
Abstract
1. Larvae of multiple click beetle species, i.e. wireworms, are important pests of agriculture worldwide, and are generally managed with insecticides. Conceivably, semiochemical-based management tactics that target male beetles and reduce the mating success of females, would reduce the formation of new larvae in the field.
2. Using two of the best studied species, Agriotes obscurus (AO) and A. lineatus (AL), we evaluated the ability of male beetles to find traps that simulate calling female beetles in field plots treated with various formulations and densities of pheromone-treated substrates. Four disorientation studies were conducted, and the response of wild and marked-released beetles inferred from frequent trap collections.
3. Beetle responses differed between male AO, female AO, and male AL. The presence of AO pheromone increased male AO movements, reduced captures in baited traps, and attracted wild AO beetles into the plots. By the final experiment, the pheromone-treated substrate effectively disoriented male AO for > 17 d. However, treatment with AL pheromone reduced male AO movements and/or repelled them from the plots. Female AO were slightly attracted to their own pheromone early in the season, but not thereafter. Treatment with AL pheromone attracted male AL into plots but did not increase their activity. Both AO and AL pheromone disrupted male AL behaviour, but less significantly than observed for male AO.
4. These results suggest pheromone-based click beetle mating disruption for wireworm management is feasible. However, further work is required to determine application rates and methods, and if this can be developed for other species.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Methods to manage insect pests in large scale agricultural production have continually diversified beyond the narrow use of synthetic chemical pesticides that gained prominence after WWII. Many pesticides used during that time were eventually found to have undesirable environmental effects (Carson 1962), so with the sanctioning of integrated pest management (IPM) in the 1950s, non-chemical pest control methods that were complimentary to pesticides became supported both culturally and legislatively (Gay 2012). While less detrimental synthetic pesticides continue to be developed, current progress in pest management is comprised largely of a variety of practices and tactics as their alternatives.
Pestilent wireworms (Coleoptera: Elateridae) are a good example to illustrate this pest management trajectory. Their control in the subterranean larval stage was highly effective using persistent organochlorines beginning in the 1940s, and in subsequent decades with carbamate and organophosphate pesticides (Vernon and van Herk 2022). These chemicals protected crops and reduced wireworm populations to such a degree that agricultural land would remain free from wireworm-related damage for several years (Strickland et al. 1962). Regulatory revisions have gradually caused the de-registration and withdrawal of almost all of the most effective wireworm control products from that era, leading to the research and development of less-persistent and less-environmentally toxic insecticides and a wide range of alternative practices congruent with IPM principles. These so-called ‘alternatives’ were most recently reviewed by Poggi et al. (2021), who covered not only the management strategies available for the pestiferous wireworm stage, but also opportunities for reducing wireworm populations in fields by targeting the egg laying adult beetle stage. Targeting elaterid beetles as an IPM approach has been enabled, to a large degree, because of the discovery of female sex pheromones (Yatsynin et al. 1996; Traugott et al. 2015) which have been shown to be strong attractants for male beetles. Their relatively easy synthesis and commercial availability have facilitated the development, demonstration and implementation of a variety of IPM tools and strategies, including: monitoring of single or multiple pest populations in fields to estimate the risk of wireworm damage (Furlan et al. 2020); direct the timing of adult sprays (Ester and Rozen 2005); facilitate mass trapping of males to reduce female mating and fecundity (Arakaki et al. 2008b; Vernon et al. 2014a); and increase biocontrol efficacy by attracting beetles to entomopathogen inoculum (Kabaluk et al. 2015).
Mating disruption is a well-recognized tactic that uses a device or substrate to dispense female sex pheromone to prevent a male insect’s orientation to its potential female mate, and is a well-employed tactic for controlling a number of insect pests (Welter et al. 2005) including scarab and cerambycid beetles (e.g. Koppenhöfer et al. 2005, Wenninger and Averill 2006, Maki et al. 2011, Arakaki et al. 2013, Sweeney et al. 2017). Research dedicated to its use in targeting elaterid beetles is limited in the literature, with notable exceptions being the work by Ivashchenko and Chernova (1995) on Agriotes spp., and Arakaki et al. (2008a) on Melanotus okinawensis Ohira. Arakaki et al’s work suggested the field-scale release of pheromones can result in significant population reductions over time, and both papers report significantly lower mating rates in females in treated areas in the year of application. Similarly, the use of synthetic pheromones in mass trapping reportedly can reduce populations of Agriotes sputator L. and A. gurgistanus (Faldermann) (Balkov and Ismailov 1991) and M. okinawensis (Arakaki et al. 2008b) when repeated over several years, and cause female M. okinawensis to mate only after the pheromone’s effect has waned (Arakaki et al. 2008b). These reports support the idea that mating disruption might be useful for click beetles, particularly for species (e.g. A. sputator, A. obscurus L) that have a relatively short activity period (approx. 3 months) whose onset is easy to predict in spring, a long (3–5 year) life history—which slows population recovery after the application of suppression tactics—and a high likelihood that beetles only mate once (Maki et al. 2011).
In developing a biologically-based attract and kill tactic targeting A. obscurus click beetles, Kabaluk et al. (2015) used corn cob grits (Kramer Industries, Inc., Conway, NH) as a substrate to dispense, en masse, the A. obscurus pheromone (Borg-Karlson et al. 1988; Yatsynin et al. 1996; Tóth et al. 2003). These pheromone granules served to attract male beetles to the Metarhizium brunneum LRC112 ‘kill’ component, with the authors recognizing the granulated formulation as having utility for click beetle mating disruption (Kabaluk 2014a, 2014b, 2017). They further tested other experimental substrate formulations including wax - paraffin ‘pellets’ used in the present study, and carnauba beads developed together with ChemTica International used in current (yet unreported) studies.
In the present research, we used corn grit pheromone granules and paraffin pheromone pellets in experiments aimed at disorienting male A. obscurus and A. lineatus (L) beetles, with the expectation that such disorientation would extend to male beetles being unable to locate mates. The following responses were anticipated in our experiments: (1) disorientation by the pheromone applied with the granules or pellets, resulting in reduced beetle catches in baited (with a point source attractant) pitfall traps. The applied pheromones could also lead to (2) agitation (i.e. increased movement; Leung et al. 2020), which would result in higher beetle captures in unbaited pitfall traps in pheromone-treated plots compared to unbaited traps in untreated plots. This increase in capture could also be due to (3) immigration into the plots due to attraction to the pheromone. By subjecting both marked-released and wild beetles to our treatments, we aimed to differentiate between an increased capture due to agitation (2, above) or immigration (3, above).
Methods and materials
Seven field experiments (Exp.) were conducted, to achieve three main objectives: (1) determine the minimum number of pheromone-treated granules (“pheromone granules”) required for attracting male A. obscurus (AO) beetles to pitfall traps in the field, i.e., to create point source attractants (Exp. 1, 2; Table 1); (2) compare the attractiveness of these granules with pheromone-treated pellets (“pheromone pellets”) that were used to disorient male AO in Exp. 6 and 7 (Exp. 3, Table 1); and (3) determine the ability of wild and marked-released male AO and male A. lineatus (AL) beetles to locate baited traps in plots to which either granules or pellets were applied (Exp. 4–7, Table 1). All experiments were run at the Agassiz Research and Development Centre (ARDC) (Agassiz, BC; Latitude: 49.242859, Longitude: -121.763138) in the spring and early summer of 2014 (Exp. 1, 4, 5) and 2017 (Exp. 2, 3, 6, 7), in a grass field (approx. 1 ha) that had not been disturbed for > 10 years other than with periodic mowing. Study areas were mowed with a lawn mower to a uniform grass height of 4 cm within 1 wk of the start of each Exp. Experiments were designed with 4–7 replicates, and arranged in either a Latin square or complete randomized block design (Table 1). Hourly temperature and precipitation data during each experiment was collected by an Environment and Climate Change Canada weather station at the ARDC (World Meteorological Organization Identifier: 71,113, approx. 100 m from the study site).
Pheromone-treated granule and pellet preparation
Pheromone-treated granules were developed from dried corn-cobs using a proprietary process by Contech Enterprises Inc. (Richmond, BC) in 2014, and by the authors in 2017, and contained 1.0% w/w pheromone for AO (1:1 ratio of geranyl hexanoate and geranyl octanoate). Granules were roughly cubical in shape. As individual granules used in Exp. 1 (part 1) varied in size (length: approx. 1.5–4 mm), and therefore in the amount of pheromone per granule, they were screened through a 2 mm sieve before use in part 2 of Exp. 1 and in Exp. 4 and 5, to ensure a consistent size of 1.5–2 mm. Granules used in Exp. 2, 3, 6, and 7 were similarly screened, but were 2–3 mm in size (Fig. 1E).

Design and placement of pitfall traps used in 2014 (A) and 2017 (B). Point source attractants were placed on the sticky surface of tape put on the bottom of the pitfall trap lids (D), and, after preliminary studies, were screened (C) to obtain a consistent size. Pheromone-treated plots for mating disruption were established by broadcast applications of treated granules in 2014 (E) and hand-placement of treated wax pellets in 2017 (F)
Pheromone pellets were developed by ChemTica Internacional SA (Heredia, Costa Rica), were cylindrical and uniform in shape and size (diameter: 3 mm, height 3 mm), and consisted of paraffin wax (80% w/w) with 15% w/w pheromone of AO (as above) or AL (1:1 ratio of geranyl butanoate and geranyl octanoate) and 5% w/w tocopherol to protect the pheromone from degradation by UV radiation (Fig. 1F). This higher concentration of pheromone in the pellets was chosen as it was expected to increase the longevity of the treatment in the field. After the granules and pellets were treated with the pheromones, they were stored at 4–5 °C to minimize pheromone dissipation until their use in the field experiments.
Pitfall traps
Traps used in 2014 (Exp. 1, 4, 5) were baited pitfall traps made by the authors, and consisted of a 590 ml plastic cup inserted snuggly into the soil so the upper lip of the cup was level with the soil surface (Fig. 1A; van Herk et al. 2022b). Each trap was placed in the centre of a 20 cm × 20 cm × 5 cm deep depression in the field created by the uniform removal of sod with a flat-edged shovel, and was covered with a 15 × 15 cm plywood lid that was elevated approx. 1 cm above the trap opening by small corner posts (Fig. 1A). Traps used in 2017 were Vernon Pitfall Traps® (VPT), inserted directly into the field without prior removal of sod (Fig. 1B; van Herk et al. 2022b). All collected beetles were identified to species, and depending on the number collected, the sex was determined of either all beetles (Exp. 1, 4–7), or of subsamples taken from all treatments (Exp. 2–3).
Beetle marking
Marked beetles were released in all four male disorientation studies (Exp. 4–7). Within one week before each release, wild male AO beetles were collected in pheromone baited traps at ARDC and stored in well ventilated Plexiglas cages (30 × 100 × 100 cm). The day before each release, an approximately 2 mm × 2 mm spot on the beetle pronotums were manually marked with various acrylic paints (Golden Artist Colors, Inc., New Berlin, NY) using fine detail paint brushes. Different colors were used for each release in Exp. 4 [release 1: Iridescent Silver (Fine) (#8573), Iridescent Copper (Fine) (#8571); release 2: Teal (#8547)] and Exp. 5 [release 1: Iridescent Gold (Fine) (#8572); release 2: Vat Orange (#2403)]. For Exp. 6 and 7, four colors were used to mark each species [Iridescent Gold (Fine), Vat Orange, Iridescent Pearl (Fine) (#8574), Cerulean Blue (#8526) for AO; Quinacridone Red (#8541), Teal, Iridescent Copper (Fine), Hansa Yellow Light (#2180) for AL], and all four were released on each release date (i.e., one color per treatment). The paints used were pre-tested on lab populations and had no detrimental effects on click beetle health or behaviour (see also van Herk and Vernon et al., 2023). During storage, beetles were fed with freshly cut apple slices every 2–3 days.
Development of pheromone-baited traps
To develop baited traps that simulated the attractiveness of calling female A. obscurus beetles, AO pheromone granules were placed on the sticky side of a folded piece of polyethylene-coated cloth duct tape attached to the bottom of pitfall trap lids (Acklands-Grainger Inc., Chilliwack, BC) (Fig. 1A and B). For Exp. 1, variable numbers of granules were attached to the wooden trap covers (Fig. 1A), so that the cumulative weight of the granules was 5, 10, 20, or 40 mg (Table 2). For Exp. 2, the tape was adhered to the bottom of VPT lids, and 1, 3, 5, or 10 granules were attached (Fig. 1B). Lids to which no granules were affixed served as blank controls for both experiments. Both Exp. 1 and 2 consisted of 25 traps (5 treatments × 5 replicates) arranged in a square grid, with individual traps spaced 5.0 m apart. Trap lids with granules were placed on 6 June and 19 June 2014, and on 21 April 2017 for parts 1 and 2 of Exp. 1, and for Exp. 2, respectively, and granules were not replaced during these experiments (Table 1). Beetles were collected 2 and 4 times in parts 1 and 2 of Exp. 1, respectively, and 6 times in Exp. 2.
The lowest amount of pheromone (i.e. fewest number or amount of granule) attractive to male AO in Exp. 1 and 2 was used to create the baited traps used in subsequent male disorientation studies (Exp. 4–5 and 6–7, respectively). Separate studies were not done to determine the lowest number of granules that was attractive to male AL, as their response was expected to be similar to that of male AO.
Pheromone attractant comparison
To compare the relative attractiveness of pheromone-treated wax pellets and corn-cob granules, a single three treatment experiment was conducted in 2017 (Exp. 3; Table 1). Treatments consisted of either a single wax pellet or a single granule affixed to the bottom of a VPT trap with duct tape (as above), with unbaited traps (tape only) serving as a control treatment. Traps were arranged in a rectangular 3 × 7 (replicate) grid, with individual traps 5.0 m apart. Traps were installed on 25 April and checked 2× per week.
Disorientation of Agriotes obscurus with pheromone granules in 2014
General methods
Two disorientation studies were conducted in 2014 (Exp. 4, 5) to determine if, and how long, field applications of pheromone granules reduced captures of male AO in pheromone-baited traps. Granules were applied to recently-mowed, circular plots (radius: 2.6 m, Fig. 3a), containing 4 pitfall traps (design and placement as in Exp. 1) arranged in a 2.5 m × 2.5 m square). Of these, 2 traps, placed diagonally from each other, were baited with 3–6 granules (i.e., 5 mg), and the other two served as unbaited controls. Trap lids in baited traps were replaced approx. 2× per week with lids containing new the same amount of granules, and were the same for all four treatments in Exp. 4 and 5. Plots were treated with one of three rates of granules, or left untreated (i.e., the control). Both experiments consisted of 16 plots (4 treatments × 4 replicates), with 15.0 m between adjacent plots and using the same randomization (i.e., the same plots were untreated, or received the lowest, middle, and highest rates). The application of granules to the plots during Exp. 4 was not thought to affect beetle behaviour in Exp. 5 as the capture data indicated the granules were no longer effective by the last collection period in Exp. 4, and as plots had been mowed between Exp. 4 and 5 (i.e., on 20 June). All traps were cleaned between Exp. 4 and 5.
Pheromone granules in both experiments were applied manually using shaker vials, consisting of 37 ml polystyrene containers fitted with a perforated lids. Starting at the centre of the plot and moving outwards at a constant speed and in a circular pattern, vials were shaken 50 cm above the field surface until empty. Disposable plastic boot covers were put over footwear when entering plots for granule applications, trap checks, and beetle releases, etc., and these were changed between treatments to prevent cross-contamination. Calculations to determine the amount of granule to broadcast were based on the amount of granule used per baited trap (5 mg) and the area of individual trap lids (0.0225 m2), which equaled 222 mg granule/ m2 (i.e., 2.22 mg pheromone/ m2, as the pheromone content was 1% w/w). The amounts of pheromone selected for treatments in Exp. 4 were 0.25×, 1×, and 2× this rate, or 0.57, 2.27, and 4.49 mg pheromone/ m2, which was equivalent to 1.2, 4.82, and 9.54 g granules per plot (area of plot = 21.2 m2). For Exp. 5 treatment rates were 0.25×, 2×, and 4.2× that of baited traps, or 0.57, 4.54, and 9.27 mg pheromone/ m2, which was equivalent to the 1.2, 9.64, and 19.67 g granules per plot.
Marked beetles were released to all plots to provide a second, standardized variable that could be measured to assess if and how long granules disrupted beetle behaviour. Beetles (30 / plot) were released twice for both Exp. 4 and 5, from uncovered, 10 cm Petri dishes placed in the centre of each plot and covered with an unbaited trap lid, so that the beetles could easily disperse. It was hypothesized that the granules would initially prevent marked and wild male beetles from finding the baited traps, which would result in similar numbers being collected in control and baited traps, and that the numbers collected with baited traps relative to the control traps would increase over time as the granules became less effective. For both Exp. 4 and 5 marked beetles were released twice to ensure capture numbers were sufficient to compare captures between traps types, both immediately after granule application, and after granules had aged.
Application dates and times
For Exp. 4, pheromone granules were applied on 11 June (4 PM), and baited trap lids were placed 1 h later. To ensure baited traps remained attractive, baited lids were replaced with newly baited ones on 16 June (4 PM) and 18 June (3 PM) (i.e., at 120 and 167 h after granule application). Marked beetles were released on 11 June (5 PM), and 16 June (4 PM) (i.e., at 1 and 120 h after granule application). All traps were checked at 8–9 AM and 3–4 PM on 12, 13, 16, 17, 18, and 19 June, and on 20 June (AM only). Captures of 12 + 13 June, and 17 + 18 June were combined to determine the number of wild and marked beetles captured within approx. 48 h of the first and second releases, respectively (Table 3). Captures of other dates were similarly combined for analysis to determine the numbers captured after the first 48 h of the first and second releases.
For Exp. 5, pheromone granules were applied on 23 June (11 AM), and baited trap lids placed 1 h later and replaced on 25 June (1 PM), 27 June (1 PM), 30 June (9 AM), and 2 July (10 AM) (i.e., at 1, 50, 98, 166, and 215 h after granule application). Marked beetles were released on 23 June (1 PM), and 27 June (1 PM) (i.e., at 2 and 98 h after granule application). All traps were checked at 8 PM on 23 June and at 8–9 AM and 3–4 PM on 24, 25, 26, and 27 June, and at 8–9 AM on 30 June and on 2 and 4 July. Captures of the first four trap checks were combined for analysis to determine the number of wild and marked beetles captured within approx. 48 h of the first release. The first trap check after the second release was conducted 3 h after the release as a considerable amount of precipitation was expected for the following day (26 mm, Fig. 2A). As a high number of beetles were captured during this collection, and the next trap check was 3 days later, these data were not combined with that of other dates. Captures from 25 June (PM) to 27 June (AM) were combined to determine the number captured for the remainder of the first release period, and captures of the last 3 checks of Exp. 5 were combined to determine the numbers captured for the remainder of the second first release (Table 4).

Mean daily temperature (line) and total precipitation (bars) during experiments in Agassiz, British Columbia in 2014 and 2017, to develop point source attractants and evaluate mating disruption tactics for Agriotes obscurus and A. lineatus
Disorientation of Agriotes lineatus and A. obscurus with pheromone pellets in 2017
General methods
Beetle disorientation studies were conducted for both AO and AL in 2017 (Exp. 6, 7; Table 1) using uniform arrays of pheromone pellets. Pellets were placed in recently-mowed, circular plots (radius: 3.1 m, Fig. 3b), containing 6 VPT traps (installed as in Exp. 2) arranged in a hexagon array with 1.2 m spacing between traps and all traps 1.2 m from the plot centre. Of these, 2 traps, placed diagonally from each other, were baited with a single pheromone granule (concentration: 1% w/w, size 2–3 mm) for AO, 2 traps were baited with a single pheromone granule for AL, and the remaining two served as unbaited controls. Baited traps were refreshed periodically by replacing the trap lids with lids containing new granules. AO and AL-baited traps were the same for all four treatments in Exp. 6 and 7. AO and/or AL pellets (concentration: 15% w/w) were manually placed in a uniform, rectangular grid with 1.0 m or 0.5 m between pellets in Exp. 6 and Exp. 7, respectively, resulting in 32 and 109 pellets per plot (Fig. 3B). For plots in which both pellet types were placed, AO and AL pellets at each location were spaced 30 cm apart. Each pellet was affixed to the soil surface with a small, brightly colored metal pin, which facilitated pellet retrieval at the end of the experiments. Sixteen plots (4 treatments × 4 replicates) were set up, with 8.0 m between adjacent plots. The same randomization was used for Exp. 6 and 7, with the same plots serving as the control treatment (i.e., no pellets), or treated with the AO, AL, or both pellet types. At the end of Exp. 6, all pellets were removed, all traps were cleaned, and plots were mowed (on 29 May), and hence the Exp. 6 treatments were not expected to interfere with beetle behaviour in Exp. 7.
Marked beetles (30 / plot) were released 1× and 2× to all plots in Exp. 6 and 7, respectively, using the release methods described for Exp. 4 and 5. Disposable plastic boot covers were again put over footwear when entering plots for pellet placement, trap checks, etc., and changed between treatments to prevent cross-contamination between plots.
Based on the mean weight of the granules used in Exp. 6 and 7 (Table 2), the concentration of pheromone per granule, and the area of a trap lid (radius = 8.25 cm), the amount of pheromone in each baited-trap was approx. 1.73 mg / m2 (i.e., 3.7 mg granule / lid, 0.0214 m2 lid size = 173 mg / m2, or 1.73 mg pheromone/ m2) (see Table 2 and below for granule weight approximations). In comparison, based on the weight of the pellets (approx. 28 mg), their pheromone content, and the density applied to plots (1 or 4 / m2), the amounts of pheromone selected per plot for Exp. 6 and Exp. 7 were 4.2 and 16.8 mg pheromone/ m2, or 2.4× and 9.7× that of the baited traps, respectively.

Placement of pitfall traps baited with point-source attractants for Agriotes obscurus (AO) and A. lineatus (AL), and unbaited control traps (C) in mating disruption experiments. Small brown hexagons in (A) represent granules broadcast in Exp. 4 and 5. White circles in (B) show placement of wax pellets in Exp. 6. In Exp. 7 wax pellets were similarly placed, but with 0.5 m spacing. Marked beetles were released at the centre of each circular plot, marked with a gray circle
Application dates and times
For Exp. 6, pheromone pellets were applied on 18 May (4 PM), and baited trap lids placed on 19 May (9 AM), and replaced with new baited trap lids on 23 May (9 AM) (i.e., at 113 h after pellet application). Marked beetles were released on 19 May (1 PM), and traps were checked at 8–9 AM on 19, 23, and 26 May. Beetles were also collected before pellet application with both baited and unbaited traps between 17 May (3 PM) and 18 May (3 PM), and after pellet application with only unbaited traps between 18 May (3 PM) and 19 May (9 AM).
For Exp. 7, pheromone pellets were applied on 30 May (10 AM), and baited trap lids placed on 31 May (9 AM), and replaced with new baited trap lids at 9 AM on 2, 5, 9, 12, 16 and 19 June. Marked beetles were released at 1 PM on 31 May and 9 June, and traps were checked at 3 PM on 31 May and 9 June (2 h after the two marked beetle releases, respectively), and at 8–9 AM on 2, 5, 9, 12, 16, 19, and 23 June. Beetles were also collected after pellet application with (only) unbaited traps between 30 May (10 AM) and 31 May (9 AM). Captures on 31 May (3 PM) and 2 June were combined for analysis, as were captures on 9 June (3 PM) and 12 June, and those on 16, 19, and 23 June.
Statistical analyses
All statistical analyses were performed in SAS v.9.04 (SAS Institute, Cary, NC, USA). For all experiments, the total number of wild or marked AO beetles collected per trap were analyzed with a generalized linear model (Proc GENMOD), using a log-link function and a negative binomial distribution. Generalized linear models with a binomial distribution and a logit link function (Proc GENMOD) were used to compare the proportion of male AO between treatments in Exp. 2 and 3, and to compare the proportion AO beetles collected in AO traps (and AL in AL traps) between treatments in Exp. 4–7. Pairwise comparisons between treatments were done with the ‘lsmeans’ statement, using Tukey’s adjustment. For Exp. 4–7, several trap collections were sometimes combined for analysis, either to determine beetle (re)capture approx. 48 hr after marked beetle releases, or to ensure numbers were robust enough for analysis.
Results and discussion
Calculation of granule amount needed for baited traps
Traps baited with screened pheromone granules collected predominantly male AO beetles, i.e., all 725 beetles in Exp. 1 (part 1), 222 of 225 beetles in Exp. 1 (part 2), and 1478 of 1577 beetles that were sexed in Exp. 2. A total of 3528 AO and 28 AL were collected in Exp. 2. For both Exp. 1 and 2, beetle numbers were significantly higher in treatments with granules than in the unbaited control, with catch numbers generally increasing with the amount of granules applied (Table 2). Since in both studies beetle captures in traps with the lowest amount of pheromone granule (5, approx. 3.7 mg, respectively, Table 2) were considerably higher than in the unbaited control traps (12.1, 5.4, 5.0× for Exp. 1 (part 1), Exp. 1 (part 2), and Exp. 2, respectively), these lowest amounts were used to create baited traps for the disorientation studies (Exp. 4–7).
Unlike for Exp. 1, granules used in Exp. 2 were not weighed. However, their weight can be approximated (Table 2) based on their similar composition to, and size (length: 2–3 mm) relative to those used in Exp. 1 (part 2) (length: 1.5–2 mm). Both granule types were similar in composition and shape, with those in Exp. 2 approximately double in volume and therefore in amount of pheromone per averaged-sized granule. Linear regression of the mean number of screened granules in Exp. 1 (part 2) (Table 1) to their weight (in mg) indicated that weight was 1.87 × granule number (R2 = 0.98), so that one average-sized granule in Exp. 2 would weigh approx. 3.7 mg. It was not determined in these experiments how similar the pheromone amounts contained in the 5 mg of granule used in 2014, or in the one granule used in 2017, were to that produced by a calling female AO in the field. However, based on the low trap catch in the baited traps relative to regular pheromone traps deployed at ARDC, the pheromone amounts used in these baited traps were considered to be low. Future work should measure both the amount emitted by female AO and AL, and the attractiveness of traps containing a live, unmated female beetle relative to the baited traps used herein. For these proof-of-concept disorientation experiments for AO and AL, however, it was not thought critical that baited traps equaled female beetles in attractiveness.
Comparison of pheromone attractants used in experiments
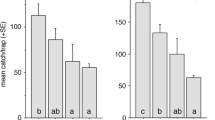
A total of 1928 AO, and 16 AL were collected in the pheromone attractant comparison experiment (Exp. 3), of which most (1000 of 1006 AO sexed) were male. Numbers of male AO differed significantly between treatments (Chi = 52.92, df = 2,18, P < 0.0001), with significantly (P < 0.0001) more beetles collected in traps baited with the pellet (mean: 232.9, SEM: 32.5) than in traps baited with a granule (mean: 37.9, SEM: 3.1) or in unbaited control traps (mean: 4.7, SEM: 1.8). Captures in the latter two treatments were also significantly different from each other. The proportion of male AO in treatments differed significantly between treatments (Chi = 17.17, df = 2,17, P = 0.0002), with more females (P < 0.01) in the control than in the pheromone treatments, but no difference (P > 0.05) in the proportion males collected in traps baited with the pellet or granule. The higher captures in pellet-baited traps was expected as they contained approx. 114× the amount pheromone (pellets weighed approx. 7.6× granules, pheromone content w/w in pellets was 15× that of granules).
Disorientation of Agriotes obscurus with pheromone granules in 2014
A relatively low percentage of marked beetles were recaptured in baited and unbaited traps (combined) during the first 48 and 44 h period (respectively) in Exp. 4 (range 5–20%) and Exp 5 (range 9–33%). There was no significant difference (P < 0.05) in the number of marked and wild male AO beetles collected in the unbaited and baited traps in the first 48 and 44 h (respectively) after granule application in both Exp. 4 and Exp. 5 for all three treatments to which granules were applied (Tables 3 and 4), indicating beetles were as likely to find baited traps as the control traps immediately after granules were applied in these treatments. In contrast, significantly more beetles (both types; 15.6× and 14.1× more wild, respectively) were collected in the baited traps in the control treatments (Tables 3 and 4). Consequently the proportion of AO collected with baited traps differed among treatments, decreasing with the amount of granules applied (for wild beetles) (Tables 3 and 4). For Exp. 4, significant differences between trap types were observed for both marked and wild male AO beetles for all treatments in all subsequent collections for which data could be analyzed, with the exception of wild beetles collected 120–168 h after granule application in the highest rate treatment (Table 3). While all beetles in the latter were collected in the baited traps, this exception, and the numerically lower numbers of both marked and wild beetles collected at the two highest rates relative to the control and low rate treatment during this trapping period, suggests some residual effect of the granules on beetle behaviour. Hence these results indicate male beetles were deterred from finding the baited traps when granules were applied at all three rates for up to 120 h after granule application in Exp. 4, but that this was significant only for the first 48 h period. For Exp. 5, there was also no significant difference between unbaited and baited traps in the number of wild beetles collected 44–93 h after granule application for the two highest pheromone rates, and a significant decrease in the proportion of wild AO collected in baited traps for the two highest granule rates relative to the control for this trapping period (Table 4). This suggests that beetles were deterred from finding the baited traps in the highest rate treatments longer in Exp. 5 than in Exp. 4, possibly due to the higher rates applied in Exp. 5, or to the lower amount of precipitation early in this experiment (Fig. 2).
Untreated control traps placed in the treatment plots captured numerically more wild (up to 10.4×) and marked beetles than control traps placed in control plots in the first 48 and 44 h (respectively) after granule application in both Exp. 4 and Exp. 5. This was also observed for wild beetles collected 48–120 h after application in Exp. 4, and for both wild and marked beetles collected 46–95 h after application in Exp. 5 (Tables 3 and 4), and indicates beetle movement increased in treated plots as a result of the granules. Total captures of marked beetles were significantly reduced in the 0–48 h after application period for all three rates in Exp. 4 (range: 67–73%; Table 3), and in the 0–44 h period for the highest rate in Exp. 5 (72%; Table 4). In contrast, the total number of wild beetles caught was similar, or higher, in the treated vs. untreated plots during both the 0–48 h and 48–120 h collection periods in Exp. 4 (Table 3) and in the 0–44 h and 95–101 h collection periods in Exp. 5. The disparity in the response of marked beetles (which have fixed numbers and were released at the centre of the plots), and the wild beetles (which are dispersed throughout and outside the plots), indicates the latter were being attracted into the plots by the granules.
During the last collection periods of Exp. 4 and 5, nearly all marked and wild beetles were collected in baited traps, the total number of wild beetles collected was similar between all treatments, and the total number of marked beetles was the same or increased with rate (i.e., the remaining marked beetles were being collected by the baited traps), all of which suggests the granules were no longer noticeably altering beetle movement.
Disorientation of Agriotes obscurus and A. lineatus with pheromone pellets in 2017
Beetle response in the absence of pellets
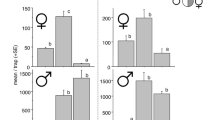
There was no significant difference between treatments in the total number of male AO (range: 30.1–38.3), female AO (range: 15.3.1–17.8), or male AL (range: 15.8–19.8), collected prior to the application of pellets in Exp. 6, indicating a relatively uniform population of both species across the study area (Table 5). As expected, more male AO and AL were collected with traps baited with their own pheromone than with unbaited traps or traps baited with the other pheromone, but the differences were more pronounced and only significant for AL (Table 5). In contrast, the number of female AO collected was similar between trap types (range: 4.0-7.8 for all trap types and treatments), indicating no significant attraction to either pheromone.
Beetle response in the presence of pellets: captures in unbaited traps
Numerically more male AO were collected in treatments to which AO, or both AO and AL-treated pellets were applied (mean = 11.6, 9.1 per plot, respectively), than in the untreated control (6.9), or AL-pheromone treated plots (4.8) at the start of Exp. 6, suggesting the AO pheromone caused beetle agitation and/or immigration into the plots, but that this was reduced by the AL pheromone (Table 5). A similar, but less pronounced trend was observed for female AO, with more beetles collected in AO and AO + AL-treated plots (4.3, 5.4, respectively), than in the control (3.3), suggesting some attraction to the AO conspecific pheromone. Trapping with unbaited traps after pellet application at the start of Exp. 7 also indicated significantly or numerically more male AO collected in treatments to which AO or both pellets were applied (6.3, 2.4, respectively) than if no (0.8) or only AL (0.9) pellets were applied, but indicated no difference in female AO captures (range 1.0–1.5) (Table 5).
Female AO response in the presence of pellets: captures in baited traps
Similar numbers of female AO were collected between treatments, and between the three trap types within each treatment, for all three collection dates in Exp. 6 (range: 0.5-2.0, 7.8–14.5, 2.0-4.8 for the three collections, Table 6), indicating female AO were not attracted to either AO or AL-baited traps, and were not noticeable affected by the presence of either pellet type.
Male AO response in the presence of pellets: captures in baited traps
In Exp. 6, significantly fewer marked AO were collected in both treatments with AO pellets during the first collection period (reduction relative to control treatment: 77–83%), and in the number of wild AO collected during the third period (77–78%) (Table 7). Additionally, 63–68% fewer wild beetles were collected during the first period. There was also no significant difference (P < 0.05) between unbaited and AO-baited traps in the number of beetles collected in the first (marked and wild) and second (wild only) periods in treatments with AO pellets, and there were significant differences between treatments in the proportion of AO collected in AO traps for both marked (collection 1) and wild beetles (all 3 collections) (Table 7). These response all indicate that the presence of the pellets disrupted male AO behaviour throughout the experiment. However, significantly more wild AO were collected in AO-baited traps than in unbaited traps by the third period (Table 7), indicating male beetles were able to locate the baited traps at the end of the experiment.
In Exp. 7, significantly fewer marked AO were collected in both treatments with AO pellets during the first and fifth periods (70–73%, 58–67%, respectively) (Table 8). Reductions in the number of wild beetles collected was less pronounced in these treatments (e.g. 23–57% in the first period), and generally not statistically significant (Table 8). The difference in response between marked and wild AO is likely due to immigration into the plots due to attraction to the pellets by the latter. Captures of wild AO in plots with AL pellets were numerically or significantly lower than in control plots for all periods (Table 8), indicating either reduced movement within the plots, or emigration from the plots. Comparison of capture reductions during the first period of wild and marked beetles (75% vs. 33% respectively), suggests both processes occurred, as marked beetles would be more concentrated in the centre of the plots (i.e., near their release points), and wild beetles were dispersed throughout and outside the plot area (Table 8).
Significantly more marked AO were collected in AO-baited than in control traps in the first and fourth collections in control plots and plots treated with AL pellets in Exp. 7, but not in plots treated with AO or both pellet types (Table 8), suggesting beetles were disoriented in the latter. Similarly, there was no significant difference in the number of wild AO collected in AO-baited vs. control traps in plots with AO (second, third, and fifth collections) or both AO and AL pellets (all collections) (Table 8).
More marked and wild AO were collected in control traps when AO or AL pellets were applied (6.7× more for both beetle types in the first collection) relative to plots without pellets. In all cases, this increase in catch was more pronounced in the two treatments with AO pellets than in the AL (alone) treatment (Table 8), suggesting that the presence of AO but not AL pellets increased male AO movements. In contrast, captures of marked and wild AO beetles in control traps in all four treatments were similar for each collection period in Exp. 6, indicating no increase in beetle movement with the lower pellet density (Table 7).
Male AL response in the presence of pellets, captures in baited traps
In Exp. 6, significantly fewer marked AL were collected in both treatments with AL pellets during the first collection period (81–86%), and with both pellet types in the second collection period (60%) relative to the untreated control treatment (Table 9). Numbers of wild AL were numerically lower in both treatments containing AL pellets for the first two collection periods, indicating the pellets were disrupting beetle movements (Table 9). Significantly more wild AL beetles (P < 0.05) were collected in AL-baited than in unbaited traps in all treatments in the second and third collections, but not in the first collection, when overall beetle numbers were low (Table 9). While captures of marked beetles were low, the proportion of both marked and wild AL collected in AL traps was consistently lower in plots treated with both AL and AO pellet types for the first two collections (Table 9), indicating AL beetles also responded to the presence of AO pheromone. The similarity between all treatments in the total wild AL catch, and in the proportion of AL collected in AL traps (Table 7), indicates that, unlike observed for AO, neither pellet type had an observable effect on male AL behaviour by the third collection period.
In Exp. 7, significantly fewer marked AL were collected in both treatments with AL pellets during the first and fourth collection periods (70–91%, 72–75%, respectively), indicating these beetles were prevented from finding the baited traps or (less likely) reduced beetle movements overall (Table 10). In contrast, numbers of wild AL in all treatments with AO or AL pellets were similar (first two collections), or numerically higher (last three collections) than those in the untreated control (Table 10), suggesting wild AL were moving into the plots and that there was no reduced beetle movement. Significantly more wild AL (P < 0.05) were collected in AL-baited traps than in unbaited traps in both treatments with AL pellets in all five collections, indicating AL beetles were able to locate the baited traps throughout the experiment (Table 10). Unlike wild AL, there was no significant difference in the number of marked AL collected in control vs. AL-baited traps in the two treatments with AL pellets during the first collection period (Table 10), suggesting some disorientation did occur. The difference between the marked and wild AL beetles suggests that immigration into the plots of the latter affected the baited:unbaited trap ratios.
With the exception of the second collection of wild AL in the combined pellet treatment in Exp. 6, captures of marked and wild AL beetles in untreated control traps in all four treatments were similar for each collection period in both Exp. 6 and 7, indicating no increase in beetle movement with the either the 0.5 or 1.0 m granule density (Table 9).
Evaluation of disorientation experiments
Our study sought to determine if broadcast applications of granulate pheromones can be used to disorient male A. obscurus and A. lineatus mate finding behaviour, and consequently disrupt mating, in the field. Based on pheromone-based mating disruption work by others (e.g., Koppenhöfer et al. 2005, Wenninger and Averill 2006, Maki et al. 2011, Sweeney et al. 2017), we anticipated three responses if disorientation occurred in our experiments: (1) reduced captures of beetles in baited traps in plots to which granules or pellets were applied, as evidence beetles were less able to find these traps, and an indicator that these treatments potentially disrupted mating; (2) increased captures of beetles in unbaited traps in plots to which granules or pellets were applied, as evidence of increased beetle movement (agitation) and/or of increased numbers in the plots (due to immigration, for wild beetles only); and (3) increased total captures in plots (baited and unbaited traps combined), as evidence of immigration into the plots due to attraction to the pheromone. Conversely, decreased total captures would suggest emigration from the plots, reduced beetle activity, or simply the reduced ability of beetles to find baited traps. All these responses were observed, but with notable differences between experiments and species.
Data from all four experiments show both that the granules and AO-pheromone treated pellets increased male AO movement inside the plots, reduced captures in AO-baited traps, and attracted wild AO beetles into the plots. While male AO could locate baited traps at the end of all four experiments, some disorientation was still evident at the end of Exp. 6 and 7. Male AO took longer to find the baited traps in Exp. 5 than in Exp. 4, possibly because of the higher rates applied and/or the higher precipitation during Exp. 4 (Fig. 2). AO-treated pellets were effective longer than the granules. Based on the collection periods in which captures were significantly reduced, we conclude the duration pellets and granules effectively disoriented male AO appeared to be > 17 d for Exp. 7, up to 7 d for Exp. 6, but only 4–5 d for Exp. 4–5.
Data from both baited and unbaited traps indicate that in the presence of AL pheromone, male AO reduce their movements within and/or moved out of the plots. Repellency of male AO to the pheromone of AL has been observed before (Vernon et al. 2014b; van Herk et al. 2022). Interestingly, female AO appeared to be attracted to their own pheromone early in the season, but not thereafter and were not noticeably attracted to AO or AL-baited traps or affected by the presence of either pellet type. Early season attraction of female AO to their own pheromone has also been observed previously (van Herk et al. 2022a).
Like AO, AL pellets appeared to attract male AL beetles into treatment plots. However, unlike AO, there was no indication that the presence of AL or AO treated pellets increased the activity of male AL. Both pellet types appeared to disrupt male AL behaviour and reduced captures in AL-baited traps. However, the reduction in male AL captures in traps baited with their pheromone was notably less than observed for male AO in both Exp. 6 and Exp. 7, and generally not statistically significant. The different response from the two species may be due in part to the higher overall activity rate of AL (Hicks and Blackshaw 2008).
Potential of pheromone-based click beetle mating disruption for wireworm management
In finding that male AO and AL beetles were disoriented in the presence of granulate pheromone (both corn grit granules (Exp. 4, 5) and paraffin pellets (Exp. 6, 7)), we expect that they will also be unlikely to locate calling (pheromone-emitting) females, providing the attracting compounds comprise, solely, of those tested in our experiments. A limitation of our study was that the pheromone emission rate from the point source attractants compared to a calling female under natural conditions was unknown. If the release rate from a calling female is equal to or less than the baited traps used in our experiments, then our granulate pheromone rates would be expected to prevent male beetles from finding her. If a higher release rate from either calling females or baited traps was shown to increase male attraction when situated among the granulate pheromone rates used in our experiments, then simply an increase in granulate rate could re-establish male disorientation. Further research is required to (i) determine pheromone emission rates from calling female beetles; (ii) establish minimum pheromone rates that would disable male beetles from locating female beetles; and (iii) determine if increased male and female movements in the presence of pheromone granules lead to increased encounters and matings.
We found some immigration of wild beetles into pheromone-treated plots and would expect that on a field scale, granulate pheromone application would similarly attract beetles from refugia. Once attracted, however, we’d expect immigrants to be equally subjected to disorientation as the beetles resident in the field. If applied to refugia directly, a granulate pheromone formulation could serve to reduce emigration from these areas – both by the attractive quality of the pheromone and over years through beetle attrition, created by mating disruption.
In laboratory studies, Leung et al. (2020) found that in comparison to ambient conditions, the presence of pheromone increased the walking speed, distance traveled, and duration of movement of A. obscurus beetles. We similarly found evidence of this, albeit in AO only. In the presence of applied granulate pheromone there were greater numbers of beetles in control (unbaited) traps, which suggested increased movement and probability of falling into a trap. Increasing beetle activity as a result of granulate pheromone application might be exploited for other purposes. Kabaluk (unpublished data) improved the targeting of click beetles with an aqueous spray of Metarhizium brunneum conidia. He found that the conidia dose of beetles in the presence of pheromone granules, applied pre-spray to agitate beetles, was significantly higher than in the absence of granules, presumably by increasing beetle encounter with spray droplets on leaves and by more beetles crawling to the ends of leaves and being exposed to direct spray (personal observation).
For click beetle mating disruption to be effective, pheromone granules need to remain effective for the duration of the mating period (i.e. approx. the first two months of their activity period), which can potentially be done by using different materials, higher concentrations, or adding compounds that retard degradation (e.g. tocopherol), provided these compounds are acceptable for use in organic agriculture. For A. obscurus and A. lineatus, this period could be up to four months in warm and cold temperate climates (e.g. van Herk and Vernon 2020, 2023), otherwise multiple application of pheromones would be required. Our study showed that the effectiveness of the pheromone granule application was decreased or lost following rain. Based on these data we have developed a carnauba wax bead pheromone granular formulation with ChemTica Internacional, which will attract beetles for up to four weeks, even after exposure to rain and sunlight (unpublished data). Due to approx. 3 month activity period of these species, however, three or four applications would still be required to effectively disrupt mating throughout the entire period of adult activity. Fewer applications, or even a single application of this particular formulation might be better-suited to other species. For example, Limonius canus (LeConte) and L. californicus (Mann.) have a shorter swarming period than AO and AL (Gries et al. 2021; van Herk et al. 2021; Lemke et al. 2022) requiring a shorter period for the applied pheromone to interfere with mating. Furthermore, exposure to limoniic acid has been shown to desensitize L. californicus male beetles to the call of females (Lilly and McGinnis 1968). Depending on the length of desensitization, the particular timing and frequency of pheromone granules, in an applied setting, would need to be considered for this species, and other potential species that might possesses the same quality. It is possible that extreme rates of applied pheromone might serve to desensitize any species, although this remains untested.
The duration of the life cycle among Elateridae is variable, with Vernon and van Herk (2022) noting the studies describing such durations for a number of pest species. For granulate pheromone to be an effective wireworm control tactic, they would need to be applied for several consecutive years, or used in conjunction with an entomopathogen (e.g. Kabaluk et al. 2015). Although theoretical, if granulate pheromone could prevent every potential mating event, they would need to be applied until every soil-borne larva emerges as an adult, in both the cropland and refugia so that immigrants could not recolonize cropped areas. In a practical setting, the efficiency of mating disruption would be imperfect, and attaining sub-threshold larval levels would be an acceptable goal, particularly if such levels can be maintained by other control measures. The efficiency of mating disruption can be assessed by yearly measurement of neonate larvae or young instars from soil samples, in either experimental or on-farm settings.
The constituent compounds for A. obscurus and A. lineatus pheromones, are the esters geranyl octanoate, geranyl hexanoate, and geranyl butanoate, and readily available and relatively inexpensive. For AO for example, the lowest pheromone rate with which we observed disorientation of male beetles equated to 5.7 g/ha (0.57 g/m2; Exp. 4). The highest rate of pheromone used in our study equated to 168 g/ha (16.8 mg/m2; Exp. 7). In 2021, Penta Manufacturing Company (Livingston, New Jersey, USA) sold 1 kg of the constituent compounds geranyl hexanoate and geranyl octanoate for USD $275 and $295, respectively (each in a 1 kg quantity). Using these figures, the cost of one application would range from USD $1.62/ha to $47.88/ha per year. Multiple applications and multiple species would increase costs proportionally, but savings would be gained by determining the lowest effective rate, creating a long-lasting formulation, and purchasing greater pheromone quantities at a lower cost per unit. However, this tactic will likely be applied only to those field areas where pest pressure warrants applications (i.e., “hotspots”), and/or to field headlands, as these serve as high-population reservoir areas from which beetles enter the fields to lay eggs (Vernon and van Herk 2018). The ultimate goal is to create an inexpensive granule that emits a male-disorienting rate of pheromone for the duration of the mating period, and can be dispensed using a granule applicator such as that used to broadcast fertilizer granules. For refugia and field margins, application by drone is possible.
With the availability of pheromones for the main European pest species (Tóth 2013), and the recent identification of semiochemicals for pestilent species of Elateridae in North America (see Serrano et al. 2018, Singleton et al. 2022, 2023; Williams et al. 2019; Pellegrino et al. 2021; Gries et al. 2022), we believe that effective mating disruption can progress from the conceptual framework we have presented in our research, and become a new and widespread tool for multiple species. Providing that both adults and larval species can be taxonomically identified, protocols can be developed with experiments on a single species for which a pheromone is available, and elements of the protocol extended to other species for which pheromones are also available or can be identified.
References
Arakaki N, Nagayama A, Kobayashi A, Hokama Y, Sadoyama Y, Mogi N, Kishita M, Adaniya K, Ueda K, Higa M, Shinzato T (2008a) Mating disruption for control of Melanotus okinawensis (Coleoptera: Elateridae) with synthetic sex pheromone. J Econ Entomol 101:1568–1574. https://doi.org/10.1093/jee/101.5.1568
Arakaki N, Nagayama A, Kobayashi A, Kishita M, Sadoyama Y, Mougi N, Kawamura F, Wakamura S, Yamamura K (2008b) Control of the sugarcane click beetle Melanotus okinawensis Ohira (Coleoptera: Elateridae) by mass trapping using synthetic sex pheromone on Ikei Island, Okinawa, Japan. Appl Entomol Zool 43:37–47. https://doi.org/10.1303/aez.2008.37
Arakaki N, Hokama Y, Nagayama A, Yasui H, Fujiwara-Tsujii N, Tanaka S, Mochizuki F, Naito T, Hongo T, Wakamura S (2013) Mating disruption for control of the white grub beetle Dasylepida ishigakiensis (Coleoptera: Scarabaeidae) with synthetic sex pheromone in sugarcane fields. Appl Entomol Zool 48:441–446. https://doi.org/10.1007/s13355-013-0202-6
Balkov VI, Ismailov VY (1991) Attractant traps for elaterids. Zashchita Rastenii (Mosc) 10:21
Borg-Karlson AK, Ågren L, Dobson H, Bergström (1988) Identification and electroantennographic activity of sex-specific geranyl esters in an abdominal gland of female Agriotes obscurus (L.) and A. lineatus (L.) (Coleoptera: Elateridae). Experientia 44:531–534. https://doi.org/10.1007/BF01958940
Carson R (1962) Silent Spring. Houghton Mifflin, Boston
Ester A, van Rozen K (2005) Monitoring and control of Agriotes lineatus and A. obscurus in arable crops in the Netherlands. IOBC/WPRS Bull 28(4):81–85
Furlan L, Contiero B, Chiarini F, Benvegnu I, Tóth M (2020) The use of click beetle pheromone traps to optimize the risk assessment of wireworm (Coleoptera: Elateridae) maize damage. Sci Rep 10:8780. https://doi.org/10.1038/s41598-020-64347-z
Gay H (2012) Before and after silent spring: from chemical pesticides to biological control and integrated pest management - Britain, 1945–1980. Ambix 59:88–108
Gries R, Alamsetti SK, van Herk WG, Catton HA, Meers S, Lemke E, Gries G (2021) Limoniic acid - major component of the sex pheromones of the click beetles Limonius canus and L. californicus. J Chem Ecol 47:123–133. https://doi.org/10.1007/s10886-020-01241-y
Gries R, van Herk W, Alamsetti SK, Catton H, Meers S, Otani J, Gries G (2022) (Z, E)-α-Farnesene?sex pheromone component of female click beetle Selatosomus aeripennis destructor with intra- and inter-sexual communication function. Entomol Exp Appl 170:344–351. https://doi.org/10.1111/eea.13142
Hicks H, Blackshaw RP (2008) Differential responses of three Agriotes click beetle species to pheromone traps. Agric For Entomol 10:443–448. https://doi.org/10.1111/j.1461-9563.2008.00397.x
Ivanshchenko II, Chernova SV (1995) Biologically active substances against click beetles. Zashchita Rastenii (Mosc) 9:16–17
Kabaluk T (2014a) Pheromone granules for the attraction and disorientation of Elateridae. Agriculture and Agri-Food Canada, INV 13918
Kabaluk T (2014b) Targeting the click beetle Agriotes obscurus with entomopathogens as a concept for wireworm biocontrol. Biocontrol 69:607–616. https://doi.org/10.1007/s10526-014-9603-x
Kabaluk T (2017) Review of patent and intellectual history relating to the use of AAFC-developed pheromone granules for wireworm control. Agriculture and Agri-Food Canada, Internal Report
Kabaluk JT, Lafontaine JP, Borden JH (2015) An attract and kill tactic for click beetles based on Metarhizium brunneum and a new formulation of sex pheromone. J Pest Sci 88:707–716. https://doi.org/10.1007/s10340-015-0661-3
Koppenhöfer AM, Polavarapu S, Fuzy EM, Zhang A, Ketner K, Larsen T (2005) Mating disruption of oriental beetle (Coleoptera: Scarabaeidae) in turfgrass using microencapsulated formulations of sex pheromone components. Env Entomol 34:1408–1417. https://doi.org/10.1603/0046-225X-34.6.1408
Lemke E, van Herk WG, Singleton K, Gries G (2022) Seasonal and diel communication periods of sympatric pest Limonius click beetle species (Coleoptera: Elateridae) in western Canada. Env Entomol 51:980–988. https://doi.org/10.1093/ee/nvac067
Leung JPS, Cory JS, Kabaluk JT, Janmaat AF (2020) Effect of collection month, visible light, and air movement on the attraction of male Agriotes obscurus L. (Coleoptera: Elateridae) click beetles to female sex pheromone. Insects 11(11):729. https://doi.org/10.3390/insects11110729
Lilly CE, McGinnis AJ (1968) Quantitative responses of males of Limonius californicus to female sex pheromone. Can Entomol 100:1071–1078. https://doi.org/10.4039/Ent1001071-10
Maki EC, Millar JG, Rodstein J, Hanks LM, Barbour JD (2011) Evaluation of mass trapping and mating disruption for managing Prionus californicus (Coleoptera: Cerambycidae) in hop production yards. J Econ Entomol 104:933–938. https://doi.org/10.1603/EC10454
Pellegrino AM, Dorman SJ, Williams L 3rd, Millar JG, Huseth AS (2021) Evaluation of 13-tetradecenyl acetate pheromone for Melanotus communis (Coleoptera: Elateridae) detection in North Carolina row crop agroecosystems. Env Entomol 50:1248–1254. https://doi.org/10.1093/ee/nvab075
Poggi S, Le Cointe R, Lehmhus J, Plantegenest M, Furlan L (2021) Alternative strategies for controlling wireworms in field crops. Agriculture 11(5):436. https://doi.org/10.3390/agriculture11050436
Serrano JM, Collignon RM, Zou Y, Millar JG (2018) Identification of sex pheromones and sex pheromone mimics for two north american click beetle species (Coleoptera: Elateridae) in the genus Cardiophorus Esch. J Chem Ecol 44:327–338. https://doi.org/10.1007/s10886-018-0940-6
Singleton K, Gries R, van Herk WG, Alamsetti SK, Lemke E, Furtado K, Gries G (2022) Identification of the major sex pheromone component of the click beetle Agriotes ferrugineipennis. J Chem Ecol 48:491–501. https://doi.org/10.1007/s10886-022-01367-1
Singleton K, van Herk WG, Saguez J, Scott I, Gries R, Gries G (2023) Sex pheromone of Nearctic Agriotes mancus, and its similarity to those of three invasive Agriotes in North America. https://doi.org/10.1111/afe.12568. Agric For Entomol (in press)
Strickland AH, Bardner HM, Waines RA (1962) Wireworm damage and insecticidal treatment of the ware potato crop in England and Wales. Plant Path 11:93–105. https://doi.org/10.1111/j.1365-3059.1962.tb00175.x
Sweeney J, Silk PJ, Rhainds M, MacKay W, Hughes C, Van Rooyen K, MacKinnon W, Leclair G, Holmes S, Kettela EG (2017) First report of mating disruption with an aggregation pheromone: a case study with Tetropium fuscum (Coleoptera: Cerambycidae). J Econ Entomol 110:1078–1086. https://doi.org/10.1093/jee/tow308
Tóth M (2013) Pheromones and attractants of click beetles: an overview. J Pest Sci 86:3–17. https://doi.org/10.1007/s10340-012-0429-y
Tóth M, Furlan L, Yatsynin VG, Ujváry I, Szarukán I, Imrei Z, Tolasch T, Franke W, Jossi W (2003) Identification of pheromones and optimization of bait composition for click beetle pests (Coleoptera: Elateridae) in Central and Western Europe. Pest Man Sci 59:417–425. https://doi.org/10.1002/ps.629
Traugott M, Benefer C, Blackshaw RP, van Herk WG, Vernon RS (2015) Biology, ecology, and control of elaterid beetles in agricultural land. Ann Rev Entomol 60:313–334. https://doi.org/10.1146/annurev-ento-010814-021035
van Herk WG, Vernon RS (2020) Local depletion of click beetle populations by pheromone traps is weather and species dependent. Env Entomol 49:449–460. https://doi.org/10.1093/ee/nvaa006
van Herk WG, Vernon RS (2023) Capture of wild and marked Agriotes obscurus in pheromone traps according to distance, wind direction and date of trapping. Arthropod Plant Interactions. https://doi.org/10.1007/s11829-023-09961-0. (in press)
van Herk WG, Lemke E, Gries G, Gries R, Serrano JM, Catton H, Wanner K, Landolt PJ, Cooper WR, Meers S, Nikoukar A (2021) Limoniic acid and its analog as trap lures for pest Limonius species (Coleoptera: Elateridae) in North America. J Econ Entomol 114:2108–2120. https://doi.org/10.1093/jee/toab154
van Herk W, Vernon B, Bourassa-Tait G, Tóth M, Kovacs E (2022) Field evaluation of selected plant volatiles and conspecific pheromones as attractants for Agriotes obscurus and A. lineatus (Coleoptera: Elateridae). Insects 13(2):173. https://doi.org/10.3390/insects13020173
van Herk WG, Vernon RS, Borden JH, Ryan K, Mercer G (2022b) Comparative evaluation of pitfall traps for click beetles (Coleoptera: Elateridae). J Econ Entomol 115:582–591. https://doi.org/10.1093/jee/toab259
Vernon B, van Herk W (2018) Wireworm and flea beetle IPM in potatoes in Canada: implications for managing emergent problems in Europe. Potato Res 60:269–285. https://doi.org/10.1007/s11540-018-9355-6
Vernon RS, van Herk WG (2022) Wireworms as pests of potato. In: Alyokhin A, Rondon S, Gao Y (eds) Insect pests of Potato: global perspectives on Biology and Management, 2nd edn. Academic Press, Elsevier, pp 103–148
Vernon RS, Blackshaw RP, van Herk WG, Clodius M (2014a) Mass trapping wild Agriotes obscurus and Agriotes lineatus males with pheromone traps in a permanent grassland population reservoir. Agric For Entomol 16:227–239. https://doi.org/10.1111/afe.12058
Vernon RS, van Herk WG, Tanaka JA (2014b) Blending of pheromone lures for two exotic european pest elaterid beetles. J Pest Sci 87:619–627. https://doi.org/10.1007/s10340-014-0596-0
Welter SC, Pickel C, Millar JG, Cave F, van Steenwyk R, Dunley J (2005) Pheromone mating disruption offers selective management options for key pests. Calif Agric 59:16–22. https://doi.org/10.3733/ca.v059n01p16
Wenninger EJ, Averill AL (2006) Mating disruption of oriental beetle (Coleoptera: Scarabaeidae) in cranberry using retrievable, point-source dispensers of sex pheromone. Env Entomol 35:458–464. https://doi.org/10.1603/0046-225X-35.2.458
Williams L, Serrano JM, Johnson PJ, Millar JG (2019) 13-Tetradecenyl acetate, a female-produced sex pheromone component of the economically important click beetle Melanotus communis (Gyllenhal) (Coleoptera: Elateridae). Sci Rep 9:1–12. https://doi.org/10.1038/s41598-019-52199-1
Yatsynin VG, Rubanova EV, Okhrimenko NV (1996) Identification of female-produced sex pheromones and their geographical differences in pheromone gland extract composition from click beetles (Col., Elateridae). J Appl Entomol 120:463–466. https://doi.org/10.1111/j.1439-0418.1996.tb01636.x
Acknowledgements
We thank our dedicated summer students Andrew Boxwell, Hafsa Salihue, Tessa Ramburn, Amber Burnett, Victoria Piollat, Marie Tissot, Damien Rabusseau, and Crystal Chan (in 2014), and Nic Ens, Alex Hylkema, Aimee McGowan, Petra Markova, and Angela Oscienny (in 2017) for their enthusiastic and very capable assistance with all aspects of the beetle trapping work.
Funding
Open access funding provided by Agriculture & Agri-Food Canada.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors have no competing interests to declare that are relevant to the content of this article.
Additional information
Communicated by Diana la Forgia.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
van Herk, W.G., Kabaluk, J.T., Vernon, R.S. et al. Disorientation of male Agriotes click beetles in the presence of granulate pheromones - a case for mating disruption. Arthropod-Plant Interactions 17, 485–506 (2023). https://doi.org/10.1007/s11829-023-09988-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11829-023-09988-3