
Abstract
Torenia fournieri, a popular potted flower crop, is frequently used as a model plant in genetic transformation studies, and transgenic plants expressing various foreign genes have been obtained in this plant. A safe and long-term conservation of the transgenic plants is necessary. In this study, shoot tips of transgenic plants of T. fournieri were successfully cryopreserved by droplet-vitrification. Growth patterns, and morphologies of leaves and flowers were identical between the plantlets recovered from cryopreservation and the in vitro stock cultures (control). No polymorphic bands and no alternations at ploidy level assessed by ISSR and FCM, respectively, were found in the transgenic plantlets recovered from cryopreservation in comparison with the in vitro stock cultures. The transgene Cry 1Ab analyzed by PCR was maintained, and its expression levels measured by qRT-PCR did not change in the cryo-derived transgenic plantlets. Therefore, cryopreservation can be considered a promising strategy for safe and long-term conservation of the transgenes.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Genetic transformation has become a complementary strategy to the conventional breeding programs. Since the first report on transgenic plants was published in the early 1980s (Zambryski et al. 1983), various transgenic plants have been obtained in a number of plant species (Teixeira da Silva 2004; Sansavini et al. 2005; Valenzuela et al. 2006; Ye et al. 2011; Cheong 2012; Chandler and Sanchez 2012). Since 1996 when the first genetically modified crops were released to commercial production, their cultivation rapidly increased, and has brought great benefits to the farmers by increasing the yield and at the same time reducing the costs (James 2010).
Obviously, continuous production of the genetically modified crops would help ensure food security required by the rapidly increasing population of the world (James 2010; Wang et al. 2014a). In the process of the transgenic plant production, once they are obtained, transgenic materials have to be maintained either in vivo (Wang et al. 2006; Mezzalama et al. 2010) or in vitro (Cao and Earle 2003; Hao and Deng 2003; Hao et al. 2005) before being analyzed, evaluated, and finally released for commercial production. In vivo maintenance of transgenic species is strictly regulated by the authorities in most countries, and may have risks of transgene loss or gene flow (Wang et al. 2006; Mezzalama et al. 2010). In vitro maintenance may also create risks of transgene loss (Menges and Murray 2004; Cho et al. 2007; Van Eck and Keen 2009) or may decrease expression levels of transgenes (Cao and Earle 2003). In addition, such maintenance needs repeated subculture, which is time-consuming and labor-cost, and may cause culture loss due to contamination or human errors (Cao and Earle 2003; Wang et al. 2014a). Therefore, availability of safe and long-term preservation techniques of transgenic materials is necessary.
Cryopreservation, which has long been considered an ideal means for long-term conservation of plant germplasm (Harding 2004; Benson 2008), has been tested for conservation of transgenes such as GUS in Oryza sativa (Meijer et al. 1991; Cornejo et al. 1995), in Pyrus pyrifolia (Gazeau et al. 1998; Hao et al. 2005), in Betula pendula (Ryynänen et al. 2002), in Citrus sinensis (Hao and Deng 2003), in Castanea sativa (Corredoira et al. 2007), and in Arabidopsis (Ogawa et al. 2008; Li et al. 2013), NPTII in O. sativa (Meijer et al. 1991), in Triticum aestivum (Fretz and Lorz 1995), and in B. pendula (Ryynänen et al. 2002), S-adenosyl-l-methionine (SAM) in Papaver somniferum (Elleuch et al. 1998), BAR in O. sativa (Cornejo et al. 1995), CYCD2;1, CYCD3;1 and ATOST1 in Arabidopsis (Menges and Murray 2004), Human cytotoxic T-lymphocyte antigen 4-immunoglobulin (hCTLA4Ig) in O. sativa (Cho et al. 2007), Papaya ringspot virus CP in Carica papaya (Tsai et al. 2009), Human serum albumin (HAS), and Escherichia coli heat labile enterotoxin (LT) protein in Nicotiana tabacum (Schmale et al. 2006; Van Eck and Keen 2009).
Cryopreservation involves not only freezing of samples in liquid nitrogen, but also in vitro culture procedures, such as maintenance of in vitro stock cultures before cryopreservation and regeneration after cryopreservation (Kulus and Zalewska 2014; Wang et al. 2014a). Furthermore, samples to be cryopreserved must be manipulated to ensure their abilities to survive in liquid nitrogen, using preculture with osmotically active compounds and exposure to cryoprotectants (Benson 2008; Kulus and Zalewska 2014). All of these steps impose osmotic or chemical stresses to the samples, which may consequently result in genetic or morphological variations of the cryo-stored material (Harding 2004; Benson 2008; Wang et al. 2014a). Therefore, genetic integrity assessments and morphological analysis in the regenerants recovered from cryopreservation are necessary. Although there have been a number of studies on the aforesaid issues in non-transgenic plants, information is still lacking on the transgenic plants.
Torenia fournieri, a popular potted flower crop, is frequently used as a model plant in genetic transformation studies, and transgenic plants expressing various foreign genes have been obtained in this plant (Aida et al. 2000; Aida 2008). The objective of the present study was, therefore, to evaluate morphologies, assess genetic integrity, and analyze foreign genes in transgenic plants of T. fournieri recovered from cryopreservation. Results obtained here would provide experimental evidence for safe and long-term conservation of genetically transformed materials.
Materials and methods
Plant materials
Torenia fournieri ‘Blue Moon’ was used in the present study. In vitro transgenic plantlets carrying Cry 1Ab gene were obtained using Agrobacterium tumefaciens-mediated transformation (Li et al. 2009). The plasmids used were pBI121 containing a reporter gene glucuronidase (GUS), a selectable marker neomycinphosphotransferase (NPTII) gene and Cry 1Ab under cauliflower mosaic virus 35S (CaMV) promoters. In vitro stock plantlets of the six transgenic lines were grown on shoot maintenance medium (SMM) composed of MS (Murashige and Skoog 1962) supplemented with 30 g l−1 sucrose and 6 g l−1 agar (Sigma Chemical Co., USA). The pH of the medium was adjusted to 5.8 prior to autoclaving at 121 °C for 20 min. The stock cultures were grown at 22 ± 2 °C under a 16-h photoperiod with a light intensity of 50 µmol s−1 m−2 provided by cool-white fluorescent tubes. Subculture was done every 8 weeks. After 6 months of in vitro maintenance of the transgenic stock cultures, shoot tips were used for cryopreservation, as described below.
Cryopreservation
Cryopreservation of shoot tips of the transgenic plantlets of T. fournieri was conducted according to Wang et al. (2014b). In brief, single nodal segments were taken from 8-week-old stock plantlets (Fig. 1a) and cultured on SMM. After 12 days of culture, axillary buds elongated to form shoots of approx. 5-mm in length. Shoot tips (2.0 mm in length) containing 5–6 leaf primordia (LPs) (Fig. 1b) were excised from the shoots and used for droplet-vitrification cryopreservation. The shoot tips were precultured on MS containing 0.5 M sucrose for 1 day. Precultured shoot tips were treated for 20 min on MS supplemented with 2 M glycerol and 0.4 M sucrose, followed by exposure to plant vitrification solution 2 (PVS2, Sakai et al. 1990) for 30 min at room temperature. PVS2 consisted of MS supplemented with 30 % (w/v) glycerol, 15 % (w/v) ethylene glycol, 15 % (w/v) dimethylsulfoxide (DMSO), and 0.4 M sucrose. Then, each shoot tip was transferred into 2.5-µl PVS2 droplets carried on aluminum foil (Fig. 1c), prior to direct immersion in liquid nitrogen for 1 h. Frozen shoot tips were rewarmed in a water bath set-up at 38 °C for 2 min, followed by unloading in MS containing 1.2 M sucrose for 20 min. Cryopreserved shoot tips were post-cultured for recovery on SMM supplemented with 0.05 mg l−1 gibberellic acid 3 (GA3) in the dark for 3 days and then transferred on the same medium under the light conditions. Shoots (>0.5 mm in length) developed after 4 weeks of post-thaw culture, and were then transferred onto the same fresh SMM for further growth. Plantlets with roots developed after 8 weeks of culture. Subculture was performed every 8 weeks. In cryopreservation experiments, at least ten shoot tips were included in each of three replicates, and the whole experiment was repeated twice.

Shoot tip cryopreservation of transgenic T. fournieri plantlets by droplet-vitrification, and recovery and morphologies of plantlets regenerated from cryopreservation. In vitro transgenic stock plantlet (a). A shoot tip used from cryopreservation (b). PVS2 droplets containing shoot tips carried on aluminum foil (c). A surviving shoot tip after 1 week of post-thaw culture (d). A shoot recovered from cryopreservation after 4 weeks of post-thaw culture (e). A plantlet recovered from cryopreservation after 8 weeks of post-thaw culture (f). Morphologies of leaf (g) and flowers (h, i) of plantlets recovered from cryopreservation (+LN) and in vitro cultures (−LN, control) after 8 weeks of post-thaw culture
Observations of recovery, growth, and morphologies
Shoot tips exhibiting green coloration after 1 week of post-thaw were considered as survival, while shoot regrowth was defined as surviving shoot tips that regenerated to shoots (≥5 mm in length) after 4 weeks of post-thaw culture, respectively. Plantlet growth pattern, and morphologies of leaves and flowers were observed in the regenerants that had been subcultured every 8 weeks for 6 months following cryopreservation.
Genetic integrity assessments
Plantlets of the six transgenic lines recovered from cryopreservation after 6 months of regeneration and the in vitro stock plantlets (control) were used for genetic integrity assessments by inter-simple sequence repeat (ISSR) and flow cytometry (FCM).
Inter-simple sequence repeat (ISSR)
Genomic DNA was extracted from 0.2 g fresh leaf tissue using a Plant Genomic DNA Kit (Tiangen, Beijing, China), according to the manufacturer’s instructions. Purified total genomic DNA was quantified and its quality was verified by ultraviolet spectrophotometry. Each sample was diluted to 50 ng l−1 in Tris–EDTA buffer and stored at −20 °C until use.
Fifty ISSR primers were screened to select suitable primers for genetic stability assessments, according to Wang et al. (2014b). PCR was performed in a 25-μl reaction solution containing 1 μl template DNA, 1 μl primer, 12.5 μl TaqMix (CoWin Biotech, Beijing, China), and 10.5 μl ddH2O. DNA amplification was performed in a PCR instrument (Eppendorf, Hamburg, Germany) using the following reaction conditions: initial denaturation step at 95 °C for 5 min, followed by 45 cycles at 94 °C for 45 s, 53 °C for 40 s, 72 °C for 70 s, and by a final extension step at 72 °C for 7 min. The PCR products were separated by electrophoresis in 1.5 % agarose gel containing 0.1 % ethidium bromide and visualized under ultraviolet light. The DNA marker DL2000 (Dongsheng, Guangzhou, P. R. China) was used to estimate the sizes of the amplified products. The experiment was repeated twice to confirm reproducibility, and only reliable and reproducible bands were included in the data analysis.
Flow cytometry
Nuclear suspensions from young leaves (0.2 g) were prepared, as described by Wang et al. (2014b), and nuclei were released from the cells by chopping samples with a razor blade in Marie’s isolation buffer (Galbraith et al. 1983). The suspension of nuclei was first filtered through a 50 μm nylon filter to remove cell fragments and large tissue debris, and then 50 μg/ml of propidium iodide (PI) (Sigma, St. Louis, Mo, USA) and 20 μg ml−1 of RNAse (Sigma, China) was added to the samples to stain the DNA. Samples were analyzed within a 15-min period in a flow cytometer. The fluorescence of samples was measured using a BD Accuri C6 flow cytometer (BD Biosciences, California, USA). Histograms were generated after analysis of at least 5000 nuclei using the software FSC 3.0 Flow Cytometry Express.
Transgene analyses
The same plantlets of the six transgenic lines recovered from cryopreservation, which were used for genetic stability assessments, were subjected to the transgene analyses by polymerase chain reaction (PCR) and real-time quantitative RT-PCR (qRT-PCR).
Polymerase chain reaction (PCR)
DNA extraction was performed as described previously (Wang et al. 2014b). Each sample was diluted to 10 ng l−1 in Tris–EDTA buffer and stored at −20 °C until use. The primer pairs for Cry1Ab (forward: 5′-GCTCTGGAACTTCTGTCG-3′ and reverse: 5′-CAGCTCATTCACCGCCTT-3′) were used for amplification of DNA products by PCR, which was performed in a 25-μl reaction solution containing 0.5 μl of each primer, 0.3 μl TaqMix (Takara, Japan), 1 μl DNA template, 2 μl dNTPs, 2.5 μl 10× PCR buffer, 1.5 μl Mg2 +, and 16.7 μl ddH2O. Conditions for PCR amplification were 94 °C for 3 min, 35 cycles each at 94 °C for 30 s, 55 °C annealing temperature for 30 s, and 72 °C for 1 min, followed by a final extension at 72 °C for 10 min. The PCR products were separated by electrophoresis in 1 % agarose gel containing ethidium bromide and visualized under ultraviolet light.
Real-time quantitative RT-PCR (qRT-PCR)
Total RNA was isolated from 0.2 g fresh leaves using TRIzol® kit (Invitrogen, China), according to manufacturer’s instructions. Ten μg of total RNA was used in reverse transcription with Moloney murine leukemia virus reverse transcriptase (Takara Biotech, China). Reverse transcription conditions were as follows: at 70 °C for 5 min, at 42 °C for 1 h, and at 95 °C for 5 min. The cDNA thereby obtained was used for real-time quantitative RT-PCR (qRT-PCR). Real-time PCR amplification used the following primer pairs: the forward (5′-GCTCTGGAACTTCTGTCG-3′) and the reverse primer (5′-CAGCTCATTCACCGCCTT-3′) of Cry1Ab, and Actin2-F (5′-GGTAACATTGTGCTCAGTGGTGG-3′) and Actin2-R (5′-AACGACCTTAATCTTCATGCTGC-3′), and was conducted in iCycler iQ5 PCR (Bio-Rad, China) in a 25-μl reaction solution containing 12.5 µl iQTM SYBR Green Supermix, 0.5 µl each primer, 1 µl cDNA, and 9.5 µl ddH2O. Thermal cycling conditions were 94 °C for 5 min, 30 cycles of 94 °C for 45 s, 55 °C annealing temperature for 45 s, 72 °C for 90 s, followed by a final extension at 72 °C for 10 min. Content of cDNA was measured by a spectrum photometer. Specific cDNA was quantified with a standard curve based on the known amounts of amplified target gene fragments. The mean value of three replicates was normalized using Actin2 as the internal control.
Results
Recovery, growth pattern, and morphology
Surviving shoot tips (Fig. 1d) showed green color and developed into shoots (Fig. 1e) after 1 and 4 weeks of post-thaw culture, respectively. Survival and shoot regrowth rates were approx. 96.7 and 66.7 %, respectively. Plantlets with well-developed roots formed after 8 weeks of post-thaw culture (Fig. 1f). Growth patterns (Fig. 1a, f), and morphologies of leaves (Fig. 1g) and flowers (Fig. 1h, i) were identical between the plantlets recovered from cryopreservation and the in vitro stock plantlets (control) after 6 months of regeneration.
Genetic integrity assessments
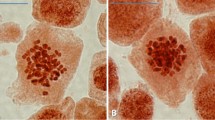
Out of the fifty ISSR primers initially screened, eight produced strong, clear, reproducible bands (Table 1), which ranged from 200 to 1500 bp in size (Fig. 2). The number of bands for each primer varied from 5 to 8, with an average of 6.4 bands produced per primer (Table 1). A total of 306 bands (number of bands obtained with eight primers × number of plantlets tested) were generated across the plantlets tested following cryopreservation. No polymorphic bands were detected across all primers and plantlets tested (Table 1; Fig. 2). Similar patterns of ploidy levels analyzed by FCM were found between in vitro stock plantlets and those recovered from cryopreservation (Fig. 3), indicating that ploidy levels were maintained in the cryopreservation-derived plantlets.

ISSR banding patterns in transgenic T. fournieri plants regenerated from cryopreserved shoot tips and in vitro shoots (control) by the primer combinations UBC811 (a) and UBC812 (b). M molecular maker; lanes 1–6 in vitro-derived plantlets (control); lanes 7–12 plantlets recovered from cryopreserved shoot tips

Ploidy level analyzed by flow cytometry (FCM) in the transgenic plantlets regenerated from cryopreservation (a) and the in vitro stock cultures (b) of T. fournieri
Transgene analyses
PCR analysis showed that a specific band of about 850 bp, which resulted from presence of Cry1Ab, was easily detected in in vitro transgenic stock cultures and plantlets recovered from cryopreservation, while such a band was never found in non-transgenic plantlets (Fig. 4). Quantitative analysis by qRT-PCR showed that there were no detected differences in expression levels of Cry1Ab between the in vitro transgenic stock cultures and plantlets recovered from cryopreservation, while such expression was not shown in in vitro non-transgenic stock cultures (Fig. 5).

PCR analysis of the Cry 1Ab transgene in the transgenic T. fournieri plantlets recovered from cryopreservation. Specific bands were detected of about 850 bp in size amplified from transgenic plantlets expressing the Cry1Ab gene. M molecular marker; lanes 1–4 in vitro non-transgenic plantlets; lanes 5–6 in vitro transgenic stock plantlets without cryopreservation; lanes 7–12 transgenic plantlets recovered from cryopreservation

Real-time quantitative RT-PCR analysis of the transgenic T. fournieri plantlets recovered from cryopreservation. Bars 1–4 in vitro non-transgenic plantlets; bars 5, 6 in vitro transgenic stock plantlets without cryopreservation; bars 7–12 transgenic plantlets recovered from cryopreservation. Bar diagram represents the expression level of the Cry 1Ab gene. Results are presented as mean ± SE
Discussion
Transgenic plants expressing various genes have been obtained in a great number of vegetatively propagated plant species, including fruit trees (Sansavini et al. 2005; Cheong 2012), ornamentals (Teixeira da Silva 2004; Chandler and Sanchez 2012), and forest species (Valenzuela et al. 2006; Ye et al. 2011). These transgenic plants can be used either directly or for further genetic improvements. Shoot tips are preferred for germplasm preservation over cells and callus, because they maintain unique strains of propagated materials and are identical to the mother plants (Kulus and Zalewska 2014; Wang et al. 2014a).
Although there have been a number of studies on cryopreservation of transgenic plants, only a few used shoot tips (Wang et al. 2012a, b). Tsai et al. (2009) successfully cryopreserved shoot tips of the transgenic papaya (Carica papaya L.) expressing coat protein gene of Papaya ringspot virus. However, they did not analyze the presence and expression of the transferred gene in the regenerants recovered from cryopreservation. Ryynänen et al. (2002) reported that shoots recovered from cryopreservation had the same copy number of the NPTII gene as the non-cryopreserved control, and the NPTII transcript levels were generally not altered by cryopreservation in the transgenic plants of B. pendula. Hao et al. (2005) found that the GUS gene assayed by PCR and Southern blotting was stably maintained, and no single base variations of the GUS gene were detected by conformation polymorphism assay in the transgenic plants recovered from cryopreservation. The studies of Ryynänen et al. (2002) and Hao et al. (2005) used transgenic plants containing either reporter gene (GUS) or selectable marker gene (NPTII). More recently, Li et al. (2013) found that the expression of the ATOST1 gene was up-regulated about 2.4-fold in the transgenic Arabidopsis plantlets recovered from cryopreserved seedling tissues, compared with non-cryopreserved controls. These data indicated that cryopreservation could increase the expression of foreign gene.
Cry 1Ab, an insecticidal crystal protein gene of Bt, encodes gene expression resistant to lepidopteran insects by selectively damaging their midgut lining (Perlak et al. 1991). Regulatory approvals for environmental release of transgenic plants expressing Cry 1Ab have been issued in several countries and the European Union (http://creativecommons.org/licenses/by-nc-nd/3.0/us/). In this study, we for the first time reported that Cry1Ab was maintained and its expression levels were not altered in the transgenic plants of T. fournieri recovered from cryopreservation. These data were consistent with the previous studies of Ryynänen et al. (2002) and Hao et al. (2005).
Morphology and genetic integrity are the most important concerns on conservation of plant germplasm including not only the wild species and cultivated crops but also the transgenic materials. Morphology evaluations and genetic integrity assessments have been extensively studied in non-transgenic plants recovered from cryopreservation (Harding 2004; Benson 2008; Kulus and Zalewska 2014; Wang et al. 2014a). The update information can be found in the two latest review papers of Kulus and Zalewska (2014), and Wang et al. (2014a). However, morphological evaluations and genetic integrity assessments have been lacking in the transgenic plants recovered from cryopreservation. The present study evaluated morphologies and assessed genetic integrity in the cryopreserved transgenic plants of T. fournieri, and found that plant growth patterns, and morphologies of leaves and flowers were identical between the transgenic plants recovered from cryopreservation and the in vitro stock cultures (control). Similar results have been obtained in non-transgenic ornamental crops by Ishikawa et al. (1997) for Bletilla striata, Hornung et al. (2001) for Primula, Suzuki et al. (2005) for Gentiana, Hitmi et al. (2000) and Wang et al. (2014b) for Chrysanthemum, Sopalun et al. (2010) for Grammatophyllum speciosum, and Zhang et al. (2015) for Argyranthemum. Furthermore, in this study, no polymorphic bands and no alternations at ploidy levels assessed by ISSR and FCM, respectively, were observed between the cryo- and in vitro-derived (control) plants, indicating that genetic stability was maintained in the transgenic plantlets following cryopreservation. These results agreed with the previous studies on cryopreservation of other non-transgenic ornamental species such as Vanda pumila (Na and Kondo 1996), Grammatophyllum speciosum (Sopalun et al. 2010), Chrysanthemum (Wang et al. 2014b), and Argyranthemum (Zhang et al. 2015).
In conclusion, the present study demonstrates that there are no detected differences in morphologies of leaves and flowers of the transgenic plantlets of T. fournieri recovered from cryopreservation and the in vitro stock cultures (control). Genetic stability assessed by ISSR and CFM was maintained in cryo-derived transgenic plants. These results indicate that cryopreservation can retain the transgene Cry 1Ab and does not alter its expression levels. Thus, cryopreservation can be considered a reliable tool for long-term and safe conservation of transgenes.
Author contribution statement
Qiaochun Wang, the Chief scientist of the present study, is responsible for the financial supports, experimental design, and preparation of the manuscript; Jingwei Li and Houhua Li for the performance of the experiments, data collection and analysis, and preparation of manuscript; and Ren-Rui Wang and Xiaoxia Gao for the assistance of experimental materials; chemical and solution preparation.
References
Aida R (2008) Torenia fournieri (torenia) as a model plant for transgenic studies. Plant Biotechnol 25:541–545
Aida R, Kishimotoa S, Tanaka Y, Shibata M (2000) Modification of flower color in torenia (Torenia fournieri Lind.) by genetic transformation. Plant Sci 153:33–42
Benson EE (2008) Cryopreservation of phytodiversity: a critical appraisal of theory & practice. Crit Rev Plant Sci 27:141–219
Cao J, Earle ED (2003) Transgene expression in broccoli (Brassica oleracea var. italica) clones propagated in vitro via leaf explants. Plant Cell Rep 21:789–796
Chandler SF, Sanchez C (2012) Genetic modification; the development of transgenic ornamental plant varieties. Plant Biotechnol J 10:891–903
Cheong EJ (2012) Biotechnological approaches for improvement and conservation of Prunus species. Plant Biotechnnol Rep 6:17–28
Cho JS, Hong SM, Joo SY, Yoo JS, Kim DI (2007) Cryopreservation of transgenic rice suspension cells producing recombinant hCTLA4Ig. Appl Microbiol Biotechnol 73:1470–1476
Cornejo MJ, Wong VL, Blechl AE (1995) Cryopreserved callus: a source of protoplasts for rice transformation. Plant Cell Rep 14:210–214
Corredoira E, San-José MC, Vieitez AM, Ballester A (2007) Improving genetic transformation of European chestnut and cryopreservation of transgenic lines. Plant Cell Tissue Org Cult 91:281–288
Elleuch H, Gazeau C, David H, David A (1998) Cryopreservation does not affect the expression of a foreign sam gene in transgenic Papaver somniferum cells. Plant Cell Rep 18:94–98
Fretz A, Lorz H (1995) Cryopreservation of in vitro cultures of barley (Hordeum vulgare L. and H. murinum L.) and transgenic cells of wheat (Triticum aestivum L.). J Plant Physiol 146:489–496
Galbraith DW, Harkins KR, Maddox JM, Ayres NM, Sharma DP, Firoozabady E (1983) Rapid flow cytometric analysis of the cell cycle in intact plant tissues. Science 220:1049–1051
Gazeau C, Elleuch H, David A, Morisset C (1998) Cryopreservation of transformed Papaver somniferum cells. CryoLett 19:147–158
Hao YJ, Deng XX (2003) GUS gene remains stable in transgenic Citrus callus recovered from cryopreservation. CryoLett 24:375–380
Hao YJ, Cheng YJ, Deng XX (2005) Stable maintenance and expression of a foreign gene in transgenic pear shoots retrieved from in vitro conservation. J Plant Physiol 162:237–243
Harding K (2004) Genetic integrity of cryopreserved plant cells: a review. CryoLett 25:3–22
Hitmi A, Barthomeuf Ch, Sallanon H (2000) Cryopreservation of Chrysanthemum cinerariaefolium shoot tips. J Plant Physiol 156:408–412
Hornung R, Holland A, Taylor HF, Lynch PT (2001) Cryopreservation of auricula shoot tips using the encapsulation/dehydration technique. CryoLett 22:27–34
Ishikawa K, Harata K, Mii M, Sakai A, Yoshimatsu K, Shimomura K (1997) Cryopreservation of zygotic embryos of a Japanese terrestrial orchid (Bletilla striata) by vitrification. Plant Cell Rep 16:754–757
James C (2010) A global overview of biotech (GM) crops: adoption, impact and future prospects GM Crops. GM Crop 1:8–12
Kulus D, Zalewska M (2014) Cryopreservation as a tool used in long-term storage of ornamental species—a review. Sci Hortic 168:88–107
Li HH, Zhao B, Seitz C, Forkmann G (2009) In vitro plant regeneration and Agrobacterium-mediated genetic transformation of Torenia fournieri. In: Zhang QX (ed) Advances in ornamental horticulture. Forest Press of China, Beijing, pp 183–187
Li ZA, Du YQ, Wang ZC (2013) Effect of cryopreservation on the efficiency of exogenous gene, genetic transformation and expression level of Arabidopsis thaliana. Electron J Biotechnol 16:6. doi:10.2225/vol16-issue6-fulltext-12
Meijer EGM, Iren E, Schrijnemakers E, Hensgens LAM, van Zijderveld M, Schilperoort RA (1991) Retention of the capacity to produce plants from protoplasts in cryopreserved cell lines of rice (Oryza sativa L). Plant Cell Rep 10:171–174
Menges M, Murray AH (2004) Cryopreservation of transformed and wild-type Arabidopsis and tobacco cell suspension cultures. Plant J 37:635–644
Mezzalama M, Crouch JH, Ortiz R (2010) Monitoring the threat of unintentional transgene flow into maize gene banks and breeding materials. Electron J Biotechnol 13:2. doi:10.2225/vol13-issue2-fulltext-5. http://www.ejbiotechnology.info/content/vol13/issue2/full/5/
Murashige T, Skoog F (1962) A revised medium for rapid growth and bioassays with tobacco cell cultures. Physiol Plant 15:473–497
Na HY, Kondo K (1996) Cryopreservation of tissue-cultured shoot primordia from shoot apices of cultured protocorms in Vanda pumila following ABA preculture and desiccation. Plant Sci 118:195–201
Ogawa Y, Suzuki H, Sakurai N, Aoki K, Saito K, Shibata D (2008) Cryopreservation and metabolic profiling analysis of Arabidopsis T87 suspension-cultured cells. CryoLett 29:427–436
Perlak FJ, Fuchs RL, Dean DA, Mcpherson SL, Fischhoff DA (1991) Modification of the coding sequence enhances plant expression of insect control protein genes (Bacillus thuringiensis/insect-resistant plants/synthetic genes). Proc Nat Acad Sci 88:3324–3328
Ryynänen L, Sillanpää M, Kontunen-Soppela S, Tiimonen H, Kangasjärvi J, Vapaavuori E (2002) Preservation of transgenic silver birch (Betula pendula Roth) lines by means of cryopreservation. Mol Breed 10:143–152
Sakai A, Kobayashi S, Oiyama I (1990) Cryopreservation of nucellar cells of navel orange (Citrus sinensis Osb. var. brasiliensis Tanaka) by vitrification. Plant Cell Rep 9:30–33
Sansavini S, Belfanti E, Costa F, Donati F (2005) European apple breeding programs turn to biotechnology. Chron Hortic 45:16–19
Schmale K, Rademacher TH, Fischer R, Hellwig S (2006) Towards industrial usefulness—cryocell-banking of transgenic BY-2 cell cultures. J Biotechnol 124:302–311
Sopalun K, Kanchit K, Ishikawa K (2010) Vitrification-based cryopreservation of Grammatophyllum speciosum protocorm. CryoLett 31:347–357
Suzuki M, Akihama T, Ishikawa M (2005) Cryopreservation of encapsulated gentian axillary buds following 2 step-preculture with sucrose and desiccation. Plant Cell Tissue Org Cult 83:115–121
Teixeira da Silva JA (2004) Ornamental chrysanthemums: improvement by biotechnology. Plant Cell Tissue Org Cult 79:1–18
Tsai SF, Yeh SD, Chan CF, Liaw SL (2009) High-efficiency vitrification protocols for cryopreservation of in vitro grown shoot tips of transgenic papaya lines. Plant Cell Tissue Org Cult 98:157–164
Valenzuela S, Balocchi C, Rodríguez J (2006) Transgenic trees and forestry biosafety. Electron J Biotechnol. http://www.ejbiotechnology.info/content/vol9/issue3/full/22/
Van Eck J, Keen P (2009) Continued expression of plant-made vaccines following long-term cryopreservation of antigen-expressing tobacco cell cultures. In Vitro Cell Dev Biol Plant 45:750–757
Wang F, Yuan QH, Shi L, Qian Q, Liu WG, Kuang BG, Zeng DL, Liao YL, Cao B, Jia SR (2006) A large-scale field study of transgene flow from cultivated rice (Oryza sativa) to common wild rice (O. rufipogon) and barnyard grass (Echinochloa crusgalli). Plant Biotechnol J 4:667–676
Wang B, Zhang ZB, Yin ZF, Feng CH, Wang QC (2012a) Novel and potential application of cryopreservation to plant genetic transformation. Biotechnol Adv 30:604–612
Wang QC, Wang RR, Li BQ, Cui ZH (2012b) Cryopreservation: a strategy for safe preservation of genetically transformed plant materials. Adv Genet Eng Biotechnol 1:1–2
Wang B, Wang RR, Cui ZH, Li JW, Bi WL, Li BQ, Ozudogru EA, Volk GM, Wang QC (2014a) Potential applications of cryobiotechnology to plant genetic transformation and pathogen eradication. Biotechnol Adv 32:583–595
Wang RR, Gao XX, Chen L, Huo LQ, Li MF, Wang QC (2014b) Shoot recovery and genetic integrity of Chrysanthemum morifolium shoot tips following cryopreservation by droplet-vitrification. Sci Hortic 176:330–339
Ye X, Busov V, Zhao N, Meilan R, Lisa M, McDonnell LM, Heather D, Coleman HD, Shawn D, Mansfield SD, Feng CF, Li Y, Cheng ZM (2011) Transgenic populus trees for forest products, bioenergy, and functional genomics. Cri Rev Plant Sci 30:415–434
Zambryski P, Joos H, Genetello C, Leemans J, Montagu MV, Schell J (1983) Ti-plasmid vector for the introduction of DNA into plant cells without alteration of their normal regeneration capacity. EMBO J 2:2143–2150
Zhang Z, Skjeseth G, Elameen A, Haugslien S, Sivertsen A, Wang QC, Blystad DR (2015) Field performance evaluation and genetic integrity assessment in Argyranthemum maderense plants recovered from cryopreserved shoot tips. In Vitro Cell Dev Biol Plant 51:505–513
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by M. Lambardi.
J.-W. Li and H.-H. Li have contributed equally to this work.
Rights and permissions
About this article

Cite this article
Li, JW., Li, HH., Wang, RR. et al. Cryopreservation for retaining morphology, genetic integrity, and foreign genes in transgenic plants of Torenia fournieri . Acta Physiol Plant 38, 8 (2016). https://doi.org/10.1007/s11738-015-2028-z
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11738-015-2028-z