
Abstract
The cultivation of cocoa is a commodity of great importance worldwide. In Mexico, cocoa is grown in Tabasco and Chiapas states in agroforestry systems since pre-Hispanic times, where cocoa is grown under the canopy of shade trees. Crops such as sugarcane and extensive livestock production have gradually reduced the area devoted to cocoa cultivation in the region. Yet, farmers keep small portions of the cocoa agroforestry systems to maintain the local floristic diversity. This practice seems to be a successful contribution to plant conservation. To determine the management practices used by farmers in the agroforestry cocoa systems and the diversity of products they harvest, a non-probabilistic sampling was carried out in 38 shade cocoa plots of 20 × 50 m each (19 in Tabasco and 19 in Chiapas). We counted and identified all trees with DBH ≥ 5 cm, recording their height. We estimated canopy cover at 20 points within each plot. Our results show no tree species richness differences between states, but there were differences at the municipality level; Pichucalco (Chiapas) had the highest tree species richness. Considering all tree species, there was a higher tree density in Tabasco than in Chiapas. Regarding only cocoa trees, there was also a higher abundance in Tabasco than in Chiapas. In both cases, farmers obtain a wide variety of products for self-consumption and local market sale from their cocoa agroforestry systems (e.g., timber, fruit, and grains). The wide floristic diversity in these agroecosystems aids native plant species conservation and could favor the increase of agroforestry plantations associated with cocoa cultivation.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Cocoa cultivation in Mexico has its origins before the arrival of Europeans, where the Mayan inherited its use from the Olmec culture. Residues of theobromine (formerly known as xantheose; an alkaloid of cacao) found in archeological pots from San Lorenzo Tenochtitlán (Veracruz, Mexico), dated between 1800 and 1000 B.C.E., provided evidence that the Olmecs were the first known culture to use cocoa (Powis et al. 2011). Furthermore, ceramic pots found in San Andrés (Tabasco, Mexico) suggest that the Olmecs also used cocoa as drinking during their main festivities (Seinfeld 2007). Subsequently, during the Late Classic period of Mesoamerica, between 800 and 1100 C.E., the Mayas from the Chontalpa region in Tabasco domesticated the cocoa and the region became an active area of commerce where cacao beans were used as currency (Attolini 2011; Millon 1955; Thompson 1997; West et al. 1985). Additionally, there is evidence that the Chontalpa region (Tabasco) and the Soconusco region (Chiapas) were the most important cocoa producers in Mesoamerica before the arrival of the conquistadors (Bergman 1969; Millon 1955; Thompson 1997; West et al. 1985; Young 2007).
The traditional management of the cocoa agroforestry systems (cocoa-AFS) in Mexico has been characterized by a diverse array of plant species other than cocoa since pre-Hispanic times (Bergman 1968; Gómez-Pompa et al. 1990; Millón 1955; Ogata 2018; Toledo et al. 2007; Young 2007). In words of Gómez-Pompa et al. (1990), “Cacao, therefore, is a shade-tolerant species that is traditionally grown under the shade of other trees, mainly of the nitrogen-fixing Leguminosae family. The prevalent genera and species of Leguminosae used for shade in the cacao orchards of today (Erythrina, Gliricidia, Inga) may in fact be the same ones used in pre-contact times.” According with Millón (1955), the most common shade tree used in cocoa plantations in Mexico and Central America was Gliricidia sepium and in Central America and the West Indies was the Erythrina sp. tree called “Bois inmortal” in Trinidad and “Madre del cacao” in Nicaragua. Young (2007, p. 24) stated that “the discovery of cacao gardens planted by the Mayas in the cenotes of the northern Yucatan Peninsula helps in understanding the domestication of the cocoa tree in the Mesoamerican region. Trees were selected in the rain forest for transplanting into small garden plots, including those of the cenotes. Other economically useful trees grown in the cenotes by the Mayas included Citrus, Cocos, Annona, and Musa, along with nitrogen-fixing tree species.” The management strategies of the cocoa-AFS used by Mesoamerican ancestors are currently believed to prevail in the states of Tabasco and Chiapas, where cocoa is grown in similar ecological conditions that the Mayan indigenous people used to do before the conquest.
The diversified management under the shade of cocoa-AFS provides multiple benefits such as food and non-food resources, shade trees promote nutrient cycling or prevent soil erosion, promote carbon sequestration and reduce greenhouse gas emissions, contribute to mitigating the effects of climate change, and support much higher biodiversity than unshaded systems (Dawoe et al. 2016; Jezeer et al. 2017; Middendorp et al. 2018; Tscharntke et al. 2011). Such agroforestry systems are management units that might include a rich assemblage of tree and plant species (Gliessman 2002), of animal species and yearly and/or perennial crops (Tscharntke et al. 2011). Such systems provide important conservation areas for different native organisms (da Silva et al. 2008). For instance, areas where shade coffee or cocoa with a species-rich canopy is promoted are more effective at protecting both migratory and resident birds (Greenberg et al. 2000; Ibarra et al. 2001; Sunshine et al. 2007), in comparison with sun coffee plantations or grazing fields where diversity generally declines (Greenberg et al. 1997; Philpott et al. 2008; MacGregor-Fors et al. 2018). Regarding economic value for farmers, in Africa, Indonesia, and Latin America, cocoa-AFS have been used as alternatives to improve local people well-being by adding commercial value and elements associated with other plant species, such as fruit trees (e.g., oranges, avocados, dragon fruit, black pepper) and others for timber purposes (e.g., cedar, mahogany) (Yamada and Gholz 2002; Rani et al. 2008; Sonwa et al. 2014; Notaro et al. 2020). Thus, agroecosystems such as those managed in traditional ways often represent integral productive systems due the inclusion of high species richness (Young 2018), representing one of the best productive options for biodiversity conservation (Anglaaere et al. 2011; Bhagwat et al. 2008) while still being economically profitable (Notaro et al. 2020; Useche and Blare 2013).
Among cocoa-AFS, a wide array of management practices exist across the globe, some of which are biodiversity friendly and others that exclude any conservation value in exchange for a higher yield, usually of lower quality produce. Technified sun cocoa-AFS are becoming more economically important in some countries because of their high productivity by using sun-resistant and high yielding varieties, allowing densities of cocoa trees above 1000 trees per hectare (Beer et al. 1998; Bentley et al. 2004; Rueda et al. 2018). This type of management has shown severe long-term ecological damage causing soil degradation, low quality and productivity, and biodiversity loss (Gyau et al. 2014; Andres et al. 2016). Recent studies in Tabasco (southeast Mexico) showed a decreasing trend in the area used for cacao production, with the loss of almost half of the territory (46.4%) in a period of 13 years (Oporto-Peregrino et al. 2019). Some of the most critical components behind this pattern are urban development, the expansion of the monoculture frontier (e.g., bananas, oil), drops in market prices of cocoa beans, and lack of interest of the new generations in cultivating cocoa (Tudela 1992; Zequeira-Larios 2014). However, there are still some farmers in southeastern Mexico that have preserved and still continue to work cocoa-AFS. At present, small cocoa farmers in Mexico suffer from heavy economic pressures to increase their production due to high domestic demand (Sandoval 2012; Zequeira-Larios et al. 2012), leading to management intensification, and thus reducing the number of tree species used as shade in order to include more cocoa plants (Somarriba and López-Sampson 2018).
Somarriba and López-Sampson (2018) established that measures to avoid declining cocoa areas to the benefit of other crops and land uses are (a) increasing on-farm profitability through higher international prices, (b) increasing crop yield, (c) intercropping and shade tree products for household consumption and sale, and (d) payments for ecosystem services. To establish government initiatives that favor these actions, the conservation of the existing cocoa plantations and the increase of the harvested area were done in countries such as Ecuador, Colombia, Brazil, Bolivia, and Peru; studies focused on knowing the different existing agricultural typologies are necessary from cocoa-AFS (Somarriba and López-Sampson 2018). In this sense, studies in Mexico were carried out in the State of Chiapas in the municipalities with the highest cocoa production (Soconusco region), where a high plant species richness is found within those cocoa agroforestry systems (Salgado-Mora et al. 2007). The most recent studies in the state of Tabasco focused on the study of the diversity, structure, and carbon content of the arboreal vegetation of the cacao agroforestry systems where Salvador-Morales et al. (2019) highlighted that regardless of the little contribution that cocoa trees have in the storage of carbon, they are important in environmental services such as the conservation of biodiversity. However, similar studies are necessary for other municipalities in the state of Tabasco to determine the tree diversity and to contribute more information on the tree structure and local uses of these agroecosystems. The present study contributes new information that can favor the establishment of public policies in favor of the conservation of cocoa-AFS in Mexico and to identify which tree species can be utilized in different agroforestry designs. Hence, in this study our objectives were to determine tree species diversity, composition, and abundance in traditional (i.e., rustic) cocoa-AFS from the states of Tabasco and Chiapas within four municipalities, two in Tabasco (Cárdenas and Huimanguillo) and two in Chiapas (Pichucalco and Ixtacomitán).
Materials and methods
Study sites
In Mexico, the area dedicated to cocoa cultivation is of + 60,000 ha with an average yield between 250 and 560 kg ha−1 (SIAP 2019). It is estimated that about 40,000 families grow cocoa in Tabasco and 11,000 in Chiapas (PRCC 2012). Approximately 70% of the Mexican cocoa production is concentrated in the state of Tabasco and 20% in the state of Chiapas, with the remaining production spread in other parts of the Mexican southeast (Ortega et al. 2015; SIAP 2019).
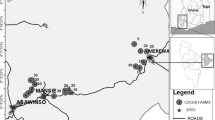
Two of our study sites are located in the state of Tabasco (Fig. 1): Huimanguillo (17° 49′ and 17° 83′ N, 93° 23′ and 93° 39′ W; ~ 20 m asl) and Cárdenas (17° 55′ and 18° 25′ N, 93° 16′, and 94° 08′ W; elevation ranges from 0 to 10 m asl). Both municipalities have a warm humid climate with abundant summer rains (2000–3000 mm/year), with temperature ranging between 22 and 28 °C. The main agricultural activity in these municipalities is sugarcane, followed by cocoa-AFS and extensive cattle ranching (INEGI 2013a,2013b).

Location of study sites in Tabasco and Chiapas states, southeastern Mexico. These two states produce > 90% of the cacao from Mexico
The other two study sites are located in the northwestern side of the state of Chiapas (Fig. 1): Pichucalco (17° 19′ and 17° 42′ N; 93° 00′ and 93° 25′ W; altitude ranges from 0 to 1300 m asl) and Ixtacomitán (17° 21′ and 17° 30′ N; 92° 02′ and 93° 11′ W; elevation ranges from 0 to 1200 m asl). The climate in these municipalities is warm humid with rains all year (2500–4500 mm/year), and temperatures range between 20 and 28 °C. Agricultural activities are mainly cocoa-AFS, followed by the cultivation of fruit trees, e.g., mamey (Pouteria sapota), citrus (Citrus spp.), and by extensive cattle ranching (INGEI, 2013a,2013b). These municipalities belong to the Zoquean area of Chiapas (INPI 2020).
Data collection
We collected information from 57 households in a non-probabilistic sample, recording data on vegetation composition and diversity of 38 plots of 20 × 50 m (5 in Cárdenas, 14 in Huimanguillo, 4 in Ixtacomitán, and 15 in Pichucalco) from January to August 2012. Given the region’s insecurity, the criteria for choosing the households included only those plantations to which we had explicit permission to access and conduct research where the farmers guaranteed researchers’ safety and support. We applied a structured questionnaire with four sections (data of farm, experience and problems, experience in farmer school, and general data about the family) that included close and open questions. Within each plot, we recorded and identified all trees with a diameter at breast height (DBH) ≥ 5 cm and measured the height and percentage of canopy cover with an Haga Altimeter and densiometer single crosshair by visual estimation, respectively (as suggested by Somarriba 2002).
Owner farmers provided common names of most of the recorded trees. We subsequently cross-referenced each specimen’s common name with its scientific name consulting Maldonado et al. (2000), Magaña (2006), and Pennington and Sarukhán (2005). Other crops cultivated outside of the cocoa-AFS but within the owner’s property were mentioned by the farmers during the interview (e.g., corn, beans), in which case we consulted their common name in Maldonado et al. (2000). All species’ identifications were validated by a botanic expert. We deposited all collected specimens in the Herbarium of the Academic Division of Biological Sciences at the Autonomous Juárez University of Tabasco (UJAT, for its acronym in Spanish).
Data analysis
We determined the species richness statistical expectation (Sest) for each municipality and state using individual-based data calculation with EstimateS 9 (Colwell 2015, EstimateS: Statistical estimation of species richness and shared species from samples, version 9, http://viceroy.eeb.uconn.edu/estimates). Given that our sample sizes differed among municipalities, we contrasted both municipalities with few samples (i.e., Ixtacomitán, 4, Cárdenas, 5) and those with more samples (i.e., Pichucalco, 15 and Huimanguillo, 14). To statistically compare expected species richness among municipalities and between states, we contrasted the 84% confidence intervals of the species richness statistical expectations, following two studies that demonstrate that overlapping 84% confidence intervals mimic α = 0.05 tests; thus, we considered significant differences whenever confidence intervals did not overlap (MacGregor-Fors and Payton 2013; Payton et al. 2003). We also performed an abundance-based Bray–Curtis multivariate cluster analysis (group average-linkage) to analyze tree species composition similarity at the municipality level, using vegan (Community ecology package “vegan”; https://cran.r-project.org, https://github.com/vegandevs/vegan). To complement these analyses, we constructed rank–abundance curves to assess the distribution of the abundances of the tree species comprising the studied assemblages, as well as to identify dominant species (Magurran 2003).
For all tree species recorded at each sampled plot, we calculated their importance value (IV, sum of relative values of frequency, density, and basal area; Newton 2007). We calculated descriptive statistics for the questions of the interviews (i.e., median, standard deviation, minimum, maximum) and conducted t tests to compare variables between states. Interval classes of the variables were calculated with the Sturges rule (Scott 2009; Sturges 1926).
According to the answers of farmers, the species used were classified in two categories: Option I-species use: (a) shade, the tree is used as permanent shade for the cocoa tree; (b) wood, species of trees the farmers preserves them for domestic use or sell; (c) food, parts of the tree are consumed; (d) medicine, some parts of the tree, such the bark or leave, are used for medicinal purposes in homes; (e) others, trees whose parts are used to make other objects or some parts are used in local gastronomy. Option II-species commercialization: (a) not commercial, (b) few commercial, the products are sold in small quantities in the local market, (c) wide commercial, the products are sold in the formal market in large quantities.
We used general linear mixed models (GLMM) to analyze tree species abundance in cocoa plots, having state (Tabasco, Chiapas) and municipality (Huimanguillo, Cárdenas, Pichucalco, Ixtacomitán) as categorical explanatory factors, and tree species as a random factor. Given the nature of our data, we used a Poisson distribution with a “log” link to run models (function glmer() from the package lme4 v1.1–15; Linear mixed-effects models using “Eigen” and S1; https://github.com/lme4/lme4/, http://lme4.r-forge.r-project.org/). We conducted a similar analysis for the response variables DBH, tree height, and tree canopy cover. Because DBH, height, and cover showed to be correlated (r > 0.75, P < 0.05), we provide only the results for DBH, as it is the variable that explained more variance in the model. We used the function rsquared from the R-package piecewiseSEM to calculate marginal (random factor) and conditional (full model) variances explained by the model (Lefcheck 2016).
Given that cocoa trees are the most abundant in all studied cocoa-AFS, we conducted a separate statistical analysis in order to assess differences in the number of cocoa trees at our study sites, at the municipality level. Given that our design is unbalanced and that we found no homogeneity of variance of cocoa trees across municipalities (Fligner-Killeen test: med χ2 = 3.32, df = 3, P = 0.34), we performed a nonparametric Kruskal–Wallis test, followed by a post hoc Dunn test to assess the cocoa density. We performed all analyses in R (v 3.3.3, http://www.R-project.org).
Results
Tree species composition
A total of 3268 individual trees of 66 species and 29 families were recorded within a surveyed area of 3.8 hectares. Tree families with the highest representation in our surveys were: Malvaceae, Fabaceae, Meliaceae, Musaceae, Rutaceae, Rhamnaceae, Boraginaceae, and Anacardiaceae. In the state of Tabasco, we recorded 1972 trees (1038/ha) of 42 species. The most abundant species were Theobroma cacao and Erythrina americana. In the state of Chiapas, we recorded a total of 1296 trees (682/ha) of 52 species, including tree species that belong to primary forest vegetation such as Guarea glabra, Ficus sp., and Bursera sp., and species of secondary vegetation (i.e., acahual) such as Pithecellobiun sp. The most abundant species were T. cacao and Gliricidia sepium (Table S1).
Tree species uses
According to the uses that farmers give to the cocoa-AFS and their land, the assessed agroforestry systems can be considered a diversified food-and-cash-crop livelihood strategy (Tscharntke et al. 2011). Results for categories: Option I-species use; showed for Tabasco that 55% of tree species are used for food, 31% of tree species are for wood, 24% of tree species are for other uses, the 14% of tree species some parts are used as a medicine, and 12% of the tree species are used as shade for cocoa tree. In Chiapas, the 48% of tree species are used for food, 25% of tree species are for wood, 15% of tree species are for other uses, the 10% of the tree species are used as shade for cocoa tree, and 8% some parts of tree species are used as a medicine (Table S1). Option II-species commercialization; in Tabasco 90% of the trees are not commercial; 31% of the tree species are few commercial and the 2% of tree species are wide commercial. In Chiapas, the 50% of the trees are not commercial; 48% of the tree species are few commercial, and the 2% of tree species are wide commercial (Table S1).
Tree diversity and structure
We recorded 27 tree species that were shared in cocoa-AFS between Tabasco and Chiapas, 24 species were recorded only in Chiapas, and 15 were unique to Tabasco (Table S1). We did not find significant differences in the number of tree species recorded in both states (Tabasco: 42.9 ± 5.1 species, Chiapas: 52.0 ± 5.7 species), neither we did between both municipalities for which we had fewer samples (Ixtacomitán: 22.5 ± 2.9 species, Cárdenas: 25.0 ± 5.6 species; Fig. 2a). Yet, we found significant differences between both municipalities with more samples (Pichucalco: 47.0 ± 4.3 species, Huimanguillo: 36.0 ± 5.5 species), with Pichucalco showing the highest tree species richness of the four studied municipalities (Fig. 2a).

a Species richness expectation between states and municipalities of Tabasco and Chiapas; n indicates the number of plots sampled per site. b Cocoa tree abundances among municipalities. c Rank–abundance curves for tree species found in the municipalities of Tabasco and Chiapas. d Multivariate cluster showing tree species composition similarities of cocoa-AFS plantations among municipalities of Tabasco and Chiapas
When analyzing cocoa tree abundance, we found their numbers to differ among municipalities (χ2 = 20.68, df = 3, P < 0.001; Fig. 2b). Cárdenas and Huimanguillo (Tabasco) showed higher cocoa tree abundance per plot and nonsignificant differences between them (Dunn test = 0.66, P = 0.25), and Ixtacomitán and Pichucalco (Chiapas) had lower cocoa tree abundance per plot and nonsignificant differences between them (Dunn test = 0.37, P = 0.35). We found significant differences in cocoa tree abundance between states, with Tabasco having more cocoa trees per plot when compared with Chiapas (Dunn tests > 2.11, P < 0.01) (Table S2, Fig. 2b).
Our analyses showed significantly higher tree abundances in Tabasco compared to Chiapas, and significantly lower tree abundances in Huimanguillo compared to Cárdenas and Pichucalco (Table S3). Tree species explained 70% of the variance in abundance, whereas state and municipality explained 3% and 4%, respectively, in the model. The most abundant species across study sites were T. cacao, E. americana, and G. sepium (Fig. 2c; Tables S4, S5). E. americana was particularly more abundant in Cárdenas and Huimanguillo (i.e., Tabasco) compared to the municipalities in Chiapas. In contrast, G. sepium was more abundant in Ixtacomitán and Pichucalco than in Cárdenas and Huimanguillo (Fig. 2c; Tables S4, S5). In Tabasco, the tree with the highest importance value as a shade tree was E. americana, but there are also other important tree species such as Cedrela odorata, Colubrina arborecens, Guazuma ulmifolia, Diphysa robinioides, Citrus sinensis, Erythrina fusca, and Mangifera indica, with more than 4% of individuals (Table S4). In Chiapas, the tree species with the highest importance value as a shade tree was G. sepium, whereas other relevant tree species with frequency ≥ 4% were: Hevea brasiliensis, Annona muricata, E. americana, G. ulmifolia, Enterolobium cyclocarpum, C. Sinensis, Pouteria sapota, Cordia alliodora, and C. odorata (Table S5).
The abundance-based multivariate cluster analysis showed two clear groupings: Huimanguillo–Cárdenas and Pichucalco–Ixtacomitán (Fig. 2d); yet, differences measured with the Bray–Curtis index showed less dissimilarity in the former group (Bray–Curtis = 0.31), and higher dissimilarity values for the latter group (Bray–Curtis = 0.54). The Bray–Curtis dissimilarity value between both groups was high (average = 0.75 ± SD 0.08), indicating very different tree species composition between Tabasco and Chiapas traditional cocoa-AFS (Fig. 2d).
Cocoa trees DBH in Tabasco was on average of 11.83 cm (± SD 3.38) and of 14.07 cm (± SD 4.61) in Chiapas (Table S6, Fig. S1). When comparing the DBH of all tree species between states, we also found that trees from plantations in Chiapas had significantly larger DBH than those in plantations from Tabasco (Table S6). The variable state explained 6.5% of the variance and tree species explained 57.7% of the DBH variance in the model. In general, the higher tree species (> 20 m) found in cocoa-AFS were: Artocarpus altilis, C. odorata, Ceiba pentandra, Chrysophyllum cainito, C. arborecens, E. fusca, Ficus tecolutensis, H. brasiliensis, P. sapota, Pterocarpus hayesii, Samanea saman, Spondias mombin, and Swietenia macrophylla. At the plot level, we detected a higher density of cocoa trees and less abundance of other trees in Tabasco than our surveys from Chiapas (Fig. 2). Mean plantation age in Tabasco was lower than in Chiapas, and lower mean species richness in Tabasco than in Chiapas (Table S2, Fig. S2). Seventy-five percent of Tabasco cocoa-AFS had less than 30 years, whereas in Chiapas, only 25% of the plantations had less than 30 years (Fig. S2).
Household characteristics
In reference to how farmers use their land, in Tabasco each farmer owns an average of 8 ha (± SD 8.6; range: 0.5–37.5 ha per interviewed farmer), whereas in Chiapas they possess an average of 9.3 ha (± SD 6.9 ha; range = 1–27 ha per farmer; Table S2, Fig. S3). From the interviewed farmers, each farmer in Tabasco uses an average of 2 ha (± SD 1.1) for cocoa-AFS (range: 0.5–5.8 ha) and in Chiapas each farmer uses 5 ha (± SD 3.3) for cocoa-AFS (range: 1–12 ha) (Table S2, Fig. S4).
Regarding land tenure, in Tabasco 8% (n = 3) of the farmers have private property and 92% (n = 34) have ejido property;Footnote 1 in Chiapas 16% (n = 3) of the farmers have private property and 84% (n = 16) have ejido property. Regarding the main source of income, 89% (n = 34) of the farmers interviewed in Tabasco live from agriculture; 11% (n = 4) have other off-farm income (e.g., construction, driving). In Chiapas, proportions were surprisingly the same, with 89% (n = 17) living from agriculture and the rest 11% (n = 2) living from other additional activities. According to land use, we identified two types of farming: (1) where land was entirely dedicated to cocoa-AFS production and (2) where besides cocoa land is used for sugarcane cultivation and/or for cattle ranching. In the first case, farmer income is mostly dependent on selling cocoa, selling other crops and other forms of work (e.g., builder, laborer, and employee). In Tabasco 25% (n = 9) and in Chiapas 42% (n = 8) of farmers correspond to this category. In the second case, farmer income mainly derives from selling sugarcane, renting grasslands as pastureland, and selling cattle and cocoa. Additionally, they receive income derived from selling products obtained from shade cocoa-AFS. In Tabasco 75% (n = 27) and in Chiapas 58% (n = 11) of the farmers correspond to this category. Although we did ask about their crop incomes, none of them could tell, as they do not keep records.
Regarding knowledge on how farmers learned to grow cocoa showed that in Tabasco 55% (n = 21) learned from their parents, 5% (n = 2) from their grandparents, 3% (n = 1) from a relative, and 37% (n = 14) learned from friends. In Chiapas, 58% (n = 11) learned from their parents, 5% (n = 1) from their grandparents, 5% (n = 1) from a relative, and 32% (n = 6) from friends (χ2 = 0.366, df = 3, P < 0.05).
In relation to the ways in which the interviewed farmers sell cocoa, in Tabasco 87% (n = 33) sell their product in fresh (the seeds of the freshly cut cob), 8% (n = 3) sell dry cocoa, and 5% (n = 2) in other forms (e.g., roasted). In Chiapas, 17% (n = 3) sell fresh cocoa, 72% (n = 13) sell dry cocoa, and 11% (n = 2) in other forms (e.g., roasted) (χ2 = 27.632, df = 2, P < 0.05). In Tabasco 100% (n = 37) of the cocoa production was sold to a cooperative society, and in Chiapas 86% (n = 12) was sold to a cooperative society and 14% (n = 2) to an intermediary (locally known as “coyote”).
Discussion
The traditional management of cocoa-AFS in Mexico is very similar in the four studied municipalities of the two most important producing states (i.e., Tabasco and Chiapas), where farmers still use the same tree species known to be used by the Mayas since pre-Hispanic times (Gómez-Pompa et al. 1990; Millón 1955), sheltering a large array of tree species that can be used not only as shade, but also as timber, food for self-consumption or crops to sell. According to Somarriba et al. (2011), cocoa-AFS from our study areas are considered as varied cocoa-AFS with shade. The diversity and composition of cocoa-AFS may benefit cocoa growth and help to fulfill, in some cases, households’ food requirements, providing additional income and material for house construction (e.g., Anim-Kwapong and Osei-Bonsu 2009; Bisseleua et al. 2009; Jagoret et al. 2014). Despite some similarities between Tabasco and Chiapas cocoa-AFS, we also found that there are differences in management and tree species diversity and composition. Different shade tree species are used in Tabasco and Chiapas, the species composition was 75% different between the states, and there is a tendency for higher tree species richness in Chiapas than in Tabasco cocoa-AFS. The interviewed farmers only recognize Erythrina sp. in Tabasco and Gliricidia sp. in Chiapas as a permanent shade necessary for the profitable growth of cocoa. The rest of the tree species are later incorporated to satisfy the tastes or nutritional needs of the home, as well as to make optimal use of the territory at their disposal; it is more of a diversification strategy (Anderzén et al. 2020). In the composition of cocoa-AFS, it is considered that there are no other species associated with plantations with large-scale commercial purposes such as cocoa.
We recorded 52 tree species in cocoa-AFS of northern Chiapas, which is 10% higher than the 47 species reported by Salgado-Mora et al. (2007) in the Soconusco region (southeastern Chiapas). The tree species used for cocoa shade in the surveyed plots was different between our study area and the Soconusco. The main species for Pichucalco and Ixtacomitán were G. sepium, E. americana, D. robinioides and C. arborescens, whereas Inga micheliana and G. sepium dominated cocoa-AFS in the Soconusco (Salgado-Mora et al. 2007). Regarding tree species used for timber, C. odorata and C. alliodora were the most common in our study sites in Chiapas, whereas C. alliodora and Tabebuia donnell-smithii were the most abundant in the Soconusco region studied by Salgado-Mora et al. (2007). Finally, the percentage of tree cover we recorded (60% ± SD 28) is lower in relation to that reported by Salgado-Mora et al. (2007; 89% ± SD 0.56). This mainly derives from the recently established practices implemented in 2011 (Zequeira-Larios et al. 2012) by the Nestle Cocoa Plan in these communities, where they promote a reduction of up to 30% of shade to increase cocoa production (compare Figs. S5 and S6). At this point, these practices are unsustainable and have been documented not to be the best option for neither families nor the environment (Andres et al. 2016; Córdova et al. 2018; Notaro et al. 2020; Middendorp et al. 2018; Santhyami et al. 2018).
In Tabasco, we recorded higher species richness of tree species (n = 42) in cocoa-AFS compared to the study by Salvador-Morales et al. (2019) carried out in Comalcalco, Cárdenas, and Jalpa de Méndez, all municipalities from Tabasco, where a total of 39 species were reported. The dominating tree species reported by Salvador-Morales et al. (2019) were D. robinioides, E. americana, C. arborescens, C. odorata, and Tabebuia rosea. We recorded the same species with the exception of T. rosea. Other studies in Latin America have shown ecological differences between regions and greater plant diversity in more advanced successional stages of cocoa-AFS (Braga et al. 2018; see also Marconi and Armengot 2020). In these case studies, we found greater diversity in Chiapas, which were older.
Differences in the abundance and floristic composition of tree species within the assessed cocoa-AFS between Tabasco and Chiapas indicated that farmers favor different species at the local level. Interestingly, we found higher species composition dissimilarities between municipalities in Chiapas than Tabasco; this could be due to social aspects and management decisions (Anderzén et al. 2020; Córdova et al. 2018). In the study cases, we found the plantations in Chiapas are more traditional while in Tabasco plantations are more commercial. Yet, in both cases, the cocoa-AFS included other tree species to satisfy specific household needs.
In Tabasco, there is a higher density of cocoa trees and less abundance of other tree species than in Chiapas; this shows that the process of intensification of cocoa production occurs mainly in Tabasco. Possibly this happens because in Tabasco farmers are more focused on the sale of cocoa; in this way, they keep cocoa trees in constant renewal and maintain the shade trees. In contrast, Chiapas’s farmers obtain income from cocoa and fruit trees. Furthermore, in Chiapas we found older plantations and cocoa trees with a larger DBH indicating less renewal, corroborating data from the interviews with local owners. The two most abundant tree species, E. americana in Tabasco and G. sepium in Chiapas, have important benefits where cocoa trees grow. On the one hand, E. americana during winter sheds its leaves and becomes covered with red flowers, but during the rest of the year, when the shade is required, E. americana is heavily covered with leaves (Millon 1955). Farmers from Tabasco said that it is advantageous to use it as mother/shade tree; in addition, it is a tree that saves moisture and has edible flowers. On the other hand, G. sepium sheds its leaves in spring and it is full with leaves during winter. Thus, its foliage increases the humus and reduces the high temperatures that destroy litter, provides protection to the soil, decreases the erosion, and with the fall of its leaves helps to conserve groundwater (CONABIO 2019). Regarding its uses, G. sepium produces firewood of excellent quality, makes good fire, and burns with little smoke (local farmers pers. comm.). Also, its seeds, leaves, bark, and roots contain toxic substances that are used locally to poison rodents in the fields. The leaves and bark are used in the form of plasters and general bath as remedy for measles and fever; the trunk is a support for climber species like vanilla and other epiphytes (CONABIO 2019). For this species, farmers from Chiapas confirmed its use as good firewood that they routinely use on household or to sell. Additionally, this tree genus has been suggested to play an important role in the nutrition of cocoa trees and support cocoa root development and distribution by provision of nitrogen (Niether et al. 2019).
Importance values for tree species in Tabasco were lower in this study than those recorded for species found by Salvador-Morales et al. (2019); yet, recorded tree species were the same between the two studies. The species with higher importance value are those with commercial relevance for farmers (e.g., C. sinensis, M. indica, C. odorata, C. arborecens). In the case of E. fusca, it was an introduced species during the 1970s as shade for cocoa-AFS via government programs. We found individuals with DBH > 250 cm, many of which were found with a ringed mark (Fig. S7). According to farmers, E. fusca is a very damaging tree for cocoa trees given that it has easily breakable branches, but it is commonly used for construction (pers. obs.). Also, C. arborecens is used for construction, which showed a 6% frequency in cocoa-AFS and farmers commonly sell it.
Two tree shade species had highest importance values in Chiapas, G. sepium and H. brasiliensis. The latter is an introduced species in Mexico and it was established in Chiapas for rubber exploitation and commercialization (Aguirre-Ríos and Santoyo 2013). The rest of the species with considerable importance value (i.e., > 8) are of commercial interest for local farmers, which include C. sinensis, P. sapota, A. muricata, C. alliodora, and C. odorata. In this study, farmers use these tree species because that is the way they were thought by their relatives and friends according to our surveys. For instance, in a new plantation farmers would cultivate the shade trees (E. american or G. sepium) at a distance of 12 m from each other (i.e., in a 12 × 12 m. grid) before they plant the cocoa trees, along with fast growing trees such as D. robinioides and C. arborescens and fast producing plants such as Musa spp. and Manihot esculenta that are used as temporary shade. When cocoa trees are mature, farmers remove temporary shade species and subsequently plant in those empty spaces fruit and/or wood trees, which many times are dispersed by birds and bats (pers. obs.). The reported shade trees transition between successional management phase of cocoa-AFS is similar to the process described by others authors in the Brazilian Amazon (Braga et al. 2018). We also recorded farmers growing Hylocereus spp. (the dragon fruit) on shade trees from the species G. sepium, E. americana, and E. fusca, which Calix et al. (2005) have reported as excellent support for this highly valued fruit worldwide (Perween et al. 2018). We identified that farmers in both Tabasco and Chiapas are convinced that cocoa needs permanent shade, and most of them resist to dismiss or break down trees without a reason. In Tabasco, farmers mentioned that the specie E. fusca causes a lot of damage to cocoa trees because its large branches are very brittle, so most of them decided to cut them down. In both entities, the density of cacao trees depends on territory availability or their strategies for managing the territory. At the time of the investigation, 100% of interviewed farmers stated that the cultivation of cocoa is no longer profitable for them and represents more expenses than profit due to low prices in the market.
The cocoa trees DBH was significantly higher in Chiapas than in Tabasco, indicating older trees in cocoa-AFS from Chiapas. Some studies suggest that in Tabasco the low productivity of cocoa trees is due to its age, arguing that they are too old (Avendaño-Arrazate et al. 2011; Córdova-Ávalos et al. 2001). However, in this investigation we showed that it is rather in the state of Chiapas where there are older cocoa trees, while in Tabasco there is evidence of renovation work in the plantations. Thus, actively managing cocoa-AFS based on regional commercial needs is in the economic benefit of local people.
Regarding the conservation value of cocoa-AFS, we recorded the same tree species (i.e., G. sepium, E. americana, D. robinioides, C. sinensis, C. odorata, G. ulmifolia, M. indica, and Cecropia obtusifolia) when contrasted with those reported by Ibarra et al. (2001) that were used by birds. Interestingly, these plant assemblages have been associated with a wide variety of avian species. Other studies, such as Muñoz et al. (2006), showed that diversity of trees presented in the studied cocoa-AFS provide suitable habitat for different species of animals, including howler monkeys (Alouatta palliata). Evidence demonstrates adverse impacts of intensively managed or sun cocoa-AFS for biodiversity (e.g., Santos-Heredia et al. 2018; Bisseleua et al. 2009). In this regard, federal and state government programs tilt the balance regarding the type of crops and species that are favored by local farmers, which are not necessarily those with long-term benefits for both local farmers and biodiversity. For instance, currently and in detriment to biodiversity, both Tabasco and Chiapas states have higher economic incentives for sugarcane and oil palm production (Xantomilla 2019; FIRA 2020a; SAGARPA, 2020). Such financial incentives, along with urbanization, have promoted the loss of large extensions of cocoa-AFS in the region (Oporto-Peregrino et al. 2019). Furthermore, the type of land tenure affects the access to government resources, where credits are available for private owners and not for those within ejidos (FIRA 2020b). Yet, as shown in this study, most of the cocoa-AFS owners in the region belong to ejidos tenure land, suggesting that very few people benefit from government programs.
This study provides valuable information in terms of description and characterization of the vegetation structure, diversity, composition, and management practices in cocoa agroforestry systems of Tabasco and Chiapas, Mexico. However, we recognize that the study’s limitations focus on the sampling design since the plots studied were not randomized. In general, the results provide a description of cocoa production systems in new territories and show some of the management practices carried out by cocoa producers in Mexico. Therefore, further studies of floristic inventories and livelihood practices of cocoa agroforestry systems established in other municipalities in both states are necessary. In other scientific areas, we suggest conducting market studies of fruit and timber species found in this study. It is recommended to carry out experimental plots with cocoa/fruit, cocoa/wood association in abandoned pastures that previously were cocoa agroforestry systems (Somarriba et al. 2018), in order to recover degraded areas and increase the cacao cultivation area, improving the living conditions of farmers and at the same time contributing to the mitigation of climate change and biodiversity conservation (Blaser et al. 2018; Middendorp et al. 2018).
Conclusions
The management practices employed by cocoa smallholders have allowed them to conserve these agroforestry systems and to obtain income from the local market by selling a great variety of products grown in this system, as well as satisfying their own food needs. The importance value recorded in this study confirms the interest of producers to keep commercially relevant timber and fruit species. This suggests individual interest in diversifying their income and family support.
In the cocoa-AFS of the two states evaluated, we recorded 29 families and 66 tree species. Tree families with the highest representation were: Malvaceae, Fabaceae, Meliaceae, Musaceae, Rutaceae, Rhamnaceae, Boraginaceae, and Anacardiaceae. The most important species were: E. americana, C. odorata, C. arborecens, G. ulmifolia, D. robinioides, C. sinensis, E. fusca, and M. indica, H. brasiliensis, A. muricata, E. cyclocarpum, P. sapota, and C. alliodora.
Differences in the abundance and floristic composition of tree species indicated that farmers favor different species at the local level and included other tree species to satisfy specific household needs. A greater tree diversity was found in Chiapas, but a greater abundance of cacao trees in Tabasco. For cocoa shade, farmers used mainly E. americana in Tabasco and G. sepium in Chiapas. The indigenous people used these tree species before the arrival of the Spanish in Mexico. The E. fusca species was introduced in Mexico and it was rather damaging to cacao trees.
The structure, diversity, and composition of the cocoa-ASF in Mexico are traditional and respond more to local needs than to a large-scale commercial strategy.
Notes
The ejido is a type of land in Mexican legislation, product of the revolutionary agrarian reform. The ejidos and agrarian communities are a type of social property or agrarian nuclei; constitute forms of exclusive ownership of land in the country (Morett-Sánchez and Cosío-Ruíz 2017).
References
Aguirre-Ríos CE, Santoyo-Cortés VH (2013) El cultivo del árbol del hule (Hevea brasiliensis Muell Arg): avances y retos en la gestión de la innovación. Colección Trópico Húmedo 1st edn. Universidad Autónoma de Chapingo, Estado de Mexico
Anderzén J, Guzmán LA, Luna-González DV, Merrill SC, Caswell M, Méndez EV, Hernández JR, Giménez CMMT (2020) Effects of on-farm diversification strategies on smallholder coffee farmer food security and income sufficiency in Chiapas, Mexico. J Rural Stud 77:33–46
Andres C, Comoé H, Beerli A, Schneider M, Rist S, Jacobi J (2016) Cocoa in monoculture and dynamic agroforestry. In: Lichtfouse E (ed) Sustainable agriculture reviews, vol 19. Springer, Switzerland, AG. https://doi.org/10.1007/978-3-319-26777-7_3
Anglaaere LCN, Cobbina J, Sinclair FL, McDonald MA (2011) The effect of land use systems on tree diversity: farmer preference and species composition of cocoa-based agroecosystems in Ghana. Agroforest Syst 81:249–265. https://doi.org/10.1007/s10457-010-9366-z
Anim-Kwapong GJ, Osei-Bonsu K (2009) Potential of natural and improved fallow using indigenous trees to facilitate cacao replanting in Ghana. Agroforest Syst 76:533–542. https://doi.org/10.1007/s10457-008-9196-4
Attolini LA (2011) Cuentas, dares y tomares del cacao: delicia, convite, rito mesoamericano. Aspectos antropológicos. Revista Digital Universitaria 12(4):3–22. http://www.revista.unam.mx/vol.12/num4/art38/art38.pdf
Avendaño-Arrazate CH, Villarreal-Fuentes JM, Campos-Rojas E, Gallardo-Méndez RA, Mendoza-López A, Aguirre-Medina JF, Sandoval-Esquivez A, Espinosa-Zaragoza S (2011) Diagnóstico del Cacao en México. SAGARPA, SNICS, SINAREFI, Universidad Autónoma de Chapingo, INIFAP, México DF
Beer J, Muschler R, Kass D, Somarriba E (1998) Shade management in coffee and cacao plantations. Agroforest Syst 38:139–164
Bentley JW, Boa E, Stonehouse J (2004) Neighbor trees: shade, intercroping and cocoa in Ecuador. Human Ecol 32:241. https://doi.org/10.1023/B:HUEC.0000019759.46526.4d
Bergman JF (1969) The Distribution of Cacao Cultivation in Pre-Columbian America. Annal Assoc Am Geograph 59:85–96
Bhagwat SA, Willis KJ, Birks HJB, Whittaker RJ (2008) Agroforestry: a refuge for tropical biodiversity? Trend Ecol Evol 23:261–267. https://doi.org/10.1016/j.tree.2008.01.005
Bisseleua DHB, Missoup AD, Vidal S (2009) Biodiversity conservation, ecosystem functioning, and economic incentives under cocoa agroforestrry intensification. Conserv Biol 23:1176–1184. https://doi.org/10.1111/j.1523-1739.2009.01220.x
Blaser W, Oppong J, Hart S, Landolt J, Yeboah E, Six J (2018) Climate-smart sustainable agriculture in low-to-intermediate shade agroforests. Nature Sustainability 1(5):234–239
Braga DPP, Domene F, Gandara FB (2018) Shade trees composition and diversity in cacao agroforestry systems of southern Pará, Brazilian Amazon. Agroforest Syst 93:1409–1421. https://doi.org/10.1007/s10457-018-0250-6
CONABIO (2019 Comisión Nacional para el Conocimiento y uso de la Biodiversidad, Gliricidia sepium. CONABIO. http://www.conabio.gob.mx/conocimiento/info_especies/arboles/doctos/29-legum19m.pdf. Accessed 12 Sept 2019
Córdova R, Hogarth NJ, Kanninen M (2018) Sustainability of smallholder livelihoods in the Ecuadorian highlands: a comparison of agroforestry and conventional agriculture systems in the indigenous territory of Kayambi people. Land 7(2):45. https://doi.org/10.3390/land7020045
Córdova-Ávalos V, Sánchez-Hernández M, Estrella-Chulím NG, Macías-Layalle A, Sandoval-Castro E, Martínez-Saldaña T, Ortiz-García CF (2001) Factores que Afectan la Producción de Cacao (Theobroma cacao L.) en el Ejido Francisco I. In: Madero del Plan Chontalpa, vol 17, no. 34. Universidad y Ciencia, Tabasco, México, pp 93–100
da Silva Moco MK, da Gama-Rodrìguez AC, Machado RCR, Baligar VC (2008) Soil and litter fauna of cacao agroforestry systems in Bahia, Brazil. Agroforest Syst 76:127–138. https://doi.org/10.1007/s10457-008-9178-6
Dawoe E, Asante W, Acheampong E, Bosu P (2016) Shade tree diversity and aboveground carbón stocks in therobroma cacao agroforestry systems: Implications for REDD+ implementation in a West African cacao landscape. Carbon Balance Manag 11(1):11–17
Calix de DH, Castillo MR, Rodríguez CA, Castañeda CR (2005) El cultivo de la Pitahaya en el trópico. Gobierno el estado de Tabasco, México
FIRA (2020a) Estímulos a la producción para Palma de aceite 2018. https://www.fira.gob.mx/Nd/estimulosPalma.jsp. Accessed 26 Feb 2021
FIRA (2020b) Fideicomiso Instituido en Relación con la Agricultura. https://www.fira.gob.mx/Nd/AcercadeNosotros.jsp. Accessed 26 Feb 2021
Gómez-Pompa A, Salvador-Flores J, Aliphat-Fernández M (1990) The sacred groves of the Maya. Lat Am Antiq 1(3):247–257
Greenberg R, Bichier P, Sterling J (1997) Bird populations in rustic and planted shade coffee plantations of Eastern Chiapas, México. Biotropica 29:501–514
Greenberg R, Bichier P, Cruz Angón A (2000) The conservation value for birds of cacao plantations with diverse plented shade in Tabasco, Mexico. Anim Conserv 3:105–112
Gyau A, Smoot K, Kouame C, Diby L, Kahia J, Ofori D (2014) Farmer attitudes and intentions towards trees in cocoa (Theobroma cacao L.) farms in Côte d’Ivoire. Agroforest Syst 88:1035–1045. https://doi.org/10.1007/s10457-014-9677-6
Ibarra AE, Arriaga WE, Estrada MA (2001) Avifauna asociada a dos cacaotales tradicionales en la región de la Chontalpa, Tabasco. Mexico Universidad y Ciencia 17(34):101–112
INEGI (2013a) Prontuario de información geográfica municipal de los Estados Unidos Mexicanos. Cárdenas, Tabasco. Clave geodésica 27008. INEGI, México
INEGI (2013b) Prontuario de información geográfica municipal de los Estados Unidos Mexicanos. Huimanguillo, Tabasco. Clave geodésica 27002. INEGI, México
INEGI (2013c) Prontuario de Información geográfica municipal de los Estados Unidos Mexicanos. Pichucalco, Chiapas. Clave geodésica 07068. INEGI, México
INEGI (2013d) Prontuario de información geográfica municipal de los Estados Unidos Mexicanos. Ixtacomitán, Chiapas. Clave geodésica 07043. INEGI, México
INPI Instituto Nacional de los Pueblos Indígenas (2020) Atlas de los pueblos indígenas de México. INPI. http://atlas.inpi.gob.mx/?page_id=5420. Accessed 16 Sept 2020
Jagoret P, Kwesseu J, Messie C, Michel-Dounias I, Malézieux E (2014) Farmers’ assessment of the use value of agrobiodiversity in complex cocoa agroforestry systems in central Cameroon. Agroforest Syst 88:983–1000. https://doi.org/10.1007/s10457-014-9698-1
Jezeer R, Verweij P, Santos M, Boot R (2017) Shaded coffee and cocoa—double dividend for biodiversity and small-scale farmers. Ecol Econ 140:136–145
Lefcheck JS (2016) piecewiseSEM: piecewise structural equation modeling in R for ecology, evolution, and systematics. Method Ecol Evol 7:573–579. https://doi.org/10.1111/2041-210X.12512
MacGregor-Fors I, Payton ME (2013) Contrasting diversity values: statistical inferences based on overlapping confidence intervals. PLoS ONE 8:e56794. https://doi.org/10.1371/journal.pone.0056794
MacGregor-Fors I, González-García F, Hernández-Lara C, Santiago-Alarcón D (2018) Where are the birds in the matrix? Avian diversity in a Neotropical landscape mosaic. Wilson J Ornithol 130:81–93. https://doi.org/10.1676/16-087.1
Magaña AMA (2006) Catálogo de nombres vulgares y científicos de plantas de Tabasco. Universidad Juárez Autónoma de Tabasco, Tabasco, México
Magurran AE (2003) Measuring biological diversity. Blackwell Science Ltd, Oxford
Maldonado MF, Vargas SG, Molina MRF, Sánchez SA (2000) Frutales tropicales de Tabasco, 2nd edn. Universidad Juárez Autónoma de Tabasco, Tabasco, México
Marconi L, Armengot L (2020) Complex agroforesty systems against biotic homogenization: the case of plants in the herbaceous stratum of cocoa production systems. Agricult Ecosyst Environ 287. https://doi.org/10.1016/j.agee.2019.106664
Middendorp R, Vanacker V, Lambin E (2018) Impacts of shaded agroforestry management on carbon sequestration, biodiversity and farmers income in cocoa production landscapes. Landscape Ecol 33(11):1953–1874
Millon RF (1955) When money grew on trees: a study of cacao in ancient Mesoamerica. Columbia University, New York, PhD. Dis
Morett-Sánchez JC, Cosío-Ruíz C (2017) Outlook of ejidos and agrarian communities in México. Agricultura, Sociedad y Desarrollo 14:125–152. http://www.scielo.org.mx/pdf/asd/v14n1/1870-5472-asd-14-01-00125-en.pdf
Muñoz D, Estrada A, Naranjo E, Ochoa S (2006) Foraging ecology of howler Monkeys in a Cacao (Theobroma cacao) plantation in Comalcalco Mexico. Am J Primatol 68:127–142. https://doi.org/10.1002/ajp.20211
Newton AC (2007) Forest ecology and conservation. A handbook of techniques. Oxford University Press, New York
Niether W, Schneidewind U, Fuchs M, Schneider M, Armengot L (2019) Below-and aboveground production in cocoa monocultures and agroforestry systems. Sci Total Environ 657:558–567
Notaro M, Gary C, Deheuvels O (2020) Plant diversity and density in cocoa-based agroforestry systems: how farmers´ income is effected in the Dominican Republic. Agroforest Syst. https://doi.org/10.1007/s10457-019-00472-7
Oporto-Peregrino S, Hidalgo-Mihart HG, Collado-Torres RA, Castro-Luna AA, Gama-Campillo LM, Arriaga-Weiss SL (2019) Effects of land tenure and urbanization on the change of land use of cacao (Theobroma cacao) agroforestry systems in southeast Mexico. Agroforest Syst. https://doi.org/10.1007/s10457-019-00453-w
Payton ME, Greenstone MH, Schenkerk N (2003) Overlapping confidence intervals or standard error intervals: What do they mean in terms of statistical significance? J Insect Sci 3:34. https://doi.org/10.1093/jis/3.1.34
Pennington TD, Sarukán J (2005) Árboles Tropicales de México. Manual para la identificacion de las principales especies, 3rd edn. UNAM FCE, México
Perween T, Mandal KK, Hasan MA (2018) Dragon fruit: an exotic super future fruit of India. J Pharmacogn Phytochem 7(2):1022–1026
Philpott SM, Arendt WJ, Armbrecht I, Bichier P, Diestch V, Gordon C, Greenberg R, Perfecto I, Reynosos-Santos R, Soto-Pinto L, Tejeda-Cruz C, Williams-Linera G, Valenzuela J, Zolotoff JM (2008) Biodiversity loss in Latin American coffee landscapes: review of the evidence on ants, birds, and trees. Conserv Biol 22:1093–1105. https://doi.org/10.1111/j.1523-1739.2008.01029.x
PRCC (2012) Plan rector cacao Chiapas. Comité Estatal Sistema Producto Cacao en Chiapas. http://dev.pue.itesm.mx/sagarpa/estatales/ept%20comite%20sistema%20producto%20cacao%20chiapas/plan%20rector%20que%20contiene%20programa%20de%20trabajo%202012/pr_cacao_chiapas_2012.pdf. Accessed 9 Sept 2020
Rani BD, Kumar KR, Jose S, Pal SH (2008) Ecological Basis of Agroforestry. CRC Press, Taylor & Francis Group, USA
Rueda X, Paz A, Gibbs-plessl T, Leon R, Moyano B, Lambin EF (2018) Smallholders at a crossroad: intensify or fall behind? Exploring alternative livelihood strategies in a globalized world. Bus Strat Env 27:215–229. https://doi.org/10.1002/bse
SAGARPA (2020) Guía práctica de los programas de apoyo de la SAGARPA para los productores de caña de azúcar, reglas de operación 2018. SAGARPA. https://www.zafranet.com/2018/01/sagarpapublico-la-guia-practica-de-los-programas-de-apoyo-para-productores-de-cana-de-azucar/. Accessed 12 Feb 2020
Salgado-Mora MG, Ibarra-Núñez G, Macías-Sámano JE, López-Báez O (2007) Diversidad arbórea en cacaotales del Soconusco, Chiapas, México. Interciencia 32(11):763–768. https://www.ceune.unach.mx/images/pdf_audes/publicaciones/11.pdf
Salvador-Morales P, Cámara-Cabrales L del C, Martínez-Sánchez JL, Sánchez-Hernández R, Valdés-Velarde E (2019) Diversity, structure and carbon of the arboreal vegetation on cocoa agroforestry systems. Madera y Bosques 25:1–14. https://doi.org/10.21829/myb.2019.2511638
Sandoval MC (2012) Al rescate del cacao criollo en México. Consultores en Difusión y Comunicación Social. http://codicsconsultores.blogspot.mx/2012/09/al-rescate-del-cacao-criollo-en-mexico.html. Accessed 15 Mar 2019
Santhyami S, Basukriadi A, Petala PM, Abdulhadi R (2018) The comparison of aboveground C-stock between cacao-based agroforestry system and cacao monoculture practice in West Sumatra. Indonesia Biodiversitas 19(2):472–479. https://doi.org/10.13057/biodiv/d190214
Santos-Heredia C, Andersen E, Zárate DA, Escobar F (2018) Dung beetles and their ecological functions in three agroforestry systems in the Lacandona rainforest of Mexico. Biodivers Conserv 27:2379–2394. https://doi.org/10.1007/s10531-018-1542-x
Scott DW (2009) Sturges Rule. Wires Comput Stat 1(3):303–306. https://doi.org/10.1002/wics.35
Seinfeld DM (2007) Molecular archaeological investigations of Olmec feasting in ceramics from San Andrés. Dissertation, Florida State University, Tabasco, Mexico
SIAP Servicio de Información Agroalimentaria y Pesquera (2019) Produccion Agricola por cultivo. SAGARPA. http://www.siap.gob.mx/cierre-de-la-produccion-agricola-por-cultivo/. Accessed 25 Sep 2019
Somarriba E (2002) Estimación visual de la sombra en cacaotales y cafetales. Agroforestería De Las Américas 9(35–36):86–94
Somarriba EJ, López-Sampson A (2018) Coffee and cocoa agroforestry systems: pathways to deforestation, reforestation, and tree cover change. Tech Rep. https://doi.org/10.13140/RG.2.2.29700.78724
Somarriba E, Quesada CF, Orozco AL, Cerda BR, Villalobos RM, Orozco-Estrada S, Astorga-Domian C, Deheuvels O, Say-Chávez E, Villegas-Cáseres R (2011) El manejo de la sombra del cacao. Serie Técnica. Materiales de Extensión no. 5. CATIE, Costa Rica
Somarriba E, Cerda R, López-Sampson A (2018) Analysis and design of the shade canopy of cocoa-based agroforestry systems. Burleigh Dodds Sci Publish. https://doi.org/10.19103/AS.2017.0021.29
Sonwa DJ, Weisw SF, Schroth G, Janssens MJJ, Shapiro HY (2014) Plant diversity management in cocoa agroforestry systems in West and Central Africa – effects of markets and household needs. Agroforest Syst 88:1021–1034. https://doi.org/10.1007/s10457-014-9714-5
Sturges HA (1926) The choice of a class interval. J Am Stat Assoc 21(153):65–66
Sunshine A, Bael V, Bichier P, Ochoa I, Greenberg R (2007) Bird diversity in cacao farms and forest fragments of western Panama. Biodivers Conserv 16:2245–2256. https://doi.org/10.1007/s10531-007-9193-3
Thompson S JE (1997) Historia y Religión de los Mayas. Siglo XXI, México
Toledo VM, Barrera Bassols N, García Frapollo E, Alarcón Chaires P (2007) Manejo y uso de la biodiversidad entre los mayas yucatecos. CONABIO Biodiversitas 70:10–15
Tscharntke T, Clough Y, Bhagwat SA, Buchori D, Faust H, Hertel D, Holscher D, Juhrbandt J, Kessler M, Perfecto I, Scherber C, Schroth G, Veldkamp E, Wanger TC (2011) Multifunctional shade-tree management in tropical agroforestry landscapes – a review. J Appl Ecol 48:619–629. https://doi.org/10.1111/j.1365-2664.2010.01939.x
Useche P, Blare T (2013) Traditional vs. Modern production systems: Price and nonmarket consideratios of cacao producers in Northern Ecuador. Ecol Economic 93:1–10. https://doi.org/10.1016/j.ecolecon.2013.03.010
West RC, Thom BG, Psuty NP (1985) Las tierras bajas de Tabasco en el Sureste de Mexico. Tabasqueña, biblioteca básica. Gobierno del Estado de Tabasco, Mexico
Xantomilla J (2019) Incorporan a caña de azúcar al programa de producción para el bienestar. Periódico la Jornada. https://www.jornada.com.mx/ultimas/sociedad/2019/04/22/incorporan-a-cana-de-azucar-alprograma-de-produccion-para-el-bienestar-5925.html. Accessed 22 May 2019
Yamada M, Gholz HL (2002) An evaluation of agroforestry systems as a rural development option for the Brazilian Amazon. Agroforest Syst 55:81–87. https://doi.org/10.1023/A:1020523107243
Young AM (2007) The chocolate tree. A natural history of cacao. Univ Florida Press, Gainesville, Florida
Young KJ (2018) Floristics of cacao agroforestry in the Mata Atlantica of southern Bahia, Brazil. Tropic Resour 37:23–28
Zequeira-Larios C (2014) La producción de cacao (Theobroma cacao) en México. Dissertation, Centro de Investigaciones Tropicales, Universidad Veracruzana, Tabasco, estudio de caso
Zequeira-Larios C, Ogata N, Gama-Campillo L, Brown D (2012) Escuelas de campo para agricultores en cultivo de cacao en México. Kuxulkab’ Revista de Divulgación XVIII (34):95–102. http://www.revistas.ujat.mx/index.php/kuxulkab/article/view/249
Acknowledgements
We thank comments provided by three anonymous reviewers, which highly improved the quality of the manuscript. To Nestle Mexico, S.A. de C.V. for the financial support, project: rescue, morphological and genetic characterization of wild and domesticated populations-abandoned pre-Hispanic possible origin of Theobroma cacao L., in Mexico. The engineers Felix Rodriguez and Ruben Jimenez Moran for their support in the field and to all farmers who provided their time and expertise. Figure 1 is prepared by Adriana Sandoval-Comte.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Zequeira-Larios, C., Santiago-Alarcon, D., MacGregor-Fors, I. et al. Tree diversity and composition in Mexican traditional smallholder cocoa agroforestry systems. Agroforest Syst 95, 1589–1602 (2021). https://doi.org/10.1007/s10457-021-00673-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10457-021-00673-z