
Abstract
Phragmites australis exhibits substantial tolerance for salt and heavy metals exposure in the environment. The purpose of this study was to evaluate some biochemical parameters of Phragmites australis irrigated with wastewater (containing nickel, zinc, copper, iron, cadmium, and lead) up to the vegetative (short term), flowering (average term), or reproductive (long term) stage. The plant samples were collected twice i.e., two days and four days after irrigation at each growth stage. The plants were irrigated with freshwater for control. The average-term application of wastewater causes to higher biomass than the control plants. Some physicochemical parameters (proline, electrical conductivity, total soluble protein, and potassium) were more strongly correlated with plant biomass. The chlorophyll a, chlorophyll b, total chlorophyll, and total soluble proteins were reduced at the reproductive stage. While, the proline, soluble sugars, and activities of antioxidant enzymes were increased. Long-term wastewater exposure led to a significant increase in sodium, potassium, and Na+/Ca2+ ratio of P. australis while magnesium contents were decreased by wastewater irrigation. The present findings suggest that P. australis possess several enzymatic and non-enzymatic defense processes that curtail oxidative stress caused by heavy metals toxicity from wastewater and protect photosynthetic pigments from damage in the flowering stage.
Similar content being viewed by others
Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
The excessive increase in the world population, human activities, mismanagement of water resources, are major causes of water scarcity and water pollution (Qadir et al. 2010). Urmia Lake is the second-largest hypersaline lake that has suffered from severe environmental degradation due to industrial pollution and water shortage (Daraine et al. 2019). Studies have predicted that a saltwater desert (400 km2) will be generated by the desiccation of the lake which will be one of the biggest challenges affecting more than 6 million people living in the area of 50 km around Urmia Lake (Nhu et al. 2020).
The reuse of wastewater is the most commonly recommended way for the optimization of water resources, particularly in water-scarce peri-urban areas (WWAP 2017). The application of wastewater in agriculture has become a worldwide accepted source of high nutrient content, including macronutrients and micronutrients, and more importantly, organic matter (Qadir et al. 2010) to improve crop production. However, wastewater may contain significant amounts of heavy metals (HMs). High accumulation of HMs in agricultural soils not only results in contamination of soil, but also leads to elevated uptake of HMs by crops, and thus affects food quality and safety (Muchuweti et al. 2006; Belhaj et al. 2016). High concentrations of HMs prevent growth and development of plant and cause physio-chemical changes (Rezania 2009). The degree of damage depends upon the toxic ions concentration, the plant species, growth stage, and soil and climate conditions (Qadir et al. 2010). To alleviate the contamination of soils with HMs and salinization land problems, some halophytic plants have been identified that can extract HMs from the soil (Moray et al. 2015). Removal of HMs and other pollutants from wastewater with the help of aquatic plants has been reported as a cost-effect technology (Kumari and Tripathi 2015). Additionally, the use of non-food crops would be helpful to reduce the risk of the transfer of HMs from wastewater into the food chain (Lajayer et al. 2019).
Phragmites australis, a common reed, from the Poaceae family has been extensively used for the treatment of wastewater containing different types of pollutants through phytoremediation and phytostabilisation (Topal 2015; Liu et al. 2016). Its wide distribution, resilient rhizome system, and long growth period facilitate the bioaccumulation of pollutants (Liu et al. 2012; Rezania et al. 2019).
Despite the unknown mechanisms of removing heavy metals by plants, enzyme activities controlling oxidative stress might be effective in the tolerance toward HM contamination (Iannelli et al. 2002). Phragmites exhibits a high level of tolerance to HMs due to enzymatic and non-enzymatic defense systems (Uruç Parlak and Demirezen Yilmaz 2012; Esmaeilzadeh et al. 2017). As a result of altered translocation of ions, such as iron (Fe), copper (Cu), zinc (Zn), lead (Pb), and nickel (Ni), HMs cause to suppress the biological functions due to ionic imbalance (Fediuc and Erdei 2002) and bind to sulphydryl groups of enzymes (Van Assche and Clijsters 1990). Magnesium (Mg) is the most abundant free divalent cation in cells, essential for cation balance and membrane stabilization in plants, and a structural component of the chlorophyll molecule and enzymes including protein kinases, ATPases, phosphatases, and carboxylases (Rengel et al. 2015; Guo et al. 2015). Cadmium (Cd) and Pb are well-known specific blockers of voltage-dependent calcium (Ca) channels (Marchetti 2013). Disturbance of the metabolism by excessive HMs causes inhibition of plant growth and respiration, reduction of chlorophyll content, change the cell ultra-structure, and alter the activity and quantity of the enzyme (Meloni et al. 2003; Guo et al. 2007).
Cadmium is a potent inhibitor of aminolevulinic acid and photosynthetic activity (Bhattacharjee and Mukherjee 2003; Sarangthem et al. 2011). Reactive oxygen species (ROS), such as hydroxyl radicals (OH.), hydrogen peroxide (H2O2), and superoxide (O2−), are produced naturally during cell metabolism (Pereira et al. 2002) but also induce by external stresses such as exposure to pollutants and herbicides (Sharma et al. 2010), and salinity and drought stress (Khalilzadeh et al. 2016; Babaei et al. 2017). Higher ROS create an imbalance in the redox homeostasis, damaging membranes, degrading DNA, and proteins (Xu et al. 2010; Babaei et al. 2017).
Despite extensive research on individual metal tolerance to plants (Perez-Romero et al. 2016), the interactive effects of several metal stress in the system of soil–plant should be extensively studied. The mismanagement of wastewater irrigation leads to toxicity problems by accumulating HMs and declining crop quality. Therefore, the study aimed to improve our knowledge of the physiological mechanism and anti-oxidative enzyme activities of P. australis under heavy metal stress at different times and duration of wastewater irrigation. We hypothesized that the native P. australis can keep up optimal growth and physiological parameters while effectively reducing the pollution of irrigation water in urban and industrialized areas can tolerate growth.
Materials and methods
Plant material and growth conditions
The experiment was conducted at the Urmia Lake coastal, Iran (average altitude of 1735 m, extends over 6000 km2 and lies between 35° 40′ and 38° 30′ N, 44° 07′ and 47° 53′ E) (Fig. 1). The average annual rainfall is 300–700 mm. The seeds of common reed plants (Phragmites australis) were sown in plastic trays on 10 July 2019. The plants were uniformly transplanted on 24 July 2019 in the field near Uremia Lake. The characteristics of the soil of field were: EC 400 μS cm−1, pH 8.77, MgCO3 varies from 0.4 to 0.5%, and CaCO3 varies from 2.8 to 6.3%.

The map of the studied area
Phragmites plants are irrigated with wastewater or freshwater (for control). The wastewater was applied up to three stages of Phragmites australis, i.e., according to the following details:
Wastewater treatments | ||
|---|---|---|
1 | 20 days after transplanting (short term) | Vegetative growth stage |
2 | 30 days after transplanting (average term) | Flowering growth stage |
3 | 40 days after transplanting (long term) | Reproductive growth stage |
The plant samples were collected at two times, i.e., two days and four days of each growth stage. The treatments were arranged in a split-plot design with four replications. Irrigation was taken as the main factor and the factorial combinations of three irrigation stages and sampling times as sub-plot factors. Each sub-plot has 3 rows of plants and each row was 100 m long.
The source of wastewater was a mixture of domestic and industrial effluents from Urmia city; and 10 cm below the substrate surface were constructed to maintain constant water level. The impact of settling or floating depending on the media type can lead to some changes in quality parameters of wastewater over time (Cortes-esquivel et al. 2012). Therefore, the physicochemical characteristics of wastewater, shown in Table 1, were taken from the average of 4-year data from June to August.
Plant analysis
Chlorophyll and carotenoids were extracted in 80% acetone and values were obtained based on the Arnon (1949) and Khalilzadeh et al. (2020).
Catalase (CAT), peroxidase (POD), and polyphenol oxidase (PPO) enzyme activities were analyzed according to Karo and Mishra (1976). The evaluation of protein was carried out by Bradford’s (1976) method,
To measure proline, a homogeneous mixture was obtained by extracting 10 ml sulpho-acetic acid solution with 0.5 g of plant fresh tissue. Then, 2 ml glacial acetic acid and 2 ml dimenhydrinate reagent were added. 4 ml toluene was added to produce two separate-phases. The absorbance was recorded at 520 nm (Bates et al. 1973).
Soluble sugars were estimated according to Dubois et al. (1956).
Electrical conductivity was assessed according to the Jodeh et al (2015) method. Relative water content (RWC) was measured as described by Tambussi et al. (2005), and calculations are performed based on the formula:
where MF is leaf fresh mass, Md is leaf dry mass, and MT is mass after saturation.
Ratios of Na/K and Na/Ca were assessed based on the content of sodium (Na), potassium (K), and calcium ions in aboveground tissues. The Na and K were prepared based on the method of Morales et al. (2012). Calcium and Mg were determined as per the method given by Kelley et al. (1946), using atomic absorption spectrometry. To measure plant dry biomass, aerial parts of the harvested plants were dried at 80 °C for 48 h.
Statistical analysis
The significance of irrigation (I), duration of irrigation (D), time of sampling (T), and their interactions were analyzed by SAS software. The means of the data were compared using the least significant difference (LSD) test at p < 0.05.
Results
The analysis of variance (ANOVA) showed the significant interactive effect of irrigation, duration, and time on chlorophyll b, proline, relative water content, catalase, peroxidase, calcium, sodium, potassium, Na/Ca, and K/Na ratios. The chlorophyll a, total chlorophyll, carotenoids, and plant biomass were significantly influenced by the interactive effect of duration and time. While, chlorophyll a, total chlorophyll, soluble sugars, total soluble protein, electrical conductivity, polyphenol oxidase, magnesium, and plant biomass were also affected by the interaction of duration × irrigation (Tables 2 and 3).
Plant biomass
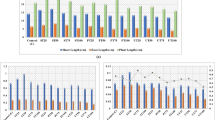
The plant biomass was significantly increased by the wastewater irrigation, duration of irrigation, and time (Table 1). The highest plant biomass (28.49 mg plant−1) was observed for the average duration of wastewater irrigation and the lowest (12.22 mg plant−1) for freshwater irrigated plants at the vegetative stage (Fig. 2a). Plant biomass was not significantly affected by short-term irrigation with wastewater at the vegetative stage, whereas, at the reproductive stage, plant biomass was significantly decreased by 13% by long-term wastewater irrigation as compared to control plants (Fig. 3a).

Concentration of plant biomass (a), chlorophyll a (b), total chlorophyll (c), soluble sugars (d), total soluble protein (e), electrical conductivity (f), polyphenol oxidase (g), and magnesium concentration (h) of Phragmites australis as affected by irrigation and duration of wastewater application. Short: 20 days after sowing (at vegetative stage); average: 30 days after sowing (at flowering stage); long: 40 days after sowing (at reproductive stage). Each bar indicates mean (n = 4) ± standard error. The different alphabets on bars indicate significant difference (P ≤ 0.05) using LSD

Concentration of plant biomass (a), chlorophyll a (b), total chlorophyll (c), and carotenoid (d) of Phragmites australis as affected by time and duration. Short: 20 days after sowing (at vegetative stage); average: 30 days after sowing (at flowering stage); long: 40 days after sowing (at reproductive stage). Each bar indicates mean (n = 4) ± standard error. The different alphabets on bars indicate significant difference (P ≤ 0.05) using LSD
Photosynthetic pigments
The changes in chlorophyll a and total chlorophyll showed the same trend for long-, short-, and average-term irrigation for control and wastewater-irrigated plants (Fig. 2b, c). The chlorophyll a content and the total chlorophyll content were even higher (but not statistically significant) for sewage-irrigated plants than the control plants at the short-term and long-term durations. The D × T interaction indicated that the maximums chlorophyll a, total chlorophyll, and carotenoids concentrations were appeared after 4 days at the flowering stage, while minimum values were observed after 4 days at the reproductive stage (Fig. 3b–d). The leaf chlorophyll b was not significantly affected by the irrigation (Table 2).
Biochemical components
Soluble sugar content was significantly higher in the wastewater-irrigated plants than the control plants. There was a 47% and 32% increase in soluble sugars by sewage irrigation at the flowering and reproductive stage, respectively when comparing to control (Fig. 2d). In comparison with the control, a significant reduction in protein content was observed after the long-term and average-term irrigation of wastewater (Fig. 2e). The proline was significantly increased in wastewater-irrigated plants at 4 days of the vegetative stage and two days of the flowering and reproductive stages as compared to control, and the highest were at the reproductive stage (Fig. 4a).

Concentration of proline (a) and relative water content (b) of Phragmites australis as affected by time and duration of wastewater application. Short: 20 days after sowing (at vegetative stage); average: 30 days after sowing (at flowering stage); long: 40 days after sowing (at reproductive stage). Each bar indicates mean (n = 4) ± standard error. The different alphabets on bars indicate significant difference (P ≤ 0.05) using LSD
Relative water content and electrical conductivity
Data showed that RWC in Salicornia was not affected by the duration of wastewater irrigation and time of sampling. Plants exposed to wastewater irrigation after 4 days at the reproductive stage result in a 5% decrease in RWC (Fig. 4b). There was no significant difference in electrical conductivity among the treatments for control plants. While plants exposed to wastewater led to an 11% increase in electrical conductivity as compared to control (Fig. 2f).
The enzymes activity
The CAT activity was unchanged for the control plant at different growth stages (Fig. 5a). A significant increase in CAT activity was detected with the average and long-term application of wastewater. The POD activity increased up to a maximum (184 µg protein min−1) after 4 days at the reproductive stage in sewage-irrigated plants. The highest PPO activity appeared at the flowering (125.5 µg protein min−1) and reproductive (119.6 µg protein min−1) stages of wastewater-irrigated plants and the lowest value (84.95 µg protein min−1) was found at the reproductive stage of control plants (Fig. 3g). In wastewater-irrigated plants, the PPO activity was 1.3- and 1.5-fold higher at the flowering and reproductive stages, respectively as compared to control.

Activities of a catalase and b peroxidase of Phragmites australis as affected by time and duration of wastewater application. Short: 20 days after sowing (at vegetative stage); average: 30 days after sowing (at flowering stage); long: 40 days after sowing (at reproductive stage). Each bar indicates mean (n = 4) ± standard error. The different alphabets on bars indicate significant difference (P ≤ 0.05) using LSD
Nutrient analysis
The magnesium (Mg) content in plants was significantly decreased when treated with the long-term duration of wastewater as compared to control plants (Fig. 2h). Whereas, no change was observed with short- and average-term exposure of wastewater as compared to control. There was no significant difference of wastewater irrigation on sodium (Na) concentration at 4 days of the flowering stage (Fig. 6a). In contrast, a significant increase in potassium (K) concentration was observed in wastewater treated plants after 4 days of the vegetative, flowering, and reproductive stages as compared to control (Fig. 6b). However, the maximum shoot Na and K concentrations were recorded after the long-term exposure of wastewater which was 51% and 26% higher than the control, respectively.

The means sodium (a) potassium (b), calcium (c), ratios of Na+/Ca2+(d) and K+/Na+ (e) of P. australis aboveground as affected by time and duration of wastewater application. Short: 20 days after sowing (at vegetative stage); average: 30 days after sowing (at flowering stage); long: 40 days after sowing (at reproductive stage). Each bar indicates mean (n = 4) ± standard error. The different alphabets on bars indicate significant difference (P ≤ 0.05) using LSD
The calcium (Ca) content of aboveground parts was not significantly affected by the wastewater; however, Ca content by average-term exposure of wastewater was higher than the long-term exposure (Fig. 6c). Furthermore, a significant increase (27%) in Ca was observed after 4 days of short-term irrigation with wastewater as compared to control. The highest Na/Ca ratio was absorbed in plants from the 4 days after the reproductive stage, and the lowest in the control plants after 4 days after the flowering stage (Fig. 6d). The Na/Ca ratio at vegetative and flowering stages was relatively lower than the long-term exposure. There was no significant difference between control and wastewater treated plants in the K/Na ratio except 4 days after the vegetative stage (Fig. 6e). The application of wastewater increased the aboveground K/Na ratio in all treatments, except 4 days after the reproductive stage.
Correlation coefficient between traits
The correlation analysis indicates that the plant biomass was significantly and positively related to EC, TSP, proline, and K contents (Fig. 7). The chlorophyll a, chlorophyll b, total chlorophyll, total soluble protein, and Ca contents were negatively related to Na/Ca ratio. The EC was positively related to the activity of CAT, POD, and PPO, and Na and K content. The Ca content was positively related to the concentration of TSP. Similarly, Mg concentration was positively related to chlorophyll b, carotenoid, and TSP.

Correlation coefficient matrix between traits. Chl a chlorophyll a, Chl b chlorophyll-b, TSP total soluble protein, EC electrical conductivity, RWC relative water content, CAT catalase, POD peroxidase, PPO polyphenol oxidase, Ca calcium, Mg magnesium, Na sodium, K potassium
Discussion
Phragmites plants subjected to different times and duration of HM polluted wastewater showed some physio-chemical changes. Based on observed results, HMs observed in irrigation treatments, are necessary for the plant growth and development at the flowering stage which cause to inhibition of plant photosynthesis which were highly accumulated at 40 days after sowing (the reproductive stage). The highest biomass was observed at the flowering stage, indicated that the optimum sewage exposure time for restoration of Phragmites should be 30 DAS. However, Phragmites contains a high amount of heavy metals accumulation and nutrient uptake from July to August, therefore, whiles the shoot biomass increase with increased development times, the Phragmites shoots weight decrease after the reproductive stage. Mishra and Behera (1991) reported that the reduction of shoot biomass in long-term exposure to wastewater may be due to the antagonistic effects of Mg and Ca with other cations (K and Na) (Najafi et al. 2012). However, there was an inverse tendency that the photosynthetic pigments and Mg content decreased significantly but the enzyme activities increased markedly, with the long-term exposure to wastewater. The lowest photosynthetic pigments under such conditions proved that long-term exposure decreased the photosynthesis rates, and then reduced the biomass production (Wen et al. 2017). Saltmarsh et al. (2006) also indicated that Phragmites was favored by average-term of duration with an increase in photosynthetic pigments which indicated that the long-term duration of wastewater was not optimal for this species.
The presence of HMs, Zn, Fe, Cu, Cd, Pb, and Ni ions exhibited a strong inhibitory effect on photosynthetic pigments and soluble protein of Phragmites. Zinc accumulation declined usually the contents of total chlorophyll, chlorophyll a, and chlorophyll b in plants (Li et al. 2013). This study suggests that chlorophyll b was more sensitive to time and duration of wastewater exposure in comparison with chlorophyll a in Phragmites plants. The plants were probably better sun-acclimatized at that time, since chlorophyll b acts as a light-harvesting complex when light conditions are not optimal (Lawlor 1987). The analysis of foliar pigments in Phragmites was consistent with average-term exposure to wastewater. During the vegetative and flowering stages, the chlorophyll content increased with increasing the HMs exposure in wastewater, mostly because metals supply in wastewater helped in the synthesis of chlorophyll. The electrical conductivity of leaves correlated negatively with RWC and Mg concentration. Therefore, declining chlorophyll could be because of the peroxidation of chloroplast membranes (Hou et al. 2007) and inhibition of other metals absorption (Wilson et al. 2000). Zhou and Qiu (2005) reported that the decrease in chlorophyll content may be due to increased Fe uptake in the presence of Cd. Iron substitutes Mg forming heme instead of chlorophyll. Increasing the chlorophyll content in plants irrigated with wastewater compared to plants irrigated with water in the reproductive growth stage suggesting that enzyme activity of CAT, POD, and PPO plays an essential role against HMs-induced oxidative damage in Phragmites plants.
Increased enzyme activity in Phragmites shows the ability to adapt under Zn (Uruç Parlak and Demirezen Yilmaz 2012), Cu, Cd (Iannelli et al. 2002; Rocha et al. 2014), and Cr contamination (Dhir et al. 2009). The higher anti-oxidative activities, such as CAT, POD, and PPO, could neutralize or eliminate free radicals (Mittler 2002; Babaei et al. 2017). Szabó et al. (2005) reported that carotenoids dissipate excitation energy by rapid internal conversion and could prevent photo-oxidation of photosynthetic membranes and destruction of the entire photosystem.
The higher proline by exposure to excess metals, such as Zn, Fe, Cu, Cd, Pb, and Ni, has been previously reported (Li et al. 2013; Uruç Parlak and Demirezen Yilmaz 2012; Khalilzadeh et al. 2020). Despite the progressive increase in proline with growth duration, the osmolyte contents under control and wastewater treatments were almost the same. The long-term exposure to HMs did not exhibit any significant effect on leaf proline content in Phragmites. Proline as an organic osmolyte has a strong ability to prevent the degeneration of enzyme inactivation, protects the structure of organelles and macromolecules (Perez-Alfocea et al. 1993; John et al. 2008). Furthermore, proline acts as a metal chelator and eliminates singlet oxygen and hydroxyl radicals, thus reduces ROS-induced cell damage (Uruç Parlak and Demirezen Yilmaz 2012). This study revealed that proline correlated better with CAT than with either POD or PPO. A previous investigation exhibited the proline accumulation in Cu (Ku et al. 2012), Ni (Lin and Kao 2007), and Zn (Li et al. 2013) treated plants.
In this study, wastewater had Ni, Zn, Cu, Fe, Cd, and Pb contents which induced the decline in soluble protein, while soluble sugar contents were significantly increased in Phragmites by long-term exposure to wastewater irrigation. A higher concentration of soluble sugar provides an adaptive mechanism under HM toxicity by maintaining favorable osmotic potential (Guo et al. 2007). The decrease in protein content after prolonged HMs exposure may be caused by oxidative damage and increased activity of CAT, POD, and PPO, which are activated by HMs. Also, the accumulation of Cd, Cu, and Pb could inhibit Mg absorption by plants on which protein synthesis system relied and induce DNA damage (John et al. 2008; Hou et al. 2007). Also, Ni- or Cu-induced proline accumulation in plant leaves could be related to protein degradation (Hou et al. 2007; Lin and Kao 2007). Our results showed the increased POD enzyme activity at the vegetative growth stage, suggesting that this enzyme might be important in alleviating lipid peroxidation induced by HM stress, as found by Guo et al. (2007).
These results showed that pollutant industrial wastewater in the environment would change the level of antioxidant enzyme activities in plants. The insufficient activities and concentration of antioxidants cause damage to pigments, nucleic acids, membrane lipids, and proteins, causing mutations, and even causing death to the plants (Xu et al. 2010). Antioxidant enzyme activities in different HMs stressed plants are highly variable, depending on the concentration of metal ions, exposure duration, and plant species (Sharma et al. 2010). Phragmites can adapt to different environments due to efficient internal mechanisms (Liu et al. 2016). Since HMs-induced cytotoxicity was exerted in most cases, after 4D of wastewater exposure, it was unknown detoxication pathways. Indeed, after 4 d of long-term exposure to wastewater, CAT and POX activities were maximum and 60% higher than the control plants. The uptake of Cd and Zn ions in the root cells occurs via the same transmembrane carriers of micronutrient metal ions (Benavides et al. 2005). Cadmium causes the deactivation of enzymes and denaturation of proteins due to its high affinity with sulfhydryl groups. Therefore, Cd is a significant threat to the plants owing to its high toxicity and mobility (Gill and Tuteja 2011). The excessive Cu induces a wide range of physicochemical changes in the plant (Påhlsson 1989) and being an oxidation–reduction (redox) metal, it is involved in the generation of free radicals in the chloroplast. Hydroxyl radicals is the most active ROS causing irreversible cell injury by modifying DNA, proteins, and lipid peroxidation. Elimination of H2O2 is therefore an essential protection mechanism for the protection of membrane integrity under HM toxicity (Van Assche and Clijsters 1990).
Peroxidase is the major H2O2-scavenging enzyme that destroys H2O2 from the cytosol and chloroplasts of plants. Catalase is most commonly used by cells to rapidly catalyze the decomposition of H2O2 (Mate´s 2000). The increased activities of CAT due to Pb, Fe, Cd, and Ni stresses have been reported in different species (Pandey and Sharma 2002; Shu et al. 2012). Induction of POD activity has been well documented under high degree toxicity of Fe, Cd, Zn, Cu, and Pb (Shu et al. 2012; Jucoski et al. 2013; Khalilzadeh et al. 2020).
Thus, the lower content of Ca in leaves from plants exposed to high HMs can be explained by a redistribution of cellular Ca for heavy metal detoxification (Jáuregui-Zúñiga et al. 2005). The role of Ca in HMs absorption and detoxification needs to be elucidated in future. In some cases, Ca alleviated the toxicity of the Ni metal ions because of the inhibition of Ni uptake by Ca (Farcasanu et al. 2018). The Cd2+ induced aquaporin luminescence observed could also be the result of Cd binding aquaporin instead of Ca because of similar physical properties. It was found that the cell exposure to high Cu-induced broad Ca waves into the cytosol which were accompanied by elevations in cytosolic Ca2+ (Ruta et al. 2016). Binding of Cd to the thiol groups of enzymes and the cell walls carbohydrates may exchange the Mg, Zn, and Fe (cofactors of enzymes with toxic effects on cell metabolism) (Das et al. 1998). In particular, Mg is necessary for the synthesis of chlorophyll and actively involved in a number of metabolic reactions as an important co-factor for enzymes involved in carbohydrate metabolism (Guo et al. 2015). Recent studies indicate that stress hormones (ethylene, abscisic acid) are involved in the signaling response to Mg in plants (Guo et al. 2015). Reasonably, high concentrations of Mg can alleviate the toxicity of metals in many plants (Kashem and Kawai 2007). Alleviation by Mg seems to be associated with physicochemical reactions at the plasma membrane (which is usually correlated with the chlorophyll a and carotenoid) and the cell wall. Magnesium enhanced protection from oxidative stress by increasing H+-ATPase activity and synthesis of organic acids (Kinraide et al. 2004; Janicka-Russak et al. 2012; Rengel et al. 2015).
The primary mechanism for the maintenance of adequate K in plant tissue under salt stress seems to be dependent upon selective K+ uptake and selective cellular K+ and Na+ compartmentation and distribution in the shoots (Carden et al. 2003). Also, the osmotic adjustment by inorganic ions is achieved to a lower energy cost than by the accumulation of organic solutes (Patakas et al. 2002). P. australis maintained a considerably higher K/Na ratio in 4 d after the vegetative stage to 2 d after the reproductive stage. However, the reduction in shoot biomass at varying times and duration of wastewater was more in the reproductive stage at the longer period of sewage irrigation. Therefore, mineral nutrients are not beneficial in large quantities at the reproductive stage of P. australis, and this plant display extremely low tolerance to NaCl in this situation. It seems that high salinity industrial wastewater in 4D after the reproductive stage decreased the Ca concentration in the plant leaves (Wei et al. 2003) and suggested that reduced Ca availability together with high Na/Ca ratios in the leaf region probably contributes to growth failure.
Conclusion
Our results show that, the tolerance of Phragmites australis to heavy metals contamination in wastewater relies on physio-chemical mechanisms. The higher levels of heavy metals through the long-term duration of wastewater irrigation caused oxidative stress in Phragmites due to the reactive oxygen species production and induced chlorophyll degradation. The higher activities of catalase, peroxidase, and polyphenol oxidase play a pivotal role in the anti-oxidative defense system of P. australis against toxicity of heavy metals. Besides, the proline contents of Phragmites were increased with increasing exposure to wastewater. The ability of Phragmites to tolerate heavy metals at the flowering stage could partly be derived from detoxifying reactive oxygen species and accumulation of soluble sugars and proline. Higher concentrations of sodium and potassium were found at the reproductive stage. Also, Phragmites produced high biomass under wastewater irrigation and biomass was significantly and positively related to proline, soluble sugars, total soluble proteins, and potassium contents.
Author contribution statement
RK performed the experiments, conducted the analysis, and wrote the manuscript; AP conceived and designed the experiments, conducted the analysis, and assisted in writing the manuscript and investigation; ES provided technical and laboratory facilities, and however co-executed the laboratory measurements; SA and SK gave the consultants about plant growth and sampling methods.
References
Arnon DI (1949) Copper enzymes in isolated chloroplast polyphenol oxidase in Beta vulgaris. Plant Physiol 24:1–15
Babaei K, Sharifi RS, Pirzad A, Khalilzadeh R (2017) Effects of bio fertilizer and nano zn-fe oxide on physiological traits, antioxidant enzymes activity and yield of wheat (Triticum aestivum L.) under salinity stress. J Plant Interact 12:381–389. https://doi.org/10.1080/17429145.2017.1371798
Bates I, Waldren R, Teare J (1973) Rapid determination of free proline for water stress studies. Plant Soil 39:205–207
Belhaj D, Elloumi N, Jerbi B, Zouari M, Abdallah FB, Ayadi H, Kallel M (2016) Effects of sewage sludge fertilizer on heavy metal accumulation and consequent responses of sunflower (Helianthus annuus). Environ Sci Pollut Res 23:20168–20177. https://doi.org/10.1007/s11356-016-7193-0
Benavides MP, Gallego SM, Tomaro ML (2005) Cadmium toxicity in plants. Braz J Plant Physiol 17:21–34. https://doi.org/10.1590/S1677-04202005000100003
Bhattacharjee S, Mukherjee AK (2003) Heavy metals alter photosynthetic pigment profiles as well as activities of Chlase and 5-aminolevulinic acid dehydratase (ALAD) in Amaranthus lividus seedlings. J Environ Biol 24:395–399
Bradford MM (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72:248–254. https://doi.org/10.1016/0003-2697(76)90527-3
Carden DE, Walker DJ, Flowers TJ, Miller AJ (2003) Single-cell measurements of the contributions of cytosolic Na+ and K+ to salt tolerance. Plant Physiol 131:676–683. https://doi.org/10.1104/pp.011445
Cortes-esquivel JA, Giácoman-vallejos G, Barceló-quintal ID, Méndez-Novelo R, Ponce-Caballero MC (2012) Heavy metals removal from swine wastewater using constructed wetlands with horizontal sub-surface flow. J Environ Prot 3:871–877. https://doi.org/10.4236/jep.2012.328102
Daraine AB, Ghasemi H, Karami F, Hatami S (2019) Urmia Lake desiccation and the signs of local climate changes. J Hydraul Struct 5:1–17. https://doi.org/10.22055/jhs.2019.30414.1116
Das P, Samantaray S, Rout GR (1998) Studies on cadmium toxicity in plants: a review. Environ Pollut 96:29–36. https://doi.org/10.1016/S0269-7491(97)00110-3
Dhir B, Sharmila P, Pardha Saradhi P, Abdul Nasim S (2009) Physiological and antioxidant responses of Salvinia natans exposed to chromium-rich wastewater. Ecotoxicol Environ Saf 72:1790–1797. https://doi.org/10.1016/j.ecoenv.2009.03.015
Dubois M, Gilles K, Hamilton I, Robers PA, Smith F (1956) Colorimetric method for determination of sugars and related substances. Anal Chem 28:350–356. https://doi.org/10.1021/ac60111a017
Esmaeilzadeh M, Karbassi A, Bastami KD (2017) Antioxidant response to metal pollution in Phragmites australis from Anzali wetland. Mar Pollut Bull 119:376–380. https://doi.org/10.1016/j.marpolbul.2017.03.030
Farcasanu IC, Popa CV, Ruta LL (2018) Calcium and cell response to heavy metals: can yeast provide an answer? Calcium Signal Transduct. https://doi.org/10.5772/intechopen.78941
Fediuc E, Erdei L (2002) Physiological and biochemical aspects of cadmium toxicity and protective mechanisms induced in Phragmites australis and Typha latifolia. J Plant Physiol 159:265–271. https://doi.org/10.1078/0176-1617-00639
Gill SS, Tuteja N (2011) Cadmium stress tolerance in crop plants: probing the role of sulfur. Plant Signal Behav 6:215–222. https://doi.org/10.4161/psb.6.2.14880
Guo TR, Zhang GP, Zhang YH (2007) Physiological changes in barley plants under combined toxicity of aluminum, copper and cadmium. Colloids Surf B Biointerfaces 57:182–188. https://doi.org/10.1016/j.colsurfb.2007.01.013
Guo W, Chen S, Hussain N, Cong Y, Liang Z, Chen K (2015) Magnesium stress signaling in plant: just a beginning. Plant Signal Behav 10:3. https://doi.org/10.4161/15592324.2014.992287
Hou W, Chen X, Song G, Wang Q, Chang CC (2007) Effects of copper and cadmium on heavy metal polluted waterbody restoration by duckweed (Lemna minor). Plant Physiol Biochem 45:62–69. https://doi.org/10.1016/j.plaphy.2006.12.005
Iannelli MA, Pietrini F, Fiore L, Petrilli L, Massacci A (2002) Antioxidant response to cadmium in Phragmites australis plants. Plant Physiol Biochem 40:977–982. https://doi.org/10.1016/S0981-9428(02)01455-9
Janicka-Russak M, Kabala K, Burzynski M (2012) Different effect of cadmium and copper on H+-ATPase activity in plasma membrane vesicles from Cucumis sativus roots. J Exp Bot 63:4133–4142. https://doi.org/10.1093/jxb/ers097
Jáuregui-Zúñiga D, Ferrer MA, Calderón AA, Muñoz R, Moreno A (2005) Heavy metal stress reduces the deposition of calcium oxalate crystals in leaves of Phaseolus vulgaris. J Plant Physiol 162:1183–1187. https://doi.org/10.1016/j.jplph.2005.03.002
Jodeh S, Alkowni R, Hamed R, Samhan S (2015) The study of electrolyte leakage from barley (Hordeum vulgare L) and Pearl millet using plant growth promotion (PGPR) and reverse osmosis. J Food Nutr Res 3:422–429. https://doi.org/10.12691/jfnr-3-7-3
John R, Ahmad P, Gadgil K, Sharma S (2008) Effect of cadmium and lead on growth, biochemical parameters and uptake in Lemna polyrrhiza L. Plant Soil Environ 54:262–270. https://doi.org/10.17221/2787-pse
Jucoski GO, Cambraia G, Ribeiro C, de Oliveira JA, de Paula SO, Oliva MA (2013) Impact of iron toxicity on oxidative metabolism in young Eugenia uniflora L. plants. Acta Physiol Plant 35:1645–1657. https://doi.org/10.1007/s11738-012-1207-4
Kashem MA, Kawai S (2007) Alleviation of cadmium phytotoxicity by magnesium in Japanese mustard spinach. Soil Sci Plant Nutr 53:246–251. https://doi.org/10.1111/j.1747-0765.2007.00129.x
Kelley OJ, Hunter AS, Sterges AJ (1946) Determination of nitrogen, phosphorus, potassium, calcium, and magnesium in plant tissue. Semimicro wet-digestion method for large numbers of samples. Ind Eng Chem Anal Ed 18(5):319–322. https://doi.org/10.1021/i560153a018
Khalilzadeh R, Seyed Sharifi R, Jalilian J (2016) Antioxidant status and physiological responses of wheat (Triticum aestivum L.) to cycocel application and bio fertilizers under water limitation condition. J Plant Interact 11:130–137. https://doi.org/10.1080/17429145.2016.1221150
Khalilzadeh R, Pirzad A, Sepehr E, Khan S, Anwar S (2020) Long-term effect of heavy metal–polluted wastewater irrigation on physiological and ecological parameters of Salicornia europaea L. J Soil Sci Plant Nutr. https://doi.org/10.1007/s42729-020-00299-7
Kinraide TB, Pedler JF, Parker DR (2004) Relative effectiveness of calcium and magnesium in the alleviation of rhizotoxicity in wheat induced by copper, zinc, aluminum, sodium, and low pH. Plant Soil 259:201–208. https://doi.org/10.1023/B:PLSO.0000020972.18777.99
Ku H-M, Tan C-W, Su YS, Chiu C-Y, Chen C-T, Jan F-J (2012) The effect of water deficit and excess copper on proline metabolism in Nicotiana benthamiana. Biol Plantarum 56:337–343. https://doi.org/10.1007/s10535-011-0098-3
Kumari M, Tripathi BD (2015) Efficiency of Phragmites australis and Typha latifolia for heavy metal removal from wastewater. Ecotoxicol Environ Saf 112:80–86. https://doi.org/10.1016/j.ecoenv.2014.10.034
Lajayer BA, Najafi N, Moghiseh E, Mosaferi M, Hadian J (2019) Micronutrient and heavy metal concentrations in basil plant cultivated on irradiated and non-irradiated sewage sludge-treated soil and evaluation of human health risk. Regul Toxicol Pharmacol 104:141–150. https://doi.org/10.1016/j.yrtph.2019.03.009
Lawlor DW (1987) Photosynthesis: metabolism control and physiology. Longman Scientific and Technical (Publ), Harlow, England, pp 31–53
Li X, Yang Y, Jia L, Chen H, Wei X (2013) Zinc-induced oxidative damage, antioxidant enzyme response and proline metabolism in roots and leaves of wheat plants. Ecotoxicol Environ Saf 89:150–157. https://doi.org/10.1016/j.ecoenv.2012.11.025
Lin YC, Kao CH (2007) Proline accumulation induced by excess nickel in detached rice leaves. Biol Plantarum 51:351–354. https://doi.org/10.1007/s10535-007-0071-3
Liu X, Huang S, Tang T, Liu X, Scholz M (2012) Growth characteristics and nutrient removal capability of plants in subsurface vertical flow constructed wetlands. Ecol Eng 44:189–198. https://doi.org/10.1016/j.ecoleng.2012.03.011
Liu L, Jie D, Liu H, Gao G, Gao Z, Li D, Li N, Qiao Z, Guo J (2016) Response of phytoliths in Phragmites australis to environmental factors in northeast China. Ecol Eng 92:119–131. https://doi.org/10.1016/j.ecoleng.2016.03.027
Marchetti C (2013) Role of calcium channels in heavy metal toxicity. ISRN Toxicol 2013:1–9. https://doi.org/10.1155/2013/184360
Matés JM (2000) Effects of antioxidant enzymes in the molecular control of reactive oxygen species toxicology. Toxicology 153:83–104. https://doi.org/10.1016/S0300-483X(00)00306-1
Meloni DA, Oliva MA, Martinez CA, Cambraia J (2003) Photosynthesis and activity of superoxide dismutase, peroxidase and glutathione reductase in cotton under salt stress. Environ Exp Bot 49(1):69–76. https://doi.org/10.1016/S0098-8472(02)00058-8
Mishra RN, Behera PK (1991) The effect of paper industry effluent on growth pigment, carbohydrate, and protein of rice seedlings. Environ Environ Pollut 72:159–168. https://doi.org/10.1016/0269-7491(91)90065-5
Mittler R (2002) Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci 7:405–410. https://doi.org/10.1016/S1360-1385(02)02312-9
Morales SG, Trejo-Téllez LI, Merino FCG, Caldana C, Espinosa-Victoria D, Cabrera BEH (2012) Growth, photosynthetic activity, and potassium and sodium concentration in rice plants under salt stress. Acta Sci Agron 34:317–324. https://doi.org/10.4025/actasciagron.34(3):13687
Moray C, Goolsby EW, Bromham L (2015) The phylogenetic association between salt tolerance and heavy metal hyperaccumulation in Angiosperms. Evol Biol. https://doi.org/10.1007/s11692-015-9355-2
Muchuweti M, Birkett JW, Chinyanga E, Zvauya R, Scrimshaw MD, Lester JN (2006) Heavy metal content of vegetables irrigated with mixtures of wastewater and sewage sludge in Zimbabwe: implications for human health. Agric Ecosyst Environ 112:41–48. https://doi.org/10.1016/j.agee.2005.04.028
Najafi N, Mardomi S, Oustan S (2012) The effect of waterlogging, sewage sludge and manure on selected macronutrients and sodium uptake in sunflower plant in a loamy sand soil. J Water Soil 26(3):619–636 ((in Persian with English abstract))
Nhu VH, Mohammadi A, Shahabi H, Shirzadi A, Al-Ansari N, Ahmad BB, Chen W, Khodadadi M, Ahmadi M, Khosravi K et al (2020) Monitoring and assessment of water level fluctuations of the lake urmia and its environmental consequences using multitemporal landsat 7 ETM+ images. Int J Environ Res Public Health 17:1–18. https://doi.org/10.3390/ijerph17124210
Påhlsson A-MB (1989) Toxicity of heavy metals (Zn, Cu, Cd, Pb) to vascular plants. Water Air Soil Poll 47:287–319. https://doi.org/10.1007/BF00279329
Pandey N, Sharma CP (2002) Effect of heavy metals Co2+, Ni2+ and Cd2+ on growth and metabolism of cabbage. Plant Sci 163:753–758. https://doi.org/10.1016/S0168-9452(02)00210-8
Patakas A, Nikolaou N, Zioziou E, Radoglou K, Noitsakis B (2002) The role of organic solute and ion accumulation in osmotic adjustment in drought-stressed grapevines. Plant Sci 163:361–367. https://doi.org/10.1016/S0168-9452(02)00140-1
Pereira GJG, Molina SMG, Lea PJ, Azevedo RA (2002) Activity of antioxidant enzymes in response to cadmium in Crotalaria juncea. Plant Soil 239(1):123–132. https://doi.org/10.3923/jbs.2009.44.50
Perez-Alfocea F, Estan MT, Santa Cruz A, Bolarin MC (1993) Effects of salinity on nitrate, total nitrogen, soluble protein and free amino acid levels in tomato plants. J Hort Sci 68:1021–1027. https://doi.org/10.1080/00221589.1993.11516443
Perez-Romero JA, Redondo-Gomez S, Mateos-Naranjo E (2016) Growth and photosynthetic limitation analysis of the Cd-accumulator Salicornia ramosissima under excessive cadmium concentrations and optimum salinity conditions. Plant Physiol Bioch 109:103–113. https://doi.org/10.1016/j.plaphy.2016.09.011
Qadir M, Wichelns D, Raschid-sally L, Drechsel P, Bahri A, Minhaset PS (2010) The challenges of wastewater irrigation in developing countries. Agric Water Manag 97:561–568. https://doi.org/10.1016/j.agwat.2008.11.004
Rengel Z, Bose J, Chen Q, Tripathi BN (2015) Magnesium alleviates plant toxicity of aluminium and heavy metals. Crop Pasture Sci 66:1298–1307. https://doi.org/10.1071/CP15284
Rezania S, Park J, Rupani PF, Darajeh N, Xu X, Shahrokhishahraki R (2019) Phytoremediation potential and control of Phragmites australis as a green phytomass: an overview. Environ Sci Pollu Res 26:7428–7441. https://doi.org/10.1007/s11356-019-04300-4
Rocha ACS, Almeida CMR, Basto MCP, Vasconcelos MTSD (2014) Antioxidant response of Phragmites australis to Cu and Cd contamination. Ecotoxicol Environ Saf 109:152–160. https://doi.org/10.1016/j.ecoenv.2014.06.027
Ruta LL, Popa CV, Nicolau I, Farcasanu IC (2016) Calcium signaling and copper toxicity in Saccharomyces cerevisiae cells. Environ Sci Pollut Res 23:24514–24526. https://doi.org/10.1007/s11356-016-6666-5
Saltmarsh A, Mauchamp A, Rambal S (2006) Contrasted effects of water limitation on leaf functions and growth of two emergent co-occurring plant species, Cladium mariscus and Phragmites australis. Aquat Bot 84:191–198. https://doi.org/10.1016/j.aquabot.2005.09.010
Sarangthem J, Jain M, Gadre R (2011) Inhibition of δ-aminolevulinic acid dehydratase activity by cadmium in excised etiolated maize leaf segments during greening. Plant Soil Environ 57:332–337. https://doi.org/10.17221/45/2011-PSE
Sharma A, Gontia I, Agarwal PK, Jha B (2010) Accumulation of heavy metals and its biochemical responses in Salicornia brachiata, an extreme halophyte. Mar Biol Res 6:37–41. https://doi.org/10.1080/17451000903434064
Shu X, Yin LY, Zhang QF, Wang W (2012) Effect of Pb toxicity on leaf growth, antioxidant enzyme activities, and photosynthesis in cuttings and seedlings of Jatropha curcas L. Environ Sci Pollut Res 19:893–902. https://doi.org/10.1007/s11356-011-0625-y
Szabó I, Bergantino E, Giacometti GM (2005) Light and oxygenic photosynthesis: energy dissipation as a protection mechanism against photo-oxidation. EMBO Rep 6(7):629–634
Tambussi EA, Nogués S, Araus JL (2005) Ear of durum wheat under water stress: water relations and photosynthetic metabolism. Planta 221:446–458. https://doi.org/10.1007/s00425-004-1455-7
Topal M (2015) Uptake of tetracycline and degradation products by Phragmites australis grown in stream carrying secondary effluent. Ecol Eng 79:80–85. https://doi.org/10.1016/j.ecoleng.2015.03.011
Uruç Parlak K, Demirezen Yilmaz D (2012) Response of antioxidant defences to Zn stress in three duckweed species. Ecotoxicol Environ Saf 85:52–58. https://doi.org/10.1016/j.ecoenv.2012.08.023
Van Assche F, Clijsters H (1990) Effects of metals on enzyme activity in plants. Plant Cell Environ 13:195–206. https://doi.org/10.1111/j.1365-3040.1990.tb01304.x
Wei W, Bilsborrow PE, Hooley P, Fincham DA, Lombi E, Forster BP (2003) Salinity induced differences in growth, ion distribution and partitioning in barley between the cultivar Maythorpe and its derived mutant Golden Promise. Plant Soil 250:183–191. https://doi.org/10.1023/A:1022832107999
Wen B, Li X, Yang F, Lu X, Li X, Yang F (2017) Growth and physiology responses of Phragmites australis to combined drought-flooding condition in inland saline-alkaline marsh, Northeast China. Ecol Eng 108:234–239. https://doi.org/10.1016/j.ecoleng.2017.08.036
Wilson C, Lesch SM, Grieve CM (2000) Growth stage modulates salinity tolerance of New Zealand spinach (Tetragonia tetragonioides, Pall.) and Red Orach (Atriplex hortensis L.). Ann Bot 85:501–509. https://doi.org/10.1006/anbo.1999.1086
WWAP (2017) The United Nations World Water Development Report 2017. Paris, France. ISBN: 978-92-3-100201-4
Xu J, Zhang J, Xie H, Li C, Bao N, Zhang C, Shi Q (2010) Physiological responses of Phragmites australis to wastewater with different chemical oxygen demands. Ecol Eng 36:1341–1347. https://doi.org/10.1016/j.ecoleng.2010.06.010
Zhou W, Qiu B (2005) Effects of cadmium hyperaccumulation on physiological characteristics of Sedum alfredii Hance (Crassulaceae). Plant Sci 169:737–745. https://doi.org/10.1016/j.plantsci.2005.05.030
Acknowledgements
This research was supported by “Iran’s National Elites Foundation”.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors have no conflicts of interest to declare.
Additional information
Communicated by S. Esposito.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Khalilzadeh, R., Pirzad, A., Sepehr, E. et al. Physiological and biochemical responses of Phragmites australis to wastewater for different time duration. Acta Physiol Plant 44, 130 (2022). https://doi.org/10.1007/s11738-022-03469-5
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11738-022-03469-5