
Abstract
This study evaluated the nephroprotective effect of kaempferol against cadmium chloride (CdCl2) -induced nephropathy in rats. It also investigated if activation of Nrf2 is a common mechanism of action. Adult male rats ((150 ± 15 g) were divided into 4 groups (n = 8/each) as a control (1% DMSO, orally), control + kaempferol (200 mg/kg, orally), CdCl2 (50 mg/l in drinking water), and CdCl2 + kaempferol (200 mg/kg)-treated rats. All treatments were conducted for 8 weeks. Kaempferol significantly attenuated CdCl2-induced weight loss, reduction in kidney weights, and the injury in the glomeruli, proximal tubules, and distal tubules in the treated rats. It also significantly lowered serum levels of urea and creatinine, increased urine output and urinary creatinine levels and clearance but reduced urinary levels of albumin urinary albumin exertion (UAER), and urinary albumin/creatinine ratio (UACR) in these rats. In parallel, kaempferol downregulated renal levels of cleaved caspase-3 and Bax and unregulated those of Bcl2. In the kidney tissues of the control animals and CdCl2 rats, kaempferol significantly attenuated oxidative stress, inflammation and significantly boosted levels of manganese superoxide dismutase and glutathione. Also, and in both groups, kaempferol suppressed the nuclear levels of NF-κB p65, downregulated Keap1, and stimulated the nuclear activation and protein levels of Nrf2. In conclusion, kaempferol is a potential therapeutic drug to prevent CdCl2-induced nephropathy due to its anti-inflammatory and anti-oxidant effects mediated by suppressing NF- NF-κB p65 and transactivating Nrf2.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Chronic kidney disease (CKD), is a medical condition that is characterized by impaired kidney function, reduced estimated glomerular filtration rate (eGFR), albuminuria, and glomerular hypertension (Afkarian et al. 2016). The recently available updates indicate a total global CKD prevalence of about 11–13% (Alsuwaida et al. 2010; Hill et al. 2016). However, the molecular mechanisms underlying CKD are complex, but include, at least, oxidative stress activation, inflammation, intestinal fibrosis, and apoptosis (Gajjala et al. 2015). Although diabetes mellitus (DM), smoking, hypertension, and cardiovascular disorders (CVDs) are major risk factors for the development of CKD, it is currently well-established that exposure to heavy metals is an independent risk factor (Gajjala et al. 2015).
Cadmium (Cd2+) ion is the most known heavy metal that is widely distributed in the soil, air, smoking, and industrial products (e.g., pigment and batteries) (El-Kott et al. 2020b, Prozialeck and Edwards 2012, Satarug 2018). As an environmental contaminant, experimental and human studies have shown that exposure to Cd2+ increases the risk for the development of nephropathies and CKD by promoting oxidative stress, inflammation, interstitial fibrosis, and apoptosis (Diamond et al. 2019; Jiao et al. 2019; Pavón et al. 2019). However, overproduction of reactive oxygen species (ROS), depletion of antioxidants, and suppression of nuclear factor erythroid 2 related factor-2 (Nrf2) antioxidant pathway are the best-described mechanisms underlying the pro-oxidant potential of Cd2 + , which is assumed to be the major mechanism behind its effect of all renal damaging pathways (Diamond et al. 2019, Hagar and Al Malki 2014, Jiao et al. 2019, Pavón et al. 2019, Satarug 2018, Wang et al. 2013, Yuan et al. 2016).
Nonetheless, oxidative stress is identified by the overproduction of reactive oxygen species (ROS) as compared to elimination (Newsholme et al. 2016). In the majority of the cells, the major source of ROS is the mitochondria (Zorov et al. 2014). However, the generation of ROS can be increased under several stressful conditions including exposure to radiation, toxic drugs, injury, ischemia, hypoxia, etc. (Zorov et al. 2014). On the other hand, animal cells are well-equipped with enzymatic and non-enzymatic antioxidant systems that are distributed in the cytoplasm and organelles to fight oxidative stress. These include glutathione (GSH), thioredoxin (TRx), heme-oxygenase-1 (HO-1), superoxide dismutase (SOD), catalase (CAT), glutathione peroxidase (GPx), and glutathione reductase (GRx) (Birben et al. 2012). The effect of ROS in the cell is well-established and includes activation of inflammation and apoptosis. Indeed, ROS can promote cell inflammation by activating the NRLP3 inflammasome and the nuclear factor kappa-beta (NF-κB) which can translocate to the mitochondria to stimulate the transcription of numerous inflammatory cytokines and mediators. In addition, NF-κB is positively crossed talk with ROS that induced persistent oxidative stress and inflammatory status within the cells (Morgan and Liu 2011). In addition, ROS can induce intrinsic (mitochondria-mediated cell death) by damaging the DNA and upregulation of p53 and Bax signaling (Redza-Dutordoir and Averill-Bates 2016).
On the other hand, Nrf2 is a ubiquitous transcription factor that plays a significant role in the majority of the cells through stimulating cell survival and inhibiting cell inflammation and apoptosis (Bellezza et al. 2018). In the cytoplasm, Nrf2 is always bound to keap-1 protein (Li et al. 2018). Under normal conditions, keap-1 stimulates the ubiquitination and proteasome degradation Nrf2. However, under stress, ROS and electrophiles can phosphorylate keap-1, thus inhibiting its association with Nrf2 (Li et al. 2018). This promotes the nuclear translocation of Nrf2 to initiate the transcription process. In this regard, Nrf2 is the best-known antioxidant transcription factor that stimulates the synthesis of GSH and other phases II antioxidant enzymes (e.g., HO-1, SOD, CAT) (Li et al. 2018; Bellezza et al. 2018). Also, Nrf2 is a potent anti-inflammatory molecule that can suppress NF-κB (Alshehri et al. 2021; Li et al. 2008). Also, Nrf2 inhibits cell apoptosis by direct upregulation of the anti-apoptotic protein Bcl2 (Niture and Jaiswal 2012). Of note, keap-1/Nrf2 signaling is extremely inhibited in the majority of renal disorders including those induced by Cd2+ whereas the activation of this pathway was protective (Alshehri et al. 2022; Liu et al. 2019; Nezu et al. 2017).
Currently, no definite treatment is available to treat CKD. Therefore, international efforts in experimental science and clinical medicine are carried out during the last decades to develop suitable and safe therapies to treat CKD. In this view, much interest in drug discovery to treat CKD from plant flavonoids is rapidly increasing and well-reported (Vargas et al. 2018). Kaempferol is a plant flavonoid that is abundantly found in vegetables and plants such as tea, tomato, grapes, broccoli, and beans (Devi et al. 2015). Kaempferol attenuated liver, lung, kidney, brain, and heart damage in a variety of animal models by silencing oxidative stress and inflammation. It also attenuated renal oxidative, inflammatory, and fibrotic damage in diabetic, aged, and cisplatin and tacrolimus-treated rodents (Al-Numair et al. 2015; Ali et al. 2020; Park et al. 2009; Sharma et al. 2020; Wang et al. 2020; Zhang et al. 2019). Similar protective effects of kaempferol were also reported in D-ribose-induced mesangial cell apoptosis (Zhang et al. 2019). In all these studies, the nephroprotective effects of kaempferol were mediated by several mechanisms, including scavenging of ROS, activation of Nrf2/antioxidants axis, inhibiting the nuclear factor kappa-beta (NF-κB), and modulating of the mitogen-activated protein kinase (MAPK) apoptotic members (i.e., such as p38, ERK, and JNK).
Despite these studies, the renoprotective effect of kaempferol against Cd2+ ions-kidney damage was poorly investigated. Therefore, in this study, we tested whether the treatment with kaempferol could attenuate Cd-chloride (CdCl2)-induced nephropathy in rats. Besides, we tested the hypothesis that this protection involves activation of Nrf2/and or inhibiting NF-κB.
Materials and methods
Animals
Mature male Wistar rats (150 ± 15 g) were included in this investigation. All rats were provided from the animal facility complex at King Khalid University (KKU), Abha, Kingdome of Saudi Arabia. The rodents were housed in groups of 4 rats/cage under stable, controlled conditions (21 ± 1 ℃, 60% humidity, and 12 h dark/light cycle and always had free access to the chow and drinking water. Rats were adapted for 1 week. All procedures were approved by the ethics committee at the College of Science at KKU.
Experimental design
All animals were randomly selected and divided into 4 groups of rats (n = 10 each) as (1) control rats: orally (gavage) administered 0.25 ml of 1% DMSO (dimethyl sulfoxide) (cat 472,301, Sigma Aldrich, St. Louis, MO, USA) prepared in phosphate-buffered saline (PBS); (2) Kaempferol-treated rats: orally administered an 0.25 ml of kaempferol (200 mg/kg) (cat K0133, Sigma) dissolved in 1% DMSO solution; (3) CdCl2-treated rats: administered CdCl2 (cat 202,908, Sigma) in the drinking water (50 mg/l) and co-treated orally with 0.25 ml of 1% DMSO 4) CdCl2 + kaempferol-treated rats: animals treated with CdCl2 as in group 3 but also co-received the kaempferol solution (200 mg/kg). All drugs were given daily for eight weeks.
Selection of the treatment regimen
The chosen regimen of CdCl2 was adopted for the study of Wang et al. (Wang et al. 2009) who confirmed nephropathy and renal fibrosis by the end of week 8 after daily administration of this dose of Cd ions. The regimen of kaempferol was approved from our previous studies (Alshehri 2021), confirming its therapeutic effect against diabetes mellitus-induced nephropathy.
24-h urine collection
On the last day of week 8, all rats were placed individually in metabolic cages (Lab Products, USA). The 24-h urine samples were collected in 50-ml tubes containing sodium azide. The volume of urine was calculated and all tubes were centrifuged (10 min/1400 × g/room temperature). The collected clear supernatants were aliquot and maintained at − 20 until use.
Collection of the blood and tissues processing
The next day, all rats were fasted overnight and then anesthetized with a mixture of xylazine hydrochloride and ketamine hydrochloride (10 mg/kg and 90 mg/kg) (Cat. No. 61763–23-3. Sigma Aldrich, MO, USA). Blood samples (1 ml) were directly collected by cardiac puncture into empty glass tubes and centrifuged (10 min/1300 × g/room temperature) to collect clear serum which then was stored at − 80 ℃. All animals were then ethically authenticated and both kidneys were isolated on ice, weighed, and cut in transverse sections. Parts of the kidneys were directly placed in 10% buffered formalin and forwarded to the pathology laboratory. All other sections were snap-frozen and kept at − 80 ℃ until use. Later, some parts of these frozen kidney parts were homogenized either in phosphate buffered saline (PBS) or in radioimmunoprecipitation (RIPA) buffer containing protease inhibitor cocktail, centrifuged (12,000 × g/20 min/4℃) to isolate total cell homogenates or protein extracts for the biochemical and western blotting analyses, respectively. All extracts were frozen at – 80 ℃ until use. In addition, the cytoplasmic and nuclear fractions were prepared to form frozen kidney parts using the NE-PER commercially available kit (cat 78,833, ThermoFisher Scientific, USA) as per the manufacturer’s instructions.
Biochemical analysis
Serum and urinary levels of creatinine (Cr) were assessed using a colorimetric-based kit (cat Ab65240, Abcam, UK). The concentrations of albumin in the serum and urine were analyzed using a rat-specific ELISA kit (cat Ab108789, Abcam, Cambridge, UK). The 24-h urinary Cr excretion was calculated using the following formula, UCrE (mg/24 h) = urinary Cr (mg/dl) × urinary volume in 24 h (dl). The 24-h creatinine clearance (Ccr) was calculated according to this formula; Ccr (ml/min) = (urinary Cr (mg/dL) × urine Vol (ml/min))/(serum Cr (mg/dl)). Urinary albumin excretion rate (UAER) and urinary albumin/creatinine ratio (UACR) were calculated using the following equations: [UACR = Urinary albumin (mg/dl) /urinary creatinine (g/dl)] and [UACR = Urinary albumin (mg/dl)/urinary creatinine (g/dl)] (Kim et al. 2016).
Biochemical measurements in total kidney homogenates
Levels of manganese superoxide dismutase (Mn SOD); tumor necrosis factor-α (TNF-α); interlukine-6 (IL-6); GSH; and MDA were analyzed using rats special ELISA kits (cat MBS729914, cat MBS175908, cat MBS175904, cat MBS046356; and cat. MBS268427, MyBioSource, CA, USA, respectively). The assessment of the nuclear concentration of NF-κB p65 and Nrf2 was conducted using commercial ELISA kits (cat. 40,096 and cat. 31,102, respectively, Active Motif, Tokyo, Japan). A commercial fluorometric kit was used to measure levels of free radicals (ROS/RNS) (cat. No. E-BC-K138-F, Elabscience, USA).
Western blots
Proteins from the nuclear and total kidney homogenates were boiled and then diluted in the loading (2 µg/µL). Equal protein concentrations were loaded and separated by SDS-PAGE (8–12%) and then transferred to nitrocellulose membranes (cat Ab133413, Abcam, Cambridge, UK), blocked by skimmed milk (5%), washed with 1X Tris-buffered saline with 0.1% Tween (TBST). The individual membranes were incubated with 1st and 2nd antibodies (prepared in TBST buffer) for 2 h at room temperature. The primary antibodies were Bcl2 (cat. 8276, 28 kDa), Bax (Cat. 20 kDa), cleaved Caspase-3 (Asp175) (cat. 9664, 17/19 kDa), cytochrome-c (cat. 11,940, 14 kDa, 1:1000), NF-κB p65 Antibody (cat. 3034, 65 kDa, 1:1000), keap1 (sc-365626, 69 kDa), Nrf2 (cat. 17,212, 100 kDa, 1:1000), and β-actin (Cat. No. 4970, 45 kDa) (Cell signaling technology), and Lamin A (Cat. No. sc-293162, 69 kDa, 1:1000) (Santa Cruz Biotechnology). After washing, the bands were developed and measured using enhanced Chemiluminescence ECL detection reagent (cat 32,109, ThermoFisher Scientific, MA, USA) and C-Di Git blot scanner (LI-COR, NE, USA). Expressions of nuclear and total proteins were presented as relative to Lamin A and β-actin, respectively.
Histological evaluation
Parts of the kidney section were fixed 10% buffered formalin for 24 h and then were dehydrated in ethanol (70–100%). All samples were then cleared with xylene, embedded in paraffin, cut into fine Sects. (3–5 μm), and then routinely stained with hematoxylin and eosin (H&E) for routine pathological morphology detection. Slides were examined and photographed under a Nikon Eclipse E200, light microscope, Tokyo, Japan.
Statistical analysis
GraphPad Prism statistical software (V8, Sydney, Australia) was used. The normality of the data was tested using the Kolmogorov–Smirnov test. Analysis was accomplished by a 2-way ANOVA analysis followed by the Tukey t-test. Data were presented as means ± SD. Significance was considered at values P ˂ 0.05.
Results
Changes in weights and other urinary and serum-renal parameters
No rat death was viewed in all experimental groups during the whole period of the treatment. Final body weight (FBW), kidney weights, and urine volume, as well as all measured renal-related markers, were not significant when kaempferol-treated rats when compared to the control rats (Table 1). However, FBW and the weights of both kidneys were significantly lower in the CdCl2-treated rats in comparison to the control rats (Table 1). Besides, CdCl2-treated rats showed a significantly less urine volume (oliguria), urinary creatinine levels, and creatinine clearance (Ccr) that was parallel with a significantly higher concentration of serum urea and creatinine (Table 1). Besides, a significant increment in the urinary concentrations of albumin, NAG, and β2-MG, and values of UAER and UACR were depicted in CdCl2-treated rats in contrast to the control rats (Table 1). The levels of all these parameters were significantly reversed in the CdCl2 + kaempferol-treated rats compared to CdCl2-treated rats. Except for FBW, kidney weights, and urine volume, which returned to their basal levels, the values of all other markers remained significantly different (higher/lower) in CdCl2 + kaempferol as compared to control rats (Table 1).
Changes in renal markers of oxidative stress and inflammatory
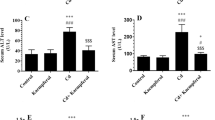
A significant increment in the renal levels of ROS, MDA, IL-6, and TNF-α, and nuclear protein levels of NF-κB p65 with a parallel reduction in the levels of MnSOD and GSH were observed in CdCl2-treated rats in contrast to the control group (Fig. 1A–D and Fig. 2A–D). A significant reduction in the ROS levels, IL-6, MDA, and TNF-α, and the nuclear protein levels of NF-κB p65 with increased levels of MnSOD and GSH were shown in the renal homogenates of both kaempferol- and CdCl2 + kaempferol-treated rodents in contrast to the control rats or CdCl2-treated rats, respectively (Fig. 1A–D and Fig. 2A–D). Interestingly, no significant differences in the values of all these markers were observed when kaempferol-treated rats CdCl2 were compared to controls.

Levels of reactive oxygen species (ROS), malondialdehyde (MDA) (B), manganese superoxide dismutase (MnSOD) (C), and total glutathione (GSH) in the kidneys of all groups of rats. Data were analyzed by two-way ANOVA followed by Tukey’s t-test. All data are presented as mean ± SD (n = 8)

Levels of tumor necrosis factor-α (TNF-α) (A) and interleukin-6 (IL-6) (B), as well as the nuclear activity of Nrf2 NF-Κb P65 (d) in the kidneys of all groups of rats. Data were analyzed by two-way ANOVA followed by Tukey’s t-test. All data are presented as mean ± SD (n = 8)
Changes in Nrf2 signaling
Significantly lower cytoplasmic and nuclear protein levels of Nrf2 with stimulated levels of keap1 and keap1/Nrf2 ratio were depicted in the renal homogenates of CdCl2-treated rats when a comparison was done versus the control group (Fig. 3A–D). On the contrary, higher cytoplasmic and nuclear protein levels of Nrf2 with significant suppression of kepa1 levels and keap1/Nfr2 ratio were observed in the kidneys of the kaempferol- and CdCl2 + kaempferol-treated animals when compared with their vehicle-treated controls (Fig. 3A–D). However, control and CdCl2 + kaempferol-treated rats showed non-significant cytoplasmic and nuclear levels of this transcription factor, as well as in the levels of keap-1 when compared against each other (Fig. 3A–D).

Total protein levels of Nrf2 and keap1 (A), the ratio of keap1/Nrf2 (B), and nuclear protein levels of Nrf2 and NF-κB p65 (D) in the kidneys of all groups of rats. Data were analyzed by two-way ANOVA followed by Tukey’s t-test. All data are presented as mean ± SD (n = 8)
Changes in apoptotic markers
Kaempferol-treated rats seen normal protein levels of all measured apoptotic/anti-apoptotic markers when compared to control animals (Fig. 4A–D). While Bcl2 protein levels were significantly repressed, significantly higher protein levels of cleaved caspase-3, Bax, and Bax/Bcl2 ratio were seen in the renal tissues of the CdCl2-treated rats (Fig. 4A–D). This was reversed in the kidneys of CdCl2 + kaempferol rats when compared to CdCl2-intoxicated animals, values which were within their normal expression depicted in the control rats (Fig. 4A–D).

Total protein levels of Bcl2 (A) and Bax (B), the ratio of Bax/Bcl2 (C), and cytoplasmic protein levels of cleaved caspase-3 in the kidneys of all groups of rats. Data were analyzed by two-way ANOVA followed by Tukey’s t-test. All data are presented as mean ± SD (n = 8)
Histological changes
Control and kaempferol-treated animals showed normal histological features including normally appeared glomeruli, glomerular membranes, proximal convoluted tubules (PCTs), and distal convoluted tubules (DCTs) (Fig. 5A, B). Severe decrease in mesangial mass with damaged glomerular membrane and degenerated proximal and distal convoluted tubules (PCTs and DCTs, respectively) were seen in CdCl2-treated rats (Fig. 5C). Almost normal histological findings like those seen in the control rats were seen in the CdCl2 + kaempferol-treated rats (Fig. 5D).

Histological images of all experimental groups of rats. A and B were taken from control and control + kaempferol-treated rats and showed normal histological features with intact glomeruli (large black arrow), and glomerular membranes (short black arrow), proximal convoluted tubules (PCTs) having brush boarders (long white arrow), and distal convoluted tubules (DCTs) (short white arrow). C was taken from a CdCl2-treated rat and showing decrease mesangial mass and damaged glomerular membrane (arrowhead) and severe damage in both the PCTs and DCTs. D was taken from a CdCl2 + kaempferol-treated rats and showed almost normal architectures like those seen in the control group. However, some abnormalities in the shape of some glomeruli are still seen (star)
Discussion
Data derived from this investigation confirm the protection of kaempferol against CdCl2-induced nephropathy in rats. In addition, it illustrates that the mechanism of protection involves suppressing oxidative stress and inflammation. In addition, the results showed that these effects are associated with activation suppression of Keap1 and NF-κB p65 and concomitant upregulation and activation of Nrf2.
Generally, the kidneys are the major target of Cd2+ ions intoxication which can lead to AKI, CKD, and renal failure (Lentini et al. 2017, Prozialeck and Edwards 2012, Satarug 2018, Wang et al. 2009). Once it reaches the kidneys, Cd2+ ions not only induce proximal tubule injury but the damage also the glamorous and distal convoluted tubules (Diamond et al. 2019, Jiao et al. 2019, Pavón et al. 2019, Prozialeck and Edwards 2012, Satarug 2018). A reduction in food intake due to decreased appetite, loss of weight gain, a decrease in kidney weights, oliguria, micro/macroalbuminuria, and reduced creatinine clearance (Ccr) are the major clinical manifestation for Cd2+ ions-induced nephropathy (Diamond et al. 2019; Pollack et al. 2015; Satarug 2018). Besides, several authors have shown Cd2+ions-induced renal injury leads to a significant release of special damaging markers named, NAG and β2-MG (Bernard et al. 1983, Hagar and Al Malki 2014, Milnerowicz et al. 2008, Prozialeck et al. 2007, Satarug 2018, Yuan et al. 2016). All these alterations were also shown in our animal model, thus validating it. Interestingly, all these changes were independent of water intake, thus dissipating the role of water consumption from the observed effect of CdCl2.
Independent of water intake or improving satiety, kaempferol was not only able to alleviate the oliguria and the alteration in all measured urinary and serum markers, but also prevented the loss in the final body and kidney weights and suppressed the renal damage of the glomeruli and tubules. These data were our first evidence that supports the direct protective impacts of kaempferol against weight loss and CdCl2-induced nephrotoxicity. Although this nephroprotective effect of kaempferol is novel to be shown in this animal model, several studies performed in other tissues of this animal model or kidneys of other animal models can support our findings. Indeed, kaempferol prevented the reduction in rats’ body weights and prevented cortical and hippocampal injury in CdCl2-treated rats without improving food intake (Al-Brakati et al. 2021). Also, kaempferol prevented renal damage and attenuated the alterations in all serum and renal kidney-related markers in diabetic and aged rats, as well as in rats exposed to cisplatin, and tacrolimus (Al-Numair et al. 2015; Ali et al. 2020; Park et al. 2009; Sharma et al. 2020; Wang et al. 2020; Zhang et al. 2019).
Nonetheless, oxidative stress and inflammation are the two leading mechanisms underlying the genotoxic effect of CdCl2 and are associated with suppression of Nrf2/antioxidant and activation of NF-κB signaling (Brzóska et al. 2016; Liu et al. 2019, 2015; Luo et al. 2017; Rani et al. 2014). This has been also confirmed in the CdCl2-treated animals in this study. Herein, the data derived from this study shows that the observed nephroprotective properties of kaempferol involve antagonizing these damaging pathways. Besides, kaempferol has a potent stimulatory potential to enhance the expression and activity of the Nrf2/GSH/SOD axis while it causes repression of NF-κB in the kidneys of control rats too. This suggests that kaempferol is a potent activator of Nrf2 and an inhibitor of NF-κB under the basal and intoxicated conditions. In addition, our data strongly inspire that the stimulatory effect of kaempferol on Nrf2 is mediated via a downregulation of kepa1, which normally stimulates the proteasome degradation of Nrf2 in the cytoplasm (Deshmukh et al. 2017). Therefore, it seems reasonable that CdCl2 suppresses the cellular antioxidants by either direct savaging through overproducing ROS or via modulating the keap1/ Nrf2 axis. So far, the antioxidant potential of kaempferol seems to be mediated by downregulating kepa1 and subsequent transactivation of Nrf2. In support, Liu et al. (Liu et al. 2019) have also shown that CdCl2 induces liver injury by activating NF-κB and keap1 and subsequent inhibition of Nrf2. Also, kaempferol is a potent stimulator of Nrf2 while being a common inhibitor of NF-κB (Saw et al. 2014). However, NF-κB and Nrf2 are negatively crossed-talked with each other (Wardyn et al. 2015). Therefore, it could also be possible that kaempferol stimulates Nrf2 by suppressing NF-κB p65 as shown here. However, the opposite is correct, and maybe kaempferol suppresses NF-κB p65 by activating Nrf2. This cannot be confirmed from these data and requires further investigation.
Supporting our report, pharmacological activation of Nrf2/antioxidant axis is a confirmed strategy to inhibit CdCl2-induced reno-hepatic damage in rats (Diamond et al. 2019; Jiao et al. 2019; Pavón et al. 2019; Wu et al. 2012). Also, accumulating data demonstrates an exceptional ability of kaempferol to attenuate renal, cardiac, hepatic, and neural injury by its antioxidant and anti-inflammatory effects [22]. Similar stimulatory effects of Nrf2 that is coincided with increased antioxidant synthesis and reduced activity of NF-κB were seen in the brains of the control and CdCl2-intoxicated rats after treatment with kaempferol (El-Kott et al. 2020a, 2020c). Also, kaempferol prevented chlorpyrifos-induced brain damage by upregulating/activating Nrf2 and subsequently increasing the antioxidant enzymes. In other animal models, kaempferol prevented doxorubicin (DOX)-induced mitochondria damage, carbon-tetrachloride CCL4-induced hepatic damage, and streptozotocin (STZ)-induced hepatic apoptosis by upregulation of antioxidant and GSH enzymes (Wang et al. 2020). Also, kaempferol prevented cisplatin-induced nephropathy, isoproterenol-induced cardiomyocytes apoptosis in diabetic rats, and oxidative stress-induced umbilical vein endothelial cells (HUVECs) injury by inhibiting NF-κB and concomitant activation of Nrf-2 (Al-Numair et al. 2015; Zhang et al. 2019). Furthermore, kaempferol prevented aged kidney disease by suppressing NF-κB inflammatory cascade and apoptotic MAPKs pathways (P38 and JNK) (Park et al. 2009).
The mitochondria-mediated (intrinsic) cell death is the dominant cell death modality in the kidneys of rodents after intoxication with CdCl2 (Almeer et al. 2019; Fujiwara et al. 2012; Joardar et al. 2019; Lee et al. 2006). Bcl2 is located on the mitochondrial outer membrane where it forms heterodimers with both Bad and Bax to inhibit their apoptotic effects and subsequent mitochondria leakiness and damage (Galluzzi et al. 2018). A higher cytoplasmic ratio of Bax/Bcl2 stimulates the opening of the mitochondria anion channel proteins which end up with the release of cytochrome-c and activating the caspase cascade (Galluzzi et al. 2018). In this investigation, we have confirmed the activation of intrinsic cell death in the kidney of CdCl2-treated rats by the more expression of Bax and cleaved caspase-3 and the concomitant reduction in Bcl2 and the increase in Bax/Bcl2 ratio. Typical data have been previously shown in the kidneys of rats and chickens after CdCl2 administration (Almeer et al. 2019; Bao et al. 2017; Joardar et al. 2019).
However, the potential of kaempferol to attenuate the increment of the expression of Bax and cleaved caspase-3 and to stimulate Bcl2 is the strongest evidence for its anti-apoptotic effect. However, kaempferol did not modulate the expression of any of these markers in the kidneys of control rats. These data suggest that the anti-apoptotic effect afforded by kaempferol is secondary to antioxidant and anti-inflammatory effects. Indeed, ROS can directly induce intrinsic cell death by upregulating p53/Bax axis and Bad (Redza-Dutordoir and Averill-Bates 2016). Also, ROS can activate mitochondria apoptosis by stimulating various MAPKs including p38 and JNK, which may also reduce the Bcl2 levels (Redza-Dutordoir and Averill-Bates 2016). In the same manner, the nuclear translocation of NF-κB triggers both intrinsic and extrinsic cell apoptosis by stimulating Bad and prompting its mitochondria translocation, as well as by raiseing the expression of FAS (Brzóska et al. 2016). Besides, TNF-α is an apoptotic cytokine that stimulates intrinsic renal cell death suppressing Bcl2 and Bcl-xL and activating of MAPKs (Campbell et al. 2008; Pastore et al. 2015).
Study limitations
Despite these data, this study still has some limitations. Importantly, our data are still observational. Besides, based on these data, it is still impossible to determine the upstream mechanism by which kaempferol affords its nephroprotection. Therefore, more focusing on these mechanisms using Nrf2 knockdown animals or cells will be more valuable. The used dose of kaempferol was based on a previously tested nephroprotective dose in diabetic rats. However, further experiments using a dose–response curve are highly recommended. In addition, identifying other pathways regulating inflammation and oxidative stress such as AMPK and SIRT1 may present an excellent scope for future studies. This could be supported by the findings of others who have demonstrated evidence that kaempferol attenuates neural oxidative damage by activating SIRT1 and AMPK (El-Kott et al. 2020a). Finally, this study examined only the preventive effect of kaempferol against CdCl2-induced renal damage. However, it could be more valuable to examine this effect on the kidney structure and function, as well as on all measured markers at different time intervals. In addition, further studies examining the therapeutic effect in rats with pre-established nephropathy of the extract will expand our knowledge about such effect and the mechanisms of action of this drug.
Conclusion
Data obtained from these findings suggest the ability of kaempferol to mitigate CdCl2-induced nephrotoxicity in animals mainly by attenuating oxidative stress and inflammation. However, these findings also demonstrate that the mechanism of action is due to activation of the keap1/Nrf2/antioxidant axis and the concomitant suppressing NF-κB p65. Given the high safety of kaempferol, these data encourage further pre-clinical and clinical studies to validate this effect in patients with CKD in a hope to discover a new safe, cheap, and effective drug. This could be correct knowing that the keap-1/Nrf2 axis is inhibited in the majority of kidney disorders.
Data availability
The data used to support the outcomes of this study are included within the article.
References
Afkarian M, Zelnick LR, Hall YN, Heagerty PJ, Tuttle K, Weiss NS, de Boer IH (2016) Clinical manifestations of kidney disease among US adults with diabetes, 1988–2014. JAMA 316:602–610
Al-Brakati A, Albarakati AJA, Lokman MS, Theyab A, Algahtani M, Menshawi S, AlAmri OD, Al Omairi NE, Essawy EA, Kassab RB, Abdel Moneim AE (2021) Possible role of kaempferol in reversing oxidative damage, inflammation, and apoptosis-mediated cortical injury following cadmium exposure. Neurotox Res 39:198–209
Ali AS, Almalki AS, Alharthy BT (2020) Effect of kaempferol on tacrolimus-induced nephrotoxicity and calcineurin B1 expression level in animal model. J Exp Pharmacol 12:397–407
Almeer RS, AlBasher GI, Alarifi S, Alkahtani S, Ali D, Abdel Moneim AE (2019) Royal jelly attenuates cadmium-induced nephrotoxicity in male mice. Sci Rep 9:5825
Al-Numair KS, Chandramohan G, Veeramani C, Alsaif MA (2015) Ameliorative effect of kaempferol, a flavonoid, on oxidative stress in streptozotocin-induced diabetic rats. Redox Rep 20:198–209
Alshehri AS, El-Kott AF, El-Kenawy AE, Khalifa HS, AlRamlawy AM (2021) Cadmium chloride induces non-alcoholic fatty liver disease in rats by stimulating miR-34a/SIRT1/FXR/p53 axis. Sci. Total Environ. 784:147182
Alshehri AS, El-Kott AF, El-Gerbed MSA, El-Kenawy AE, Albadrani GM, Khalifa HS (2022) Kaempferol prevents cadmium chloride-induced liver damage by upregulating Nrf2 and suppressing NF-κB and keap1. Environ Sci Pollut Res Int 29:13917–13929
Alshehri AS (2021): Kaempferol attenuates diabetic nephropathy in streptozotocin-induced diabetic rats by a hypoglycaemic effect and concomitant activation of the Nrf-2/Ho-1/antioxidants axis. Arch. Physiol. Biochem., 1–14
Alsuwaida AO, Farag YM, Al Sayyari AA, Mousa D, Alhejaili F, Al-Harbi A, Housawi A, Mittal BV, Singh AK (2010) Epidemiology of chronic kidney disease in the Kingdom of Saudi Arabia (SEEK-Saudi investigators) — a pilot study. Saudi J Kidney Dis Transpl 21:1066–1072
Bao RK, Zheng SF, Wang XY (2017) Selenium protects against cadmium-induced kidney apoptosis in chickens by activating the PI3K/AKT/Bcl-2 signaling pathway. Environ Sci Pollut Res Int 24:20342–20353
Bellezza I, Giambanco I, Minelli A, Donato R (2018) Nrf2-Keap1 signaling in oxidative and reductive stress. Biochimica et biophysica acta. Mol Cell Res 1865:721–733
Bernard A, Viau C, Lauwerys R (1983) Renal handling of human beta 2-microglobulin in normal and cadmium-poisoned rats. Arch Toxicol 53:49–57
Birben E, Sahiner UM, Sackesen C, Erzurum S, Kalayci O (2012) Oxidative stress and antioxidant defense. World Allergy Organ J 5:9–19
Brzóska MM, Borowska S, Tomczyk M (2016) Antioxidants as a potential preventive and therapeutic strategy for cadmium. Curr Drug Targets 17:1350–1384
Campbell MT, Dagher P, Hile KL, Zhang H, Meldrum DR, Rink RC, Meldrum KK (2008) Tumor necrosis factor-alpha induces intrinsic apoptotic signaling during renal obstruction through truncated bid activation. J Urol 180:2694–2700
Deshmukh P, Unni S, Krishnappa G, Padmanabhan B (2017) The Keap1-Nrf2 pathway: promising therapeutic target to counteract ROS-mediated damage in cancers and neurodegenerative diseases. Biophys Rev 9:41–56
Devi KP, Malar DS, Nabavi SF, Sureda A, Xiao J, Nabavi SM, Daglia M (2015) Kaempferol and inflammation: from chemistry to medicine. Pharmacol Res 99:1–10
Diamond GL, Thayer WC, Klotzbach JM, Ingerman LD (2019) Urinary cadmium clearance, its relationship to glomerular filtration rate and implications for cadmium epidemiology. J Toxicol Environ Health A 82:1187–1198
El-Kott AF, Abd-Lateif AM, Khalifa HS, Morsy K, Ibrahim EH, Bin-Jumah M, Abdel-Daim MM, Aleya L (2020) Kaempferol protects against cadmium chloride-induced hippocampal damage and memory deficits by activation of silent information regulator 1 and inhibition of poly (ADP-Ribose) polymerase-1. Sci Total Environ. 728:138832
El-Kott AF, Alshehri AS, Khalifa HS, Abd-Lateif AM, Alshehri MA, El-Maksoud MMA, Eid RA, Bin-Meferij MM (2020b) Cadmium chloride induces memory deficits and hippocampal damage by activating the JNK/p(66)Shc/NADPH oxidase axis. Int J Toxicol 39:477–490
El-Kott AF, Bin-Meferij MM, Eleawa SM, Alshehri MM (2020c) Kaempferol protects against cadmium chloride-induced memory loss and hippocampal apoptosis by increased intracellular glutathione stores and activation of PTEN/AMPK induced inhibition of Akt/mTOR signaling. Neurochem Res 45:295–309
Fujiwara Y, Lee JY, Tokumoto M, Satoh M (2012) Cadmium renal toxicity via apoptotic pathways. Biol Pharm Bull 35:1892–1897
Gajjala PR, Sanati M, Jankowski J (2015) Cellular and molecular mechanisms of chronic kidney disease with diabetes mellitus and cardiovascular diseases as its comorbidities. Front Immunol 6:340
Galluzzi L et al (2018) Molecular mechanisms of cell death: recommendations of the Nomenclature Committee on Cell Death 2018. Cell Death Differ 25:486–541
Hagar H, Al Malki W (2014) Betaine supplementation protects against renal injury induced by cadmium intoxication in rats: role of oxidative stress and caspase-3. Environ Toxicol Pharmacol 37:803–811
Hill NR, Fatoba ST, Oke JL, Hirst JA, O’Callaghan CA, Lasserson DS, Hobbs FD (2016) Global prevalence of chronic kidney disease — a systematic review and meta-analysis. PLoS ONE 11:e0158765
Jiao D, Jiang Q, Liu Y, Ji L (2019) Nephroprotective effect of wogonin against cadmium-induced nephrotoxicity via inhibition of oxidative stress-induced MAPK and NF-kB pathway in Sprague Dawley rats. Hum Exp Toxicol 38:1082–1091
Joardar S, Dewanjee S, Bhowmick S, Dua TK, Das S, Saha A, De Feo V (2019): Rosmarinic acid attenuates cadmium-induced nephrotoxicity via inhibition of oxidative stress, apoptosis, inflammation and fibrosis. Int J Mol Sci 20
Kim YG, Byun J, Yoon D, Jeon JY, Han SJ, Kim DJ, Lee KW, Park RW, Kim HJ (2016) Renal protective effect of DPP-4 inhibitors in type 2 diabetes mellitus patients: a cohort study. J Diabetes Res 2016:1423191
Lee WK, Abouhamed M, Thévenod F (2006) Caspase-dependent and -independent pathways for cadmium-induced apoptosis in cultured kidney proximal tubule cells. Am J Physiol Renal Physiol 291:F823–F832
Lentini P, Zanoli L, Granata A, Signorelli SS, Castellino P, Dell’Aquila R (2017) Kidney and heavy metals — the role of environmental exposure (Review). Mol Med Rep 15:3413–3419
Li W, Khor TO, Xu C, Shen G, Jeong WS, Yu S, Kong AN (2008) Activation of Nrf2-antioxidant signaling attenuates NFkappaB-inflammatory response and elicits apoptosis. Biochem Pharmacol 76:1485–1489
Li C, Cheng L, Wu H, He P, Zhang Y, Yang Y, Chen J, Chen M (2018) Activation of the KEAP1-NRF2-ARE signaling pathway reduces oxidative stress in Hep2 cells. Mol Med Rep 18:2541–2550
Liu L, Yang B, Cheng Y, Lin H (2015) Ameliorative effects of selenium on cadmium-induced oxidative stress and endoplasmic reticulum stress in the chicken kidney. Biol Trace Elem Res 167:308–319
Liu C, Zhu Y, Lu Z, Guo W, Tumen B, He Y, Chen C, Hu S, Xu K, Wang Y, Li L, Li S (2019): Cadmium induces acute liver injury by inhibiting Nrf2 and the role of NF-κB, NLRP3, and MAPKs signaling pathway. Int. J. Environ. Res. Public Health 17
Luo T, Liu G, Long M, Yang J, Song R, Wang Y, Yuan Y, Bian J, Liu X, Gu J, Zou H, Liu Z (2017) Treatment of cadmium-induced renal oxidative damage in rats by administration of alpha-lipoic acid. Environ Sci Pollut Res Int 24:1832–1844
Milnerowicz H, Bizoń A, Witt K, Antonowicz-Juchniewicz J, Andrzejak R (2008) Urinary N-acetyl-beta-D-glucosaminidase and its isoenzymes in smoking and non-smoking workers at copper foundry occupational co-exposed to arsenic cadmium and lead. Przegl Lek 65:518–521
Morgan MJ, Liu ZG (2011) Crosstalk of reactive oxygen species and NF-κB signaling. Cell Res 21:103–115
Newsholme P, Cruzat VF, Keane KN, Carlessi R, de Bittencourt PI, Jr. (2016) Molecular mechanisms of ROS production and oxidative stress in diabetes. Biochem J 473:4527–4550
Nezu M, Suzuki N, Yamamoto M (2017) Targeting the KEAP1-NRF2 system to prevent kidney disease progression. Am J Nephrol 45:473–483
Niture SK, Jaiswal AK (2012) Nrf2 protein up-regulates antiapoptotic protein Bcl-2 and prevents cellular apoptosis. J Biol Chem 287:9873–9886
Park MJ, Lee EK, Heo HS, Kim MS, Sung B, Kim MK, Lee J, Kim ND, Anton S, Choi JS, Yu BP, Chung HY (2009) The anti-inflammatory effect of kaempferol in aged kidney tissues: the involvement of nuclear factor-kappaB via nuclear factor-inducing kinase/IkappaB kinase and mitogen-activated protein kinase pathways. J Med Food 12:351–358
Pastore D, Della-Morte D, Coppola A, Capuani B, Lombardo MF, Pacifici F, Ferrelli F, Arriga R, Mammi C, Federici M, Bellia A, Di Daniele N, Tesauro M, Donadel G, Noto D, Sbraccia P, Sconocchia G, Lauro D (2015) SGK-1 protects kidney cells against apoptosis induced by ceramide and TNF-α. Cell Death Dis 6:e1890
Pavón N, Buelna-Chontal M, Macías-López A, Correa F, Uribe-Álvarez C, Hernández-Esquivel L, Chávez E (2019) On the oxidative damage by cadmium to kidney mitochondrial functions. Biochem Cell Biol 97:187–192
Pollack AZ, Mumford SL, Mendola P, Perkins NJ, Rotman Y, Wactawski-Wende J, Schisterman EF (2015) Kidney biomarkers associated with blood lead, mercury, and cadmium in premenopausal women: a prospective cohort study. J Toxicol Environ Health A 78:119–131
Prozialeck WC, Edwards JR (2012) Mechanisms of cadmium-induced proximal tubule injury: new insights with implications for biomonitoring and therapeutic interventions. J Pharmacol Exp Ther 343:2–12
Prozialeck WC, Vaidya VS, Liu J, Waalkes MP, Edwards JR, Lamar PC, Bernard AM, Dumont X, Bonventre JV (2007) Kidney injury molecule-1 is an early biomarker of cadmium nephrotoxicity. Kidney Int 72:985–993
Rani A, Kumar A, Lal A, Pant M (2014) Cellular mechanisms of cadmium-induced toxicity: a review. Int J Environ Health Res 24:378–399
Redza-Dutordoir M, Averill-Bates DA (2016) Activation of apoptosis signalling pathways by reactive oxygen species. Biochim Biophys Acta 1863:2977–2992
Satarug S (2018): Dietary cadmium intake and its effects on kidneys. Toxics 6
Saw CL, Guo Y, Yang AY, Paredes-Gonzalez X, Ramirez C, Pung D, Kong AN (2014) The berry constituents quercetin, kaempferol, and pterostilbene synergistically attenuate reactive oxygen species: involvement of the Nrf2-ARE signaling pathway. Food Chem Toxicol 72:303–311
Sharma D, Kumar Tekade R, Kalia K (2020) Kaempferol in ameliorating diabetes-induced fibrosis and renal damage: an in vitro and in vivo study in diabetic nephropathy mice model. Phytomedicine 76:153235
Vargas F, Romecín P, García-Guillén AI, Wangesteen R, Vargas-Tendero P, Paredes MD, Atucha NM, García-Estañ J (2018) Flavonoids in kidney health and disease. Front Physiol 9:394
Wang SH, Shih YL, Lee CC, Chen WL, Lin CJ, Lin YS, Wu KH, Shih CM (2009) The role of endoplasmic reticulum in cadmium-induced mesangial cell apoptosis. Chem Biol Interact 181:45–51
Wang L, Lin SQ, He YL, Liu G, Wang ZY (2013) Protective effects of quercetin on cadmium-induced cytotoxicity in primary cultures of rat proximal tubular cells. Biomed Environ Sci 26:258–267
Wang Z, Sun W, Sun X, Wang Y, Zhou M (2020) Kaempferol ameliorates cisplatin induced nephrotoxicity by modulating oxidative stress, inflammation and apoptosis via ERK and NF-κB pathways. AMB Express 10:58
Wardyn JD, Ponsford AH, Sanderson CM (2015) Dissecting molecular cross-talk between Nrf2 and NF-κB response pathways. Biochem Soc Trans 43:621–626
Wu KC, Liu JJ, Klaassen CD (2012) Nrf2 activation prevents cadmium-induced acute liver injury. Toxicol Appl Pharmacol 263:14–20
Yuan Y, Ma S, Qi Y, Wei X, Cai H, Dong L, Lu Y, Zhang Y, Guo Q (2016) Quercetin inhibited cadmium-induced autophagy in the mouse kidney via inhibition of oxidative stress. J Toxicol Pathol 29:247–252
Zhang N, Zhao S, Hong J, Li W, Wang X (2019) Protective effects of kaempferol on D-ribose-induced mesangial cell injury. Oxid Med Cell Longev 2019:7564207
Zorov DB, Juhaszova M, Sollott SJ (2014) Mitochondrial reactive oxygen species (ROS) and ROS-induced ROS release. Physiol Rev 94:909–950
Funding
This study was supported by the deanship of Scientific Research at King Khalid University, Abha, KSA, under grant number (R.G.P2 /35/43). Also, this research was funded by the Taif University Researchers Supporting program under grant number (TURSP-2020/99), Taif University, Taif, Saudi Arabia.
Author information
Authors and Affiliations
Contributions
ASA, AFE, and AEE: Conceptualization, Methodology, Validation, Formal analysis, Investigation, Supervision, Project administration, Funding acquisition, Writing—original draft. MSAZ, KM, RAG, MAD, MSM, and ETS: Conceptualization, Validation, Formal analysis, Investigation, Writing—original draft. ERE, EHI, HSK and AEA, MSM, HIMA: Conceptualization, Investigation, Methodology, Writing—review & editing. ASA, AFE, AEE, and EMA: Investigation, Methodology, Writing—review & editing.
Corresponding author
Ethics declarations
Ethics approval
All applicable international, national, and/or institutional guidelines for the care and use of animals and cell lines were followed and approved by the ethics committee at King Khalid University.
Consent to participate
All authors equally participate in the study.
Consent for publication
All authors allow the publication of the paper.
Conflict of interest
The authors declare no competing interests.
Additional information
Responsible Editor: Mohamed M. Abdel-Daim
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Alshehri, A.S., El-Kott, A.F., El-Kenawy, A.E. et al. The ameliorative effect of kaempferol against CdCl2- mediated renal damage entails activation of Nrf2 and inhibition of NF-kB. Environ Sci Pollut Res 29, 57591–57602 (2022). https://doi.org/10.1007/s11356-022-19876-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11356-022-19876-7