
Abstract
To date, all known or suspected pheromones of click beetles (Coleoptera: Elateridae) have been identified solely from species native to Europe and Asia; reports of identifications from North American species dating from the 1970s have since proven to be incorrect. While conducting bioassays of pheromones of a longhorned beetle (Coleoptera: Cerambycidae), we serendipitously discovered that males of Cardiophorus tenebrosus L. and Cardiophorus edwardsi Horn were specifically attracted to the cerambycid pheromone fuscumol acetate, (E)-6,10-dimethylundeca-5,9-dien-2-yl acetate, suggesting that this compound might also be a sex pheromone for the two Cardiophorus species. Further field bioassays and electrophysiological assays with the enantiomers of fuscumol acetate determined that males were specifically attracted by the (R)-enantiomer. However, subsequent analyses of extracts of volatiles from female C. tenebrosus and C. edwardsi showed that the females actually produced a different compound, which was identified as (3R,6E)-3,7,11-trimethyl-6,10-dodecadienoic acid methyl ester (methyl (3R,6E)-2,3-dihydrofarnesoate). In field trials, both the racemate and the (R)-enantiomer of the pheromone attracted similar numbers of male beetles, suggesting that the (S)-enantiomer was not interfering with responses to the insect-produced (R)-enantiomer. This report constitutes the first conclusive identification of sex pheromones for any North American click beetle species. Possible reasons for the strong and specific attraction of males to fuscumol acetate, which is markedly different in structure to the actual pheromone, are discussed.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Click beetles (Coleoptera: Elateridae) constitute a large family with ~10,000 described species, many of which are economically important (Barsics et al. 2013; Johnson 2002; Traugott et al. 2015). They are distributed worldwide and are some of the most abundant beetles in terrestrial habitats (Douglas 2011), including forests, grasslands, and croplands with organic matter in the soil. Elaterid larvae, called wireworms for some species, can inhabit the soil, plant litter, or dead and decaying wood (Johnson 2002; Traugott et al. 2015). Larvae can be phytophagous, predaceous, saprophagous, or omnivorous depending on species and availability of resources (Traugott et al. 2008; Vernon et al. 2009). Forest-inhabiting larvae can be saprophagous or predaceous on small invertebrates, and can be important natural enemies of many forest pests such as the pine looper, Bupalus piniarius (Dajoz 2000; Morris 1951; Turnock 1969; Zhang et al. 2008). Species which are agricultural pests are usually generalists, feeding on seedlings, roots, and harvested parts (e.g. tubers) of a variety of crops such as wheat, corn, and potatoes (Evans and Hogue 2006; Traugott et al. 2015).
The life histories of adult click beetles also differ with species and environmental variables. Most adults can fly and are able to disperse considerable distances (Schallhart et al. 2009). Species that feed as adults mainly consume rotting fruit, flowers, nectar, pollen, fungi, and sap from wounds, although some adult elaterids are predatory and will feed on honeydew-producing hemipterans and the larvae of wood-boring insects (Evans and Hogue 2006; Johnson 2002). The majority of elaterid adults are nocturnal with activity periods correlated with temperature thresholds (Crozier et al. 2003; Edwards and Evans 1950; Evans and Hogue 2006). Some elaterid species mate only once (van Herk and Vernon 2014; Vernon and van Herk 2013), whereas others can mate multiple times (Furlan 1996).
To date, only a few pheromones and pheromone candidates have been identified from the estimated 10,000 species of Elateridae. Thus, pheromones or sex attractants have been identified from approximately 40 species native to Europe and Asia (König et al. 2015; Tolasch et al. 2013; Tóth 2013). All examples to date indicate that the family uses female-produced sex pheromones (König et al. 2015; Tolasch et al. 2013; Tóth 2013; Tóth et al. 2015). Practical applications of the pheromones for monitoring click beetles have been developed for species endemic to Europe and Asia (Tóth 2013). Ivashchenko and Adamenko (1980) showed that a paired ball-like structure in the abdomen of females of the species Selatosomus latus (Fabricus) is the likely source of pheromones. The first pheromones that were identified, from Agriotes gurgistanus (Faldermann), A. litigiosus Rossi, and A. lineatus L., were found in extracts of dissected pheromone glands (Yatsynin et al. 1980). Since then, a number of pheromones have been identified from Agriotes spp. (reviewed in Tóth 2013). Approximately two-thirds of the existing elaterid pheromone data are from this genus, most likely because many Agriotes spp. are important pests in Europe, and are of increasing importance as invasive species in North America (Furlan 2005; Furlan et al. 2001; Parker and Howard 2001; Vernon et al. 2005). Sex pheromones have also been identified and developed to monitor rare or endangered species in Europe, such as Elater ferrugineus L. (Svensson et al. 2012; Tolasch et al. 2007; Zauli et al. 2014), and Idolus picipennis (Bach) (Tolasch et al. 2013). In Japan, pheromones are used for monitoring and management of species in the genus Melanotus, whose larvae are serious pests of sugarcane (Arakaki et al. 2008a, b, c; Iwanaga and Kawamura 2000; Kishita et al. 2003; Nagamine and Kinjo 1990).
To date, pheromones have not been identified from any of the estimated 965 North American elaterid species (Johnson 2002; Marske and Ivie 2003). Two putative pheromones, caproic and valeric acid, were reported for Limonius californicus (Mannerheim) and L. canus L. respectively (Butler et al. 1975; Jacobson et al. 1968), but field trials of these compounds in several US states failed to attract click beetles of any species (unpublished data: J. Serrano in California, P. Landolt and D. Horton in Washington state, G. Reddy in Montana), suggesting that these identifications were not correct. Although pheromones have not been identified, attraction of males to female beetles and/or extracts of females has been demonstrated in a few species, including Agriotes ferrugineipennis L. (Lilly and McGinnis 1965), Ctenicera destructor (Brown) (Doane 1961; Lilly and McGinnis 1965), C. sylvatica (Van Dyke) (Lilly and McGinnis 1965), Hemicrepidius decoloratus (Say) (Hayes and Wheeler 1968), H. morio L. (Chapman 1964), L. californicus (Jacobson et al. 1968; Lilly 1959; Lilly and McGinnis 1965; Lilly and McGinnis 1968; Shirck 1942;), and L. canus (Onsager et al. 1968). These data strongly suggest that females of these species are the pheromone-producing sex, in concordance with the data from European and Asian species.
While conducting field bioassays of longhorned beetle (Coleoptera: Cerambycidae) pheromones, we observed that male click beetles were sex-specifically attracted to traps baited with (E)-6,10-dimethylundeca-5,9-dien-2-yl acetate (fuscumol acetate). Fuscumol acetate is a known male-produced aggregation-sex pheromone for a number of species in the cerambycid subfamily Lamiinae (Hanks and Millar 2016). The click beetles were identified as Cardiophorus tenebrosus L. and C. edwardsi Horn (Fig. 1) based on the key by Blanchard (1889). Cardiophorus tenebrosus is reported to have a broad geographic range, encompassing much of the western United States, and with many morphological variations (Blanchard 1889). The C. tenebrosus that we collected matched the description of the California variety based on the structure of the male genitalia (Fig. 1) (Blanchard 1889). On the other hand, Blanchard (1889) reported that the range of C. edwardsi appears to be limited to California and Nevada, and it is not known to have any morphological or geographical variations.

Dorsal images of Cardiophorus tenebrosus male (a) and C. edwardsi male (b)
Little is known about the biology of these two Cardiophorus species, especially C. edwardsi. Some notes on the basic biology and life history of C. tenebrosus were documented by Stone (1957). Cardiophorus tenebrosus has been reported from alfalfa and lima bean fields in Los Angeles and Ventura counties in California (Stone 1941; Stone 1957), in association with L. californicus. Based on these reports, it is unclear whether larvae of C. tenebrosus cause damage to alfalfa, but they do cause damage to lima beans (Stone 1957). No additional reports of damage caused by C. tenebrosus have been published, and their overall impact on crops remains unknown.
There is a clear need to expand on what little is known about elaterid pheromones, especially for North American species which are reemerging as important pests in some crops due to changes in pesticide use (Traugott et al. 2015; Vernon et al. 2009). Thus, the goal of this work was to follow up on the captures of C. tenebrosus and C. edwardsi in traps baited with fuscumol acetate. Our initial objectives were: (1) to determine which enantiomer of fuscumol acetate was most attractive, and whether the other enantiomer might interfere with attraction; (2) to analyze female-emitted volatiles to determine if females produce fuscumol acetate and/or other compounds. However, once it became clear that females did not produce fuscumol acetate, we added a third objective, that is, to identify, synthesize, and field test the true sex pheromones of these two species.
Materials and Methods
Insects and Collection of Volatile Compounds
Adult beetles were collected from 9 June to 18 July 2016 and again from 21 April to 4 July 2017 at two sites in the San Bernardino National Forest in San Bernardino Co., CA, USA, near Jenks Lake (34°09′45.8”N 116°54′08.6”W and 34°09′48.1″N 116°54′13.0″W). The sites are dominated by Ponderosa pine (Pinus ponderosa Douglas) and white fir (Abies concolor [Gordon]) (Pinales: Pinaceae), with some western black oak (Quercus kelloggii Newbury), canyon live oak (Quercus chrysolepis Liebm.) (Fagales: Fagaceae), big-cone Douglas-fir (Pseudotsuga macrocarpa [Vasey]) (Pinales: Pinaceae), and incense cedar (Calocedrus decurrens Torr.) (Pinales: Cupressaceae). Adult female C. tenebrosus were collected 21 April to 18 May 2017 from the ground around the periphery of other elaterid pheromone traps, and C. edwardsi females were collected June 20–26 2017 from the ground and as random captures in flight intercept traps.
Headspace volatiles were collected from 1 to 4 live adult females for 1–7 d (Table 1) while they were held in 250 ml glass canning jars, with the metal lids fitted with an air inlet and outlet. Emitted compounds were trapped on a collector made from a glass tube (0.5 cm ID) with a 1-cm-long bed of activated charcoal (50–200 mesh; Fisher Scientific, Pittsburgh, PA, USA), held in place by glass wool plugs. Air was pulled through the jars at ~250 ml/min, and air entering the chamber was cleaned by passage through a copper pipe filled with granulated activated charcoal. The beetles were provided with a small metal screen to perch on, and sugar water for nutrition. Adsorbed compounds were eluted from the activated charcoal with 0.5 ml of dichloromethane. The aeration extracts were then stored at −20 °C until used in analyses.
It had been reported that the pheromone glands of female click beetles are located in the abdomen (Ivashchenko and Adamenko 1980; Merivee and Erm 1993), but we were unable to locate possible pheromone glands by dissections of females of our two study species. As a backup for C. tenebrosus females, following aerations, volatiles were collected from crushed abdomens of freeze-killed females, using solid phase microextraction (SPME). Thus, the abdomens of two C. tenebrosus females were placed in individual vials and crushed with a glass rod, the vials were capped with foil, and SPME fibers (100 μm PDMS; Supelco, Bellefonte, PA, USA) were inserted through the foil. Volatiles were collected onto the fibers for 2 h, then analyzed as described below.
Analyses of Extracts
Extracts of volatiles were analyzed with an HP 6890 gas chromatograph (Agilent, Santa Clara CA, USA) fitted with a DB-17 column (30 m × 0.25 mm ID × 0.25 μm film; J&W Scientific, Folsom CA, USA), coupled to an HP 5973 mass selective detector (Agilent). The temperature program used was 100 °C/1 min, then increased at 10 °C/min to 280 °C. SPME fibers were desorbed in the injector port (250 °C) in splitless mode for 30 s prior to starting the run. Compounds were tentatively identified by mass spectral interpretation and matches with database spectra (NIST 98, Agilent), and then confirmed by matching retention times and mass spectra with those of authentic standards.
An aliquot of an extract containing the C. tenebrosus female-specific compound was catalytically reduced with hydrogen and 5% palladium-carbon catalyst. After reduction, the catalyst was removed by filtration through a 5 mm bed of Celite in the tip of a Pasteur pipette plugged with glass wool. The reduced compound was then analyzed by GC-MS as described above.
The absolute configuration of the insect-produced methyl (6E)-2,3-dihydrofarnesoate was determined by base hydrolysis of the methyl ester, followed by formation of the diastereomeric amide with (S)-(−)-α-methylbenzylamine as described by Ho and Millar (2001b), with the exception that 1-ethyl-3-[(3-dimethylamino)propyl]carbodiimide hydrochloride (EDC) was used as the coupling reagent instead of dicyclohexylcarbodiimide (DCC) in the coupling step. Extracts of volatiles from C. edwardsi females were pooled to provide approximately 650 ng of methyl (6E)-2,3-dihydrofarnesoate. Samples of authentic racemic methyl (6E)-2,3-dihydrofarnesoate and the (3R)-enantiomer were similarly derivatized to provide standards. The products were analyzed by GC and GC-MS on DB-5 and DB-5MS columns (20 m × 0.32 or 0.2 mm ID, respectively, J&W Scientific) in splitless mode (250 °C) with the oven temperature programmed from 150 °C to 275 °C at 4 °C/min.
Gas Chromatography-Electroantennogram Detection (GC-EAD)
GC-EAD analyses were conducted on an HP 5890 Series II GC (Agilent) fitted with a DB-17 column (30 m × 0.25 mm ID × 0.25 μm film; J&W Scientific). A glass X-cross split the effluent between the flame-ionization and EAD detectors, with helium being added through the fourth arm of the cross at 3 ml/min as makeup gas. The column effluent directed to the EAD was diluted in a humidified air stream (650 ml/min) in a glass tube (15 mm ID) that was directed over the antennal preparation. The antennal preparation consisted of an antenna from a male beetle, which was removed by pulling it off at the scape. The distal tip of the antenna was cut off with a razor blade, and then the antenna was placed between two saline-filled (7.5 g NaCl, 0.21 g CaCl2, 0.35 g KCl, and 0.20 g NaHCO3 in 1 l Milli-Q purified water) glass capillary electrodes. The electrodes were fitted with 0.2 mm diam gold wires that were connected to a custom-built EAD amplifier. The signals from the GC and the amplifier were recorded simultaneously using Peak-Simple software (SRI International, Menlo Park CA, USA). The temperature program used for analyses was 100 °C/1 min, then at 10 °C/min to 275 °C, with an injector temperature of 250 °C. Extracts of volatiles were analyzed in splitless mode, and synthetic standards were analyzed in split mode. SPME fibers were thermally desorbed directly into the GC injector as described above.
GC-EAD was also used to test the responses of the antennae of males of C. edwardsi to the enantiomers of fuscumol acetate, by alternating injections of equal amounts of each enantiomer at 3 min intervals, using an isothermal oven temperature (175 °C) in split mode, so that a single antenna was alternately exposed to each synthetic enantiomer up to five times within a given run.
Chemicals
Racemic (5E)-6,10-dimethyl-5,9-undecadien-2-ol (fuscumol) and (E)-fuscumol acetate were purchased from Bedoukian Research (Danbury CT, USA). (R)- and (S)-fuscumol acetate were synthesized by enzymatic kinetic resolution of the racemic compound as described in Hughes et al. (2013). Racemic methyl (6E)-2,3-dihydrofarnesoate (henceforth “MDF”) and methyl (3R,6E)-2,3-dihydrofarnesoate [henceforth “(R)-MDF”] were synthesized as previously described by Ho and Millar (2001a, b). A sample of methyl 3,7,11-trimethyldodecanoate was prepared by catalytic reduction of MDF, as described above.
Field Bioassays of Fuscumol Acetate
The first field bioassays in which it was discovered that male Cardiophorus beetles were selectively attracted to fuscumol acetate were carried out at two sites approximately 500 m apart at the Jenks Lake site (see under Insect Collection), from 12 May to 24 June 2016. Traps consisted of black cross-vane panel traps (AlphaScents, West Linn OR, USA) painted with Teflon emulsion (Fluon®, Bioquip Products Inc., Rancho Dominguez CA, USA) to render trap surfaces slippery, hung on 1.5 m tall, L-shaped stands made from PVC pipe. The treatments, which were meant to target cerambycids, consisted of a blend of host plant volatiles (HPV) + ethanol as a positive control, and HPV and ethanol with racemic fuscumol, racemic fuscumol acetate, or racemic fuscumol + fuscumol acetate. The pheromones (1 ml of a 50 mg/ml solution in isopropanol) were dispensed from 5 × 7.5 cm (~0.05 mm thickness) low-density polyethylene zipper seal bags (Fisher Scientific, #01–816-1A). The HPV lure consisted of 10 ml of a synthetic conifer volatiles blend developed by Collignon et al. (2016), released from an open-topped 25 ml glass jar (4.3 cm tall × 4.3 cm outer diameter × 3.1 cm opening). Ethanol was released from 10 × 15 cm low-density polyethylene resealable bags (~0.05 mm wall thickness; Fisher Scientific) loaded with 50 ml ethanol. Traps were checked twice weekly and their order was rerandomized at every check. Pheromone lures were replaced weekly, the HPV blend was replenished twice per week, and ethanol was replenished as needed.
In a follow-up bioassay to determine which enantiomer of fuscumol acetate was attractive, the treatments initially were isopropanol solutions of (R)-fuscumol acetate (25 mg/ml), (S)-fuscumol acetate (25 mg/ml), racemic fuscumol acetate (50 mg/ml), or isopropanol as the control. Lures contained 1 ml of the lure solution in isopropanol dispensed with the zipper seal bags as described above. This experiment was conducted from June 9–20, 2016 at the Jenks Lake site, using two spatial replicates that were separated by at least 300 m, and counted twice per week. After several trap checks, it was apparent that (S)-fuscumol acetate was not attractive, and so this treatment was eliminated for the remainder of the season. The bioassay was continued from 20 June to 31 August, 2016 to determine whether there was any difference in attraction to racemic fuscumol acetate versus the (R)-enantiomer, i.e., whether the (S)-fuscumol acetate in the racemic material might interfere with attraction to the insect-produced (R)-enantiomer.
Field Bioassays of Methyl (6E)-2,3-Dihydrofarnesoate
Modified intercept traps better suited to the ecological niches of adult elaterids were designed and deployed in field bioassays conducted in 2017 (Fig. 2). In particular, trap design and placement were adjusted because most adult elaterids are ground-dwelling or inhabit fallen decaying trees (Kabalak and Sert 2011), rather than flying higher in the canopy. Trap tops (30.5 cm × 30.5 cm) and vanes (30.5 cm × 10.2 cm) were made of corrugated plastic (McMaster-Carr, Santa Fe Springs CA, USA) and three vanes were used instead of the traditional four. Trap surfaces were again coated with Fluon® to improve trapping efficiency. Tops and vanes were held together by plastic zip ties. The bottom funnel (30 cm top diameter × 5.5 cm spout diameter × 12.1 cm height) was wired to the vanes, directing beetles into the collecting jar below. A ~8 cm hole was cut into the threaded lid of the collecting jar, and the funnel spout was hot-glued to the lid. Traps were not suspended above ground as in previous bioassays. Instead, the lower half of the trap was placed into a buried piece of PVC drain pipe (20.3 cm × 16 cm OD), up to the midpoint of the funnel.

Schematic diagram of the modified intercept trap
For all experiments, traps were placed 10–15 m apart in transects, with treatments initially assigned randomly to traps. Traps were checked twice weekly, with lures replaced weekly, at which time the trap order was rerandomized. Lures consisted of low-density polyethylene zipper seal bags as described above, filled with 1 ml solutions of test compounds in isopropanol (doses listed below). Beetles were live trapped so that they could be brought back to the laboratory for sex determination, and for use in GC-EAD analyses.
The first bioassay of the likely sex pheromone of C. tenebrosus was deployed May 15–26 2017, with two spatial replicates. Treatments included racemic MDF (20 mg/ml) and fuscumol acetate (20 mg/ml), and an isopropanol control. On 26 May, the (R)-MDF treatment (4 mg/ml) was added to the field trial, the doses of racemic MDF and fuscumol acetate were correspondingly lowered to 8 mg/ml each, and bioassays were continued until 13 June. Doses were lowered because only small amounts of the (R)-enantiomer were available.
Field bioassays with C. edwardsi were conducted 9 June to 4 July 2017. Treatments consisted of racemic and (R)-MDF (8 and 4 mg respectively), racemic fuscumol acetate (8 mg), all in 1 ml of isopropanol in zipper seal bags, and an isopropanol control. All experiments were conducted at the Jenks Lake site using traps and lures as described above.
Voucher specimens of both Cardiophorus species have been deposited in the Entomology Research Museum at UC Riverside with the following voucher numbers: C. tenebrosus males UCRC ENT 501449–501451; C. tenebrosus females UCRC ENT 184083–184087; C. edwardsi males UCRC ENT 502171–502172; and C. edwardsi females UCRC ENT 502991–502995.
Statistical Analyses
Replicates for each field bioassay were based on both spatial and temporal replicates, with temporal replicates being the number of times the traps were checked. All statistical analyses were conducted with R version 3.4.2, used with RStudio version 1.1.383 (R-Development-Core-Team 2017; RStudioTeam 2016). Differences among treatment means were first assessed by the Kruskal-Wallis rank sum test, followed by Dunn’s multiple comparison test using the FSA package (Dunn 1964; Ogle 2016).
Results
Identification of Fuscumol Acetate as a Sex Attractant for C. tenebrosus and C. edwardsi
The first indication that fuscumol acetate might be a sex pheromone of C. tenebrosus and C. edwardsi was obtained during field trials of a number of cerambycid beetle pheromone components, when we noticed that large numbers of click beetles were caught only in traps baited with racemic fuscumol acetate. Further investigation determined that the trapped beetles were of two Cardiophorus species, and that all were males. In GC-EAD assays, antennae of males of both species responded strongly to racemic fuscumol acetate (data not shown). Further GC-EAD analyses showed that the antennae of male C. edwardsi responded only to (R)-fuscumol acetate (Fig. 3). Analogous trials with antennae of male C. tenebrosus were not possible because the flight period of that species had ended, as evidenced by no further catches of that species in traps.

Representative GC-EAD analysis of alternating injections of (R)- and (S)-fuscumol acetate, stimulating an antenna from a male Cardiophorus edwardsi. Top trace: GC chromatogram; bottom, inverted trace: antennal responses. Letters on tops of peaks indicate which enantiomer was injected; large peaks are from the solvent used to make sequential injections
During the period June 9–20 2016, a total of 227 C. edwardsi males were captured in traps baited with racemic and (R)-fuscumol acetate. Traps baited with (S)-fuscumol acetate did not capture any beetles during this period. Because there were no antennal responses to (S)-fuscumol acetate and no beetles had been captured in traps baited with (S)-fuscumol acetate, a second experiment was carried out to compare the attraction of racemic versus (R)-fuscumol acetate, to determine if the presence of the (S)-enantiomer was antagonistic. Traps baited with (R)- and racemic fuscumol acetate captured totals of 513 and 212 C. edwardsi males, whereas solvent-baited controls caught no beetles at all. Although traps baited with (R)-fuscumol acetate captured more than twice as many beetles as traps baited with racemic fuscumol acetate, there was no statistically significant difference between the two treatment means (Kruskal-Wallis, χ2(2) = 29.51, P < 0.05; Dunn’s test, P > 0.05) (Fig. 4). No female Cardiophorus were caught in any of the test treatments, indicating that attraction was sex-specific. In sum, these data suggested that (R)-fuscumol acetate was a possible sex pheromone of the two Cardiophorus species.

Mean (±SE) number of male Cardiophorus edwardsi caught in traps baited with (R)- or racemic fuscumol acetate 20 June – 31 August 2016. The solvent control was isopropanol. Means with the same letter are not significantly different (Dunn’s test, P > 0.05)
Identification of Sex Pheromone
In 2017, females of both C. tenebrosus and C. edwardsi were collected from field sites. Analyses of extracts of volatiles collected from live females of both species revealed a peak that elicited strong responses from antennae of conspecific male beetles in GC-EAD analyses (Fig. 5). The peak was detected in only one of five aerations of female C. tenebrosus, and in three of seven aerations of C. edwardsi (Table 1). The retention times and mass spectra of the compound from females of both species were identical, suggesting that both species utilized the same compound as their pheromone. In addition, GC-EAD analyses showed that the aeration extracts and SPME collections of volatiles from crushed female abdomens from two C. tenebrosus females contained the same single active component (data not shown).

Representative GC-EAD analyses of extracts of odors from females of two Cardiophorus species. Top trace shows the GC chromatogram and the bottom, inverted trace shows the responses from antennae of conspecific males. (a) GC-EAD of C. tenebrosus; (b) GC-EAD of C. edwardsi
The retention time and mass spectrum of the active component in the extracts were quite different from those of fuscumol acetate (Fig. 6a, b), with the retention indices of fuscumol acetate and the active component on a DB-17 column being 1725 and 1874, respectively. The mass spectrum of the female-produced compound was characterized by a probable molecular ion at m/z 252 Da, as evidenced by the loss of a methyl group to give a small ion at m/z 237 Da, for a possible molecular formula of C16H28O2, as compared to the molecular formula of fuscumol acetate of C15H26O2. After catalytic hydrogenation, the reduced product displayed a small molecular ion at m/z 256 (Fig. 6c), indicating the presence of two C=C double bonds (or possibly one alkyne) in the parent compound. The mass spectrum and retention time of the reduced compound matched those of an authentic sample of methyl 3,7,11-trimethyldodecanoate, prepared by catalytic hydrogenation of methyl (6E)-2,3-dihydrofarnesoate.

EI mass spectra (70 eV) of (a) compound emitted by female C. tenebrosus; (b) fuscumol acetate; (c) insect-produced compound after catalytic hydrogenation
This crucial finding established the carbon skeleton of the pheromone. The positions of the two double bonds, and the stereochemistry of the double bond at C6 were then confirmed by matching the retention time and mass spectrum of the insect-produced compound with an authentic standard of methyl (6E)-2,3-dihydrofarnesoate. Finally, the absolute configuration of the insect-produced MDF was determined by hydrolysis of the methyl ester to the free acid, then formation of the amide with (S)-(−)-α-methylbenzylamine. The chromatograms of the resulting amide diastereomer from a composite extract from female C. edwardsi, and of the corresponding amides prepared from synthetic racemic and (R)-MDF, demonstrated that female C. edwardsi produced exclusively the (R)-enantiomer (Fig. 7).

GC analyses of the (S)-(−)-α-methylbenzylamide derivatives of (a) synthetic racemic MDF; (b) synthetic (R)-MDF; (c) insect-produced MDF from C. edwardsi females
We then tested the responses of antennae of males to MDF and fuscumol acetate in GC-EAD analyses. Remarkably, the antennal responses to the two compounds were essentially the same (Fig. 8).

Representative GC-EAD analysis of synthetic fuscumol acetate (FuscAc) and MDF using an antenna from a male Cardiophorus tenebrosus. Top trace shows the GC chromatogram and the bottom, inverted trace shows the antennal responses
Field Bioassays of Methyl (6E)-2,3-Dihydrofarnesoate (MDF)
Field bioassays were then carried out with both racemic fuscumol acetate and racemic MDF. In the first bioassay, a total of 90 C. tenebrosus males were caught. Traps baited with MDF caught significantly more beetles (81 males) than traps baited with racemic fuscumol acetate (9 males) (Kruskal-Wallis, χ2(2) = 13.20, P < 0.05; Dunn’s test, P < 0.05). No beetles were captured in the isopropanol-baited controls.
In the second bioassay, comparing racemic MDF, (R)-MDF, and racemic fuscumol acetate, 71 C. tenebrosus and 169 C. edwardsi males were caught in total. The two MDF treatments attracted more male C. edwardsi than the fuscumol acetate (Kruskal-Wallis, χ2(3) = 19.20, P < 0.05; Dunn’s test, P < 0.05), whereas all three treatments were statistically equivalent for C. tenebrosus males (Kruskal-Wallis, χ2(3) = 8.19, P < 0.05; Dunn’s test, P > 0.05) (Fig. 9). Control traps caught only one C. edwardsi male and no C. tenebrosus males.

Mean (±1 SE) numbers of male beetles caught in traps baited with (R)-MDF, racemic MDF, or fuscumol acetate in isopropanol, 9 June – 4 July 2017. The solvent control was isopropanol. Within each species, means with the same letter are not significantly different (Dunn’s test, P > 0.05)
Discussion
Data from our preliminary field bioassays and GC-EAD analyses suggested that (R)-fuscumol acetate was a possible female-produced sex pheromone for C. tenebrosus and C. edwardsi. However, subsequent analyses of volatiles emitted by females of both species showed that C. tenebrosus and C. edwardsi females do not produce (R)-fuscumol acetate, but instead, produce a compound with a markedly different structure, methyl (6E)-2,3-dihydrofarnesoate (MDF). Thus, fuscumol acetate appears to act as an effective mimic of the actual pheromone, as evidenced by the strong attraction of male beetles to fuscumol acetate, and by the responses elicited from antennae of male beetles when challenged with fuscumol acetate. Further analyses unequivocally revealed that female C. tenebrosus and C. edwardsi produce (R)-MDF as their sex pheromone, as confirmed by both GC-EAD analyses and field bioassays. This compound had been previously identified as a male-produced sex pheromone component for two stink bug species in the genus Chlorochroa (Ho and Millar 2001b).
Furthermore, GC-EAD analyses of the enantiomers of both MDF and fuscumol showed that males of the two Cardiophorus species apparently only detect the (R)-enantiomer of each of these two compounds. This was corroborated by the bioassay data, in which the racemate or the (R)-enantiomer of each compound were equally attractive, with no apparent decrease in trap catches being attributable to the (S)-enantiomer in the racemate of either compound.
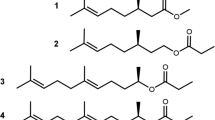
Careful comparison of the structures of (R)-fuscumol acetate and (R)-MDF revealed structural similarities which might contribute to the mimicking effect (Fig. 10). In particular, the spatial relationships between the methyl group on C2 and the carbonyl group of the acetate of (R)-fuscumol acetate, and the methyl on C3 and the carbonyl on C1 of (R)-MDF appear to be crucial, because inversion of the chiral center in each molecule completely eliminated both the EAG and the behavioral activity, despite the fact that the terpenoid hydrocarbon chain remains unchanged. This suggests that the pheromone receptor neurons are highly specific for that particular three-dimensional methyl-carbonyl structural fragment in males of both species. It remains to be seen whether other molecules containing this particular structural fragment, but different hydrocarbon chains, will also act as pheromone mimics for these two species.

Comparison of the chemical structures of (R)-fuscumol acetate and (R)-MDF
Prior to this study, all known elaterid sex pheromones had been identified from European and Asian species in the subfamilies Elaterinae and Melanotinae. Based on the structures of their pheromones, three tribes within the Elaterinae can be divided into two groups (König et al. 2015; Tolasch et al. 2007). In the tribe Elaterini, Elater ferrugineus and Sericus spp. produce esters of methyl-branched alcohols with even numbered acids (König et al. 2015; Tolasch et al. 2007), such as 7-methyloctyl octanoate, with the methyl branch on the penultimate carbon of the alcohol fragment. In contrast, in the tribe Agriotini, several species in the genus Agriotes produce esters of acyclic terpenoid alcohols (i.e. geraniol, nerol, and farnesol) coupled with even numbered carboxylic acids (König et al. 2015; Tóth 2013). Recent work from König et al. (2015) and Tolasch et al. (2013) has shown that species in the Idolus and Betarmon genera (tribe Pomachiliini) produce some of the same terpenoid esters as members of the Agriotini.
So far, pheromones have only been identified from a single genus, Melanotus, in the subfamily Melanotinae. Several species in this genus have been shown to produce esters of 12 and 14 carbon saturated and unsaturated alcohols. For example, three Melanotus species from Japan produce dodecyl acetate and related esters with terminally conjugated alcohols, (E)-9,11-dodecadien-1-ol butyrate and hexanoate (Tamaki et al. 1986; Tamaki et al. 1990; Yen and Chen 1998). Four European Melanotus species were reported to produce a variety of esters based on 14 carbon unsaturated alcohols, such as tetradecenyl butyrate (Tolasch et al. 2007; Yatsynin et al. 1996), but structure identifications were incomplete and to our knowledge, no field bioassays were ever performed to determine the actual active pheromone components.
Fuscumol acetate as a sex attractant, and MDF as a sex pheromone, do not fit exactly into any of the chemical groups discussed above. However, they are both terpenoid esters with similarities to those produced by species in the tribes Agriotini and Pomachiliini (subfamily Elaterinae). The genus Cardiophorus Esch. currently is placed in the subfamily Cardiophorinae with no tribal designation (Douglas 2017), and does not belong to any of the elaterid taxonomic groups for which pheromones had previously been identified. In addition, these are the first confirmed identifications of pheromones for any North American click beetle species. Further research on the chemical ecology of other closely related species would help to determine whether members of this subfamily represent another distinct chemotaxonomic group of pheromone producers.
It is also interesting to speculate how C. tenebrosus and C. edwardsi remain reproductively isolated, given that they are at least partially sympatric, and appear to share exactly the same sex pheromone. One such mechanism may be temporal isolation due to different seasonal activity cycles. In particular, during our field studies, although we occasionally caught both species simultaneously, it was clear that C. tenebrosus was dominant earlier in the season, and then disappeared as we continued to catch C. edwardsi. It is also possible that the two species may have somewhat different diel activity cycles. Furthermore, at close range, it is likely that species-specific contact pheromones in the cuticular lipids of the two species provide a further barrier to heterospecific mating attempts.
In summary, we have presented data showing the serendipitous but highly effective mimicry of the sex pheromone of two click beetle species, C. tenebrosus and C. edwardsi. Further work then identified the actual pheromone of both species as methyl (3R,6E)-2,3-dihydrofarnesoate. We hope that this first successful identification of sex pheromones for North American click beetle species will stimulate further work on the chemical ecology of these insects, for which a number of species are reemerging as pests of numerous agricultural crops due to changes in pest management practices.
References
Arakaki N, Nagayama A, Kobayashi A, Hokama Y, Sadoyama Y, Mogi N, Kishita M, Adaniya K, Ueda K, Higa M, Shinzato T, Kawamitsu H, Nakama S, Wakamura S, Yamamura K (2008a) Mating disruption for control of Melanotus okinawensis (Coleoptera: Elateridae) with synthetic sex pheromone. J Econ Entomol 101:1568–1574
Arakaki N, Nagayama A, Kobayashi A, Kishita M, Sadoyama Y, Mougi N, Kawamura F, Wakamura S, Yamamura K (2008b) Control of the sugarcane click beetle Melanotus okinawensis Ohira (Coleoptera: Elateridae) by mass trapping using synthetic sex pheromone on Ikei Island, Okinawa, Japan. Appl Entomol Zool 43:37–47
Arakaki N, Nagayama A, Kobayashi A, Tarora K, Kishita M, Sadoyama Y, Mougi N, Kijima K, Suzuki Y, Akino T, Yasui H, Fukaya M, Yasuda T, Wakamura S, Yamamura K (2008c) Estimation of abundance and dispersal distance of the sugarcane click beetle Melanotus sakishimensis Ohira (Coleoptera: Elateridae) on Kurima Island, Okinawa, by mark-recapture experiments. Appl Entomol Zool 43:409–419
Barsics F, Haubruge E, Verheggen J (2013) Wireworms’ management: an overview of the existing methods, with particular regards to Agriotes spp. (Coleoptera: Elateridae). Insects 4:117–152
Blanchard F (1889) Revision of the species of Cardiophorus Esch. of America north of Mexico. Trans Am Entomol Soc 16:1–27
Butler LI, McDonough LM, Onsager JA, Landis BJ (1975) Sex pheromones of the Pacific coast wireworm, Limonius canus (Coleoptera: Elateridae). Environ Entomol 4:229–230
Chapman JA (1964) Evidence for a sex attractant in the elaterid beetle, Hemicrepidius morio (LeConte). Can Entomol 96:909–910
Collignon RM, Swift IP, Zou F, McElfresh JS, Hanks LM, Millar JG (2016) The influence of host plant volatiles on the attraction of longhorn beetles to pheromones. J Chem Ecol 42:215–229
Crozier S, Tanaka A, Vernon RS (2003) Flight activity of Agriotes lineatus L. and A. obscurus L. (Coleoptera: Elateridae) in the field. J Entomol Soc British Columbia 100:91–92
Dajoz R (2000) Insects and forests. The role and diversity of insects in the forest environment. Intercept Ltd., Paris
Doane JF (1961) Movement on the soil surface of adult Ctenicera aeripennis destructor (Brown) and Hypolithus bicolor Esch. (Coleoptera: Elateridae), as indicated by funnel pitfall traps, with notes on captures of other arthropods. Can Entomol 93:636–644
Douglas H (2011) Phylogenetic relationships of Elateridae inferred from adult morphology, with special reference to the position of Cardiophorinae. Zootaxa 2900:1–45
Douglas HB (2017) World reclassification of the Cardiophorinae (Coleoptera, Elateridae), based on phylogenetic analyses of morphological characters. Zookeys 655:1–130
Dunn OJ (1964) Multiple comparisons using rank sums. Technometrics 6:241–252
Edwards EE, Evans JR (1950) Observations on the biology of Corymbites cupreus F. (Coleoptera: Elateridae). Ann Appl Biol 37:249–259
Evans AV, Hogue JN (2006) Field guide to beetles of California. University of California Press, Berkeley
Furlan L (1996) The biology of Agriotes ustulatus Schaller (Col., Elateridae). I. Adults and oviposition. J Appl Entomol 120:269–274
Furlan L (2005) An IPM approach targeted against wireworms: what has been done and what still has to be done. IOBC/wprs Bull 28:91–100
Furlan L, Tóth M, Parker WE, Ivezic M, Pancic S, Brmez M, Dobrincic R, Barcic JI, Muresan F, Subchev M, Toshova T, Molnar Z, Ditsch B, Voigt D (2001) The efficacy of the new Agriotes sex pheromone traps in detecting wireworm population levels in different European countries. Proc. XXI IWGO Conference, Venice, Oct. 27–Nov. 3, 2001, 293–303
Hanks LM, Millar JG (2016) Sex and aggregation pheromones of cerambycid beetles: basic science and practical applications. J Chem Ecol 42:631–654
Hayes JT, Wheeler AG (1968) Evidence for a sex attractant in Hemicrepidius decoloratus (Coleoptera: Elateridae). Can Entomol 100:207–208
Ho H-Y, Millar JG (2001a) Identification and synthesis of a male-produced sex pheromone from the stink bug Chlorochroa sayi. J Chem Ecol 27:1177–1201
Ho H-Y, Millar JG (2001b) Identification and synthesis of male-produced sex pheromone components of the stink bugs Chlorochroa ligata and Chlorochroa uhleri. J Chem Ecol 27:2067–2095
Hughes GP, Zou Y, Millar JG, Ginzel MD (2013) (S)-Fuscumol and (S)-fuscumol acetate produced by a male Astyleiopus variegatus (Coleoptera: Cerambycidae). Can Entomol 145:1–6
Ivashchenko II, Adamenko EA (1980) Place of pheromone formation in females of Selatosomus latus (Coleoptera: Elateridae). Zool Z 59:225–228
Iwanaga S, Kawamura F (2000) Trapping efficacy of funnel-vane and water pan traps baited with synthetic sex pheromone of the sugarcane wireworms Melanotus sakishimensis Ohira and Melanotus okinawensis Ohira (Coleoptera: Elateridae). Appl Entomol Zool 35:283–285
Jacobson M, Lilly CE, Harding C (1968) Sex attractant of sugar beet wireworm: identification and biological activity. Science 159:208–210
Johnson PJ (2002) Elateridae. In: Arnett RH, Thomas MC, Skelley PE, Frank JH (eds) American beetles, vol 2. CRC Press, Boca Raton, pp 160–173
Kabalak M, Sert O (2011) Faunistic composition, ecological properties and zoogeographical composition of the family Elateridae (Coleoptera) of the central Anatolian region of Turkey. J Insect Sci 11(57):1–36
Kishita M, Arakaki N, Kawamura F, Sadoyama Y, Yamamura K (2003) Estimation of population density and dispersal parameters of the adult sugarcane wireworm, Melanotus okinawensis Ohira (Coleoptera: Elateridae), on Ikei Island, Okinawa, by mark-recapture experiments. Appl Entomol Zool 38:233–240
König C, Szallies A, Steidle JLM, Tolasch T (2015) Sex pheromone of the rare click beetle Betarmon bisbimaculatus. J Chem Ecol 42:55–59
Lilly CE (1959) Response of males of Limonius californicus (Mann.) (Coleoptera: Elateridae) to a sex attractant separable by paper chromatography. Can Entomol 91:145–146
Lilly CE, McGinnis AJ (1965) Reactions of male click beetles in the laboratory to olfactory pheromones. Can Entomol 97:317–321
Lilly CE, McGinnis AJ (1968) Quantitative responses of males of Limonius californicus (Coleoptera: Elateridae) to female sex pheromone. Can Entomol 100:1071–1078
Marske KA, Ivie MA (2003) Beetle fauna of the United States and Canada. Coleopt Bull 57:495–503
Merivee E, Erm A (1993) Studies on sex pheromone gland morphology and pheromone components in female elaterid beetles Agriotes obscurus L. and Agriotes lineatus L. (Coleoptera: Elateridae). Proc Estonian Acad Sci Biol Ecol 42:108–117
Morris RF (1951) The larval Elateridae of eastern spruce forests and their role in the natural control of Gilpinia hercyniae Htg. Can Entomol 83:133–147
Nagamine M, Kinjo M (1990) Report on the establishment of the control technique for the sugarcane wireworm Melanotus okinawensis. Okinawa-ken Togyo Sinkou Kyokai, Naha (in Japanese)
Ogle DH (2016) FSA: fisheries stock analysis. R package version 0.8.8
Onsager JA, McDonough LM, George DA (1968) A sex pheromone in the Pacific coast wireworm. J Econ Entomol 61:691–693
Parker WE, Howard JJ (2001) The biology and management of wireworms (Agriotes spp.) on potato with particular reference to the UK. Agric For Entomol 3:85–98
R-Development-Core-Team (2017) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna
RStudio-Team (2016) RStudio: integrated development for R. RStudio, Inc., Boston
Schallhart N, Wallinger C, Juen A, Traugott M (2009) Dispersal abilities of adult click beetles in arable land revealed by analysis of carbon stable isotopes. Agric For Entomol 11:333–339
Shirck FH (1942) The flight of sugar-beet wireworm adults in southwestern Idaho. J Econ Entomol 35:423–427
Stone MW (1941) Life history of the sugar-beet wireworm in southern California. US Dep Agric Tech Bull 744:1–88
Stone MW (1957) Notes on the biology of three elaterids injurious to vegetable crops (Coleoptera). Pan-Pac Entomol 33:79–85
Svensson GP, Liedtke C, Hedenström E, Breistein P, Bång J, Larsson MC (2012) Chemical ecology and insect conservation: optimizing pheromone-based monitoring of the threatened saproxylic click beetle Elater ferrugineus. J Insect Conserv 16:549–555
Tamaki Y, Sugie M, Nagamine M, Kinjo M (1986) Female sex pheromone of the sugarcane wireworm Melanotus okinawensis Ohira (Coleoptera: Elateridae). Jpn Kokai Tokkyo Koho JP 61–12601 (in Japanese)
Tamaki Y, Sugie M, Nagamine M, Kinjo M (1990) 9,11-Dodecadienyl butyrate and 9,11-dodecadienyl hexanoate, female sex pheromone of the sugarcane wireworm Melanotus sakishimensis Ohira (Coleoptera: Elateridae). Jpn Kokai Tokkyo Koho JP 2–53753 (in Japanese)
Tolasch T, von Fragstein M, Steidle JLM (2007) Sex pheromone of Elater ferrugineus L. (Coleoptera: Elateridae). J Chem Ecol 33:2156–2166
Tolasch T, König C, von Fragstein M, Steidle JLM (2013) Identification of the sex pheromone of Idolus picipennis (Bach, 1852) revealed the presence of a cryptic sibling species. J Chem Ecol 39:1433–1440
Tóth M (2013) Pheromones and attractants of click beetles: an overview. J Pest Sci 86:3–17
Tóth M, Furlan L, Vuts J, Szarukán I, Ujváry I, Yatsynin VG, Tolasch T, Francke W (2015) Geranyl hexanoate, the female-produced pheromone of Agriotes sordidus Illiger (Coleoptera: Elateridae) and its activity on both sexes. Chemoecology 25:1–10
Traugott M, Schallhart N, Kaufmann R, Juen A (2008) The feeding ecology of elaterid larvae in central European arable land: new perspectives based on naturally occurring stable isotopes. Soil Biol Biochem 40:342–349
Traugott M, Benefer CM, Blackshaw RP, van Herk WG, Vernon RS (2015) Biology, ecology, and control of elaterid beetles in agricultural land. Annu Rev Entomol 60:313–334
Turnock WJ (1969) Predation by larval Elateridae of pupae of the pine looper Bupalus piniarius (L.) Neth J Zool 19:393–416
van Herk WG, Vernon RS (2014) Click beetles and wireworms (Coleoptera: Elateridae) of Alberta, Saskatchewan, and Manitoba. Arthropods Can Grasslands 4:87–117
Vernon RS, van Herk WG (2013) Wireworms as pests of potato. In: Giordanengo P, Vincent C, Alyokhin A (eds) Insect pests of potato: global perspectives on biology and management. Academic Press, Elsevier, Amsterdam, pp 103–164
Vernon RS, van Herk WG, Tolman J (2005) European wireworms (Agriotes spp.) in North America: distribution, damage, monitoring, and alternative integrated pest management strategies. IOBC/WPRS Bull 28:73–79
Vernon RS, van Herk WG, Clodius M, Harding C (2009) Wireworm management I: stand protection versus wireworm mortality with wheat seed treatments. J Econ Entomol 102:2126–2136
Yatsynin VG, Oleschenko IN, Rubanova EV, Ismailov VY (1980) Identification of active components of the sex pheromones of click beetles Agriotes gurgistanus, A. litigiosus, and A. lineatus. Khim Sel’sk Khoz Edition Moskova, Khimiya, pp 33–35 (in Russian)
Yatsynin VG, Rubanova EV, Okhrimenko NV (1996) Identification of female-produced sex pheromones and their geographical differences in pheromone gland extract composition from click beetles (Col. Elateridae). J Appl Entomol 120:463–466
Yen YP, Chen PH (1998) Simple and stereoselective synthesis of attractant of the sugarcane wireworm Melanotus tamsuyensis. Synth Commun 28:4561–4569
Zauli A, Chiari S, Hedenstrom E, Svensson GP, Carpaneto GM (2014) Using odour traps for population monitoring and dispersal analysis of the threatened saproxylic beetles Osmoderma eremita and Elater ferrugineus in central Italy. J Insect Conserv 18:801–813
Zhang YN, Yang ZQ, Huang HH, Fan JX (2008) Cryptalaus berus (Coleoptera: Elateridae): an important predator of Monochamus alternatus (Coleoptera: Cerambycidae), the vector of pine wilt disease in China. Chin J Biol Control 24:215–219
Acknowledgements
We thank Stacy Hishinuma and Tom Coleman for providing access to field sites, and Sean Halloran and J. Steven McElfresh for assistance with laboratory and field work. We also thank Rochelle Hoey-Chamberlain and Eric Gordon for assistance with specimen imaging. This work was funded in part by financial support from the Robert van den Bosch Scholarship to JMS, and a UC Academic Senate research grant to JGM.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Serrano, J.M., Collignon, R.M., Zou, Y. et al. Identification of Sex Pheromones and Sex Pheromone Mimics for Two North American Click Beetle Species (Coleoptera: Elateridae) in the Genus Cardiophorus Esch.. J Chem Ecol 44, 327–338 (2018). https://doi.org/10.1007/s10886-018-0940-6
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10886-018-0940-6