. It is the political and administrative capital of the Argentinian province of Tierra del Fuego, Antarctica and Islands of the South Atlantic, located in the biogeographic region of the Andean-Patagonian forest (Fig. 16.1.)

Map of Tierra del Fuego, Argentina, showing Ushuaia city, which is located on the coastal Nothofagus forests of the Beagle Channel
Ushuaia’s urban development has had a short and hurried history since its foundation, marked by a penal colony established in 1896 (closed down in 1947) that started to bring people to the city, and by the Industrial Promotion Law (1972) that stimulated regional economic development. During 1972–1990, the city underwent a population boom, which tripled its number of residents and expanded the urban area into the surrounding natural forests to settle new neighborhoods. After the 1990s, the unexpected increase of the population strongly promoted building-up areas (houses, schools, etc.) and installing public services, which reduced the forested areas around the unplanned urban layout, a situation that continues in present days. Therefore, existing forest patches inside the city are currently remnants of old or secondary native forests (~50 years old) originated from old cuts or fires (Fig. 16.2). The urbanization growth also gradually surrounded and isolated the existing peatlands within the city (Fig. 16.2). Small peatlands (<0.3 ha) were filled with soil, rubble and rocks, and therefore irreversibly transformed to building areas. But the larger ones, deemed non-suitable for building were left as “brown areas” immersed in the urbanization, exposed to degrading uses (e.g., substrate extraction, artificial drainage, urban waste accumulation, refuge of abandoned domestic animals, human trespassing). A similar situation is currently found in some patches of urban forests, which are also used to install gardens or squares, or exceptionally preserved areas (e.g., the Parque Yatana Urban Reserve, located in the center of Ushuaia).

Inside view from urban Nothofagus forests (a) and Sphagnum peatlands (b) in Ushuaia city, Tierra del Fuego, Argentina. (Photos: Julieta Benítez and Francisco Sola)
Due to the unplanned nature of the city’s sprawl, recreational and protected areas are also unplanned and scarce, resulting in the misuse and/or overuse of pre-existent natural patches inside and outside the city. This is the consequence of a cultural and social necessity, where the lack of accessible leisure outlets is compensated with available natural resources, resulting in its unregulated use and eventual deterioration. There is still the possibility to restore these sites as natural biodiversity refuges while still making them available to the public under sustainable conditions. Although Ushuaia city is surrounded by natural ecosystems, the natural green areas in the most densely urbanized area barely reach 10% of the surface. While these forests and peatlands inside the urban area were not created consciously to increase the proportion of green areas available to residents, they can provide the areas of green spaces that are necessary for a healthier and more sustainable city.
Unfortunately, these urban natural green areas are underappreciated and administratively abandoned. Moreover, we have a limited understanding of how green spaces are used for refuge, foraging and reproduction by different animal species. Urban green areas, especially if they are patches of pre-existing native forests and peatlands, retain part of the local diversity. In this chapter we aim to determine the role of natural urban green areas as biodiversity islands within Ushuaia city as a case study from Patagonia, Argentina. We present results of studies on species assemblages (species richness, relative abundance, diversity, evenness and similarity indices) of understory vascular plants and birds in forests, and vascular plants and insects in peatlands of Ushuaia city. We also compare species composition of patches inside the urban matrix (two levels of urbanization surrounding these green areas) with similar and near undisturbed patches outside the urban matrix (non-urban patches). We propose these natural urban islands could potentially serve as places to promote conservation of local biodiversity, bringing native forest and peatland species closer to people. This can contribute to biodiversity conservation in a broad sense, which could also help to preserve traditional cultural values, improve education and enhance touristic attractions.
2 Methods
2.1 Study Area
Ushuaia city is located in the north bank of the Beagle Channel, Argentina (Figs. 16.1 and 16.3) in the southern end of the South American Cone (54°48′S, 68°19′W). Mountains and glaciers surround the city, and the natural vegetation is primarily comprised of forests that reach the coast. These native forests were the main source of firewood and timber since the city foundation. Most of them are deciduous forests dominated by Nothofagus pumilio (lenga, Nothofagaceae), which develop mainly in mountain slopes and valleys. Mixed deciduous-evergreen forests of lenga and N. betuloides (guindo) also occur on the coast and mid slopes and include other woody species such as Drymis winterii (canelo), Embothrium coccineum (notro) and Maytenus magellanica (leña dura) as small trees in the understory (Moore 1983). On the other hand, N. antarctica (ñire) is found surrounding peatbogs and inhabiting mixed coastal forests. The vascular plants in the understory usually include few shrubs (e.g., Berberis and Ribes) that produce berries, and abundant forbs, graminoids, fungi, mosses, liverworts and ferns. In the whole landscape, forest masses are intermingled with peatlands and grasslands of different size and composition according to elevation above sea level. Peatlands formed by accumulation of Sphagnum spp. mosses during geological times support other species of vascular plants like Cyperaceae (e.g., Carex spp.), Astelia sp., and Drosera sp. (Moore 1983). The climate in Ushuaia city is sub-antarctic (Tuhkanen 1992) with short, cold summers (7 °C average temperature) and long winters (−1 °C average temperature) with regular snow and frost. Mean annual rainfall is 611 mm near the coast and more than 1000 mm in the mountains.

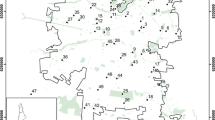
Location of the study area in Ushuaia city, Tiera del Fuego, Argentina. Circles = forest patches, squares = peatlands. The color indicates the urban categories studied: urban (white), peri-urban (gray) and non-urban (black)
For our study, we classified forests and peatlands in Ushuaia city according to the current degree of surrounding urbanization (i.e., their respective locations in the urban matrix): (1) Urban: patches inside the urban matrix, 100% surrounded by buildings, streets or waste disposal areas; (2) Peri-urban: peripheral patches but still inside the urban matrix, and partially surrounded (approximately 50%) by the urban matrix; and (3) Non-urban: patches outside the urban matrix (i.e., without direct contact with urbanization but still close to the city), with low-impact recreational use (Fig. 16.3). The degree of urbanization is also determined by the main anthropic uses of such natural habitats, the use frequency or intensity, and consequently, the potential pollution and habitat degradation (Table 16.1).
2.2 Sampling Design: Surveys of Plant, Bird and Insect Species
For each forest type, we selected four forest patches (replicates) located at the same elevation range (38 to 170 m above sea level (m.a.s.l.). These were at least 150 m apart from each other and from other types of structures, as well as at least 50 m from the edge of the same forest type, in order to avoid the influence of other environments. In the case of peatlands, we selected 12 urban, 4 peri-urban and 4 non-urban locations (elevation range 20 to 200 m.a.s.l.) separated by at least 150 m from each other.
Vegetation surveys were carried out in each patch (forests and peatlands) during 2016–2017, using a circular plot of approximately 30 m radius centered in each sampling site. In each plot, we identified vascular plants (Dicotyledonae, Monocotyledonae and Pteridophytae) at the species level (Moore 1983; Correa 1969–1998). We estimated ground cover of vascular plants using a modification of the relevé method of Braun-Blanquet proposed by Pauchard et al. (2000). The relevé method involves describing recognizable units in the vegetation of a region by the characterization of the vegetation in a single representative standard plot—a relevé—within each unit. The relevés from many units are then analyzed to develop descriptions and classifications of the vegetation in the study region and have been increasingly used in other kinds of vegetation studies as a practical, relatively fast means of collecting information on vegetation (Minnesota Department of Natural Resources 2013). Ground cover was estimated for each species separately and then values were added to obtain family and total vascular plant cover. The complementary covers to reach 100% ground cover (data not shown) were: bryophyte (mosses and liverworts), woody debris (> 3 cm diameter), and bare soil including litter. Species richness was calculated as the total number of vascular plant species identified in each replica of treatments. Voucher specimens were deposited in the Herbarium of Tierra del Fuego at the Centro Austral de Investigaciones Científicas (Austral Center of Scientific Research, CADIC-CONICET) in Ushuaia, Argentina.
Bird surveys in forest patches were conducted during the 2016–2017 breeding season in a 4-h period at sunrise, always under similar climatic conditions (i.e., avoiding rainy or foggy days). We identified bird assemblages by using the point-count method with unlimited distances, which had been used widely in previous studies in Tierra del Fuego (see Deferrari et al. 2001; Lencinas et al. 2005, 2014, 2019). We selected one observation point in each forest patch (n = 12), with two observers during each survey conducting direct sightings (with binoculars) during 13 min of effective observation period (leaving 2 min prior to recording to allow for birds’ habituation to the observers). We registered the birds to the species level according to the classification proposed by Remsen et al. (2019). From these surveys, we calculated bird species richness and relative frequency (number of individuals). Forests patches were revisited three times per month for bird observations during the complete breeding season (from October to February). Species richness represent the total number of species recorded during that period, and the relative abundance was calculated as the sum of the total number of individuals observed for the same species during visits.
Epigean insects were collected in peatlands during January–February 2016. Sampling was done using pitfall traps (plastic containers 12 cm in diameter and 14 cm height) buried and filled to 1/3 of their volume with soapy water (300 ml) to trap and kill arthropods which fell in. We used pitfall traps in sets of five in each plot, and contents of the five traps in a plot were pooled and used as a single sample in order to calculate total capture per sample (Lencinas et al. 2014). Traps were arranged placing one at the center and the remaining four at 5 m from the first, and at 90 degrees from each other, and were left open at ground level for one week before being collected. We obtained 20 samples from 100 pitfall traps. All samples were taken to the laboratory for specimen identification and quantification. Identifications were performed under a binocular dissecting microscope to genus or species level when possible (Roig-Juñent et al. 2002; Marvaldi and Lanteri 2005; Posadas 2012). Due to a lack of complete taxonomic data on Patagonian beetles, some specimens could not be determined to the species level. We employed the recognizable taxonomic unit or morphospecies concept (Oliver and Beattie 1993; Gerlach et al. 2013) when the former could not be determined (hereafter, “species”). This has been demonstrated to be a good tool for insect diversity studies in Patagonian ecosystems, such as Nothofagus forests (Lencinas et al. 2014; Sola et al. 2016; Cárcamo et al. 2019).
2.3 Statistical Analyses
Data collected were used to determine species richness, relative abundance (ground cover for plants, and number of individuals for birds and insects), Shannon-Wiener diversity (H′) and Pielou evenness (J’) indices of plants and birds in each forest, and plants and insects in each peatland types (Pielou 1975). We used one-way ANOVAs after validation of the statistical assumptions of homoscedasticity (homogeneity) and normality, to evaluate species richness, relative abundance, Shannon-Wiener diversity and Pielou evenness indices of vascular plants, birds and insects, considering the category of forests and peatlands (urban, peri-urban and non-urban) as the main factor.
Also, differences in assemblage composition of plants, birds and insects along the urban categories were examined by the non-Metrical Multidimensional Scaling (NMDS) ordination method using a manual methodology, with a Bray-Curtis distance and 250 iterations. A Monte Carlo test was used to evaluate stress in randomized data; probability was presented for each axis.
Finally, we calculated the abundance-based Chao-Sørensen similarity index (Chao et al. 2005) to determine differences among categories of urbanization, both for forests and peatlands, analyzing each taxonomic group separately. This index, varying between zero and one, considers explicitly the relative abundance of both common and rare species, and estimates the extent of shared species taking into account unseen shared species, based on the number of observed rare (singletons and doubletons, as well as unique and duplicate), shared species between two sites. The use of this index is recommended when study sites could be under-sampled and contain only a substantial fraction of the rare species (Chao et al. 2005). We used the software EstimateS (Colwell 2009) for each pairwise evaluation and compared averages and standard deviation for each urbanization category.
3 Results and Discussion
Plants, birds and insects have been widely used as bio-indicators of human impact on natural ecosystems (Lencinas et al. 2005, 2014, 2017; Gerlach et al. 2013; Rozzi and Jiménez 2014; Contador et al. 2015) and urban biodiversity islands (Noreika et al. 2015; Matthies et al. 2017). These three groups, due to high latitude and extreme climatic conditions of Southern Patagonia, sustain the greatest biological diversity in the natural ecosystems, with many endemic species and guilds (Roig-Juñent et al. 2002; Díaz 2012; Rozzi and Jiménez 2014; Cárcamo et al. 2019). Plants are among the more characteristic and recognized organisms for people, both in forests and peatlands, while birds are the most conspicuous groups in forests, and arthropods are more visible and easily distinguishable in peatlands (open areas).
3.1 Vascular Plants
In Ushuaia city, vascular plants were affected differently in forest patches and peatlands. Species richness did not strongly differ along the urbanization categories, but the relative abundance of vegetation and species diversity indexes were strongly modified in urban and peri-urban peatlands (Table 16.2). Urbanization significantly increased the relative abundance of vascular plants, due to the increase of some plant species favored by microclimatic modifications (e.g., light, soil moisture). According to our data, forbs and graminoids such as Gunnera magellanica, Acaena magellanica, and Carex sp. doubled their cover, while other species (e.g., the native fern Blechnum penna-marina and the exotic herbs Achilea millefolium, Rumex crispus, Taraxacum officinale, Poa annua), which were absent in peri-urban and non-urban peatlands, entered in the urban ones. Soil drainage, building and protection against extreme temperatures and winds could favor herbaceous development on peatlands (Pinceloup et al. 2020).
On the other hand, urban and peri-urban forests showed changes in their community composition in some patches (Fig. 16.4a), by the introduction of exotic cultivated shrubs (Ribes idaeus and Ribes rubrum) and potentially invasive forbs (Ranunculus acris and Hypochaeris radicata), which could be intentionally planted by people or escaped from home gardens, orchards and parks. Urbanization also differently influenced the Shannon diversity and Pielou evenness indices for vascular plants in forests patches and peatlands. While both indices were quite similar in forest patches along the urban categories, Shannon diversity and Pielou evenness indices of urban peatlands decreased by half compared to those in non-urban peatlands. Plant composition in urban peatlands was similar to vegetation in peri-urban areas but differed from non-urban peatlands located far from urbanization (Fig. 16.4b).

Results of analysis of differences in assemblage composition of plants examined by the Non-Metrical Multidimensional Scaling (NMDS) ordination method, based on understory plants composition in different urban categories of forest patches (a) and peatlands (b)
3.2 Birds
With respect to birds, urban assemblages in Ushuaia’s forests are conformed by bird species commonly observed in Nothofagus forests (Deferrari et al. 2001; Lencinas et al. 2005), which correspond to thirteen families of Passeriformes, and one family of five orders (Charadriiforme, Falconiforme, Pelecaniforme, Piciforme, Psittaciforme (Benítez et al. 2020). Some species detected in these urban forests are endemic to Patagonia (i.e. Aphrastura spinicauda, Phrygilus patagonicus, Pygarrichas albogularis, Enicognathus ferrugineus and Campephillus magellanicus) (Deferrari et al. 2001). Insectivore and omnivore birds are the most important groups, being more than a half of all resident species (Benítez et al. 2020).
In our study, species richness remained similar in peri-urban and non-urban patches (Table 16.2). However, bird abundance significantly increased in urban forests, with slight changes in peri-urban and non-urban forest patches. This was explained by the high abundance of generalist bird species and/or the occurrence of non-specialist forest birds in altered urban patches (pers. obs.). We observed that urban forests offered different food sources (e.g., waste, fruits/seeds) and nesting alternatives (e.g., open places, canopy openings) compared to native forests, which coincides with other studies (Marzluff and Rodewald 2008). This favored the occurrence of bird species from open habitats such as Vanellus chilensis (Marín 2014).
The availability of extra food and greater hunting possibilities favored the occurrence of scavenging raptors such the Milvago chimango, as was also reported by Biondi et al. (2005). Moreover, urbanization in Ushuaia increased the frequency of exotic bird species (e.g., Passer domesticus). Contrary to this, native birds such as woodpeckers and hole-nesters birds (C. magellanicus, P. albogularis, Troglodytes aedon and A. spinicauda) showed an increasing trend in richness and abundance from the urban to the periphery environments. These charismatic species prefer forests with dense vegetation (Vergara and Schlatter 2006), but they can be seen while feeding in urban habitats (pers. obs.). Analyzing the whole species assemblages of urban forests (Fig. 16.5a), great differences in biodiversity along the urbanization categories were possible to distinguish. Plant and bird communities in urban forests differed from the communities of non-urban forests (Figs. 16.4a and 16.5a).

Results of analysis of differences in bird abundance in different urban forests (a), and insect abundance in different urban peatlands (b), examined by the Non-Metrical Multidimensional Scaling (NMDS) ordination method
3.3 Invertebrates
Peatland invertebrate communities are almost unknown, and even more the effects that anthropic disturbances have on their diversity. Recent surveys in urban and non-urban peatlands of Ushuaia city (Sola et al. 2018) indicate the presence of a wide variety of invertebrates: snails (Gastropoda), spiders (Araneae), and harvestmen (Opiliones), mites (Acari), woodlice (Isopoda), and many orders of insects. Due to their diversity, abundance, easy sampling, and rapid response to environmental changes, insects are useful bio-indicators of habitat degradation (Bolger et al. 2000; Gerlach et al. 2013; Moreno et al. 2013), and have been widely used to study the effects of habitat modification even in south Patagonia (Sola et al. 2016; Lencinas et al. 2014, 2017, 2019; Cárcamo et al. 2019). In particular, the study of ground dwelling insects has important functional implications because of their ecological roles, as they include predators, scavengers, and herbivores.
According to our results, the main insect orders present in all categories included beetles (Coleoptera), moths (Lepidoptera), true bugs (Heteroptera), springtails (Collembola), leafhoppers (Auchenorrhyncha), and wasps (Hymenoptera). Insect richness in peatlands (including Coleoptera, Lepidoptera, Heteroptera, Auchenorrhyncha, Collembola and Hymenoptera) remained similar throughout the urbanization categories (Table 16.2). However, insect abundance significantly decreased in urban peatlands compared with those in non-urban; while peri-urban peatlands showed intermediate values. The most affected groups were Coleoptera, Auchenorrhyncha and Collembola, which reduced by half the number of individuals within these orders. All three of these groups are weaker fliers (in the case of beetle species sampled), making them more susceptible to local impacts. Furthermore, Auchenorrhyncha are herbivorous and although the ecology of local species is unknown, they could be susceptible to changes observed in local flora at each site. Collembolans, as other detritivores, can be associated with available organic matter; a fall in their abundance could be indicative of reduced productivity, and may also result in slowed decomposition rates and nutrient cycling, essential in these cold, high latitude landscapes. Finally, beetles sampled fell under the categories of scavengers or predators (e.g., Curculionidae, Carabidae) indicating a higher trophic position. These can ultimately serve as indicators of the health of plants and prey in the ecosystem, both in diversity and abundance. Although richness values recorded in our study did not significantly differ among the three urbanization levels, they hide changes in assemblages (Fig. 16.5b). Nevertheless, the proximity between points entails the occurrence of some species both in urban and non-urban sites.
The Chao-Sørensen similarity index varied among taxonomic groups and urban categories (Fig. 16.6), showing different tendencies among urbanization levels. Plants showed less similarity for all urban categories in forests (0.57 in average) than in peatlands (0.73 on average), with the lower values in urban patches. Greatest similarities in plants occurred in peri-urban forests and in non-urban peatlands, where we also found the lowest variation. In contrast, birds showed great similarity values (0.85 in average) in all urban categories, with slightly higher values in urban than in peri-urban than in non-urban forests. On the other hand, insects showed high similarity only in non-urban peatlands (0.90), with very low values in urban and peri-urban patches (0.26 in average), highlighting the greater sensitivity of this group compared with plants and birds.

Abundance-based Chao-Sorensen similarity index (mean ± SE) for forest plants and birds, and peatland plants and insects among the urban categories (urban, peri-urban, and non-urban)
3.4 Main Factors Affecting Biodiversity in Urban Forests and Peatlands in Ushuaia City
Despite some differences in richness, abundance, biodiversity indices, and composition for vascular plants, birds and insects, urban and peri-urban forests and peatlands still conserve several species and characteristics similar to non-urban ecosystems. However, the occurrence of new species, mainly plants, introduced into these urban ecosystems, modified the original communities into new assemblages, which offer another food supply to the fauna (i.e., birds and insects) (Marzluff and Rodewald 2008). The probability of greater dispersion of these newly introduced exotic plants in the non-urban ecosystem could depend on the mobility of dispersing agents, and on the connectivity and distance between urban forests and peatland patches to non-urban ecosystems. Previous studies affirm that urban green areas are ecological islands isolated from others in and around the city (Ceplová et al. 2017), but this depends on the size of cities and green areas. In addition, the relative importance of local- versus larger-scale variables in influencing urban biodiversity remains under discussion and will vary with the scale dependencies of different taxa (Goddard et al. 2010). The influence of local factors, especially patch size and quality, for urban forests and peatlands in Ushuaia city has not been analyzed, either for urban planning or conservation strategies.
The preservation of forests and peatlands as biodiversity islands within the urban matrix generates benefits for the environment and also for citizens living there, promoting ecological and socio-cultural functions (Rozzi et al. 2012) that would be completely lost if such natural islands were not preserved. Unfortunately, the link between the identity of inhabitants and care of regional habitats in Ushuaia is weak (Orzanco 1999; Kizman 2012) and much of the ancestral knowledge about natural ecosystems and their species has been lost. Moreover, the inadequate planning of public services (e.g., streets, sewage) to attend to the growth rate of the city, and bulky waste management are considered the main conflicts between urbanization and sustainability in Ushuaia city (Orzanco 1999). All these factors negatively influence the structure and functioning of biodiversity islands in the city (Fig. 16.7).

Conceptual framework to understand the main threats (top panels) affecting biodiversity (species richness and abundance) in urban forests and peatlands in Ushuaia city. Biodiversity islands serve as refuge for some native species that manage to survive despite structural changes and isolation from other natural environments, although they are also prone to invasion by introduced species (e.g., exotic plants) dispersed from homegardens, parks, etc
Because urban development is guided by human values, there is a need to inform people about the relevance of natural ecosystems supporting multiple services that increase human well-being and encourage caring for the environment. Ushuaia is a unique city in Argentina where peatlands are part of the urban space. However, their value as part of the natural heritage has not yet been recognized by society, and peatlands are considered obstacles to urban development rather than areas of natural, recreational and/or educational interest. However, peatlands were recently declared Environmental Reserves for their conservation and protection as Historical and Cultural Heritage by the Municipality of Ushuaia (Environment Office of Ushuaia 2018).
Although in Ushuaia urban forests and peatlands are connected to the natural undisturbed forests and peatlands occurring in the surrounding landscape, especially by birds and also by some insects (e.g., wasps) and some plants, this is generally not common knowledge for local people (Environment Office of Ushuaia 2018). This generates a twofold problem: on one hand, urban green areas do not receive the attention they rightfully deserve and are frequently modified irreversibly; and on the other, natural landscapes on the periphery are in danger from freely introduced exotics, either by negligence (weeds, insects) or intent (ornamental plants, pets). We recommend controlling and avoiding the installation of exotic plants, especially trees and bushes with fruits eaten by birds, in these urban green areas, to limit their spread and/or invasion to undisturbed environments.
4 Conclusions and Recommendations
Our results confirm that the urban forests and peatlands of Ushuaia are valuable reservoirs of biodiversity, even under anthropogenic influence. We recommend preserving existing natural green areas for their role as biodiversity islands in the urban landscape, due to their function as a refuge for native plants, birds and insects. In order to increase biodiversity and improve the connectivity of biodiversity islands within Ushuaia city we consider necessary the preservation and/or delimitation of new natural green areas (e.g., within new neighborhoods). Moreover, we highlight their value compared with parks and squares since these are non-natural, and mostly contain exotic vegetation (e.g., Salix sp., Populus sp., Pseudotsuga sp.).
According to our results, the urban peatlands would be more susceptible to deterioration and loss of species typical of these habitats than the urban forests. Therefore, it is important to implement urgent conservation plans to avoid the loss of these remaining natural habitats (indeed, some of the studied green areas have nowadays already been transformed and urbanized). Moving forward, the level and magnitude of the urbanization impact (i.e., distance to the urban center, transformation degree in the surroundings) on biodiversity deserve more analyses.
Urban islands must contribute to strengthening the link between society and nature, while still offering refuge, food and habitat for plant and animal species. These ecosystems also play an important role in maintaining the minimum environmental (e.g., carbon sinks, water purification) and socio-cultural functions (e.g., recreation, sense of belonging).
We also recommend including environmental education programs to facilitate the understanding of the importance of these biodiversity islands in the city, spread knowledge about native species (especially those inconspicuous or less charismatic), and promote the sustainable use of these environments. For local governments and planners, it is essential to plan the retention of natural green spaces when planning new neighborhoods, buildings, schools and parks. Moreover, conserving, designing, and managing urban green spaces requires balancing human perceptions, needs, and use with ecological requirements for preserving and enhancing biodiversity.
References
Benítez J, Pizarro JC, Blazina AP, Lencinas MV (2020) Response of bird communities to native forest urbanization in one of the southernmost city of the world. Urban Urban Gree 126887. https://doi.org/10.1016/j.ufug.2020.126887
Biondi LM, Bó MS, Favero M (2005) Dieta del chimango (Milvago chimango) durante el periodo reproductivo en el sudeste de la provincia de Buenos Aires, Argentina. Ornitol Neotrop 16:31–42
Bolger DT, Suarez AV, Crooks KR, Morrison SA, Case TJ (2000) Arthropods in urban habitat fragments in southern California: area, age and edge effects. Ecol Appl 10:1230–1248
Burghardt KT, Philips CR, Tallamy DW, Shropshire KJ (2010) Non-native plants reduce abundance, richness, and host specialization in Lepidoptera communities. Ecosphere 1(5):11. https://doi.org/10.1890/ES10-00032.1
Cárcamo JR, Contador T, Gañán M, Convey P, Kennedy J, Rozzi R (2019) Altitudinal gradients in Magellanic sub-Antarctic lagoons: the effect of elevation on freshwater macroinvertebrate diversity and distribution. Peerj 7:e7128–e7128. https://doi.org/10.7717/peerj.7128
Ceplová N, Kalusová V, Lsosová Z (2017) Effects of settlement size, urban heat island and habitat type on urban plant biodiversity. Landsc Urban Plan 159:15–22
Chao A, Chazdon RL, Colwell RK, Shen TJ (2005) A new statistical approach for assessing similarity of species composition with incidence and abundance data. Ecol Lett 8:148–159
Colwell RK (2009) EstimateS: statistical estimation of species richness and shared species from samples. http://purl.oclc.org/estimates
Contador T, Kennedy JH, Rozzi R, Villarroel JO (2015) Sharp altitudinal gradients in Magellanic sub-Antarctic streams: patterns along a fluvial system in the Cape Horn biosphere reserve. Polar Biol 38:1853–1866
Correa MN (1969–1998) Flora Patagónica. Colección Científica INTA Tomo 8. Partes II, III, IVb, V, VI y VII. Buenos Aires
Deferrari G, Camilión C, Martínez Pastur GM, Peri PL (2001) Changes in Nothofagus pumilio forest biodiversity during the forest management cycle. 2: birds. Biodivers Conserv 10:2093–2108
Díaz S (2012) Biología y conservación de la cachaña (Enicognathus ferrugineus) en Argentina. El Hornero 27:17–25
Environment Office of Ushuaia (2018) Plan de manejo de la Reserva Natural Urbana Bahía Encerrada, Municipalidad de Ushuaia, Tierra del Fuego, 98 p
Gerlach J, Samways M, Pryke J (2013) Terrestrial invertebrates as bioindicators: and overview of available taxonomic groups. J Insect Conserv 17:831–850
Goddard MA, Dougill AJ, Benton TG (2010) Scaling up from gardens: biodiversity conservation in urban environments. Trends Ecol Evol 25:90–98
Gómez-Baggethun E, Barton DN (2013) Classifying and valuing ecosystem services for urban planning. Ecol Econ 86:235–245
Hartley SE, Jones TH (2008) Insect herbivores, nutrient cycling and plant productivity. In: Weisser WW, Siemann E (eds) Insects and ecosystem function. Springer, Berlin, pp 27–52
INDEC (2010) Censo Nacional De Población, Hogares y Viviendas 2010. Documento Metodológico Preliminar. Available at: http://censo2010.indec.gov.ar/metodologia.asp. Accessed 1 Apr 2020
Kizman S (2012) Diagnóstico participativo: desarrollo de actividades sustentables. Parque Nacional Tierra del Fuego. APN, Tierra del Fuego. www.argentina.gob.ar.tierradelfuego
Lencinas MV, Martínez Pastur G, Medina M, Busso C (2005) Richness and density of birds in timber Nothofagus pumilio forests and their unproductive associated environments. Biodivers Conserv 14:2299–2320
Lencinas MV, Martínez Pastur G, Gallo E, Cellini JM (2014) Decreasing negative impacts of harvesting over insect communities using variable retention in southern Patagonian forests. J Insect Conserv 18:479–495
Lencinas MV, Sola FJ, Martínez Pastur G (2017) Variable retention effects on vascular plants and beetles along a regional gradient in Nothofagus pumilio forests. For Ecol Manag 406:251–265
Lencinas MV, Sola FJ, Cellini JM, Peri PL, Martínez Pastur GJ (2019) Land sharing in South Patagonia: conservation of above-ground beetle diversity in forests and non-forest ecosystems. Sci Total Environ 690:132–139
Lososová Z, Horsák M, Chytry M, Cejka M, Danihelka T, Fajmon J et al (2011) Diversity of Central European urban biota: effects of human-made habitat types on plants and land snails. J Biogeogr 38:1152–1163
MacArthur RH, Wilson EO (1967) The theory of island biogeography. Princeton University Press, Princeton
Maestre FT, Castillo-Monroy AP, Bowker MA, Ochoa-Hueso R (2012) Species richness effects on ecosystem multifunctionality depend on evenness, composition and spatial pattern. J Ecol 100:317–330
Marín M (2014) Distribución, fenología reproductiva, e historia natural del queltehue (Vanellus chilensis) en la zona central de Chile. Bol Mus Nac Hist Nat 63:119–126
Marvaldi AE, Lanteri AA (2005) Key to higher taxa of South American weevils based on adult characters (Coleoptera, Curculionoidea). Rev Chil Hist Nat 78:65–87
Marzluff J, Rodewald A (2008) Conserving biodiversity in urbanizing areas: nontraditional views from a bird’s perspective. Cities Environ 1(2):6–27
Marzluff JM, Bradley G, Shulenberger E, Endlicher W, Alberti M, Bradley G, Ryan C, ZumBrunnen C, Simon U (eds) (2008) Urban ecology. An international perspective on the interaction between humans and nature. Springer, Berlin
Matthies SA, Rüter S, Schaarschmidt F, Prasse R (2017) Determinants of species richness within and across taxonomic groups in urban green spaces. Urban Ecosyst 20:897–909
McGregor-Fors I, Morales-Pérez L, Schondube JE (2011) Does size really matter? Species–area relationships in human settlements. Divers Distrib 17:112–121
Millennium Ecosystem Assessment (2005) Ecosystems and human well-being. Island Press, Washington
Miller JR (2005) Biodiversity conservation and the extinction of experience. Trends Ecol Evol 20:430–434
Minnesota Department of Natural Resources (2013) A handbook for collecting vegetation plot data in Minnesota: The relevé method. 2nd ed. Minnesota Bio-logical Survey, Minnesota Natural Heritage and Nongame Research Program, and Ecological Land Classification Program. Biological Report 92. St. Paul, Minnesota Department of Natural Resources
Montagnini F, Levin B, Berg KE (2022) Introduction. Biodiversity Islands. Strategies for conservation in human-dominated environments. In: Montagnini F (ed) Biodiversity islands: strategies for conservation in human-dominated environments. Topics in Biodiversity and Conservation, Springer, Cham, pp 1–35
Moore DM (1983) Flora of Tierra del Fuego. Anthony Nelson
Moreno ML, Fernández MG, Molina SI, Valladares G (2013) The role of small woodland remnants on ground dwelling insect conservation in Chaco Serrano, Central Argentina. J Insect Sci 13:40. https://doi.org/10.1673/031.013.4001
Murphy D (1999) Challenges to biological diversity in urban areas. In: Wilson EO (ed) Biodiversity. National Academy Press, Washington, pp 71–76
Nilon CH, Aronson MFJ, Cilliers SS, Dobbs C, Frazee LJ, Goddard MA, O’Neill KM, Roberts D, Stander EK, Werner P, Winter M, Yocom KP (2017) Planning for the future of urban biodiversity: a global review of city-scale initiatives. Bioscience 67:332–342
Noreika N, Pajunen T, Kotze DJ (2015) Urban mires as hotspots of epigaeic arthropod diversity. Biodivers Conserv 24(12):2991–3007
Oliver I, Beattie AJ (1993) A possible method for the rapid assessment of biodiversity. Conserv Biol 7(3):562–568
Orzanco MG (1999) Problemas ambientales detectados por la población de Ushuaia (Tierra del Fuego, Argentina). Invest Geogr 40:85–98
Pauchard A, Ugarte E, Millán J (2000) A multiscale method for assessing vegetation baseline of environmental impact assessment (EIA) in protected areas of Chile. In: McCool SF, Cole DN, Borrie WT, O’Loughlin J (eds) Wilderness science in a time of change conference. USDA Forest Service, Rocky Mountain Research Station (United States), pp 111–116
Pielou EC (1975) Ecological diversity. Wiley, New York
Pinceloup N, Poulin M, Brice M-H, Pellerin S (2020) Vegetation changes in temperate ombrotrophic peatlands over a 35 years period. PLoS One 15(2):e0229146. https://doi.org/10.1371/journal.pone.0229146
Posadas P (2012) Species composition and geographic distribution of Fuegian Curculionidae (Coleoptera: Curculionoidea). Zootaxa 3303:1–36
Reich PB, Tilman D, Isbell F, Mueller K, Hobbie S, Flynn D (2012) Impacts of biodiversity loss escalate through time as redundancy fades. Science 336(6081):589–592
Remsen JV, Areta JI, Cadena CD, Claramunt S, Jaramillo A, Pacheco JF, Robbins MB, Stiles FG, Stotz DF, Zimmer KJ (2019) A classification of the bird species of South America. American Ornithologists’ Union. http://www.museum.lsu.edu/~Remsen/SACCBaseline.htm
Roig-Juñent S, Crisci JV, Posadas P, Lagos S (2002) Áreas de distribución y endemismo en zonas continentales. In: Costa C, Vanin SA, Lobo JA, Melic A (eds) Red Iberoamericana de Biogeografía y Entomología Sistemática (PrIBES). Sociedad Entomológica Aragonesa, Zaragoza, pp 247–266
Rozzi R, Jiménez JE (2014) Magellanic sub-antarctic ornithology: first decade of long-term bird studies at the Omora Ethnobotanical Park, Cape Horn biosphere reserve, Chile. University of North Texas Press, Dallas
Rozzi R, Massardo F, Silander J, Dollenz O, Connolly B, Anderson C, Turner N (2003) Native and exotic tree species in the squares of the Magellan Region. An Inst Patagon 31:27–42
Rozzi R, Armesto JJ, Gutiérrez JR, Massardo F, Likens GE, Anderson CB, Poole A, Moses KP, Hargrove E, Mansilla AO, Kennedy JH, Willson M, Jax K, Jones CG, Callicott JB, Arroyo MTK (2012) Integrating ecology and environmental ethics: earth stewardship in the southern end of the Americas. Bioscience 62(3):226–236
Sola FJ, Peri PL, Huertas L, Martínez Pastur GJ, Lencinas MV (2016) Above-ground arthropod community structure and influence of structural-retention management in southern Patagonian scrublands, Argentina. J Insect Conserv 20:929–944
Sola FJ, Soler R, Huertas A, Lencinas MV (2018) Urbanización y diversidad de invertebrados en turbales de Tierra del Fuego. In: Proc XXVIII Reunión Argentina de Ecología, 29 Oct - 2 Nov, Mar del Plata, Argentina
Tallamy DW, Ballard M, D’Amico VD (2010) Can alien plants support generalist insect herbivores? Biol Invasions 12:2285–2292
Thomas K, Geller L (2013) Urban forestry: toward an ecosystem services research agenda. The National Academies Press, Washington
Tuhkanen S (1992) The climate of Tierra del Fuego from vegetation geographical point of view and its ecoclimatic counterpartes elsewhere. Acta Bot Fenn 145:1–64
Vergara PM, Schlatter RP (2006) Aggregate retention in two Tierra del Fuego Nothofagus forests: short-term effects on bird abundance. For Ecol Manag 225:213–224
Zerbe S, Maurer U, Schmitz S, Sukopp H (2003) Biodiversity in Berlin and its potential for nature conservation. Landsc Urban Plan 62:139–148
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2022 The Author(s), under exclusive license to Springer Nature Switzerland AG
About this chapter

Cite this chapter
Soler, R., Benítez, J., Sola, F., Lencinas, M.V. (2022). Biodiversity Islands at the World’s Southernmost City: Plant, Bird and Insect Conservation in Urban Forests and Peatlands of Ushuaia, Argentina. In: Montagnini, F. (eds) Biodiversity Islands: Strategies for Conservation in Human-Dominated Environments. Topics in Biodiversity and Conservation, vol 20. Springer, Cham. https://doi.org/10.1007/978-3-030-92234-4_16
Download citation
DOI: https://doi.org/10.1007/978-3-030-92234-4_16
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-030-92233-7
Online ISBN: 978-3-030-92234-4
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)