
Abstract
In southern Bahia, Brazil, cabrucas are the traditional agroforests in which cacao trees are planted under thinned-out native forests. To analyze the role of cabrucas in tree species conservation, we inventoried the non-cocoa trees in 1.0 ha plots of cabruca in 16 cocoa farms and compared our results with a similar survey undertaken in the early 1960s in the same region to analyze the long term changes. We also interviewed 160 cocoa farmers to investigate their preferences for species and the main practices used in managing shade trees. The cabrucas showed high levels of tree diversity for an agroforestry system (Shannon index ranging from 2.21 to 3.52) and also high variation in structure and composition among the different farms. Forest specialist trees accounted for most species (63.9%) in the survey and were among the species most preferred by the farmers, although we found evidence that some of these trees are gradually being replaced by other species. Our results indicate that cabrucas are poor substitutes for undisturbed forests in terms of tree species richness, but their presence in human-altered landscapes is of utmost importance to the conservation of forest tree species as they increase overall heterogeneity and may serve as ecological corridors, additional habitats, and buffer zones.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
The conversion of tropical forests to agricultural and pasture lands is a major cause of deforestation, resulting in biodiversity losses and reductions in ecosystem services throughout the world (Geist and Lambin 2002). To meet the challenges of maximizing conservation in human-altered landscapes it has become increasingly necessary to examine the ability of different forms of land-use to minimize biodiversity losses and to manage these landscapes for conservation purposes (Vandermeer and Perfecto 2007). Complex agroforests are forest-like agroforestry systems that are more amenable than other forms of human land use to the conservation of biodiversity, soils and water, as well as carbon storage (Rice and Greenberg 2000; Schroth et al. 2001, 2004; Gordon et al. 2007; Delabie et al. 2007; Sambuichi and Haridasan 2007; Jose 2009; Méndez et al. 2009). Agroforests can provide habitat for many forest species and function as ecological corridors and buffer zones, and their presence in anthropogenic landscapes can minimize the impacts of human activities on forest remnants (Bhagwat et al. 2008; Perfecto and Vandermeer 2008). However, the contribution of agroforests to biodiversity conservation depends greatly on their structure, composition, management, their proximity to forest patches, the quality and extension of intact forests in the landscape, as well as the species groups considered (Faria et al. 2006, 2007; Cassano et al. 2009; Pardini et al. 2009).
Traditional cocoa agroforests are complex, multilayered agroforestry systems that combine cocoa tree crops (Theobroma cacao) with many diverse shade trees (Rice and Greenberg 2000). These cocoa agroforests occur in tropical regions in Central and South America, Africa and Asia and have traditionally been established in the shade of thinned-out forests after removing the understory and some of the canopy trees (Ruf and Schroth 2004). Cabruca is the term used to designate the traditional cocoa cultivation system found in southern Bahia State, Brazil’s main cocoa-producing region. The original vegetation in this region was the Brazilian Atlantic Forest. This tropical rainforest covered in the past all the east coast of Brazil, but has been reduced to only about 7% of its original coverage, with the remaining areas being highly disturbed and fragmented (Fundação SOS Mata Atlântica, Instituto Nacional de Pesquisas Espaciais 2008). It is characterized by high species richness and endemism and is considered a priority area (hotspot) for global biodiversity conservation efforts (Myers et al. 2000). In southern Bahia, this forest was largely replaced by cabrucas, and the current landscape is a mosaic composed mainly of small forest fragments immersed in a matrix of shaded cocoa plantations (Saatchi et al. 2001).
Cabrucas were intensively installed in southern Bahia since the mid-nineteenth century and have passed through several alternating periods of reduction or expansion of cultivated areas. In the early 1960s, the Brazilian government (through the Executive Committee for Cocoa Farming Plan—CEPLAC) promoted a technological package aimed at intensifying cocoa production systems through the reduction of shade levels and the replacement of cabrucas by cocoa plantations with monospecific shade trees. However, due to risk-aversion attitudes predominant among many farmers, the program was not successful in promoting the widespread replacement of cabrucas or reduction in shade levels (Johns 1999). The farmers perceived that replacing cabrucas would be expensive, and that reducing the shade levels in cocoa plantations would increase production costs by demanding greater expenditures for fertilizers, herbicides and insecticides. In light of the wide price fluctuations of international cocoa markets, farmers have generally found it more prudent to maintain the cabruca shade trees, since increased productivity would not compensate the extra costs involved in more intensive and less-shaded production systems (Johns 1999).
A major crisis hit the local cocoa production sector in the 1980s as a result of low cocoa prices and the introduction of the “witches’ broom” disease caused by the fungus Moniliophthora perniciosa, and many cabruca areas were replaced by more intensive cultivation systems or pastures, while others were simply abandoned. Environmental (climate, topography, soils) and socioeconomic (commercialization channels and logistics) factors in the cocoa-growing region did not favor cattle raising or the cultivation of other crops, however, and cabrucas have remained the principal landscape component. In the early 1990s, it was estimated that 70% of the 6,800 km2 of cocoa plantations in the region are cabrucas (Alger and Caldas 1994). No new cabruca areas are currently being established in forested areas as Brazilian law no longer allows the conversion of Atlantic Forest vegetation into agricultural land.
Studies on the shade trees component of cabrucas have shown that these agroforests have high structural and floristic diversity (including many endemic and endangered tree species) in comparison to other agricultural systems (Sambuichi et al. 2008). However, some studies have demonstrated that the forest trees shading the cocoa plantations are gradually being replaced by early successional and exotic arboreal species, indicating that the cabrucas are suffering alterations of their species composition over time (Rolim and Chiarello 2004; Sambuichi 2006; Sambuichi and Haridasan 2007).
In order to evaluate the role of cabrucas in the conservation of Atlantic Forest tree species and to contribute to conservation efforts, it will be necessary to understand the factors that affect the composition and dynamics of tree species in these agroforests. We investigated the value of cabrucas for tree species conservation by analyzing the influence of vegetation structural parameters (density and basal area) on the richness, diversity and composition of cabruca shade tree species. We carried out a survey of the non-cocoa trees in cabrucas in the cocoa-growing region of southern Bahia and compared these data with the results of an earlier survey undertaken in the early 1960s by Alvim and Pereira (1965) to access the long-term changes. We also investigated the preferences of farmers for certain species and the principal management practices for shade trees to aid in our understanding of the composition and dynamics of cabruca systems. Based on these results, we discussed possible strategies to conciliate profitability and tree species conservation in cabrucas.
Materials and methods
Study area
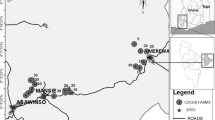
The present study was conducted from July 2008 to January 2009 in the cocoa zone of southern Bahia State, Brazil (Fig. 1), which retains some of the last Atlantic Forest remnants in northeastern Brazil. In southern Bahia, these forest remnants are considered of extremely high biological importance, showing some of the highest values of woody species richness in the world and very high indices of plant endemism (Thomas et al. 1998), being a hotpoint for biodiversity conservation in the Atlantic Forest hotspot (Martini et al. 2007). Shaded cocoa plantations are the main matrix of the landscape, which is also composed of fragments of forest, successional vegetation, pastures and other crops.

Study area in southern Bahia State, Brazil, indicating the locations of the farms where the tree surveys were carried out
The climate in the study area is tropical, hot, and humid (Af by the Köppen system) without a well-defined dry season. There is a rainfall gradient that decreases from the coast towards the interior, and from north to south, with annual totals exceeding 1,200 mm (and reaching over 2,000 mm in the rainiest areas). Mean annual temperatures range from 22° to 25°C. The predominant soils are Alfisols and Ultisols, with Histosols being found at lower altitudes and Inceptisols at higher elevations. Altitudes range from sea level to near 600 m.
Tree survey
We surveyed the non-cocoa trees (diameter at breast height—DBH ≥ 10 cm) in cabrucas on 16 cocoa farms located in 13 municipalities in the region (Fig. 1). We sampled a 1.0 ha area in each farm using four 50 × 50 m plots at different topographic positions. The species were identified by comparison with collections held in the Cocoa Research Center (CEPEC) and the State University of Santa Cruz (UESC) herbaria, or with the assistance of experienced taxonomists.
We considered the total 16 ha area surveyed for each species and calculated tree density, basal area, frequency and importance value (IV) (the sum of relative values of density, basal area and frequency) (Curtis and McIntosh 1951). For each farm we considered the total 1.0 ha area surveyed and calculated tree density, basal area, Shannon diversity index (using the natural logarithms of the numbers of individuals) and Pielou’s evenness index (Magurran 1988). Floristic similarities among the farms were measured using the Sørensen index (Mueller-Dumbois and Ellenberg 1974). Diameter distributions of individuals were compared among the 16 farms using the Kolmogorov–Smirnov test. The Pearson’s correlation method was used to correlate floristic parameters (diversity, richness and composition) of cabrucas with their vegetation structure [tree densities and basal areas (BA)]. As the species/individuals relationships are usually logarithmic, we also correlated the floristic parameters with the natural logarithm of density. Simple linear regression analysis was used to fit the curves among selected parameters.
Native species were classified according to their status in the successional gradient, observing the seedling shade tolerance, growth rate and average life span of the tree in nature. This classification was based on the field experience of the present authors, supplemented by the opinion of other regional experts and consulting the information available in literature. We used four ecological groups, following Budowski (1965) with few modifications: Pioneer species: shade intolerant, fast growing, life cycles usually less than 20 years; Early secondary species: intolerant or only partially shade tolerant, rapid growth, average life spans from 20 to 50 years; Late secondary species: partially shade tolerant, intermediate growth rates, life spans over 50 years; Climax species: shade tolerant, slow growth, life spans over 50 years. We also differentiate between open environment species (pioneer and early secondary species) and forest specialist species (late secondary and climax species).
In order to access the long-term changes in the arboreal component of cabrucas we compared our results with those of Alvim and Pereira (1965). These authors surveyed all shade trees (DHB ≥ 10 cm) in 1.0 ha plots of cabruca in 61 cocoa farms in 30 municipalities in the cocoa-growing region of southern Bahia in the early 1960s and recorded the total number of species; average densities and their ranges; and provided a table with the numbers of trees of the 24 major species.
Survey of farmers’ preferences and management practices
A total of 160 farms located in 14 municipalities with the largest cocoa production in southern Bahia were visited (Fig. 2). We adopted a stratified sampling for selecting cocoa farms that was based on the proportions of farms that cultivated cocoa in these municipalities under differing land tenure systems (family farms, patronal farms, and commercial farms) and differing sizes (very small, small, medium, and large) within the set of farms registered by CEPLAC. Most of the farms surveyed belonged to the “patronal agriculture” land tenure type (84.5%) (where the landowner does not work on the farm, but rather hires workers or contracts sharecroppers), followed by “family agriculture” (12.3%) (most farm labor being provided by the landowners and their family members), and “commercial agriculture” (3.2%) (farms owned by companies). More than half of the farms (53%) belonged to the small (20–80 ha) size category, 24% were medium sized (80–300 ha), 16% were very small (less than 20 ha), while only 7% were large (more than 300 ha). Most family-run farms in the sample were in the “very small” size category, while most patronal farms were small or medium sized. Commercial farms were mostly represented in the medium and large size categories. The farms in all of the land tenure and size categories were randomly selected.

Locations of farms visited during the surveys of farmers’ preferences and management practices
A questionnaire was submitted to the person responsible for the farm at the time of our visit and included farm managers (37.3%), landowners (34.8%), sharecroppers (21.7%), and farm workers (6.2%). The questionnaire was used to identify the frequency of adoption of management practices that affected the composition and diversity of shade trees in cabruca areas such as: selective weeding (selection of certain seedlings during weeding to allow their natural regeneration); cutting shade trees; and planting shade trees. For each management practice reported we enquired about the farmer’s personal preference for certain species and the reasons why those species were preferred over others. We also asked about their perception of the level of shade in their cabruca; the uses of cabruca tree species for food, firewood, timber, and medicinal purposes; and the commercialization of shade tree products. The respondents were also asked to indicate three tree species they would want to save in a hypothetical situation in which all the other non-cocoa trees had to be eliminated. We used the χ2 statistical test to analyze whether there was a significant difference between the classes of tenure system, size of the farm and function of person responding the questionnaire in the frequency of adoption of the management practices and frequency of citation of the two most cited tree species for each practice.
Results
Vegetation structure and composition
A total of 1,933 non-cocoa trees belonging to 216 species and 49 families were found in the 16 ha of cabrucas surveyed. The families with the greatest varieties of species were: Fabaceae (51 species), Moraceae (16) and Euphorbiaceae (15); these three families also had the highest numbers of individuals (654, 330 and 153, respectively). The most common species in the survey was Artocarpus heterophyllus Lam. (jackfruit tree) (IV = 27.1), an exotic fruit tree with the largest number of individuals (269), the highest frequency of occurrence (72% of plots), as well as the largest basal area (26.5 m2) (Table 1). Other species of significant importance were: Plathymenia foliolosa Benth (“vinhático”) (IV = 14.1), a native timber species; and Spondias mombin (“cajá”) (IV = 13.4), a fruit tree from the Amazon rainforest. The 30 most common species accounted for 69% of the individuals encountered; 44% of the species were represented by only a single individual in the survey; 46 species were endemic to Brazil, and 22 endemic to the Brazilian Atlantic Forest.
Native trees accounted for most individuals (73.9%) and species (92.6%) in the survey, comprising 314 pioneer trees, 455 early secondary trees, 391 late secondary trees, and 258 climax trees (Table 2). The climax class had the highest number of species (98), followed by the late secondary (40), early secondary (33), and pioneer (18) classes. A total of 16 exotic species (504 trees) were encountered, surpassing the numbers of trees in any of the native successional classes. The late secondary class had the largest total basal area (109.9 m2) (Table 2). Open environment species (pioneer and early secondary) accounted for 39.7% of all trees, forest specialist species (late secondary and climax) for 33.5%, and exotics for 26.1%.
There were large variations in most structural and floristic parameters of the trees among the 16 cabruca farms surveyed. The highest coefficient of variation (CV) among all the farms (the sixteen 1.0 ha plots) was observed for density (60.7%), which ranged from 43 to 284 (average 121 ± 73) individuals per ha (Table 3). Basal area varied less than density (CV = 23.5%), ranging from 12.3 to 30.9 m2 ha−1; there was no significant correlation between these two structural parameters on the farms. We did observe, however, a highly significant negative correlation between basal area per individual and logarithm of density (r = −0.829, p < 0.001). In most cases, the distribution of individuals per diameter class was significantly different among the different farms (Online Resource 1 and 2).
The floristic similarity (Sørensen qualitative) among the farms (1.0 ha plots) ranged from 0.07 to 0.49, averaging 0.29 ± 0.09. Species diversity (Shannon) ranged from 2.21 to 3.52 (average 3.03 ± 0.39) and showed no significant correlation with tree density. The average evenness (Pielou) was 0.86 ± 0.06 and tended to decrease as logarithm of density increased (r = −0.587, p = 0.017). Total richness ranged from 16 to 60 (average 36 ± 13) species per ha, and there was a strong significant positive correlation with the logarithm of density (r = 0.841, p < 0.001). The richness of open environment species (r = 0.874, p < 0.001) and forest specialist species (r = 0.647, p = 0.007) are also positively correlated with the logarithm of density.
We observed an increase in total species richness and in forest specialist species richness with increasing density (in a logarithmic relationship), however, forest specialist richness increased less than the total species richness (Fig. 3). The proportions of forest specialist trees on farms decreased with increasing density, while the proportions of open environment trees increased (Fig. 4)—showing that denser areas had proportionally more early successional trees. The richness of exotic species and the proportions of exotic trees were not significantly correlated with density. Basal area showed no significant correlation with any of the floristic parameters examined.

Increase in total species richness and forest specialist species richness with increasing trees densities in 16 cabruca areas in southern Bahia, Brazil

Percentages of forest specialist trees (late secondary + climax) and open environment trees (pioneer + early secondary) versus tree densities in 16 cabruca areas in southern Bahia, Brazil
Vegetation changes
In comparing the results of the present study with those of Alvim and Pereira (1965) some differences were observed indicating some changes that occurred in cabrucas over the past 45 years. Alvim and Pereira (1965) found 171 tree species in their survey of 61 ha. Tree densities on the farms ranged from 25 to 323 trees per ha, with an average of 76 trees per ha. The mean tree density and total tree species richness found in the present survey were much higher than that of the previous survey. The numbers of species in the survey carried out by Alvim and Pereira (1965), however, was greatly underestimated due to difficulties encountered in species identification, and those authors considered as a single species all the taxa of some very rich genera (such as Ficus spp., Nectandra spp. and Ocotea spp.).
Of the 24 major species (or species groups) found by Alvim and Pereira (1965), 16 were also among the most important species in our current survey, and only the early secondary species Apeiba tibourbou Aubl. was not encountered by us. The observed decrease in density of this species, which is used by farmers to make rafts, may be attributed to excessive logging. Most species (17) increased in density between surveys. The largest increase was observed for the exotic species A. heterophyllus, which increased from 3.3 to 16.8 ind ha−1 (Table 4). Among the 24 major species, the total density of exotics increased from 9.1 to 28.6 ind ha−1 and of natives from 19.8 to 39.8 ind ha−1. Among the native species, the early secondary class demonstrated the largest increase in total density (from 10.5 to 20.8 ind ha−1), followed by the late secondary class (from 6.4 to 14.1 ind ha−1), and pioneers (from 1.0 to 3.6 ind ha−1); the climax species class was the only one that demonstrated any reduction in density (from 1.9 to 1.2 ind ha−1).
Management of the tree component and farmers’ preferences
Most farmers (78%) reported they managed the natural regeneration of trees in the plantation through selective weeding, with the young seedlings of some species that naturally germinated in the plantation being spared during weeding so that they could occupy gaps and progressively restore shading. A total of 41 species recognized and selected during weeding were mentioned (among 302 citations). Most of these species were native (31), of which 14 were climax species (Fig. 5a). The greatest percentage of citations mentioned late secondary species (64%), with Cedrela odorata (rose-colored cedar) (23%), Plathymenia foliolosa (13%) and Cariniana legalis (rose-colored jequitibá) (12%) being the most frequently cited species. The most cited exotic species were Spondias mombin (4%), Erythrina fusca (“eritrina-da-baixa”) (3%) and A. heterophyllus (3%). Erythrina species found in cabrucas are exotic nitrogen-fixing legume trees introduced in the region by CEPLAC to serve as a monospecific shade for cocoa.

Numbers of species cited and percentages of citations of native and exotic shade trees for each of the main management practices adopted and individual preferences of farmers in cabruca agroforests in southern Bahia, Brazil. a selection of seedlings spared during weeding to allow their natural regeneration. b Removal of mature trees for shade thinning. c Planting of trees. d Preferred species to keep the mature trees in the cabrucas. Successional status of native trees: pioneer (Pi), early secondary (ES), late secondary (LS), climax (Cl)
About half of the farmers surveyed (54%) classified the level of shade in their plantations as good, while 46% judged it to be excessive and less than 1% considered it to be low. Reducing shade by deliberately removing some established trees was reported by 63% of the farmers. The farmers mentioned a total of 34 species (in 246 citations) that they preferably cut during thinning. Most citations referred to exotic species (36%), pioneers (31%), and early secondary species (28%) (Fig. 5b). The most frequently mentioned species were A. heterophyllus (20%) and Erythrina fusca (13%). The respondents justified the removal of A. heterophyllus by alleging that this species has a low canopy and provides very dense shade that hinders cocoa development and production. The respondents also stated that Erythrina fusca has weak wood that would often falls and damages their cocoa trees, has spines on their branches, and that this species would significantly reduce soil humidity levels.
Less than half of the respondents (43%) stated that they planted trees in their cocoa plantations. A total of 36 planted species were mentioned by farmers (107 citations). The greatest number of species cited (19) (and the most citations, 64%) were exotic species (Fig. 5c), especially Spondias mombin (15%), Erythrina fusca (11%), and Hevea brasiliensis (rubber tree) (9%). Among the native species, the most cited was Caesalpinia echinata Lam. (Brazil wood) (8%). The emphasis on planting this latter threatened climax species is probably related to the “Pau-Brasil Program”, a conservation project developed by CEPLAC that distributes seedlings of this species to farmers. In terms of the uses of the planted species, the highest numbers of citations referred to fruit trees (46.3%), followed by timber species (25.4%) and nitrogen-fixing legumes (mainly species of Erythrina spp. and Inga spp.) (16.4%).
When asked about the three species they would most prefer to keep in the cabruca if all the other trees had to be eliminated, farmers mentioned a total of 45 species, 31 native and 14 exotic (in 342 citations). Among the native species, the largest numbers of species mentioned were climax species (14) (Fig. 5d). The highest percentages of citations concerned late secondary species (58%), especially timber species, with the most cited species being Cariniana legalis (17% of the interviewees), Cedrela odorata (15%) and Plathymenia foliolosa (11%). The next most cited species were the exotic fruit species Spondias mombin (10%) and A. heterophyllus (5%), and the nitrogen-fixing legume Erythrina fusca (6%).
There was no significant difference in the frequency of adoption of the management practices between classes of land tenure, farm size and function of the person responding the questionnaire. However, among the most cited species we found a higher frequency of citation in the practice of saving seedlings of Cedrela odorata and Plathymenia foliolosa during weeding among very small and small farms in relation to medium and large farms (χ2, p < 0.05). For classes of land tenure and function of the person responding the questionnaire there was no significant difference in the frequency of citation of species for all management practices analyzed.
Species uses
In addition to shading the cocoa plantations, the main uses of trees were for food, timber, firewood, and medicinal applications (Online Resource 3). Farmers mentioned 35 species used for food, in 505 citations. The greatest numbers of species (24) and percentages of citations (93%) concerned exotic species. The most-cited species were A. heterophyllus (30%), Citrus sinensis (orange tree) (16%), and Spondias mombin (10%).
For timber, farmers mentioned 46 species in 218 citations. Most of the species were climax species (24). The late secondary class had the highest percentage of citations (57%), and the most-cited species were Plathymenia foliolosa (13%) and Nectandra sp. (soap-laurel) (13%). In terms of firewood, 53 species were mentioned in 215 citations. Many were climax species (14), but the vast majority of the citations were of early secondary species (98%). The most-cited species were Inga spp. (16%) and Senna multijuga (cobi) (15%), two fast-growing trees very common in cabrucas that the farmers consider good fuel wood species.
Thirty-six species were mentioned for medicinal applications, in 170 citations. Although many exotic species were mentioned (14), the highest percentage of citations referred to climax species (57%). The most-cited species were Hymenaea oblongifolia (“jatobá”) (23%), Pradosia glaziovii (“buranhém”) (14%) and Gallesia integrifolia (“pau-d’alho”) (8%). Farmers mentioned more than one use for 34% of the species. The most-cited species for multiple uses was H. oblongifolia (medicine, food and timber), A. heterophyllus (food, firewood and timber) and Inga spp. (firewood, food and timber).
Only 29% of farmers said they commercialized some other product from their cabrucas in addition to cocoa. The most frequently cited products were bananas, with 34 citations. Banana plants are used to shade young cocoa trees and are very common in cabrucas. However, since it is not a tree its density was not estimated in our survey. Among the shade trees, 14 species were mentioned in 38 citations; all of them were exotic, and the majority was fruit trees, especially Spondias mombin (42%) and A. heterophyllus (16%).
Discussion
Cabrucas and forest comparisons
The cabrucas surveyed showed a high diversity of shade trees for an agroforestry system, which is a common feature in traditional cocoa agroforests. Sambuichi and Haridasan (2007) assessed 15 ha in five cabruca farms in Southern Bahia, with different ages and degrees of abandonment of management practices, and found 293 species (DBH ≥ 10 cm), with diversity (Shannon) ranging from 3.31 to 4.22 in plots of 3.0 ha. Rolim and Chiarello (2004) found 105 species in cabrucas of the Northeastern region of Espirito Santo state, Brazil, by sampling trees with DBH ≥ 5 cm in 4.8 ha of 20 farms. In Nigeria, Oke and Odebiyi (2007) found 45 species (DBH ≥ 10 cm) in an inventory of 21 ha in three cocoa farms. Sonwa et al. (2007), studying the dense and complex agroforests of Southeast Cameroon, sampled trees and pseudo-trees (like banana) with DBH ≥ 2.5 cm and found 206 species in 9.1 ha surveyed in 60 cocoa farms, with diversity indices (Shannon) ranging between 3.1 and 4.2 per agroforest. The results of these studies, however, are not directly comparable with ours due to differences in survey methodologies employed.
The high diversity of trees in cabrucas in southern Bahia is a reflection of the high natural diversity of trees in that region. Many of the shade trees in cabrucas are remnants of original forests that were thinned to plant cocoa, while others may have regenerated from seeds that came from surrounding forest patches. Thomas et al. (2008) encountered 264 species and 988 individual trees with DBH ≥ 10 cm in a 1 ha forest plot in this same region. It must however be noted that the average richness of tree species in our survey was only 13.6% of that found in this natural forest, and the average richness of forest specialist species was only 7.3%, indicating the significant loss of tree species richness during the conversion of forests into cabrucas. This loss is partly the result of the intense thinning of trees (with the average density observed for cabrucas being only 12.2% of that observed in the intact forest). Studies on other traditional cocoa agroforests have also shown intense reductions of total tree richness resulting from the conversion of native forests to agroforests (Oke and Odebiyi 2007; Asase and Tetteh 2010; Wade et al. 2010).
Besides harboring far fewer tree species than intact forests, cabrucas also demonstrate different species compositions, with relatively higher proportions of early successional trees and exotic species. Important tree families in the Atlantic Forest (such as Myrtaceae, Sapotaceae and Chrysobalanaceae) that comprise many endemic and threatened climax species are less common in cabrucas (Sambuichi and Haridasan 2007). Also, cabrucas do not provide habitat for small trees and understory species, as these are eliminated to make room for the cocoa crop. These results make it clear that cabrucas are not substitutes for intact forests in terms of their conservation value, and that replacing forests with cabrucas is not appropriate for conservation purposes. However, the presence of cabrucas within the landscape can favor the conservation of species occupying forest remnants by acting as permeable matrixes, additional habitats, and buffer zones (Cassano et al. 2009; Pardini et al. 2009). The value of cabrucas for species conservation therefore depends on the quantity and quality of forest remnants in the landscape. Faria et al. (2007), working with assemblages of ferns, frogs lizards, birds and bats, demonstrated that cabrucas in landscapes with less forest areas were more biologically impoverished than cabrucas in landscapes with greater forest cover.
The basal area values obtained for cabrucas were also high for cultivated systems, and indicated that cabrucas retain considerable biomass. Other studies showed basal area of non-cocoa trees in cabrucas ranging from 11.8 to 28.2 m2 ha−1 (Sambuichi 2006). Comparing these values with those of 46.3 m2 ha−1 (Mori et al. 1983) and 39.6 m2 ha−1 (Thomas et al. 2008) encountered in surveys of native forests, it can be seen that the reductions in BA from forests to cabrucas are much smaller than reductions in richness. This is due to the fact that thinning primarily affects the smallest trees, while a higher proportion of large trees are retained. Cabrucas, of course, also contain cocoa trees, although their basal area was not quantified in the present study. This data suggests that cabrucas may be interesting areas for carbon storage, and additional studies to quantify their carbon stocks will be needed (Sambuichi 2006).
Density changes, diversity and shade management
To evaluate the role of cabrucas in tree species conservation it is important to determine not only their current species composition but also what is happening in these agroecosystems over time and identify the factors affecting their species composition. The conversion of forests to shaded plantations is often only one stage in the process of their degradation, and the intensification of these agricultural practices can lead to reductions of shading in agroforests until they are totally replaced by unshaded monocultures (Ruf and Schroth 2004; Perfecto and Vandermeer 2008). The establishment of cabrucas in the cocoa-growing region of southern Bahia was initiated a long time ago, however, and cabrucas continue to be the main component of the landscape in spite of various economic crises and efforts to intensify their management (Alger and Caldas 1994).
Alvim and Pereira (1965) performed the first survey of non-cocoa trees in cabrucas and concluded that there was too much shade in the plantations; these authors recommended thinning to reduce the number of trees to about 25 shade trees per hectare. In spite of the efforts of governmental agricultural agencies to promote the removal of trees to reduce shade levels, we found higher average densities of trees in cabrucas than Alvim and Pereira (1965) did more than 45 years earlier. In addition to the resistance of farmers to adhere to thinning recommendations (Johns 1999), this higher shade tree density can be explained by the reduction of the weeding frequency in many farms (which allowed more tree regeneration) after the last cocoa crisis. Unlike the situation in Africa and Indonesia where cocoa production is dominated by small landholders farming on less than 20 ha areas (Ruf and Schroth 2004), most farms in southern Bahia are larger than this, and the landowners often have other outside income sources. Thus when crises occur, workers are laid off and management efforts (including weeding) are partially abandoned.
We found more species in our 16 ha survey than Alvim and Pereira (1965) encountered in 61 ha of cabrucas. Although the number of species in this older survey may be an underestimate due to incomplete species identifications, this increase in richness may also reflect increased tree density. We observed a strong positive correlation between species richness and density in cabrucas, as was also observed by López-Gómez et al. (2008) in complex agroforests of shaded coffee in Mexico. The logarithmic increase in species richness with increasing numbers of trees is a common pattern in natural forests but is not commonly observed in planned agroforestry systems, as the numbers of species are usually more closely determined by human choice (Perfecto and Vandermeer 2008). This unplanned diversity seen in cabrucas not just reflects the presence of the remnant individuals from the original thinned-out forest, but probably results mainly from the natural regeneration of trees influenced by the diversity of adjacent forest remnants.
Management practices are currently being revived on cocoa farms, and agronomists are again recommending reducing shade levels to increase cocoa yields. Most of the farmers interviewed reported removing trees to reduce the level of shade in their cabrucas. The richness/density relationship observed in the present study indicates that reducing density by thinning will lead to tree richness losses in the cabrucas. However, these losses may not be very significant for forest specialist species because denser areas tend to have more open environment tree species, and farmers preferentially cut these trees (and exotics) during thinning. In addition, current Brazilian legislation prohibits cutting forest trees even in cabrucas, which may inhibit farmers from removing them.
Unlike species richness, species diversity showed no significant correlation with tree density in the present study. This was because diversity is composed of two components: richness and evenness (Magurran 1988). In the case of the cabrucas, evenness showed the opposite trend of richness—and tended to increase as density decreased (i.e., the denser areas showed higher relative abundances of just a few species, while the more intensely thinned areas showed more equitable distributions of individual species). The cabrucas with fewer trees had higher numbers of species per individual plant, in some cases even higher than natural forests. We found the average number of species per individual in the 1.0 ha plots (0.30) to be slightly higher than that found by Thomas et al. (1998) in a native forest (0.27). For this reason, even with the intense loss in species richness caused by thinning, cabrucas still have high levels of shade tree diversity, with Shannon index values comparable to areas of natural forest (Sambuichi and Haridasan 2007).
The diversity of tree species in agroforests is important for conservation purposes because it favors associated diversity, and can influence the ability of these agroecosystems to function as a permeable matrix and facilitate the flux of pollinators and seed dispersers (Perfecto and Vandermeer 2008). Agroforests also contribute to biodiversity conservation by promoting heterogeneity at the landscape level because different farms usually have significantly different tree species compositions (Bhagwat et al. 2008). Apart from the high diversity found within each cabruca (alpha diversity), we found high diversity among the different cabrucas (beta diversity)—which has also been reported in other cabruca surveys in the region (Sambuichi 2006; Sambuichi and Haridasan 2007). The low floristic similarities among cabrucas may partly reflect the spatial heterogeneity of the original forest due to local differences in soil and climate and the tree population dynamics (Sambuichi 2006). In addition, historic differences in management practices and the preferences of individual farmers may also have influenced the low similarity among these agroforests.
The cabrucas studied were also very diverse in tree vegetation structure. Most farms showed different tree diameter distributions, and there were large variations in tree densities among the farms. Other studies of the tree components of cabrucas in southern Bahia have also reported these structural variations (Alvim and Pereira 1965; Sambuichi 2002, 2006; Sambuichi and Haridasan 2007). Density variations were strongly negatively correlated with differences in the sizes of individuals. This pattern was expected because areas with larger trees require fewer individuals to maintain the same shade levels. On the other hand, the densities and BA on the farms were not correlated, due to differences in the diameter distributions of the trees. These differences in tree sizes and densities among cabrucas are probably related to historical differences in their establishment, management intensities, and the regeneration stages of their arboreal components. These results indicate that recommendations for thinning the shade based only on the density of shade trees are not well-founded.
Species status, management and species preferences
Most of tree species found in the present survey were native, especially the climax and late secondary species, as was observed in other surveys in the region (Sambuichi 2002, 2006; Sambuichi and Haridasan 2007). Nonetheless, there were greater numbers of open environment and exotic trees than forest specialists, indicating that the floristic composition of cabrucas is changing over time and that the remnant trees of the original forests are gradually being substituted. In comparing old and new cabruca areas, Sambuichi and Haridasan (2007) found higher proportions of exotic trees in the older areas. In their study of cabrucas in Espirito Santo State, Rolim and Chiarello (2004) found a predominance of early successional stage species and warned of the gradual replacement of native forest species in these areas.
The presence of exotic and open environment species among the main tree species in cabruca is not a recent feature, however. The 24 main species found in the survey undertaken by Alvim and Pereira (1965) included more early secondary and exotic species than climax species (Table 4). Additionally, most of the main tree species in this previous survey were also among the main tree species in our current survey, indicating that the composition of main tree species of cabrucas had not experienced major changes over the past 45 years. These results indicate that the cabrucas were already very different from the original thinned forests at the time of this first survey, and that many of the original trees of these forests apparently had already been replaced by other species. Surprisingly, cabrucas retain even today higher numbers of forest specialist species than other species. This suggests that, despite the dominance of exotic and open environment trees, cabrucas can have the ability to maintain at least part of their richness in forest specialist species over time.
The existence of nearby intact forest remnants and the high diversity and permeability of the landscape matrix favor the supply and dispersal of seeds of forest species and may partly explain the permanence of these species in cabrucas. However, the establishment of these trees in these areas depends on agro-environmental conditions, which are mainly determined by the management practices applied. Therefore, to better understand the dynamics of tree cover in cabrucas it is necessary to analyze farmers’ practices and preferences for different ecological groups of trees. Farmers generally preferred the species they believed to provide good shade for cocoa production and the species they use for purposes other than shade. We observed that most of the species listed as preferred by farmers were also among the most cited especially for use as timber and food. Moreover, the practice of saving seedlings of the two most cited forest species was more frequent on very small and small farms, suggesting that smallholders, which commonly know and utilize more agroforestry products, might be more likely to apply practices to conserve useful forest species in cabrucas.
An important change that was observed between the past and current surveys was a great increase in the density of the exotic species A. heterophyllus, Erythrina spp. and Spondias mombin. These trees were among the main species found in many surveys of cabrucas in southern Bahia and Espirito Santo states (Alvim and Pereira 1965; Sambuichi 2002; Rolim and Chiarello 2004; Sambuichi and Haridasan 2007). These are species that regenerate very well in cabrucas (especially A. heterophyllus), and their expressive increases in density may be related to reductions in weeding frequencies due to the cocoa crisis. Additionally, the seedlings of some of these species are selected by rural workers who consume their fruits or are preferentially planted for shade. Many landowners, however, do not like to use A. heterophyllus and Erythrina spp. as shade tree and remove them during thinning for shade because they believe these trees are prejudicial to cocoa production. It should therefore be expected that the densities of these exotic trees will decrease in cabrucas with the resumption of active management practices and the reduction of existing shade levels.
We also observed increases in the densities of open environment species between surveys, especially in relation to early secondary species. The high numbers of open environment species in cabrucas cannot be explained by the preference of the farmers, for they generally avoid these plants (especially the pioneer species), and eliminate them during selective weeding and when reducing shade levels. Early secondary species are less rejected than pioneers because they can be used as firewood. The principal advantage of these species to the farmers is their rapid growth, and they will sometimes allow their temporary establishment in open spaces where there is a more immediate need for shade. However, we found a positive correlation between the proportions of open environment species and tree densities in cabrucas, indicating that these species were principally regenerating in denser areas. This reinforces the view that a dense presence of these species in cabrucas occurs mostly because of a partial or total abandonment of weeding. This same situation was observed by Sambuichi and Haridasan (2007) during their comparisons of managed and abandoned cabruca areas in the municipality of Ilhéus. The densities of these open environment species likewise tend to decrease with the resumption of active management practices.
Most of the species that the farmers preferred to keep in cabrucas were forest specialist. Farmers tend to value these species and adopt active measures to maintain them in the plantations. Besides avoiding cutting these trees when thinning for shade, the farmers will also spare their seedlings during weeding activities. Their appreciation for these species is mainly due to their longevity and considerable height. Farmers prefer more resistant trees that will not easily fall and damage their cocoa plants, and these trees are also taller and give sparser shade, thereby allowing higher cocoa productivity and reducing the incidence of cocoa diseases. Late secondary species have an advantage over climax species in that they regenerate better in cabrucas and grow faster. All of the late secondary species on the list prepared by Alvim and Pereira (1965) showed higher densities in the present survey.
The greatest increase in density among the late secondary species was recorded for Plathymenia foliolosa (Table 4), and this species also demonstrated the highest density, basal area, and IV among the native species in our survey. Interestingly, this species was not found in many other cabruca surveys in southern Bahia (Sambuichi 2002, 2006; Sambuichi and Haridasan 2007) because it does not occur in the wetter areas along the coast where most of the earlier cabruca inventories were concentrated. Nonetheless, more extensive studies as Alvim and Pereira (1965) and the current study demonstrated that this species is regionally important and highly appreciated for the quality of its wood. These results reinforce the importance of undertaking more extensive studies in cabrucas to access the present composition of these agroforests (Sambuichi 2006).
Our results indicated that cabrucas can serve as additional habitat for many forest specialist tree species. Late secondary trees like Cariniana legalis, Cariniana estrellensis, Cedrela odorata, and Plathymenia foliolosa have expressive populations in cabrucas and new trees have become established in these agrosystems, with increases in their densities between the two surveys. Farmers can usually recognize their seedlings and spare them during weeding procedures. These are large trees that usually have low densities of mature individuals in forest remnants and often fall victim to illegal logging (R.H.R. Sambuichi, personal observations). Therefore, the existing populations of these species in cabrucas constitute an important bank of trees that can provide pollen and seeds to enhance genetic diversity in forest fragments and restoring degraded lands (Sambuichi and Haridasan 2007).
Cabrucas may not be suitable habitats, however, for most shade tolerant climax tree species, and the high percentages of climax species observed in cabrucas does not mean that all of these species will persist in these areas over time. Among the main species found in the past and in the current surveys, the climax group was the only one that did not increase in density. Despite the fact that farmers indicated that they preferred to keep many climax species in their cabrucas, the number of species that they are able to recognize and select may be relatively low in relation to the immense richness of species in southern Bahia. Farmers usually prefer large trees and timber species, and cannot recognize most of the rare and non-useful taxa. Furthermore, the slow growth of these species makes their seedlings more vulnerable to elimination during successive weeding operations. These results are worrying because most of the threatened tree species belong to this successional group, and there are indications that tropical shade tolerant trees are likely to be the most vulnerable species group to changes in the landscape (Pardini et al. 2009), so cabrucas may be losing important tree richness over time.
To minimize this loss of species richness, programs could be developed by governmental and non-governmental agencies to distribute seedlings and otherwise promote the planting and conservation of threatened tree species in cabrucas. The “Pau-Brasil Program” demonstrated that this type of strategy could be successful (especially for climax timber species that farmers are interested in planting for future harvesting). This could be a very important strategy to avoid the extinction of threatened timber species such as the endemic species of Manilkara spp. (“maçaranduba”) which suffer from illegal logging in forest remnants.
Strategies for increasing profitability and conservation implications
In order to increase their income, farmers strive to increase cocoa production by intensifying land management. Several studies have shown that intensification of agricultural management to increase production leads to biodiversity reductions in agroforestry systems (García-Fernández et al. 2003; Steffan-Dewenter et al. 2007; Hervé and Vidal 2008). Among the different management practices used to enhance cocoa productivity, the one that most directly affects the tree component is the thinning of shade trees. Our results indicate that this practice leads to losses of tree species richness (although current thinning practices are mostly affecting exotic and pioneer species). One manner to avoid richness losses is to reduce shade by simply pruning tree branches, thus raising the canopies without cutting the trees. Another important long-term strategy to avoid tree density reductions would be genetic improvement of cocoa trees and the selection of genetic varieties that produce well under high-shade conditions.
Another way to increase the profitability of the cabrucas would be to diversify their production (Leakey and Tchoundjeu 2001), and this could be done by increasing the economic utilization of their shade tree components. The commercialization of these secondary products is apparently hampered by a lack of ready markets and low production volumes. In order to promote this trade, however, it would be necessary to increase the numbers of trees of the harvested species (Looy et al. 2008). This practice may lead to reductions in tree species diversity and increases in the densities and frequencies of exotic tree species growing in cabrucas, which would not be positive for conservation purposes.
Timber extraction is another alternative strategy for diversifying regional income. This timber harvesting would focus mainly on late secondary timber species that regenerate well under cabrucas conditions. Additional studies on the population dynamics of these trees in cabrucas will be necessary, however, to adapt their management techniques to minimize impacts on biodiversity conservation. Current Brazilian legislation does not permit the cutting and commercialization of native trees in the Atlantic Forest region (to avoid deforestation and the depletion of forests and cabrucas by illegal logging) and it would be necessary to alter the current laws in order to exploit native woods in these agroforestry systems. A viable short-term option is the use of fallen timber, which can be legally harvested and could be used in special craft applications.
Perhaps the best way to increase regional incomes and promote biodiversity conservation would be through “environmental certification” to obtain better prices for biodiversity-friendly cocoa (Steffan-Dewenter et al. 2007; Cassano et al. 2009). This strategy could stimulate cocoa farmers to adopt more biodiversity-friendly management practices and could help reduce the economic impacts caused by fluctuations of cocoa prices. Environmental certification could also stimulate the farmers to comply with the law that determines that all private farmlands within the Atlantic Forest region must maintain 20% of their total area (as well as riparian zones) as protected reserves (Sparovek et al. 2010), contributing for increase the amount of protected forest in the region.
Conservation value of cabrucas and concluding remarks
The findings of the present investigation were consistent with those of other complex cocoa agroforests (Bhagwat et al. 2008; Asase and Tetteh 2010) and demonstrated that cabrucas have an important role in the conservation of native forest tree species. Although they are poor substitutes for undisturbed forests in terms of tree species richness, these agroforests contribute to heterogeneity at the landscape level and thus favor biodiversity conservation. Additionally, due to the high diversity of their shade tree component, cabrucas can function as ecological corridors, avoiding the isolation of plant and animal species in forest fragments. These agroforests can also provide additional habitat for some forest tree species and reduce anthropogenic pressure on forests remnants by providing firewood and timber to meet the needs of rural families. Furthermore, cabrucas can be valuable sources of seeds for enriching Atlantic Forest remnants and restoring degraded lands.
All of these results, however, reinforce the indication that the value of cabrucas for biodiversity conservation is additional and dependent on the quantity and quality of forest remnants in the landscape (Faria et al. 2006, 2007), so it is very important to increase the amount of protected forest in the region. Moreover, the maintenance of cabrucas in southern Bahia depends on the economic sustainability of the farms, and the strategies adopted to increase the profitability of these agroecosystems can affect their conservation value, therefore it is also important to find and promote strategies that can better reconcile conservation and profitability.
Abbreviations
- CEPLAC:
-
Executive Committee for Cocoa Farming Plan
- CEPEC:
-
Cocoa Research Center
- DBH:
-
Diameter at breast height
- IV:
-
Importance value
- UESC:
-
State University of Santa Cruz
References
Alger K, Caldas M (1994) The declining cocoa economy and the Atlantic Forest of Southern Bahia, Brazil: conservation attitudes of cocoa planters. Environmentalist 14:107–119. doi:10.1007/BF01901304
Alvim P de T, Pereira CP (1965) Sombra e espaçamento nas plantações de cacau no Estado da Bahia. In: Relatório Anual do CEPEC 1964. CEPLAC, Ilhéus, pp. 18–19
Asase A, Tetteh DA (2010) The role of complex agroforestry systems in the conservation of forest tree diversity and structure in southeastern Ghana. Agrofor Syst 79:355–368. doi:10.1007/s10457-010-9311-1
Bhagwat SA, Willis KJ, Birks HJB et al (2008) Agroforestry: a refuge for tropical biodiversity. Trends Ecol Evol 23:261–267. doi:10.1016/j.tree.2008.10.005
Budowski G (1965) Distribution of tropical American rainforest species in the light of successional processes. Turrialba 15:40–42
Cassano CR, Schroth G, Faria D et al (2009) Landscape and farm scale management to enhance biodiversity conservation in the cocoa producing region of southern Bahia, Brazil. Biodivers Conserv 18:577–603. doi:10.1007/s10531-008-9526-x
Curtis JT, McIntosh RP (1951) An upland forest continuum in the prairie-forest border region of Wisconsin. Ecology 32:476–496. doi:10.2307/1931725
Delabie JHC, Jahyny B, Nascimento IC et al (2007) Contribution of cocoa plantations to the conservation of native ants (Insecta: Hymenoptera: Formicidae) with a special emphasis on the Atlantic Forest fauna of southern Bahia, Brazil. Biodivers Conserv 16:2359–2384. doi:10.1007/s10531-007-9190-6
Faria D, Laps RR, Baumgarten J et al (2006) Bat and bird assemblages from forests and shade cacao plantations in two contrasting landscapes in the Atlantic rainforest of southern Bahia, Brazil. Biodivers Conserv 15:587–612. doi:10.1007/s10531-005-2089-1
Faria D, Paciencia MLB, Dixo M et al (2007) Ferns, frogs, lizards, birds and bats in forest fragments and shade cacao plantations in two contrasting landscapes in the Atlantic forest, Brazil. Biodivers Conserv 16:2335–2357. doi:10.1007/s10531-007-9189-z
Fundação SOS Mata Atlântica, Instituto Nacional de Pesquisas Espaciais (2008) Atlas dos remanescentes florestais da Mata Atlântica período 2000–2005. Fundação SOS Mata Atlântica/INPE, São Paulo
García-Fernández C, Casado MA, Pérez MR (2003) Benzoin gardens in North Sumatra, Indonesia: effects of management on tree diversity. Conserv Biol 17(3):829–836. doi:10.1046/j.1523-1739.2003.01487.x
Geist HJ, Lambin EF (2002) Proximate causes and underlying driving forces of tropical deforestation. Bioscience 52:143–150. doi:10.1641/0006-3568(2002)052[0143:PCAUDF]2.0.CO;2
Gordon C, Manson R, Sundberg J et al (2007) Biodiversity, profitability, and vegetation structure in a Mexican coffee agroecosystem. Agric Ecosyst Environ 118:256–266. doi:10.1016/j.agee.2006.05.023
Hervé BDB, Vidal S (2008) Plant biodiversity and vegetation structure in traditional cocoa forest gardens in southern Cameroon under different management. Biodivers Conserv 17:1821–1835. doi:10.1007/s10531-007-9276-1
Johns ND (1999) Conservation in Brazil’s chocolate forest: the unlikely persistence of the traditional cocoa agroecosystem. Environ Manage 23:31–47. doi:10.1007/s002679900166
Jose S (2009) Agroforestry for ecosystem services and environmental benefits: an overview. Agrofor Syst 76:1–10. doi:10.1007/s10457-009-9229-7
Leakey RRB, Tchoundjeu Z (2001) Diversification of tree crops: domestication of companion crops for poverty reduction and environmental services. Expl Agric 37:279–296. doi:10.1017/S0014479701003015
Looy TV, Carrero GO, Mathijs E et al (2008) Underutilized agroforestry food products in Amazonas (Venezuela): a market chain analysis. Agrofor Syst 74:127–141. doi:10.1007/s10457-008-9110-0
López-Gómez AM, Williams-Linera G, Manson RH (2008) Tree species diversity and vegetation structure in shade coffee farms in Veracruz, Mexico. Agric Ecosyst Environ 124:160–172. doi:10.1016/j.agee.2007.09.008
Magurran AE (1988) Ecological diversity and its measurements. Croom Helm, London
Martini AMZ, Fiaschi P, Amorim AMA et al (2007) A hot-point within a hot-spot: a high diversity site in Brazil’s Atlantic Forest. Biodivers Conserv 6(3111–3128):3128. doi:10.1007/s10531-007-9166-6
Méndez VE, Shapiro EN, Gilbert GS (2009) Cooperative management and its effects on shade tree diversity, soil properties and ecosystem services of coffee plantations in western El Salvador. Agrofor Syst 76:111–126. doi:10.1007/s10457-009-9220-3
Mori SA, Boom BM, Carvalho AMV et al (1983) Southern Bahian moist forests. Bot Rev 49:155–232. doi:10.1007/BF02861011
Mueller-Dumbois D, Ellenberg H (1974) Aims and methods of vegetation ecology. Wiley, New York
Myers N, Mittermeier RA, Mittermeier CG et al (2000) Biodiversity hotspots for conservation priorities. Nature 403:853–858. doi:10.1038/35002501
Oke DO, Odebiyi KA (2007) Traditional cocoa-based agroforestry and forest species conservation in Ondo State, Nigeria. Agric Ecosyst Environ 122:305–311. doi:10.1016/j.agee.2007.01.022
Pardini R, Faria D, Accacio GM et al (2009) The challenge of maintaining Atlantic forest biodiversity: a multi-taxa conservation assessment of specialist and generalist species in an agro-forestry mosaic in southern Bahia. Biol Conserv 142:1178–1190. doi:10.1016/j.biocon.2009.02.010
Perfecto I, Vandermeer J (2008) Biodiversity conservation in tropical agroecosytems: a new conservation paradigm. Ann N Y Acad Sci 1134:173–200. doi:101196/annals.1439.011
Rice RA, Greenberg R (2000) Cacao cultivation and the conservation of biological diversity. Ambio 29:167–173. doi:10.1579/0044-7447-29.3.167
Rolim SG, Chiarello AG (2004) Slow death of Atlantic forest trees in cocoa agroforestry in southeastern Brazil. Biodivers Conserv 13:2679–2694. doi:10.1007/s10531-004-2142-5
Ruf F, Schroth G (2004) Chocolate forests and monocultures: a historical review of cocoa growing and its conflicting role in tropical deforestation and forest conservation. In: Schroth G, da Fonseca GAB, Harvey CA et al (eds) Agroforestry and biodiversity conservation in tropical landscapes. Island Press, Washington, pp 107–134
Saatchi S, Agosti D, Alger K et al (2001) Examining fragmentation and loss of primary forest in the southern Bahian Atlantic Forest of Brazil with radar imagery. Conserv Biol 15:867–875. doi:10.1046/j.1523-1739.2001.015004867.x
Sambuichi RHR (2002) Fitossociologia e diversidade de espécies arbóreas em cabruca (Mata Atlântica raleada sobre plantação de cacau) na região sul da Bahia, Brasil. Acta Bot Bras 16:89–101. doi:10.1590/S0102-33062002000100011
Sambuichi RHR (2006) Estrutura e dinâmica do componente arbóreo em área de cabruca na região cacaueira do sul da Bahia, Brasil. Acta Bot Bras 20:943–954. doi:10.1590/S0102-33062006000400018
Sambuichi RHR, Haridasan M (2007) Recovery of species richness and conservation of native Atlantic forest trees in the cacao plantations of southern Bahia in Brazil. Biodivers Conserv 16:3681–3701. doi:10.1007/s10531-006-9017-x
Sambuichi RHR, de Oliveira RM, Mariano Neto E et al (2008) Conservation status of ten endemic trees from the Atlantic Forest in the south of Bahia—Brazil. Nat Conserv 6:208–225
Schroth G, Lehmann J, Rodrigues MRL et al (2001) Plant–soil interactions in multistrata agroforestry in the humid tropics. Agrofor Syst 53:85–102. doi:10.1023/A:1013360000633
Schroth G, Harvey CA, Vincent G (2004) Complex agroforests: their structure, diversity and potential role in landscape conservation. In: Schroth G, da Fonseca GAB, Harvey CA et al (eds) Agroforestry and biodiversity conservation in tropical landscapes. Island Press, Washington, pp 227–260
Sonwa DJ, Nkongmeneck BA, Weise SF et al (2007) Diversity of plants in cocoa agroforests in the humid forest zone of Southern Cameroon. Biodivers Conserv 16:2385–2400. doi:10.1007/s10531-007-9187-1
Sparovek G, Berndes G, Klug ILF et al (2010) Brazilian agriculture and environmental legislation: status and future challenges. Environ Sci Technol 44:6046–6053. doi:10.1021/es1007824
Steffan-Dewenter I, Kessler M, Barkmann J et al (2007) Tradeoffs between income, biodiversity, and ecosystem functioning during tropical rainforest conversion and agroforestry intensification. Proc Natl Acad Sci USA 104:4973–4978. doi:10.1073/pnas.0608409104
Thomas WW, de Carvalho AMV, Amorim AMA et al (1998) Plant endemism in two forests in southern Bahia, Brazil. Biodivers Conserv 7:311–322. doi:10.1023/A:1008825627656
Thomas WW, de Carvalho AMV, Amorim AMA et al (2008) Diversity of woody plants in the Atlantic coastal forest of southern Bahia, Brazil. In: Thomas WW (ed) The Atlantic coastal forest of northeastern Brazil. The York Botanical Garden Press, New York, pp 21–66
Vandermeer J, Perfecto I (2007) The agricultural matrix and a future paradigm for conservation. Conserv Biol 21:274–277. doi:10.1111/j.1523-1739.2006.00582.x
Wade ASI, Asase A, Hadley P et al (2010) Management strategies for maximizing carbon storage and tree species diversity in cocoa-growing landscapes. Agric Ecosyst Environ 138:324–334. doi:10.1016/j.agee.2010.06.007
Acknowledgments
We are thankful to the Dutch Buffer Stock Fund (Dutch Ministry of Economic Affairs, Agriculture and Innovation), the Instituto Cabruca, and the Universidade Estadual de Santa Cruz for their financial support; the CEPEC/CEPLAC and HUESC herbaria for help in identifying the tree species; and to all those who collaborated with this project. We thank Dr. G. Schroth and Dr. C.D. Foy for their comments on an earlier version of this paper.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article
Cite this article
Sambuichi, R.H.R., Vidal, D.B., Piasentin, F.B. et al. Cabruca agroforests in southern Bahia, Brazil: tree component, management practices and tree species conservation. Biodivers Conserv 21, 1055–1077 (2012). https://doi.org/10.1007/s10531-012-0240-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10531-012-0240-3