
Abstract
We examined the effect of subchronic hypobaric hypoxia in rat heart. Adult male Sprague–Dawley rats were exposed at 25,000 ft for different time periods (2 and 5 days). Susceptibility of their hearts to oxidative stress as well as modulation in gene expression was evaluated. The results showed a crosstalk between reactive oxygen species (ROS) and nitric oxide (NO), initial response was accompanied by increase in ROS generation and development of oxidative stress as confirmed by increased lipid peroxidation, protein oxidation and accumulation of 2, 4-dinitrophenyl hydrazine and 4-hydroxy-2-nonenal adducts. At the same time, glutathione activity decreased; however, antioxidant enzymatic activities of superoxide dismutases, glutathione-S-transferase, and glutathione peroxidase rose in response to 5-days hypoxia. Interestingly, NO level increased till 5 days, however ROS decreased after 5 days; this observation suggests that ROS/NO balance plays an important role in cardioprotection. This observation is further supported by upregulation of antioxidant genes hemeoxygenase (HO-1) and metallothionein (MT). In addition, hypoxia also induces gradual upregulation of hypoxia-inducible transcription factor (HIF-1α), which in turn induces the expression of adaptive genes erythropoiesis, vascular endothelial growth factor, glucose transporter-1, nitric oxide synthase. Collectively, our data suggests a reciprocal regulation of ROS and NO and this effect is mediated by the increase in antioxidant proteins HO-1 and MT. Along with this HIF-1-mediated induction of various cardioprotective genes also plays an important role in acclimatization.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
A constant supply of oxygen is essential for cardiac viability and function. At high altitudes, the lower partial pressure of oxygen makes it more difficult to enter our cardio vascular systems. The heart ceases to contract with in seconds from the onset of oxygen deprivation (hypoxia) a pathological condition. As a common fact, reactive oxygen species (ROS) are generated in hypoxic heart [1, 2] and this may depress myocardial contraction through interaction with membrane lipids, proteins (by oxidation) as well as DNA and causes cell damage and death [3, 4]. There are several cellular mechanisms that counterbalance the production of ROS and inhibit the ROS-related damage of cells. Many enzymatic and nonenzymatic pathways, viz. glutathione (GSH), glutathione peroxidase (GPx), the superoxide dismutases (SODs), and vitamin (C and E) protects the cells against oxidative damage [5]. Hypoxia also influences nitric oxide (NO), which affects on the cellular respiration and ROS production, and could either contribute to or protect cells from hypoxia injury. NO does react and interact with ROS, and this crosstalk also resulted in significant effects on cardiac functions [6].
The hypoxia-inducible transcription factor (HIF-1) composed of α and β subunit plays a central role in controlling genes expression in response to oxygen levels [7, 8]. This heterodimeric transcription factor is composed of an oxygen-regulated hypoxia-inducible transcription factor (HIF-1α) subunit and a constitutively expressed HIF-1β subunit. The activation of HIF-1 is mainly dependent on the hypoxia-induced stabilization of the HIF-1α subunit, whereas the HIF-1β subunit is independent of the oxygen partial pressure. HIF-1α binds a hypoxia response element in HIF-responsive genes. These genes are mainly involved in angiogenesis {vascular endothelial growth factor (by VEGF), glucose metabolism (by Glut), vasodilators [hemeoxygenase (HO-1) and nitric oxide synthase (NOS)] and erythropoiesis}. Most of the genes of this class are involved in either the delivery of oxygen and nutrients to cells or controlling cellular utilization of these substrates [7, 9]. In addition many studies suggested that ROS provides a redox signal for hypoxic HIF-1 induction [10].
In the present study, we investigate the effect of sub chronic hypoxia on oxidative stress induced by hypobaric hypoxia on ROS generation, NO levels, antioxidant enzymes (SOD, GST, and GPx) activity, antioxidant genes (HO-1 and MT), as well as HIF-1-regulated genes. Our work is an elaborate study on the biochemical and molecular changes in the cardiac tissue that has not been studied so far on subchronic hypoxic exposure. It proposes a novel and potential phenomenon of acclimatization at high altitude.
Methods
Animals and Reagents
Male Sprague–Dawley rats (175 ± 25 g) were used for all experiments. Animals were maintained in a 12:12-h light–dark cycle at constant temperature (24 ± 2 °C) in our in-house animal house facility with food (Lipton India Ltd.) and water ad libitum. The experimental protocol was carried out in accordance with the guidelines of the Ethics Committee of this Institute and was in compliance with the European Communities Council directive 1986.
Hypoxic Exposure
The animals were randomly subdivided into three experimental group of 10 animals each: (1) control, (2) 2 days, and (3) 5 days and exposed to a simulated altitude of 25,000 ft (subchronic) (as rats have metabolic rate approximately six times more than humans) in a decompression chamber where temperature and humidity were maintained at 28 °C and 55–60 %, respectively. The rate of ascent was at the rate of 300 m/min. The rats were taken out of hypoxic chamber once after every 24-h exposure for 15 min for replenishing food and water.
Biochemical Analysis
After hypoxic exposure, the rats were sacrificed and heart was dissected out after perfusion with 20 ml sterile PBS (pH 7.4) to remove the blood and stored at −80 °C for further analysis. Later, the heart was homogenized in cold 0.154 M KCl fortified with protease inhibitors cocktail, PMSF and DTT (Sigma, USA), to obtain 10 % homogenate (w/v). One part of homogenate was then centrifuged at 2,900×g for 10 min at 4 °C. The pellet-containing tissue/cell debris were discarded and supernatant was used to determine ROS, GSH, lipid peroxidation, and protein oxidation. The other part of homogenate was centrifuged at 14,000×g for 20 min at 4 °C and supernatant was used for estimation of antioxidant enzyme activity like GST, GPx, and SOD levels. The protein content in the homogenate was determined by Lowry’s method [11].
Reactive Oxygen Species
ROS generation was measured in supernatant with 2,7-dichlorofluorescein-diacetate (DCFH-DA) by the method of LeBel and Bondy [12], as modified by Kim et al. [13]. In brief, 50 μl of homogenate was added to a cuvette containing 2,938 μl of 0.1 M PBS (pH 7.4) and 12 μl of 1.25 mM, 2,7-dichlorofluorescein-diacetate (DCF-DA) prepared in methanol. The assay mixture was incubated for 15 min at 37 °C and 2,7-dichlorofluorescein formation was determined fluorimetrically using spectrofluorimeter (Varion, USA). The fluorescent intensity represent to the amount of ROS formed in the individual sample. The results were expressed as relative fluorescent unit (rfu).
Lipid Peroxidation
Lipid peroxidation was measured by estimating malondialdehyde (MDA) formed by thiobarbituric acid reaction and measuring the absorbance at 531 nm [14]. 1,1,3,3-Tetraethoxy propane was used as standard, and the levels of lipid peroxides were expressed as nanomole per milligram protein (nmol MDA/mg protein). Also, 4-hydroxy-2-nonenal (HNE) protein adducts were detected by western blotting using rabbit polyclonal HNE antibody (Alexis, Farmingdale, NY, USA) and anti-rabbit-IgG-HRP conjugate as secondary antibody(Santa Cruz, CA, USA).
Protein Oxidation
Protein oxidation was measured by determining the carbonyl groups after derivatization with 2,4-dinitrophenyl hydrazine (DNPH) [15]. Briefly, equal volume of supernatant 10 mM DNPH/2 M HCl was incubated for 60 min at 50 °C. Protein was then precipitated with 20 % TCA, and unreacted DNPH was removed by centrifugation at 1,400×g for 10 min. The precipitates were washed with cold ethanol–ethyl acetate (1:1) mixture three times, and finally precipitates were dissolved in 1 N NaOH. The absorbance was measured at 450 nm, and the carbonyl content was expressed as nanomole per milligram of protein. Protein associated carbonyl groups were also determined by western blotting using rabbit polyclonal DNPH antibody (Alexis) as described in “Lipid Peroxidation” section.
Enzymatic and Nonenzymatic Antioxidants
Reduced GSH levels were measured fluorimetrically by the method of Hissin and Hilf [16]. Activities of GPx (EC 1.11.19) and total SOD (EC 1.15.1.1) were determined using commercial kits (RANDOX, UK) as per manufacturer’s instructions. GST (EC 2.5.1.1.8) was determined using protocol described by Habig et al. [17]. Briefly, 100 μl sample was mixed with 2.790 ml 0.1 M potassium phosphate buffer (pH 6.5) and 100 μl of 100 mM GSH. Reaction was initiated by adding 1-chloro 2,4-dinitrobenzene (40 mg/ml in ethanol) and optical density was recorded at 340 nm.
NO Levels
Nitrite, a biological metabolite of NO was measured in heart tissue homogenate using Griess reagent [18]. Briefly, 100-mg heart tissue was homogenized in 1 ml PBS and centrifuged at 2,500×g for 10 min. Supernatant was mixed with equal volume (100 μl) of Griess reagent (Sigma) and incubated for 30 min in dark. A chromophore azo-derivative thus formed was measured at 540 nm in a microplate reader (Molecular Devices, CA, USA). Standard curve was prepared by using different dilutions of sodium nitrite with each assay.
Histology
Hearts from all the rats from each group were excised post exposure. The tissues were fixed in 10 % buffered formalin and after proper fixation, were dehydrated in graded series of alcohol, cleared in xylene, and embedded in paraffin wax. Multiple sections from each block were prepared at 5 μm, and stained with haematoxylin and eosin.
Protein Expression Studies
Preparation of Nuclear and Cytosolic Extract
For nuclear and cytosolic fractionation, frozen heart tissues were homogenized in an ice-cold buffer A [0.5 M sucrose, 10 mM HEPES, 10 mM KCl, 1.5 mM MgCl2, 10 % glycerol, 1 mM EDTA, 1 mM DTT, 1 mM PMSF fortified with protease inhibitors (Sigma)]. Homogenates were kept on ice for 15 min, and 0.6 % nonidet P-40 was added, and then centrifuged for 10 min at 2,000×g at 4 °C. The supernatant with cytosolic fraction was collected, stored and pellet was dissolved in cold buffer B (20 mM HEPES, 1.5 mM MgCl2, 0.3 mM NaCl, 0.2 mM EDTA, 20 % glycerol, 0.5 mM DTT, 0.5 mM PMSF, and cocktail of protease inhibitors) for nuclear fraction. It was incubated for half an hour on ice followed by centrifugation at 20,000×g at 4 °C for 15 min. The supernatant containing nuclear fraction was aliquot and stored at −80 °C for further analysis.
Western Blotting
Nuclear and cytosolic cell extracts as prepared above were made to detect various proteins present in different cellular fraction. Transcription factor HIF-1α was analyzed using nuclear extract and VEGF, erythropoietin (EPO), glucose transporter-1 (Glut-1), HO-1, MT-1, iNOS, nNOS and eNOS (Santa Cruz) were quantified in cytosolic extract by western blot analysis. Protein (40 μg) was separated by SDS-polyacrylamide gel electrophoresis and transferred onto a nitrocellulose membrane (HYBOND Millipore, USA). The membranes were blocked with 3 % BSA in PBS containing 0.1 % Tween 20 (Sigma, USA), washed and probed with respective mouse/rabbit polyclonal antibodies (Santa Cruz). The membranes were then incubated with anti-mouse/rabbit-IgG HRP conjugate (Santa Cruz), respectively. The membrane was washed and incubated with chemiluminiscent substrate (Sigma) and the bands were developed using X-ray films (Kodak, UK). Quantification was performed by scanning desitometry using Labworks software (UVP Bio-imaging systems, UK).
Statistical Analysis
All the experiments were performed at least thrice and the data are presented as mean ± SD. Statistical analysis was carried out by analysis of variance followed by post hoc Newman Keul’s test using SPSS version 14.0 statistical software (SPSS Inc., Chicago, IL, USA). P < 0.05 was considered as significant.
Results
Reactive Oxygen Species Generation
To assess the effect of hypobaric hypoxia-induced oxidative stress in rat heart; ROS levels were determined using a fluorescent probe DCF-DA. Exposure for 2 days caused a significant increase in ROS generation. A twofold increase in ROS generation was observed in the animal kept in hypobaric hypoxia condition relative to control animal (201.41 ± 1.25 vs. 115.76 ± 2.1 rfu/min/mg protein, p < 0.01). Surprisingly, ROS generation decreased significantly (163.79 ± 2.5 rfu/min/mg protein, p < 0.01) after 5 days of hypoxic exposure relative to 2-days exposure, indicating reduced oxidative stress (Fig. 1).

ROS as an indicator of oxidative stress measured using dichlorofluorescein (DCF) as a probe in fresh heart homogenates and values were expressed as relative fluorescence unit (rfu) is depicted in the figure above. ROS generation increased significantly after 2 days of hypoxic exposure. It deceased after 5 days of continuous hypobaric hypoxia. Data expressed as mean ± SD of three independent experiments. Data analyzed by analysis of variance followed by post hoc Newman Keul’s test. a P < 0.05, compared to control; b P < 0.05, compared to 2-days hypoxia
Lipid Peroxidation
Lipid peroxidation products include both saturated aldehydes, such as malondialdehyde and unsaturated aldehydes, such as 4-hydroxy-nonenal (HNE). Exposure of animals to 2-days hypobaric hypoxia resulted in marked increase in mean tissue MDA levels (10.71 ± 0.099 vs. 2.59 ± 0.08 nmol/mg protein, p < 0.01) relative to control rats. Further, only marginal increased was observed in mean tissue MDA level after 5 days of hypoxic exposure (12.36 ± 0.87 nmol/mg protein; Fig. 2a).

Lipid peroxidation in heart homogenate observed after 2 and 5 days of continuous hypoxia shown by a spectrophotometric measurement of thiobarbituric acid reactive substances (TBARS) and by b immunoblot for formation of HNE adducts and relative optical density (ROD) of control and exposed rats for 2 and 5 days continuous hypoxia. Exposure led to a significant increase in both TBARS production and HNE adduct formation. Data expressed as mean ± SD of three independent experiments. Data analyzed by analysis of variance followed by post hoc Newman Keul’s test. a P < 0.05, compared to control
The unsaturated aldehydes have a well-documented toxicity to organic molecules and their detection may be of greater biological importance than that of malondialdehyde [19]. Therefore, we determined production of HNE adduct by immunoblotting. A significant increase in 4-HNE adduct levels was detected in 2 and 5 days (p < 0.05) relative to control but no significant difference was observed between 2- and 5-days exposure (Fig. 2b).
Protein Oxidation
Reactive protein carbonyl derivatives have often been measured as an index of tissue damage by oxidative stress [20]. Hence, the effect of hypoxia on protein oxidation was measured by determining protein carbonyl content in heart homogenate after derivatization with DNPH. A marked increase was observed in protein oxidation in rats after 2 days (175.42 ± 1.9 vs. 129.32 ± 2.89 nmol/mg protein, p < 0.01) of hypoxic exposure compared to control. However, only little increase was observed further after 5 days (187.75 ± 2.07 nmol/mg protein) of hypoxic exposure compared to 2-days hypoxic group (Fig. 3a). We also determined the protein carbonyl group in heart by immunoblotting using anti-DNP antibodies. Significant increase in oxidized proteins was detected after 2 and 5 days (p < 0.05) of exposure to hypoxia as compared to control animals (Fig. 3b). However, no significant change was observed in DNP expression between 2- and 5-days exposed rats.

The level of protein oxidation in rat heart as determined by: a spectrophotometric measurement of the carbonyl groups after derivatisation of proteins with dinitrophenylhydrazine (DNPH); b immunoblot for formation of carbonyl group on oxidized proteins and its relative optical density (ROD). Protein oxidation increased significantly when exposed to simulated altitude of 7,622 m for 2 and 5 days. Data expressed as mean ± SD of three independent experiments. Data analyzed by analysis of variance followed by post hoc Newman Keul’s test. a P < 0.05, compared to control
Glutathione Status
Glutathione, the major non-protein thiol in living cells, plays an important role as an antioxidant. GSH acts as a free-radical scavenger and helps in regenerating other antioxidants [21]. The changes in GSH and GSSG levels in heart tissue homogenate are depicted in Table 1. Exposure to hypoxia for 2 days caused a significant decrease in mean tissue GSH levels and increase in GSSG levels relative to control animals (p < 0.05). Also the GSH/GSSG ratio was reduced significantly after 2 days of hypoxia exposure. However GSH, GSSG levels, and GSH/GSSG ratio changed marginally after 5 days of hypobaric hypoxia exposure relative to 2-days exposure.
Antioxidant Enzyme Activity
SOD, GST, and GPx are regarded as the first line of the antioxidant defense system against ROS generated during hypoxia. As shown in Table 1. SOD activity significantly increased after 2 days of hypoxia (2.99 ± 0.01 vs. 1.01 ± 0.01 U/mg protein, p < 0.01) compared to control group. It increased further after 5 days of exposure (4.38 ± 0.01, p < 0.01). There was little change in GPx and GST, activity after 2 days of hypoxic exposure; however, significant increase was observed after 5 days (p < 0.01) of hypoxia exposure compared to control animals (Table. 1).
Nitrite Production
Nitrite as an indicator of NO, which is one of the most important regulatory factors of cardiovascular system [22] was measured. Exposure of rats to hypobaric hypoxia resulted in an appreciable increase in NO production after 2 days (83.03 ± 0.91 vs. 57.89 ± 1.1 μmol/mg protein, p < 0.05) relative to control, the nitrite level increased further after 5 days (102.38 ± 2.07 μmol/mg protein, p < 0.01.) of exposure (Fig. 4).

Nitrite as an indicator of nitric oxide as measured using Griess reagent in fresh heart homogenates is depicted in the figure above. There was a significant increase in nitrite after 2 and 5 days of continuous hypobaric hypoxia. Data expressed as mean ± SD of three independent experiments. Data analyzed by analysis of variance followed by post hoc Newman Keul’s test. a P < 0.05, compared to control; b P < 0.05, compared to 2-days hypoxia
Histology
To further validate the effects of hypobaric hypoxia on heart histopathology was conducted. Figure 5 presents histoarchitecture of heart in control and hypobaric hypoxia-exposed rats. Exposure to hypobaric hypoxia demonstrated an increase in myocyte size from right ventricle after day 2 of exposure which restored towards normal after day 5 of exposure.

Histology of male rats heart tissue after 2 and 5 days of hypobaric hypoxia exposure. a Histoarchitecture of heart in control male rats. Exposure to hypobaric hypoxia demonstrated an increase in myocyte size from right ventricle after 2 days of exposure (b) which restored towards normal after 5 days of exposure (c)
Expression of Hemeoxygenase and Metallothionein
One of the less well-known but critical defenders of cellular homeostasis is the microsomal enzyme HO-1. It is induced by exposure to various forms of oxidative stress, including heat shock, hypoxia, hyperoxia, UV light, hydrogen peroxide [23]. The levels of HO-1 protein expression was elevated significantly after 2 days (p < 0.05) and increased further up to 5 days (p < 0.01) of hypoxic exposure (Fig. 6a).

Induction of hemeoxygenase-1 (HO-1) and metallothionein-1 (MT-1) in rat heart, a representative immunoblot of HO-1 with its relative optical densities (ROD), b immunoblot of MT-1 with its ROD normalized with actin. There was a significant increase in HO-1 and MT-1 levels after 5 days of continuous hypobaric hypoxia. Data expressed as mean ± SD of three independent experiments. Data analyzed by analysis of variance followed by post hoc Newman Keul’s test. a P < 0.05, compared to control; b P < 0.05, compared to 2-days hypoxia
Metallothionein (MTs) are intracellular, low molecular weight, cystein-rich proteins that provide protection against heavy metals and oxidant damage. We determined the levels of MT-1 in rat heart by immunoblotting. Upregulation of MT-1 levels was observed after 2 and 5 days of exposure as compared to control group (p < 0.05; Fig. 6b).
Effect of Hypoxia on HIF-1α and Regulated Genes
The hypoxia-inducible factor (HIF-1) is a key regulator of oxygen homeostasis. Immunoblotting showed a virtually undetectable level of HIF-1α protein in control animals however, exposure to hypoxia for 2 days (p < 0.05) and 5 days (p < 0.05) resulted in a significant increase in HIF-1α protein levels (Fig. 7a).

Immunoblot analysis of a HIF-1α- and HIF-1-regulated genes, viz. b EPO, c Glut-1, d VEGF in rat heart along with their respective relative optical densities (ROD). ROD was normalized with actin. Data represent the mean ± SD of three independent experiments. a P < 0.05, compared to control; b P < 0.05 compared to 2-days hypoxia
The continuous formation of new blood cell is regulated by glycoprotein hormone EPO. A substantial increase was observed in EPO level in the heart of rats exposed for 2 days (p < 0.05) it remained elevated after 5 days (p < 0.05) of continuous hypoxia relative to control (Fig. 7b).
During hypoxia, oxidative metabolism decreases greatly, while anaerobic glycolysis increases to compensate for ATP generation. The effect of hypoxia on Glut-1 expression, a glucose transporter was therefore, determined in heart of rats. A significant change in Glut-1 expression was observed after 2 days (p < 0.05) and it increased marginally after 5 days (p < 0.05) of hypoxic exposure as compared to control rats (Fig. 7c).
Hypoxia has been shown to be very important stimulus for the new vessel formation seen in coronary artery disease and tumor angiogenesis [24]. A considerable increase in expression of VEGF a potent angiogenic factor was observed after 2 days, it increased marginally after 5 days (p < 0.05) of hypoxia compared to control group (Fig. 7d).
Expression of Nitric Oxide Synthase
NO synthase (NOS) catalyzes the oxygen and NADPH-dependent oxidation of l-arginine, leading to the production of nitric oxide. We observed an apparent increase in expression of eNOS after 2 and 5 days (p < 0.05) of hypoxic exposure as compared to control rats (Fig. 8a). Increased expression was also observed in iNOS after 2 and 5 days (Fig. 8b). Neuronal NOS (nNOS) was also examined but the result was not significant (data not shown).

Effect of continuous hypoxia on the source of nitric oxide as observed by immunoblot analysis of nitric oxide synthase (NOS) isoforms. a Representative western blot of endothelial NOS (e-NOS) and b representative western blot of inducible NOS (i-NOS) along with their respective relative optical density (ROD). ROD was normalization with actin. Data represent the mean ± SD of three independent experiments. a P < 0.05, compared to control; b P < 0.05 compared to 2-days hypoxia
Discussion
To our knowledge, this study represents the first attempt to address how heart reacts to subchronic hypobaric hypoxia stress using in vivo model. Continuous exposure results in increased endogenous antioxidant defense system and HIF-mediated induction of various adaptive responses. The salient finding of the present study are; (1) exposure for 2 days caused a marked increase in ROS generation, lipid peroxidation and protein oxidation; (2) the NO level markedly increase after 2 and 5 days; (3) enzymatic and nonenzymatic antioxidants levels increases in heart tissue after 5 days; (4) exposure for 5 days attenuates ROS generation; (5) the expression of HIF-1α and its regulated gene such as HO-1, MT-1, EPO, VEGF, Glut-1, eNOS and iNOS significantly increased.
High altitude is characterized by hypobaric hypoxia, which is considered as an acute physiological stress leading to oxidative stress, consequently causing potential damage to protein, lipid and DNA [25]. Oxidative stress is the result of excessive production of ROS species and or depletion of intracellular antioxidant defense, leading to imbalance in the redox status of the cell. A significant number of in vitro and animal studies have demonstrated ROS activation in the cardiovascular system in response to various stressors and in the heart failure [3].
Therefore, present study is an attempt to investigate the possible biochemical and molecular cellular mechanism(s) by which an animal acclimatized to hypoxic condition. Our data clearly suggest that initial or short-duration exposure (2 days) of hypobaric hypoxia resulted in significant ROS generation in heart tissue but interestingly at similar exposure condition for long duration we did not observed any further increase in ROS generation. These results provide a strong reason to investigate whether a decrease in ROS level after five days exposure is due to of its saturation or is a result of acclimatization of the animal in hypoxic condition.
ROS generation after hypobaric hypoxia exposure caused further damages to membrane lipid composition as observed by increase in lipid peroxidation and protein oxidation. Our results are in agreement with many reports which showed that lipid peroxidation, is a well-characterized effect of ROS that results in damage to the cell membrane as well as the membranes of cellular organelles [26]. Polyunsaturated lipids are susceptible to free-radical attack that results in peroxidation of membrane and generation of various aldehyde, alkenals, and hydroxyalkenals such as MDA and HNE [19]. HNE, a major product of lipid peroxidation, is thought to be the most reactive of these compounds and is, therefore, an important mediator of free-radical damage [19]. Thus, hypoxia-induced free-radical events are likely to be mediated, in part, by HNE-derived modifications. In the present study too we observed an increase in the MDA and HNE adducts after 2 and 5 days of hypobaric hypoxia-exposed rats. This view of increased MDA levels and HNE adduct formation is supported by our previous studies in brain and lung tissue [27, 28]. Also, Nakanishi et al. [29] showed increased MDA levels in serum, heart, lung, and kidney of hypoxic rats.
Modification of proteins by ROS can cause inactivation of critical enzymes and can induce denaturation that renders proteins nonfunctional [30]. HNE generated during lipid peroxidation also reacts with protein at the sulfhydryl group of cystein, the imidazole nitrogen(s) of histidine, and/or the έ-amine of lysine [31, 32], resulting in enzyme inactivation [33]. It was therefore of interest to determine the extent of oxidative modification of protein in heart of hypoxic rats. A significant increase in protein oxidation was observed in rat’s heart after 2 days of hypobaric hypoxia further only a little increase was observed in protein oxidation after 5 days.
To correlate with the observed fall in oxidative stress after 5 days, we monitored the antioxidants such as GSH, GPx, GST, and SOD which are responsible to scavenge ROS. GSH directly scavenges ROS and protects cell or tissue from free-radical damage. GSH is pivotal in various protective systems such as GPx and GST [34]. In our study, a decrease in GSH level with concomitant increase in GSSG levels was observed in hearts of rats exposed to hypoxia was in accordance with previous reports by Ilavazaghan et al. [35]. However antioxidant enzyme activity of GPx and GST were increased after 2 and 5 days of hypoxic exposure. This result may be a consequence of increased GSH depletion due to increased oxidative stress in hypoxic group. Additionally, GSH may be depleted by organic peroxidases through GPx activity, and certain GST can also catalyze such reaction. GPx decomposes peroxides using the GSH as its co-substrate [36, 37]. Moreover, a part of the system is GST, which catalyzes the conjugation of GSH to nucleophilic xenobiotics or cellular components damaged by oxy-radical attack to result in their detoxification reaction [36, 38]. SOD, an intracellular antioxidant enzymes, present in all aerobic cells has antitoxic effect against superoxide anion. In the present study we have shown that hypoxia is associated with increased SOD concentrations in rat’s heart exposed to 2 and 5 days of continuous hypoxia, which would help to protect the heart from oxidative damage. The accumulation of damaged products is probably partially prevented by the elevated activities of antioxidant enzymes (GPx and SOD).
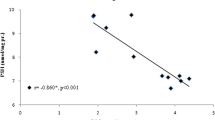
To fully elucidate the acclimatization process in the animal, we next study the NO, one of the main candidates to accord the cardioprotection effect of acclimatization at high altitude [39]. Our data indicate that NO level increase consistently till 5-days exposure in the animal. As one of the main finding of present study, here we report a reciprocal regulatory relationship between of ROS and NO during hypoxic exposure which not only show signaling complexity of these two functionally different molecules but also provide strong evidence of acclimatization at high altitude. Initial response is accompanied by reversible increase in the generation of ROS and is blocked by enzymatic and nonenzymatic antioxidants and interventions that increases the tissue levels of NO, suggesting that continuous hypoxia leads to an alteration of ROS/NO balance which is eventually restored during the acclimatization process [40]. Therefore significant increase in nitric oxide after 2 and 5 days of hypobaric hypoxia might lead to cardioprotection.
To support our biochemical results, we also see the levels of antioxidant proteins like hemeoxygenase and metallothionein. HO involves in the formation of bilirubin an efficient scavenger of ROS and has been reported to attenuate free-radical-mediated damage to serum albumin [41]. Furthermore, increased bilirubin levels by HO in various tissues have been regarded as an important cellular defense mechanism against oxidative injury [42]. Metallothionein are also very efficient hydroxyl radical scavenger and many in vivo studies indicate that MT indeed provide protection against oxidative injury in multiple organ system, strongly implicating its antioxidant function. We observed increase in HO-1 and MT-1 activity after 2 days of hypoxia exposure which further increases after 5-days exposure, showing a defensive response against hypobaric hypoxia-induced oxidative stress.
Under hypoxic condition acclimatization response at molecular level is characterized by upregulation of HIF-1α expression in rat heart. This increase in nuclear HIF-1α concentration leads to elevated transcriptional activation of the HIF-1 target genes [43] responsible for erythropoesis, angiogenesis and glucose transport. In our study, we found increased expression of EPO, Glut-1, and VEGF in heart tissue of rats exposed to continuous hypoxia for 2 and 5 days. Increased EPO and Glut-1 level with hypoxia in heart leads to higher performance, a cause by a higher O2 transport capacity to heart muscles. Increased angiogenesis is advantageous since the given amount of blood will pass the muscle of heart at slower flow rate, and therefore increase the time for gas and metabolite exchange across the tissue. Thus, an increased expression of these genes could be a logical adaptive mechanism to chronic exposure. VEGF exerts a long-lasting stimulatory effect on endothelial nitric oxide generation by increasing constitutive nitric oxide synthase expression [44]. Nitric oxide has been proposed to play a role as a mediator of increased tolerance to ischemia in the chronically hypoxic hearts [45]. Increased VEGF expression might therefore contribute to the protective effect of chronic hypoxia on the cardiac muscle by these two mechanisms.
Based on the effects of NO synthase inhibitors or NO donors, it has been proposed that increased generation of NO plays a positive role in the protective mechanism induced by chronic hypoxia in neonatal rabbit [45] and rat heart [46]. It still remains elusive whether NO produced by the hypoxic myocardium originates from constitutive eNOS [45] or iNOS [47, 48]. Our study also reports that the source of increased NO production may be both iNOS and eNOS as shown by their upregulation after 2 and 5 days of hypobaric hypoxia. However, it should be perceived that there is an optimal concentration of NO for protection: too little or too much may be detrimental. The role of NO in myocardial I/R injury and in adaptive protective responses of chronically hypoxic heart is extremely complex [49, 50] and further studies may be required to understand the complex mechanism (Fig. 9).

Putative corelation among various factors and mechanism of induction of carioprotection by hypoxia
Conclusion
In conclusion, the present study showed that excessive formation of ROS is a primary cause of tissue injury which decreased with the increase in production of NO, antioxidant enzymes, antioxidant genes, and activation of adaptive genes. Therefore, it is an important mechanism by which sustained hypoxia triggers long-term adaptive responses, which may in turn lead to cardioprotection. The identification of the biochemical and molecular basis of the phenomenon would provide a conceptual framework for developing novel therapeutic strategies aimed at mimicking the cardioprotective effect of preconditioning with pharmacological agents (hypoxia mimetic) and genetic approaches (transfer of cardioprotective genes) that can maintain the heart in a sustained or defensive (preconditioned) state for coping with stress.
References
Damerau, W., Ibel, J., Thurich, T., Assadnazari, H., & Zimmer, G. (1993). Basic Research in Cardiology, 88, 141–149.
Vanden Hoek, T. L., Li, C., Shao, Z., Schumaker, P. T., & Becker, L. B. (1997). Journal of Molecular and Cellular Cardiology, 29, 2571–2583.
Suematsu, N., Tsutsui, H., Wen, J., Kang, D., Ikeuchi, M., Ide, T., et al. (2003). Circulation, 107, 1418–1423.
Hensley, K., Robinson, K. A., Gabbita, S. P., Salsman, S., & Floyd, R. A. (2000). Free Radical Biology & Medicine, 28, 1456–1462.
Halliwell, B., & Gutteridge, J. M. C. (1998). Free radical in biology and medicine (3rd ed.). Oxford: Oxford University Press.
Arteel, G. E., Briviba, K., & Sies, H. (1999). FEBS Letter, 445, 226–230.
Giordano, F. J., & Johnson, R. S. (2001). Current Opinion in Genetics and Development, 11, 35–40.
Carmeliet, P., Dor, Y., Herbert, J. M., Fukumura, D., Brusselmans, K., Dewerchin, M., et al. (1998). Nature, 394, 485–490.
Semenza, G. L. (2001). Pediatric Research, 49, 614–617.
Kietzmann, T., & Gorlach, A. (2005). Seminars in Cell & Developmental Biology, 16, 474–486.
Lowry, O. H., Rosebrough, N. J., Faro, A. L., & Randell, R. J. (1951). The Journal of Biological Chemistry, 193, 265–275.
LeBel, C. P., & Bondy, S. C. (1990). Neurochemistry International, 17, 435–440.
Kim, J. D., McCarter, R. J. M., & Yu, B. P. (1996). Aging Clinical And Experimental Research, 8, 123–129.
Ohkawa, H., Ohishi, N., & Yagi, K. (1979). Analytical Biochemistry, 95, 351–358.
Levine, R. L., Garland, D., & Oliver, C. N. (1990). Methods in Enzymology, 186, 464–478.
Hissin, P. J., & Hiff, R. A. (1976). Analytical Biochemistry, 74, 214–226.
Habig, W. H., Pabst, M. J., & Jocoby, B. W. (1974). The Journal of Biological Chemistry, 249, 7130–7139.
Barrias, J. A. G., Escalante, B., Valdes, J., Bertha, A., Chavez, L., & Fong, D. M. (2002). Brain Research, 945, 114–122.
Esterbauer, H., Schaur, R. J., & Zollner, H. (1991). Free Radical Biology & Medicine, 1, 181–128.
Radak, Z., & Goto, S. (2000). In Z. Radak (Ed.), Free radicals in exercise and aging (pp. 177–209). Champaign: Human Kinetics.
Meister, A., & Anderson, M. E. (1983). Annual Review of Biochemistry, 52, 711–760.
Moncada, S., & Higgs, A. (1993). The New England Journal of Medicine, 3(29), 2002–2012.
Lee, P. J., Jiang, B. H., Chin, B. Y., Iyer, N. B., Alam, J., Semenza, G. L., et al. (1997). The Journal of Biological Chemistry, 272, 5375–5381.
Shweiki, D., Itin, A., Soffer, D., & Keshet, E. (1992). Nature, 359, 843–845.
Baily, D. M., & Davies, B. (2001). High Altitude Medicine & Biology, 2, 21–29.
Rathore, N., John, S., Kale, M., & Bhatnagar, D. (1998). Pharmacological Research, 38, 297–303.
Shrivastava, K., Shukla, D., Bansal, A., Mustoori, S., Banerjee, P. K., & Ilavazhagan, G. (2008). Neurochemistry International, 103, 54–59.
Shukla, D., Saxena, S., Jayamurthy, P., Mustoori, S., Mrinalini, S., Jain, S. K., et al. (2009). High Altitude Medicine & Biology, 10, 57–69.
Nakanishi, K., Tajima, F., Nakamura, A., Yagura, S., Ookawara, T., Yamashita, H., et al. (1995). The Journal of Physiology, 489, 869–876.
Stadtman, E. R., & Levine, R. L. (2003). Amino Acids, 25, 207–218.
Szweda, L. I., Uchida, K., Tsai, L., & Stadtman, E. R. (1993). The Journal of Biological Chemistry, 268(5), 3342–3347.
Tsai, L., & Sokoloski, E. A. (1995). Free Radical Biology & Medicine, 19, 39–44.
Sabri, M. N., DiSciascio, G., Coeley, M. J., Alpert, D., & Vetroveck, G. W. (1991). American Heart Journal, 121, 876–880.
Bast, A., Haenen, G. R. M. M., & Doelman, C. J. A. (1991). American Journal Of Medicine, 91(3C), 2S–13S.
Ilavazhagan, G., Bansal, A., Prasad, D., Thomas, P., Sharma, S. K., Kain, A. K., et al. (2001). Aviation, Space, and Environmental Medicine, 72, 899–903.
Halliwell, B. (1994). Nutrition Reviews, 1, 253–256.
Saez, G. T., Bannister, W. H., & Banister, J. V. (1990). In J. Vina (Ed.), Glutathione metabolism and physiological functions (pp. 237–254). Boca Raton: CRC.
Singal, S. S., Saxema, M., Ahmad, H., Awasthi, S., Haque, A. K., & Awasthi, Y. C. (1992). Archives of Biochemistry and Biophysics, 299, 232–241.
Kolar, F., & Ostadal, F. (2004). Physiological Research, 53(1), S3–S13.
Gonzalez, N. C., & Wood, J. G. (2001). Advanced in Experimental Medicine and Biology, 502, 39–60.
Neuzil, J., & Stocker, R. (1993). FEBS Letters, 331, 281–284.
Llesuy, S. F., & Tomaro, M. L. (1994). Biochimica et Biophysica Acta, 223, 9–14.
Hopfl, G., Ogunshola, O., & Gassmann, M. (2003). Advances in Experimental Medicine and Biology, 543, 89–115.
Vander Z, R., Murohara, T., Luo, Z., Zollmann, F., Passeri, J., Lekutat, C., et al. (1997). Circulation, 95, 1030–1037.
Baker, J. E., Holmen, P., Kalyanaraman, B., Griffith, O. W., & Pritchard, K. A. (1999). Annals of the New York Academy of Sciences, 874, 236–253.
Ostadalova, I., Ostadal, B., Jarkovska, D., & Kolar, F. (2002). Pediatric Research, 52, 561–567.
Ding, H. L., Zhu, H. F., Dong, J. W., Zhu, W. Z., Yang, W. W., Yang, H. T., et al. (2005). Acta Pharmacologica Sinica, 26, 315–322.
Ferreiro, C. R., Chagas, A. C., Carvalho, M. H., Dantas, A. P., Jatene, M. B., Bento de Souza, L. C., et al. (2001). Circulation, 103, 2272–2276.
Manukhina, E. B., Downey, H. F., & Mallet, R. T. (2006). Experimental Biology and Medicine, 231, 343–365.
Zaobornyi, T., Gonzales, G. F., & Valdez, L. B. (2007). Frontiers in Bioscience, 12, 1247–1259.
Acknowledgments
We are thankful to Dr. Y. K. Sharma and Dr. S. Anand for providing the help in statistical analysis. We also thank Mr. B. Singh for providing the help during animal handling and experimentation.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Singh, M., Thomas, P., Shukla, D. et al. Effect of Subchronic Hypobaric Hypoxia on Oxidative Stress in Rat Heart. Appl Biochem Biotechnol 169, 2405–2419 (2013). https://doi.org/10.1007/s12010-013-0141-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12010-013-0141-2