
Abstract
In order to understand the molecular characteristics of the Chinese radish, the mitochondrial DNA structure and sequence were analyzed in Chinese wild radish and cultivated varieties. A total of four male-sterile lines, four maintainer lines, 17 cultivars, and a single Chinese wild radish were used, along with 25 male-sterile individuals and 159 fertile plants. We found that the cytoplasm of Chinese radishes could be classified into two types: the normal type and the Ogura type. The Ogura-type cytoplasm was detected in 25 male-sterile plants. The 159 fertile plants had normal cytoplasm. Both the Ogura cytoplasm and the normal cytoplasm were detected in the male-sterile “RA”. The orf138 gene in mitochondrial DNA was detected in cultivated Chinese radish cultivars but not in the wild radish. The Chinese radish orf138 nucleotide sequence was determined in four male-sterile lines and displayed complete homology to the known orf138 type A nucleotide sequence. Three types of mitochondrial orfB (type 1, type 2 and type 3) were found in Chinese radishes. Type 1 was only present in the male-sterile lines. Chinese cultivated radishes were divided into type 2 and type 3, while the Chinese wild radish only had type 3 cytoplasm.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The radish (Raphanus sativus L.) is cultivated worldwide but mainly in eastern Asia where it is used as a vegetable, fodder, grass and oilseed crop (Lu et al. 2008). The radish has a long history of cultivation in China, with an approximate planted acreage of 1,200,000 ha. Radish is the third most consumed vegetable domestically, and the edible organs include the leaves, pod and root (Wang and He 2005). The genetic resource of radish is very abundant in China. To date, more than 2,100 radish accessions have been conserved in the National Mid-term Genebank for Vegetables located in the Institute of Vegetables and Flowers, Chinese Academy of Agricultural Sciences (Kong et al. 2011). Radish breeding research started in the 1950s in China. Before the 1980s, open pollination radish cultivars were dominant; however, since 1990, Chinese breeders have released more than 100 hybrid cultivars, which widespread use has improved yield, quality and disease resistance (He 1993).
Male sterility (MS) and self-incompatibility (SI) are two common methods used to produce hybrid seeds. Compared with SI, MS can improve the purity of hybrids and has been widely employed in commercial production of F1 hybrid radish seed. MS conveys the advantage of plants whose anthers cannot produce functional pollen but retain normal female fertility. MS is classified into two types: nuclear male sterility (NMS) and cytoplasmic male sterility (CMS). In the radish, pollen sterility occurs via CMS as a result of to gain-of-function mutations in the plant mitochondrial genome (Chase and Gabay-Laughnan 2004; Hanson and Bentolila 2004). The CMS is determined by the mitochondrial genome and is associated with a pollen sterility phenotype that can be suppressed or counteracted by nuclear genes known as restorer-of-fertility genes (Rf genes) (Chase 2006, 2007; Chase et al. 2010; Budar and Berthomé 2007; Wise and Pring 2002; Gulyas et al. 2010). The mitochondrial genome is maternally inherited in most plant species (Hagemann 2004). Maternally inherited CMS genes do not segregate; therefore, uniform populations of male-sterile plants can be obtained by simple cross pollination. However, the transcription of plant mitochondrial genes is complex and occasional paternal inheritance can occur in some plants (Azhagiri and Maliga 2007; Elansary et al. 2010; McCauley and Olson 2008).
Radish CMS was first discovered by Dr. Ogura in a Japanese wild radish in 1968 (Ogura 1968). The Ogura CMS is characterized by a mitochondrial male-sterile gene and a nuclear chromosomal gene to restore fertility, which has proved useful in hybrid radish breeding. The Ogura CMS has now been introduced to other cruciferous crops including rapeseed, cabbage, Chinese cabbage, cauliflower and broccoli by intergenetic crosses and protoplast fusion (Bannerot et al. 1977; Sakai and Imamura 1990). Several other CMS types including 77-01A (He et al. 1981), Kos CMS (Ikegaya 1986), NWB-CMS (Nahm et al. 2005) and DCGMS (Lee et al. 2008), have been discovered in China, Japan and South Korea where they have been utilized in the production of F1 hybrid seeds. The molecular and genetic basis of Ogura CMS has been further characterized. The examination of fertile and sterile radish cybrids, a fusion of the Brassica nuclear genome and radish cytoplasm, revealed a region of Ogura cytoplasmic mtDNA that contained a unique transcribed gene, termed orf138, upstream of the orfB (or atp8, encoding ATP synthase subunit 8), which correlates with CMS (Bonhomme et al. 1991,1992; Krishnasamy and Makaroff 1993). Although strong evidence has identified the role of orf138 in pollen abortion, the Ogura-related cytoplasm of wild plants in European natural populations carries an orf138 locus that has lost its ability to induce male sterility due to processing of its transcript, which disrupts the orf138 coding sequence (Giancola et al. 2007). In contrast to many sterility-inducing proteins, the ORF138 protein is not a chimeric polypeptide composed of fragments of conventional mitochondrial proteins. Rather, it has recently been shown to reside in the inner membrane of mitochondria where it is likely assembled as a homopolymer. The mechanism by which it interferes with pollen production is still unclear (Duroc et al. 2005). Nevertheless, because the expression of orf138 has been shown to strongly inhibit bacterial growth, ORF138 is presumed to produce a certain level of toxicity to mitochondrial activity in the tapetum of anthers (Duroc et al. 2005).
Furthermore, orfB, a homolog of atp8 is found in both Ogura and normal cytoplasms. The sequence comparison of orfB showed that the structure in this region can be classified into three types, two of which occur in normal cytoplasm (Terachi et al. 2001). Recently, molecular markers for specific amplification of the orf138 and orfB genes have been used to classify diverse radish germplasms based on their cytoplasmic type (Yamagishi and Terachi 1994; 1996; Yamagishi 2004). Ogura-CMS can be suppressed or counteracted by nuclear Rf genes. Several groups recently succeeded in cloning the Ogura CMS restorer locus, Rfo, in radish by positional cloning (Brown et al. 2003; Desloire et al. 2003; Koizuka et al. 2003). The Rfo locus contains three genes that are organized in tandem, arbitrarily named PPR-A, PPR-B, and PPR-C. These genes are predicted to encode highly similar proteins. PPR-B was genetically defined as the restorer gene and is predicted to encode a pentatricopeptide repeat (PPR) protein that belongs to the P subfamily of PPR genes and comprises 17 PPR repeats (Lurin et al. 2004). The primary role of PPR-B in restoring fertility is to inhibit ORF138 synthesis in the tapetum of young anthers (Uyttewaala et al. 2008).
CMS in Chinese radish is known to consist of male-sterile cytoplasm and two recessive nuclear alleles (Zhang et al. 1999). However, few molecular studies have investigated the CMS of Chinese radish in detail. In this study, we report the identification of Chinese radish cytoplasm types using a PCR assay of mitochondrial DNA. The results will contribute to the breeding of new male-sterile lines and also provide a better understanding of the genetic origin of the radish.
Materials and Methods
Plant Materials
A total of four male-sterile lines, four maintainer lines, 17 cultivars, and a single Chinese wild radish were used in this study, as shown in Table 1. The lines RA, RB, WA, WB, SA and SB were created by Zhang. QA and QB were developed by He et al. (1981). The 17 cultivars are traditional types that are widely grown in China. The wide radish was gifted from Dr. Liu of Nanjing Agricultural University. The population of materials included 25 male-sterile individuals and 159 fertile individuals. The plants were grown in growth chambers with a 16-h photoperiod, at 20°C day and 15°C during the night. The fresh leaves were harvested in liquid nitrogen before use for DNA extraction.
Isolation of Total DNA
Total genomic DNA was extracted from fresh leaves of individual plants using a commercial DNA extraction kit (DNeasy Plant Mini Kit, QIAGEN, Germany) according to the manufacturer’s instructions. The quantity and the quality of DNA extraction were determined by 1.0% agarose gel electrophoresis in 1× TAE buffer using λ-HindIII as a marker (Invitrogen, USA)
Cytoplasm Type Identification Using PCR
Two primer pairs (138-5′KM/138-3′KM and NorfBF/orfBR) were used for the identification of the diverse cytoplasm types (Table 2). ‘MS-Gensuke’ was used as a control for the Ogura cytoplasm and ‘Comet’ as a normal cytoplasm (Yamagishi 2004). PCR amplification was performed by GeneAmp PCR system 9700 (Applied Biosystems, USA). The reaction systems were 25-μl reaction mixtures containing 40 ng template DNA, 2.5 μl of 2 mM 10× TAE buffer, 2.5 μl 2 mM dNTPs, 2.5 μl 25 mM MgCl, 0.2 μl Taq polymerase (Invitrogen, USA), 1.25 μl 10 μM forward and reverse primer solutions and 13.8 μl MilliQ water. PCR was performed with an initial denaturation at 94°C for 3 min followed by 30 cycles of 94°C for 30 s, 55°C for 30 s and 72°C for 1 min. The amplified products were analyzed by 1.5% agarose gel electrophoresis in 1× TAE buffer and stained with ethidium bromide.
PCR-RFLP of the Mitochondrial orfB Region
According to the orfB nucleotide sequence (Terachi et al. 2001), normal and Ogura cytoplasm can be classified into three types (types 1, 2 and 3). Ogura cytoplasm contained only the type 1 sequence, whereas plants with normal cytoplasm had either type 2 or type 3. A nucleotide substitution in orfB sequence at +782 bp gives a type 3-specific SnaBI restriction site. The PCR products amplified using the NorfBF/orfBR primers were digested with SnaBI (NEB, Hertfordshire, UK) following the manufacturer’s instructions and the products were examined by 1% agarose gel electrophoresis.
Male-Sterile Cytoplasm Nucleotide Sequencing Analysis
The orf138 locus was directly sequenced from the PCR product. PCR amplification was performed with the 138,158fw and orfB,rv3 primers (Table 2) and the products were purified to remove unincorporated primers and dNTPs using the PCR-M™ Clean Up System (Viogene-Biotek Corp., Sijhih City, Taiwan). The products were sequenced using the orfB5′RKM, 138,158/fw, 138/xbaI/rv and 138Fo7 primers (Table 2) using the GenomeLab™ Dye Terminator Cycle Sequencing with the Quick Start Kit (Beckman Coulter, Inc., Brea, CA). The sequencing results were analyzed using the Gene Work 2.5.1 software (Beckman Coulter, Inc., Brea CA).
Results
Chinese Radish Cytoplasm Type Detection
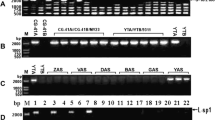
Using specific primers for the orf138 mtDNA gene, associated with Ogura cytoplasm in radish, we observed a 500-bp PCR product in “MS-Gensuke”. Conversely, a 1.4-kb DNA fragment corresponding to amplification of the normal cytoplasm mtDNA sequence was observed in “Comet” (Fig. 1). The two PCR primers were used to identify the diverse cytoplasm in Chinese radish. The 500-bp PCR fragment was found in 25 Chinese radish male-sterile individual plants. The orf138 amplification product was not detected in any of the 159 fertile individuals. Conversely, a 1.4-kb DNA fragment corresponding to amplification of the normal cytoplasm mtDNA sequence was observed in 163 individuals including 159 fertile plants and four sterile plants. The normal-specific band was also absent from 21 individuals from three male-sterile lines (Table 3). These results indicate that all the fertile Chinese radish cultivars tested in this study had normal cytoplasm. Additionally, male-sterile cytoplasm in the Chinese radish is linked to the same mitochondrial gene as Ogura CMS in radish, and the orf138 gene is distributed in Chinese radishes.

PCR amplification of orf138 and normal PCR products from cytoplasmic mtDNA in “MS-Gensuke” and “Comet”. The 500-bp DNA of ‘MS-Gensuke’ was amplified with the primer pair 1385′KM/1383′KM. The 1.4-kb DNA of ‘Comet’ was amplified with the primer pair NorfBF/orfBR. (1) PCR with 1385′KM/1383′KM; (2) PCR with NorfBF/orfBR. M MS-Gensuke, C Comet, MW molecular marker (Marker II, Tiagen, China)
Interestingly, we discovered that the normal cytoplasm-specific band was observed in the male-sterile line RA, a population from the northeast of China (Table 3, Fig. 2). The presence of both the normal cytoplasm and orf138 CMS bands in one individual cultivar suggests the mitochondrial DNA of RA is heterogeneous.

PCR amplification of orf138 and NorfBF PCR products from cytoplasmic mtDNA in four different individual plants of the Chinese radish “RA” cultivar using (1) primer pair 1 (138-5′KM/138-3′KM) to amplify a 500-bp region of the Ogura orf138 gene and (2) primer pair 2 (NorfBF/orfBR) to amplify a 1.4-kb region of the orfB gene. Marker: ФX174/HindIII (Invitrogen, USA)
Distribution of Three Types of orfB in Chinese Radish
To identify the three types (types 1, 2 and 3) of orfB sequence in cytoplasmic mtDNA, PCR amplification was performed in Chinese radishes with normal cytoplasm using the NorfBF/orfBR primer pair and SnaBI PCR-RFLP analysis. Type 2 mitochondrial orfB PCR product does not contain the SnaBI restriction site and produces a single 1.4-kb DNA fragment, whereas type 3 mitochondrial orfB is cleaved by SnaBI to produce two bands at 1.2 kb and 200 bp (Fig. 3). Type 1 (male-sterile cytoplasm) is not amplified using the NorfBF/orfBR primer pair (Fig. 1, Table 2).

PCR amplification and PCR-RFLP of orfB in Chinese radishes with normal cytoplasm. Left: PCR amplification profile showing the 1.4-kb orfB PCR amplification product. Right: PCR-RFLP profile using SnaBI digestion to distinguish between type 2 mitochondrial orfB which does not contain the SnaBI restriction site and type 3 mitochondrial orfB which is cleaved by SnaBI to produce 1.2-kb and 200-bp bands. Type 1 (male-sterile cytoplasm) is not amplified using the NorfBF/orfBR primer pair. (1) Maintainer line ‘WB’; (2) Chinese cultivated radish ‘Beijing Dahongpao’
Table 4 summarizes the distribution of orfB types in Chinese radish, in which all three types of orfB were observed. Among the four male-sterile lines, type 1 was detected in three male-sterile lines, WA, QA and SA, with a frequency of 84.0%, while type 2 was in RA with a frequency of 16.0%. The maintainers possessed type 2 and type 3 with a frequency of 56% and 44%, respectively. In the Chinese cultivated radishes, type 2 and type 3 orfB types was detected and most of the plants (97.4%) possessed type 3. The Chinese wild radish was classified as type 3.
In contrast to the other three male-sterile lines, the populations of the male-sterile line “RA” tested were all type 2. Its maintainer line, the “RB” population, was composed of two orfB types, with two plants having type 2 and five plants having type 3. Most of the Chinese radish cultivars were of only one type, while polymorphisms of both type 2 and type 3 genes were observed in the “RB” maintainer and “Nan Pan Zhou” cultivar (Table 4).
orf138 Nucleotide Sequence in Cytoplasmic Male-sterile Chinese Radishes
A large-scale sequence analysis has previously indicated the orf138 sequence can be classified into nine types, termed A–I (Yamagishi and Terachi 2001). To confirm the orf138 types in Chinese radishes with CMS, the nucleotide sequences of the PCR-amplified fragments from the four male-sterile lines were analyzed. One plant from each male-sterile line was sequenced and the orf138 sequence in each was identical to the published type A orf138 (Fig. 4).

Sequence alignment showing complete homology between the orf138-A type nucleotide sequences and orf138 sequences in the Chinese radish RA cultivar
Discussion
Since the first report of Ogura male-sterile cytoplasm (Ogura 1968), mitochondrial genomes in the radish have been classified into Ogura or normal cytoplasm. Many molecular markers, based on the presence or absence of gene-specific PCR products, have previously been used to distinguish Ogura and normal cytoplasms in different radish cultivars (Yamagishi and Terachi 1994; 1996; Lee et al. 2005). In this study, the cytoplasm type was distinguished using specific markers, and in contrast to the previous studies in radish, we observed two types of cytoplasm in Chinese radish: the Ogura type and the normal type.
Heterogeneous cytoplasm, which was identified by the presence of both Ogura and normal cytoplasm-type PCR amplification products, was found only in the “RA” male-sterile line. The nucleotide sequence of the PCR-amplified fragments from “RA” was identical to orf138, which indicated that the normal PCR-amplified fragment in “RA” was not due to the presence of normal mitochondrial sequences. The heterogeneous cytoplasm may have had a copy of the normal orfB that was introduced in the mt genome. Further investigations should be performed to identify the normal cytoplasm type PCR amplification products in “RA”.
The original “RA” male-sterile plants were found in a single cultivar from the northeast region of China. Using the male-sterile plants as female parents, the male-sterile line was developed by continuous backcrossing of the fertile plants as male parents. Chimeric individuals have been observed in this variety, and the male sterility ratio varies from 97% to 100% in different seasons (data not shown). It may be possible that the heterogeneous cytoplasm observed in this variety can lead to unstable fertility, or that the environmental conditions may influence the expression of mitochondrial genes associated with fertility.
Orf138, the Ogura-specific mitochondrial gene, was found in the four male-sterile lines of Chinese cultivated radish, which have different morphologies and originated from different regions of China. For example, the RA male-sterile line was transferred from the Chinese cultivar ‘Dinglonghong’ which has a large spherical root, red skin, white flesh and is grown in the northeast of China. WA, which has a long cylindrical root, white skin and flesh, was from the south of China. QA was bred by backcrossing from the ‘Qingyuancui’ cultivar and SA arose from the ‘Xiaowuying’ cultivar. The orf138 gene sequence in the cytoplasm of these four male-sterile lines is identical to the orf138 type A sequence with no detected polymorphisms. It was identical to the ‘Chibetto Kei Daikon’ cultivar from the southwest of China. However, a previous study showed that five different Chinese radish cultivars from southern China contained the type H orf138 sequence (Yamagishi and Terachi 2001). It is notable that cultivars with Ogura mtDNA have been found in diverse geographical regions in south China and Japan, and they have very different morphologies (Yamagishi and Terchi 1996). It is difficult to assume that these cultivars with Ogura mtDNA had a common ancestor.
Terachi et al. (2001) described three types of cytoplasm in wild and cultivated radishes that differed in their orfB 5′ flanking sequences. One cytoplasm type was found to be strictly linked to orf138 and found in Ogura and Ogura-related cytoplasm, while the other two types were present in normal cytoplasm. We analyzed the mitochondrial orfB locus using PCR-RFLP in Chinese radish Ogura-related cytoplasm and normal-related cytoplasm, and investigated the classification of orfB in these populations (Table 5). The majority of Chinese radishes were type 3 and type 2, with these types distributed in several cultivated radishes including a male-sterile line, three maintainers and one cultivated radish from southern China. Two cultivars, RB and Nan Pan Zhou, showed polymorphic type 2 and 3 cytoplasms. Most Japanese radishes and Japanese cultivars are type 2, with the other types of orfB present less frequently (Yamagishi 2004). Three types of orfB have been described in cultivated Chinese radishes, and our study indicates that the frequency of the three types of cytoplasm varies in Chinese radishes. Additionally, these findings suggest that the mtDNA of cultivated radishes in China and Japan is highly similar but that a significant amount of variation has occurred in the course of evolution.
Male-sterile plants are very useful in hybrid radish seed production, and consequently, radish breeders have produced a significant amount of experimental data on the process of obtaining and maintaining CMS. The identification of their cytoplasm will help to identify the initial material for the new CMS lines.
Abbreviations
- CMS:
-
Cytoplasmic male sterility
- MS:
-
Male sterility
- NMS:
-
Nuclear male sterility
- Rf:
-
Restorer-of-fertility
- SI:
-
Self-incompatibility
References
Azhagiri AK, Maliga P (2007) Exceptional paternal inheritance of plastids in Arabidopsis suggests that low-frequency leakage of plastids via pollen may be universal in plants. Plant J 52:817–823
Bannerot H, Boulidard L, Chupeau Y (1977) Unexpected difficulties met with the radish cytoplasm in Brassica oleracea. Eucarpia Cruciferae Newslett 2–16
Bonhomme S, Budar F, Férault M, Pelletier G (1991) A 2.5 kb NcoI fragment of Ogura radish mitochondrial DNA is correlated with cytoplasmic male-sterility in Brassica cybrids. Curr Genet 19:121–127
Bonhomme S, Budar F, Lancelin D, Small I, Defrance MC, Pelletier G (1992) Sequence and transcript analysis of the Nco2.5 Ogura-specific fragment correlated with cytoplasmic male sterility in Brassica cybrids. Mol Gen Genet 235:340–348
Brown G, Formanová N, Jin H, Wargachuk R, Dendy C, Patil P, Laforest M, Zhang J, Cheung WY, Landry BS (2003) The radish Rfo restorer gene of Ogura cytoplasmic male sterility encodes a protein with multiple pentatricopeptide repeats. Plant J 35:262–272
Budar F, Berthomé R (2007) Cytoplasmic male sterilities and mitochondrial gene mutations in plants. In: Logan DC (ed) Plant mitochondria. Blackwell, Oxford, pp 278–307
Chase CD (2006) Genetically engineered cytoplasmic male sterility. Trends Plant Sci 11:7–9
Chase CD (2007) Cytoplasmic male sterility: a window to the world of plant mitochondrial–nuclear interactions. Trends Genet 23:81–90
Chase CD, Gabay-Laughnan S (2004) Cytoplasmic male sterility and fertility restoration by nuclear genes. In: Daniell H, Chase CD (eds) Molecular biology and biotechnology of plant organelles. Springer, Netherlands, pp 593–622
Chase CD, Ribarits A, Herberle-Bors E (2010) Male sterility. In: Pua EC, Davey MR (eds) Plant developmental biology–biotechnological perspectives. Springer, Berlin, pp 437–457
Desloire S, Gherbi H, Laloui W, Marhadour S, Clouet V, Cattolico L, Falentin C, Giancola S, Renard M, Budar F, Small I, Caboche M, Delourme R, Bendahmane A (2003) Identification of the fertility estoration locus, Rfo, in radish, as a member of the pentatricopeptide-repeat protein family. EMBO Rep 4(6):588–594
Duroc Y, Gaillard C, Hiard S, Defrance M, Pelletier G, Budar F (2005) Biochemical and functional characterization of ORF138, a mitochondrial protein responsible for Ogura cytoplasmic male sterility in Brassiceae. Biochimie 87:1089–1100
Elansary HO, Müller K, Olson MS, Štorchová H (2010) Transcription profiles of mitochondrial genes correlate with mitochondrial DNA haplotypes in a natural population of Silene vulgaris. BMC Plant Biol 10:11–16
Giancola S, Rao Y, Chaillou S, Hiard S, Martin-Canadell A, Pelletier G, Budar F (2007) Cytoplasmic suppression of Ogura cytoplasmic male sterility in European natural populations of Raphanus raphanistrum. Theor Appl Genet 114:1333–1343
Gulyas G, Shin Y, Kim H, Lee JS, Hirata Y (2010) Altered transcript reveals an orf507 sterility-related gene in chili pepper (Capsicum annuum L.). Plant Mol Biol Rep 28:605–612
Hagemann R (2004) The sexual inheritance of plant organelles. In: Daniell H, Chase C (eds) Molecular biology and biotechnology of plant organelles. Springer, Dordrecht, pp 93–114
Hanson MR, Bentolila S (2004) Interactions of mitochondrial and nuclear genes that affect male gametophyte development. Plant Cell 16:154–169
He QW (1993) Hybrid breeding of cruciferae vegetables. Agricultural Publishing House, Beijing, pp 151–214
He QW, Shi H, Liu E (1981) The development of 77-01A CMS in Chinese radish. Shangdong Agric Sci 1:13–16
Ikegaya Y (1986) Frequent appearance of cytoplasmic male sterile plants in a radish cultivar Kosena. Jpn J Breed 36(Suppl 2):106–107
Koizuka N, Imai R, Fujmoto H, Hayakawa T, Kimura Y, Kohno-Murase J, Sakai T, Kawasaki S, Imamura J (2003) Genetic characterization of a pentatricopeptide repeat protein gene, orf687, that restores fertility in the cytoplasmic male-sterile Kosena radish. Plant J 34:407–415
Kong QS, Li XX, Xia CP, Wang HP, Song JP, Zhi HY (2011) Genetic diversity of radish (Raphanus sativus L.) germplasm resources revealed by AFLP and RAPD markers. Plant Mol Biol Rep 29:217–223
Krishnasamy S, Makaroff CA (1993) Characterization of the radish mitochondrial orfB locus: possible relationship with male sterility in Ogura radish. Curr Genet 24(1–2):156–163
Lee H, Lee S, Joo G, Harn C, Yang S, Min B (2005) Development of a molecular marker specific to a novel CMS line in radish (Raphanus sativus L.). Theor Appl Genet 111:1191–1200
Lee Y, Park S, Lim C, Kim H, Lim H, Ahn Y, Sung S, Yoon M, Kim S (2008) Discovery of a novel cytoplasmic male-sterility and its restorer lines in radish (Raphanus sativus L.). Theor Appl Genet 117:905–913
Lu N, Yamane K, Ohnishi O (2008) Genetic diversity of cultivated and wild radish and phylogenetic relationships among Raphanus and Brassica species revealed by the analysis of trnK/matK sequence. Breeding Sci 58:15–22
Lurin C, Andres C, Aubourg S, Bellaoui M, Bitton F, Bruyere C, Caboche M, Debast C, Gualberto J, Hoffmann B, Lecharny A, Le Ret M, Martin-Magniette ML, Mireau H, Peeters N, Renou JP, Szurek B, Taconnat L, Small I (2004) Analysis of Arabidopsis pentatricopeptide repeat proteins reveals their essential role in organelle biogenesis. Plant Cell 16:2089–2103
McCauley DE, Olson MS (2008) Do recent findings in plant mitochondrial molecular and population genetics have implications for the study of gynodioecy and cytonuclear conflict? Evolution 62:1013–1025
Nahm S, Lee H, Lee S, Joo G, Harn C, Yang S, Min B (2005) Development of a molecular marker speciWc to a novel CMS line in radish (Raphanus sativus L.). Theor Appl Genet 111:1191–1200
Ogura H (1968) Studies on the new male sterility in Japanese radish, with special reference to the utilization of this sterility towards the practical raising of hybrid seeds. Mem Fac Agric Kagoshima Univ 6:39–78
Sakai T, Imamura J (1990) Intergenetic transfer of cytoplasmic male sterility between Raphanus sativus (cms line) and Brassica napus through cytoplast–protoplast fusion. Theor Appl Genet 80:421–427
Terachi T, Yamaguchi K, Yamagishi H (2001) Sequence analysis on the mitochondrial orfB locus in normal and Ogura male sterile cytoplasms from wild and cultivated radishes. Curr Genet 40:276–281
Uyttewaala M, Arnala N, Quadradoa M, Martin-Canadella A, Vrielyncka N, Hiarda S, Gherbic H, Bendahmanec A, Budara F, Mireaua H (2008) Characterization of Raphanus sativus pentatricopeptide repeat proteins encoded by the fertility restorer locus for Ogura cytoplasmic male sterility. Plant Cell 20:3331–3345
Wang LZ, He QW (2005) Chinese radish. Scientific and Technical Document Publishing House, Beijing, pp 6–13
Wise RP, Pring DR (2002) Nuclear-mediated mitochondrial gene regulation and male fertility in higher plants: light at the end of the tunnel? PNAS 99:10240–10242
Yamagishi H (2004) Assessment of cytoplasmic polymorphisms by PCR-RFLP of the mitochondrial orfB region in wild and cultivated radishes (Raphanus). Plant Breed 123:141–144
Yamagishi H, Terachi T (2001) Intra- and inter-specific variations in the mitochondrial gene orf138 of Ogura-type male-sterile cytoplasm from Raphanus sativus and Raphaus raphanistrum. Theor Appl Genet 103:725–732
Yamagishi H, Terchi T (1994) Molecular and biological studies on male-sterile cytoplasm in the Cruciferae. I. The origin and distribution of Ogura male-sterile cytoplasm in Japanese wild radishes (Raphanus sativus L.) revealed by PCR-aided assay of their mitochondrial DNAs. Theor Appl Genet 87:996–1000
Yamagishi H, Terchi T (1996) Molecular and biological studies on male-sterile cytoplasm in the Cruciferae. I. Distribution of Ogura male-sterile cytoplasm among Japanese wild radishes and Asian radish cultivars. Theor Appl Genet 93:325–332
Zhang L, Sheng XQ, Zhao GY (1999) Inheritance of male sterility in spring–summer radish. Acta Horticulturae Sinica 26:238–243
Acknowledgements
This work was supported by a grant form the Visiting Scholarship Program of China Scholarship Council. We would like to thank E. Tomioka, Y. Yamashita and Y. Matsumoto for their technical assistance.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Zhang, L., Yasumoto, K. & Yamagishi, H. Identification of Cytoplasmic Male Sterility in Chinese Radish Following PCR Analysis of Mitochondrial DNA. Plant Mol Biol Rep 30, 817–826 (2012). https://doi.org/10.1007/s11105-011-0391-5
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11105-011-0391-5