
Abstract
Biological pest control in greenhouses generally depends on the release of natural enemies. Macrolophus pygmaeus (Hemiptera: Miridae) is one of the most used predators in European vegetable crops. However, its long establishment time is considered as one of the main drawbacks in regard to pest control. Pre-plant release in nurseries and the use of banker plants are strategies frequently used to enhance the response of predatory mirids. The aim of this work was to determine how the length of the pre-plant release period in banker plants (PRP) and banker plant species influenced the establishment and population dynamics of M. pygmaeus and its pest control in tomato crops. Two PRPs (24 h and 30 days) and two banker plants (tomato, Solanum lycopersicum—Solanaceae, and Ballota hirsuta—Lamiaceae) were assayed in a complete factorial design with three replicates, in individual greenhouses. The proportion of individuals established, founder population, abundance and population growth rate of M. pygmaeus were significantly higher for the long than for the short PRP. For the long PRP, the founder population and abundance of M. pygmaeus in greenhouses were higher when B. hirsuta, rather than tomato, was used as banker plant. Lower incidences of Bemisia tabaci (Hemiptera: Aleyrodidae) and Tuta absoluta (Lepidoptera: Gelechiidae) were registered in greenhouses with the long PRP, while the effect of the banker plant was not significant. The higher proportions of individuals established and higher population growth rates of M. pygmaeus in the greenhouses with the largest founder populations suggest the implication of Allee effects.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Key Message
-
Macrolophus pygmaeus is a key predator for greenhouse pests, but its long establishment limits its potential.
-
The effects of the pre-plant release period (PRP) and banker plants on the population dynamics and pest control of M. pygmaeus in tomato greenhouses were tested.
-
The abundance of M. pygmaeus in greenhouses was higher when B. hirsuta, rather than tomato, was used.
-
Lower incidences of Bemisia tabaci and Tuta absoluta were registered in greenhouses with the long PRP.
Introduction
The colonization of new areas by arthropods generally starts from a low number of individuals, and there is a great risk of extinction due to stochasticity and Allee effects (Grevstad 1999a; Drake 2004; Simberloff 2009; Duncan et al. 2014). Climatic conditions, diet breadth, and the availability of host plants for herbivores and of hosts for parasitoids are some of the factors known to influence the establishment of insects in new areas (Stiling 1990; Bacon et al. 2014; Rossinelli and Bacher 2015). Several colonization events are usually needed for the establishment, with the probability of success depending on the size of the founding population and environmental factors (Simberloff 2009; Duncan et al. 2014). Small populations are very sensitive to stochasticity and Allee effects, and environmental conditions influence the probability of survival of newcomers and the potential population growth (Dennis 2002; Duncan et al. 2014). Many agricultural crops are temporary habitats which are periodically removed or replaced, or regularly manipulated by cultural practices. Regarding the establishment of insects that live on plants, temporary crops correspond very well to the concept of “Darwinian islands” (Gillespie and Roderick 2002), in the sense that the communities in both, crops and islands, depend on the colonization by and establishment of organisms from external sources. As in the case of islands, the colonization of crops is expected to depend on isolation and time, with greenhouse crops suffering from a greater degree of isolation than open-field crops. The isolation of greenhouses is intended to reduce the colonization of the crops within them by insect pests but, as a side-effect, this also limits the immigration of their natural enemies (Albajes and Alomar 1999). Although conservation biological control is being assayed and practiced by enhancing greenhouse colonization by naturally occurring parasitoids or predators, biological pest control in greenhouses generally depends on the introduction and successful establishment of pest antagonists (van Lenteren 2012; Messelink et al. 2014). Several releases are generally needed before natural enemies become established and their populations start to build up (van Lenteren 2012). This reiteration of releases acts as a source of individuals that sustain the initial population and facilitate the establishment of the species (Lockwood et al. 2005).
Several species of predatory hemipterans are released as biological pest control agents in greenhouses (van Lenteren 2012). Macrolophus pygmaeus (Rambur) (Hemiptera: Miridae), Orius laevigatus (Fieber) (Hemiptera: Anthocoridae), and Nesidiocoris tenuis (Reuter) (Hemiptera: Miridae) are probably the predatory hemipterans released most for pest control in European vegetable greenhouses (Castañé et al. 2011; van Lenteren 2012). Several Macrolophus species, such as M. pygmaeus and Macrolophus caliginosus Wagner—syn. Macrolophus melanotoma (Costa)-(Hemiptera: Miridae), have been used mainly for the control of whiteflies, although they may also prey on other small arthropods such as thrips, spider mites, leafminers, aphids, and lepidopterans (Alvarado et al. 1997; Barnadas et al. 1998; Riudavets and Castane 1998; Margaritopoulos et al. 2003; Urbaneja et al. 2012; Jaworski et al. 2013; Sylla et al. 2016; Sanchez et al. 2018). Although several Macrolophus species have been reported as biological control agents in tomato crops, it is quite likely that all these citations refer to a single species, M. pygmaeus (Martínez-Cascales et al. 2006; Castañé et al. 2013).
The long establishment time of predatory mirids has been considered as one of their main drawbacks with regard to pest control (Lenfant et al. 2000; Fischer and Terrettaz 2003; Calvo et al. 2012b; Trottin-Caudal et al. 2012; De Backer et al. 2014; Nannini et al. 2014). The presence of prey and the existence of the right host plants are two of the main factors that may influence the establishment of omnivorous mirids in new areas. Most predatory mirids have low development and reproduction rates when feeding only on plants (Perdikis and Lykouressis 2000; Sanchez et al. 2004; Perdikis et al. 2007; Lykouressis et al. 2008; Ingegno et al. 2011; Mollá et al. 2014; Biondi et al. 2016; Sanchez et al. 2016). In addition, omnivorous mirids are very restricted in their host preference (Sanchez et al. 2004; Ingegno et al. 2011); therefore, their colonization of new habitats and their performance may greatly depend on the presence of the right host plants (Naselli et al. 2016). Several release strategies aimed at improving the establishment of mirids have been assayed, such as: (1) the release on seedlings in nurseries—pre-plant release (Lenfant et al. 2000; Fischer and Terrettaz 2003; Calvo et al. 2012a, b; Nannini et al. 2014; Perdikis et al. 2015); (2) the confinement on plants by the use of netting during the establishment period (Lenfant et al. 2000; Fischer and Terrettaz 2003; Nannini et al. 2014); (3) the use of banker plants (Arnó et al. 2000; Sanchez et al. 2003a; Frank 2010; Huang et al. 2011; Bresch et al. 2014; Biondi et al. 2016; Balzan 2017); (4) the addition of factitious prey or sugar to the crop (Put et al. 2012; Urbaneja-Bernat et al. 2013).
In spite of the release of omnivorous mirids being a common practice in many greenhouse vegetable crops grown with biological pest control, very little is known about the mechanisms governing their establishment. Adaptation and host plants are two factors known to influence the establishment and population dynamics of insects in new places (Kim and McPheron 1993; Bacon et al. 2014). The aim of this work was to determine how the length of the pre-plant release period (PRP) in banker plants (i.e., as a way of adaptation) and the use of different banker plant species influenced the establishment, population dynamics, and pest control response of M. pygmaeus in greenhouse tomato crops. Ballota hirsuta Bentham (Lamiaceae) was assayed as banker plant because it is a natural host on which M. pygmaeus usually reaches high populations in natural conditions (Sanchez et al. 2003b; Martínez-Cascales et al. 2006). Ballota hirsuta was tested in relation to tomato—Solanum lycopersicum L. (Solanaceae)—as banker plant, to determine whether a banker plant species with characteristics different from those of the crop (e.g., hairiness and volatile profile—Ingegno et al. 2016), and for which the preference of M. pygmaeus could differ, influenced the establishment and population dynamics of the mirid.
Materials and methods
Crop setting
The assay was carried out from 14 February to 5 June 2018 in 12 polycarbonate greenhouses (5 × 10 m each), with individual meshed openings at the front and on top for ventilation (10 × 14 threads/cm), at the IMIDA Torreblanca research station in Campo de Cartagena, Murcia (Spain). The temperature was controlled independently in each greenhouse by the Mithra Clima System (Nutricontrol, S.L., Cartagena, Murcia); openings were activated automatically at 20 °C and no heating or artificial lighting was provided. The temperature and relative humidity were recorded by the system every minute. In each greenhouse, four rows of 20 tomato plants (cv. Optima, Seminis, Murcia) were transplanted into soil on 14 February 2018. The separation of plants within and between rows was 0.5 and 1 m, respectively. The plants were 45 days old and approximately 20–25 cm tall at transplanting. They were watered and fertilized when needed, by drip irrigation. Insecticides and fungicides were not applied during the trial.
Experimental design and sampling of insects
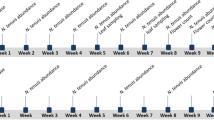
The effect of the length of the PRP in banker plants and the banker plant species on the population dynamics of M. pygmaeus in a tomato crop was tested in a complete factorial design, with the PRP at two levels (i.e., short—24 h—and long—30 days) and two banker plant species (tomato—cv. Optima—and B. hirsuta). Each combination of the two PRP levels and the two plant species was replicated three times in individual greenhouses, following a randomized block design. The protocol of the experiment is summarized in Fig. 1. Individuals of M. pygmaeus (320 adults, approx. 7–10 days old, half females and half males), provided by Agroscience (Essex, UK), were released into each of the six wooden-frame muslin cages containing 16 tomato plants (approx. 30–40 cm tall) and each of the six cages containing 16 B. hirsuta plants (approx. 30–40 cm tall). The cages with M. pygmaeus and the plants were kept in walk-in climatic chambers at 25 ± 2 °C, 60–70% RH, and a 16L:8D photoperiod. In the long PRP treatment, M. pygmaeus adults were released into the cages 30 days before the transplanting of tomato and B. hirsuta plants in their respective greenhouses. Sixteen banker plants of either tomato or B. hirsuta were transplanted in each greenhouse, being interspaced regularly among the 80 tomato plants already present (i.e., four plants in each of the four rows). For the short PRP, M. pygmaeus was released into the cages 24 h prior to the transplanting of tomato and B. hirsuta. Each cage was supplied with 24 g of Ephestia kuehniella Zeller (Lepidoptera: Pyralidae) eggs on the first day; thereafter, the cages of the long PRP period were supplied with 24 g of E. kuehniella eggs every 3–4 days. In the cages of the long PRP treatment the number of M. pygmaeus was counted on six plants per cage before their transplanting in the greenhouse. The total number of M. pygmaeus mobile forms (nymphs and adults) present on the banker plants at the time of their introduction into the greenhouse was estimated as 306 ± 47 and 671 ± 31 (mean ± SE) individuals for the tomato and B. hirsuta plants, respectively. In the cages of the short PRP treatment M. pygmaeus was not counted because it was supposed that the numbers had varied very little since its introduction 24 h before; the careful inspection of plants and cages at transplanting revealed no signs of mortality. In all the treatments the plants with the mirids were transplanted in the greenhouses on 27 March 2018; in the 2 weeks following this the banker plants were supplemented twice with 5 g of E. kuehniella eggs per greenhouse. Bemisia tabaci Gennadius (Hemiptera: Aleyrodidae) (400 adults, approx. 1 week old, per greenhouse and date of release) was introduced on 13 and 21 March 2018. Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae) was not released but colonized naturally all the greenhouses.

Protocol of the experiment to test the effect of banker plants and the length of pre-plant release periods (PRP) on the establishment and population dynamics of M. pygmaeus in tomato greenhouses
In each greenhouse, the number of M. pygmaeus (nymphs and adults) was counted visually on 15 whole tomato crop plants and on six whole banker plants (i.e., tomato or B. hirsuta) selected at random. The total number of M. pygmaeus on the tomato crop and banker plants was estimated by multiplying the average number of M. pygmaeus per plant by the number of tomato crop plants or banker plants, as appropriate. The total number of M. pygmaeus in each greenhouse was calculated as the sum of the number of individuals on the tomato crop plants plus those on the bankers. For the sampling of B. tabaci, three leaves—one from the top, middle, and bottom part- of each of the 15 tomato crop plants and five leaves from each of the six banker plants were inspected for the counting of the nymphs and adults. Tuta absoluta galleries were counted in all the leaves of each of the 15 whole tomato crop plants. All samplings were carried out with the help of a magnifying glass (1.8–4.8 x). Six samplings were carried out, approximately every 10 days, from 8 days after the transplanting of the banker plants with M. pygmaeus until the end of the assay.
Statistical analyses
For each greenhouse, the estimate of the total number of M. pygmaeus 8 days after their introduction into the greenhouses was considered as the founder population. In general, there is a high dispersion of individuals after releases and 8 days was considered as a reasonable period of time for the dispersion rate to stabilize. The proportion of M. pygmaeus established was calculated by dividing the size of the founder population by the number of M. pygmaeus introduced in each greenhouse. The effects of the length of the PRP and the banker plant species on both the size of the founder population and the proportion of M. pygmaeus established were tested by a two-way ANOVA using the function “aov”, and the separation of the means was performed with “Tukey HSD”, both functions being available in the “stats” package (R-Development-Core-Team 2017).
Generalized linear mixed models (GLMM) were used to test the effects of the PRP and banker plant on the number of M. pygmaeus in the greenhouses throughout the duration of the experiment. This variable was found to be normally distributed; thus, the “lmer” function (“lme4” package) was used to perform the analyses (Bates et al. 2015). GLMM were also the models of choice to test the effects of the PRP and banker plant on the overall proportion of M. pygmaeus found in the tomato crop. In this case the data fitted log-normal distributions and their analyses were performed using “glmmPQL” (“MASS” package) (Venables and Ripley 2002) set to the Gaussian distribution with the link “log” (R-Development-Core-Team 2017). In these two cases, date of sampling was introduced in the models as a random factor. The contrast among the treatments was tested using “Tukey” with the function “glht” in the “multcomp” package (Hothorn et al. 2008).
The logistic function (1) (Smith 1980) was used to calculate the population growth rate from the introduction of M. pygmaeus into the greenhouses until the time at which the population peak was reached, for each greenhouse independently.
where Nt is the total number of M. pygmaeus in the greenhouse at time t, k is the carrying capacity, a = (k − No)/No (No being the number of M. pygmaeus in the greenhouse at time t = 0 as estimated by the model), and r is the population growth rate of M. pygmaeus. Because all the greenhouses had the same characteristics, we assumed the same carrying capacity for all of them. Some preliminary analyses were performed to estimate k independently for each greenhouse and the highest value (k = 22,444) was included in the models to estimate No and r in the following analyses. This k value was considered realistic as in our case (i.e., 80 plants per greenhouse) it is equivalent to 280 individuals per plant (14 individuals per leaf in a plant with 20 leaves). The effects of the PRP and banker plants on the population growth rate were tested by a two-way ANOVA, using the function “aov” (R-Development-Core-Team 2017). The Tukey test, as explained above, was used for the separation of the means.
GLMM were also used to compare the number of B. tabaci (nymphs and adults) and the number of T. absoluta galleries per plant among treatments. PRP and banker plant were introduced in the models as fixed factors and date of sampling as a random factor. The numbers of B. tabaci (nymphs and adults) per leaf fitted log-normal distributions; accordingly, their analyses were performed with the “glmmPQL” function (“MASS” package) (R-Development-Core-Team 2017), using the Gaussian family with the log normal link. The data for T. absoluta galleries fitted a negative binomial distribution; thus, the “glmmPQL” function (“MASS” package) set to the negative binomial family was used to perform this analysis. The contrast among treatments was tested using the function “glht”, in the same way as in the analyses of the population dynamics of M. pygmaeus.
Results
Effect of pre-plant release and banker plants on M. pygmaeus establishment and population dynamics
The size of the founder population was greater in the greenhouses with 30 days of pre-plant release of M. pygmaeus in banker plants (PRP) than in those with one day (F = 169.9; df. 1, 8; P < 0.001), the effect of the banker plant being significant also (F = 75.9; df. 1, 8; P < 0.001) (Fig. 2a). A significant interaction was denoted between the PRP and banker plant (F = 56.2; df. 1, 8; P < 0.001). This interaction means that no significant differences in the size of the founder population were registered between the greenhouses with either B. hirsuta or tomato as the banker plant when the PRP was 1 day (Tukey test, P = 0.825), while when the PRP was 30 days the founder population in greenhouses with B. hirsuta was bigger than in those with tomato as banker plant (Tukey test, P < 0.001). In addition, no significant differences in the founder population were found between greenhouses with B. hirsuta and 1 day of PRP and those with tomato—as the banker plant—and 30 days of PRP (Tukey test, P = 0.061) (Fig. 2a). The length of the PRP had a significant effect on the proportion of M. pygmaeus established in the greenhouses (F = 40.6; df. 1, 8; P < 0.001), being higher with 30 days than with 1 day (Fig. 2b). In contrast, the banker plant did not have a significant effect (F = 2.97; df. 1, 8; P = 0.123), and the interaction between the PRP and the banker plant was not significant (F = 0.407; df. 1, 8; P = 0.541).

Size of the founder population—number of M. pygmaeus in the greenhouses 8 days after its introduction (mean ± SE) (a) and proportion of Macrolophus pygmaeus established—founder population/number of M. pygmaeus introduced (mean ± SE) (b) in tomato greenhouses with short (1 day) and long (30 days) pre-plant release periods (PRP) for M. pygmaeus and two banker plants (Ballota hirsuta and tomato). The same letter at the top of bars indicates no significant differences on the mean values (Tukey test, P > 0.05)
Overall, a higher number of M. pygmaeus was registered throughout the duration of the experiment in the greenhouses with 30 days of PRP than in those with only 1 day, independently of the banker plant. At long PRP, the abundance of M. pygmaeus increased progressively until day 51, reaching density peaks of 2662 ± 181 and 1916 ± 36 individuals in greenhouses with B. hirsuta and tomato as the banker plant, respectively (Fig. 3a). At short PRP, the abundance of M. pygmaeus was lower, reaching at day 61 peaks of 1112 ± 159 and 952 ± 163 individuals in the greenhouses with tomato and B. hirsuta as the banker plant, respectively (Fig. 3a). The total number of M. pygmaeus in the greenhouses was significantly influenced by the PRP period (χ2 = 70.7; df. 1; P < 0.001) and banker plant (χ2 = 5.61; df. 1; P = 0.018). Besides, there was a significant interaction between the PRP period and banker plant (χ2 = 4.90; df. 1; P = 0.027). This means that no significant differences were found in the population dynamics of M. pygmaeus between greenhouses with tomato as the banker plant and those with B. hirsuta when the PRP period was 1 day (Tukey contrast, P = 0.9995), while significant differences were found when the PRP was 30 days (Tukey contrast, P = 0.006). The logistic models described satisfactorily the population dynamics of M. pygmaeus in the 12 greenhouses (Fig. 3a) (Supplementary material, Table 1). The population growth rate of M. pygmaeus was significantly higher (F = 12.5; df. 1, 8; P = 0.008) for the 30-day (B. hirsuta: 0.018 ± 0.001; tomato: 0.017 ± 0.001) than for the 1-day PRP (B. hirsuta: 0.011 ± 0.002; tomato: 0.013 ± 0.002); in contrast, the population growth rate was not significantly influenced by the factor banker plant (F = 0.780; df. 1, 8; P = 0.403).

Population dynamics (a) and proportion of individuals in the crop (b) (mean ± SE) for Macrolophus pygmaeus in tomato greenhouses with short (1 day) and long (30 days) pre-plant release periods (PRP) for M. pygmaeus and two banker plants (Ballota hirsuta and tomato). Time in days since the introduction of the banker plants with the M. pygmaeus in the greenhouses. In graph A, the dots represent the experimental data and the lines the predictions according to the logistic function (1)
In the first week after the introduction of M. pygmaeus in the greenhouses, the proportion of M. pygmaeus was higher in the greenhouses with tomato as the banker plant (Short PRP: 0.863 ± 0.039; long PRP: 0.909 ± 0.027) than in those with B. hirsuta (Short PRP: 0.744 ± 0.131; long PRP: 0.651 ± 0.075) (Fig. 3b). 3 weeks after the introduction of M. pygmaeus in the greenhouse, the proportion of M. pygmaeus in the tomato crop in all the treatments, with the exception of that with short PRP and B. hirsuta, was above 0.95 (Fig. 3b). The banker plant had a significant effect on the proportion of M. pygmaeus registered on the tomato crops (χ2 = 8.77; df. 1; P = 0.003). In contrast, the PRP (χ2 = 0.284; df. 1; P = 0.594) and its interaction with the banker plant (χ2 = 0.607; df. 1; P < 0.436) were not found to influence significantly the proportion of M. pygmaeus on the tomato crops.
Abundance of phytophagous insects
The highest abundance of B. tabaci was registered at the short PRP, with density peaks of 1.22 ± 0.45 and 1.49 ± 0.34 adults per leaf in greenhouses with B. hirsuta and tomato as the banker plant, respectively (Fig. 4a); similar peaks of abundance of 14.8 ± 2.6 and 14.2 ± 3.3 nymphs per leaf were reached in greenhouses with tomato and B. hirsuta as the banker plant, respectively (Fig. 4b). The abundance of B. tabaci at the long PRP was lower than at the short PRP, with density peaks of 0.491 ± 0.140 and 0.526 ± 0.155 adults per leaf (Fig. 4a), and 5.78 ± 0.63 and 2.99 ± 0.53 nymphs per leaf (Fig. 4b) in greenhouses with tomato and B. hirsuta as the banker plant, respectively. The abundance of B. tabaci was significantly higher in greenhouses with the short M. pygmaeus PRP than in those with the long PRP (Adults: χ2 = 24.9; df. 1; P < 0.001; nymphs: χ2 = 64.6; df. 1; P < 0.001). In contrast, the banker plant—tomato or B. hirsuta—did not significantly affect the abundance of B. tabaci (Adults: χ2 = 1.237; df. 1; P = 0.266; nymphs: χ2 = 1.678; df. 1; P = 0.195). The interaction between the PRP and the banker plant was not significant (Adults: χ2 = 0.310; df. 1; P = 0.578; nymphs: χ2 = 3.480; df. 1; P = 0.062). A low abundance of B. tabaci was registered in banker plants, being the density higher in tomato (short PRP: 0.213 ± 0.040; long PRP: 0.169 ± 0.054 adults + nymphs per leaf—overall average) than in B. hirsuta (short PRP: 0.011 ± 0.011; long PRP: 0.004 ± 0.004 adults + nymphs per leaf).

Population dynamics of Bemisia tabaci adults (a) and nymphs (b) per leaf (mean ± SE) in tomato greenhouses with short (1 day) and long (30 days) pre-plant release periods (PRP) for Macrolophus pygmaeus and two banker plants (Ballota hirsuta and tomato). Time in days since the introduction of the banker plants with the M. pygmaeus in the greenhouses
The highest abundance of T. absoluta was registered at the short PRP, with density peaks of 31.0 ± 12.5 and 15.3 ± 5.0 galleries per plant in greenhouses with tomato and B. hirsuta as the banker plant, respectively (Fig. 5). The lowest abundance was registered at long PRP with density peaks of 1.00 ± 0.56 and 3.19 ± 1.50 galleries per plant in greenhouses with tomato and B. hirsuta as the banker plant, respectively (Fig. 5). The number of T. absoluta galleries was significantly influenced by both the PRP (χ2 = 49.0; df. 1; P < 0.001) and the banker plant (Adults: χ2 = 7.36; df. 1; P = 0.007), with a significant interaction between them (Adults: χ2 = 19.3; df. 1; P < 0.001). Significant differences in the number of galleries per plant were found between the greenhouses with B. hirsuta and tomato as bankers at long but not at short PRP (Tukey contrast, P < 0.001). No galleries of T. absoluta were registered in the banker plants.

Number of galleries per plant of Tuta absoluta (mean ± SE) in tomato greenhouses with short (1 day) and long (30 days) pre-plant release periods (PRP) for Macrolophus pygmaeus and two banker plants (Ballota hirsuta and tomato). Time in days since the introduction of the banker plants with the M. pygmaeus in the greenhouses
Discussion
The establishment of insects in new areas is a probabilistic event which depends, among other variables, on the biology of the species, intra- and inter-specific interactions, the size of the founder population, the environment, and stochastic factors (Simberloff 2009; Bacon et al. 2014; Duncan et al. 2014). In this work, it has been demonstrated that the establishment and population dynamics of the predatory mirid M. pygmaeus in tomato greenhouses depend on the size of the founder population and the banker plant species. A long PRP (i.e. 30 days), with M. pygmaeus confined in cages with banker plants supplemented with E. kueniella eggs, produced an increase in the populations before their introduction into the greenhouses. Also, larger founder populations were achieved using B. hirsuta, rather than tomato, as the banker plant. These differences in the founder population size were reflected in the population dynamics: the greenhouses with the highest initial numbers reached the highest population peaks. Besides, the higher proportions of individuals established and the higher population growth rates of M. pygmaeus in the greenhouses with the largest initial populations suggest the implication of Allee effects.
Pre-plant release of mirids has previously been reported to give better results than classical direct introduction but until now the mechanisms implicated in the establishment process have remained largely unexplored. The release of N. tenuis in nurseries several days before the transplanting of the crop is known to enhance its establishment in tomato greenhouses (Calvo et al. 2012a, b; Perdikis et al. 2015), and this is currently a strategy adopted by professional growers in southern Spain. This strategy has also been assayed, with good results, for the establishment of M. caliginosus in tomato greenhouses (Lenfant et al. 2000; Fischer and Terrettaz 2003). Pre-plant release strategies may operate at two levels. Firstly, by improving the probability of establishment when the mirids are introduced into greenhouses; populations that are already established may have lower dispersal rates than those that are introduced without previous adaptation into new environments (Kim and McPheron 1993). In this regard, the confinement of Macrolophus species (i.e., M. pygmaeus and M. caliginosus) with nettings that reduce the dispersal of the insects after their release has been reported to give better results than classical direct introductions (Lenfant et al. 2000; Fischer and Terrettaz 2003; Nannini et al. 2014). Secondly, pre-plant release strategies allow mirids to increase their initial population by reproducing in an environment that is more optimal than the one to which they will be transferred. Bigger founding populations are known to establish more successfully than smaller ones, among other reasons, because the impact of stochasticity and the Allee effects are stronger on small than on big populations (Grevstad 1999a, b; Dennis 2002; Lockwood et al. 2005; Simberloff 2009; Duncan et al. 2014). Allee effects may result from several processes such as a decline in the fecundity of females if finding mate gets difficult at low population densities (Courchamp et al. 2008; Duncan et al. 2014). Gemeno et al. (2007) reported that the encounters between females and males of M. caliginosus were apparently random, with most of the females accepting mating after several contacts. This behavior could result in limited encounters between the two sexes and reduced reproduction rates in small populations.
The population dynamics of M. pygmaeus were also influenced by the species of banker plant. However, differences between greenhouses with B. hirsuta and those with tomato as the banker plants were only registered for the long PRP. In this case, greenhouses into which M. pygmaeus was introduced on B. hirsuta had bigger founder populations and reached higher population peaks than those with tomato as the banker plant. The absence of differences in the population growth rate indicates that the variations in the population dynamics between the greenhouses with B. hirsuta and tomato as the banker plants were mainly due to the size of the founder population. In fact, the number of M. pygmaeus on B. hirsuta after the 30 days of PRP and prior to their into the greenhouses was three times that on tomato seedlings. During the PRP, the mirids were fed with the same amount of factitious prey (i.e., E. kuehniella eggs); thus, the variations that the populations experienced during the 30 days of PRP were likely due to the performance (e.g., development rates and/or fecundity) of the mirid on the different plant species. Great variations in the fitness of omnivorous dicyphines have been registered depending on the host plant species, both in the presence and absence of prey (Sanchez et al. 2004; Ingegno et al. 2011; Biondi et al. 2016). Several factors may account for this variation in performance in relation to the plant species. For instance, the hardness of plant tissues is known to limit the oviposition in species such as M. caliginosus, on Pelargonium peltatum (L.) (Geraniaceae) (Constant et al. 1996), and D. hesperus, on corn (Sanchez et al. 2004). Differences may also arise from variations in the specific nutritional values of plants (Sanchez et al. 2004, 2016; Ingegno et al. 2011). Plant structure may also influence population growth rates (Wheeler and Krimmel 2015). For example, Tupiocoris notatus (Distant) (Hemiptera: Miridae) showed better nymphal performance and oviposition rates, which led to higher population growth rates, on glandular than on non-glandular types of Datura wrightii (Regel) (Solanaceae) (Van Dam and Hare 1998).
Other authors have also assayed the use of host plants to enhance the response of dicyphines in tomato crops. The use of mullein as an alternative host plant is known to contribute to the early establishment of D. hesperus in tomato greenhouses and to the maintenance of the population of the predator after the prey is depleted in the crop (Sanchez et al. 2003a). It has been proven several times that tobacco (Nicotiana tabacum L.—Solanaceae) enhances the early establishment of M. caliginosus and its conservation during crop-free periods in tomato greenhouses (Arnó et al. 2000; Fischer and Terrettaz 2003; Bresch et al. 2014). However, the use of tobacco has not been recommended because it serves as an optimum host for the multiplication of the whitefly B. tabaci and because of the risk of virus transmission (Bresch et al. 2014). Tamarillo [Cyphomandra betacea (Cav.) Sendtn.—Solanaceae], as banker for M. caliginosus, has also been reported to provide shorter establishment times and quicker numerical responses in comparison with classical direct releases (Fischer and Terrettaz 2003).
One aspect to be considered in relation to the banker system is that highly preferred host plants could act as sinks for predatory mirids (Sanchez et al. 2003a). In the present work, during the first part of the assay, in greenhouses with B. hirsuta a lower proportion of M. pygmaeus was registered in the crop in comparison with those greenhouses with tomato as banker; this suggests a higher preference of M. pygmaeus for B. hirsuta than for tomato, which could interfere with the colonization of the crop. However, this effect seems to have been patent mainly in the first weeks after the introduction of M. pygmaeus into the greenhouses and at low abundances; 3 weeks after the introduction, the proportion of the mirid in the crop in greenhouses with B. hirsuta was similar to that in greenhouses with tomato as banker.
The abundance of B. tabaci and T. absoluta in the tomato crops was inversely related to that of M. pygmaeus. Lower abundances of B. tabaci and T. absoluta were registered in the greenhouses with the long PRP, which were those with the highest M. pygmaeus abundances. The degree of control of B. tabaci was highest in the greenhouses with B. hirsuta and the long PRP; however, no significant differences were found in comparison to the greenhouses with tomato as the banker plant and the long PRP. Macrolophus species have been extensively reported as predators of whiteflies, with a great impact on their population dynamics (Barnadas et al. 1998; Albajes and Alomar 1999; Perdikis and Lykouressis 2002; Alomar et al. 2006; Lykouressis et al. 2009; Sylla et al. 2016; Sanchez et al. 2018; Ferracini et al. 2019). These mirids have also been cited among the main predators of T. absoluta in the Mediterranean area (Arnó et al. 2009; Mollá et al. 2009; Urbaneja et al. 2009; Biondi et al. 2018; Jaworski et al. 2013; Zappala et al. 2013; Chailleux et al. 2013; Nannini et al. 2014; Sylla et al. 2016; Sanchez et al. 2018). Earlier studies reported that the pre-planting release of N. tenuis in nurseries provided satisfactory control of both B. tabaci and T. absoluta in tomato crops (Calvo et al. 2012a, b). The use of banker plants is also known to support the population of predatory mirids, with an impact on pest control. For instance, tobacco, as a banker plant for M. pygmaeus, contributed to the control of T. vaporariorum in tomato plants (Bresch et al. 2014). Sanchez et al. (2003a) argued that mullein plants serve as a reservoir of D. hesperus that enhances the performance of D. hesperus for the control of whitefly in tomato greenhouses.
In this work we provide some insights into the processes behind the establishment of omnivorous dicyphines (i.e., M. pygmaeus) in tomato crops. The effects of the length of the PRP and the impact of natural hosts (i.e., B. hirsuta), used as banker plants, on the establishment and population dynamics of M. pygmaeus were investigated. Pre-plant release may influence the establishment in crops in distinct ways: (1) by increasing the size of the founder population during the PRP; (2) by increasing the probability that individuals settle in crops, most likely because long PRP on banker plants serve as a period of adaptation that reduces the dispersal of individuals when they are introduced into greenhouses. It has to be taken into account also that, in this case, part of the M. pygmaeus introduced into the greenhouses was in the form of nymphs, which could have lower dispersion rates than adults. The banker plants also had an effect on the establishment and population dynamics of M. pygmaeus, but this effect can be attributed mainly to the bigger population size achieved in B. hirsuta, relative to tomato plants, during the PRP. Bigger initial populations led to higher population growth rates and high population peaks, suggesting the implication of stochastic and Allee effects. The use of B. hirsuta for the PRP of M. pygmaeus prior to its introduction into tomato crops may reduce the cost of and enhance biological pest control in commercial tomato greenhouses. The fact that part of the M. pygmaeus population resides on the natural host, and its apparent higher preference for B. hirsuta, may help to preserve the mirid during adverse periods (e.g., winter, absence of crop, prey scarcity, etc.). If evidence of a sink effect is provided, this may be overcome by cutting and removing the banker plants to force the mirids to migrate to the crop. In relation to the role of banker plants in harboring tomato pests, a very low number of B. tabaci was registered in B. hirsuta plants.
Author contribution
JAS conceived and designed the research, analyzed the data, and wrote the manuscript. JAS, ELG, MPM, and LPF conducted experiments. All authors read and approved the manuscript
References
Albajes R, Alomar O (1999) Current and potential use of polyphagous predators. In: Albajes R, Gullino ML, Van Lenteren JC, Elad Y (eds) Integrated pest and disease management in greenhouse crops. Kluwer Academic Publishers, Dordrecht, pp 265–275
Alomar O, Riudavets J, Castane C (2006) Macrolophus caliginosus in the biological control of Bemisia tabaci on greenhouse melons. Biol Control 36:154–162. https://doi.org/10.1016/j.biocontrol.2005.08.010
Alvarado P, Balta O, Alomar O (1997) Efficiency of four Heteroptera as predators of Aphis gossypii and Macrosiphum euphorbiae (Hom.: Aphididae). Entomophaga 42:215–226
Arnó J, Ariño J, Español R et al (2000) Conservation of Macrolophus caliginosus Wagner (Het.Miridae) in commercial greenhouses during tomato crop-free periods. IOBC wprs Bull 23:241–246
Arnó J, Sorribas R, Prat M et al (2009) Tuta absoluta, a new pest in IPM tomatoes in the northeast of Spain. IOBC wprs Bull 49:203–208
Bacon SJ, Aebi A, Calanca P, Bacher S (2014) Quarantine arthropod invasions in Europe: the role of climate, hosts and propagule pressure. Divers Distrib 20:84–94. https://doi.org/10.1111/ddi.12149
Balzan MV (2017) Flowering banker plants for the delivery of multiple agroecosystem services. Arthropod Plant Interact 11:743–754. https://doi.org/10.1007/s11829-017-9544-2
Barnadas I, Gabarra R, Albajes R (1998) Predatory capacity of two mirid bugs preying on Bemisia tabaci. Entomol Exp Appl 86:215–219. https://doi.org/10.1023/A:1003197320977
Bates D, Maechler M, Bolker B, Walker S (2015) Fitting linear mixed-effects models using lme4. J Stat Softw 67:1–48. https://doi.org/10.18637/jss.v067.i01
Biondi A, Zappalà L, Di Mauro A et al (2016) Can alternative host plant and prey affect phytophagy and biological control by the zoophytophagous mirid Nesidiocoris tenuis? Biocontrol 61:79–90. https://doi.org/10.1007/s10526-015-9700-5
Biondi A, Guedes RNC, Wan F-H, Desneux N (2018) Ecology, worldwide spread, and management of the Invasive South American Tomato Pinworm, Tuta absoluta: past, present, and future. Annu Rev Entomol 63:239–258. https://doi.org/10.1146/annurev-ento-031616-034933
Bresch C, Ottenwalder L, Poncet C, Parolin P (2014) Tobacco as Banker Plant for Macrolophus Pygmaeus to control Trialeurodes Vaporariorum in tomato crops. Univers J Agric Res 2:297–304. https://doi.org/10.13189/ujar.2014.020803
Calvo FJ, Bolckmans K, Belda JE (2012a) Release rate for a pre-plant application of Nesidiocoris tenuis for Bemisia tabaci control in tomato. Biocontrol 57:809–817. https://doi.org/10.1007/s10526-012-9455-1
Calvo FJ, Lorente MJ, Stansly PA, Belda JE (2012b) Preplant release of Nesidiocoris tenuis and supplementary tactics for control of Tuta absoluta and Bemisa tabaci in greenhouse tomato. Entomol Exp Appl 143:111–119. https://doi.org/10.1111/j.1570-7458.2012.01238.x
Castañé C, Arnó J, Gabarra R, Alomar O (2011) Plant damage to vegetable crops by zoophytophagous mirid predators. Biol Control 59:22–29. https://doi.org/10.1016/j.biocontrol.2011.03.007
Castañé C, Agustí N, Arnó J et al (2013) Taxonomic identification of Macrolophus pygmaeus and Macrolophus melanotoma based on morphometry and molecular markers. Bull Entomol Res 103:204–2015. https://doi.org/10.1017/S0007485312000545
Chailleux A, Bearez P, Pizzol J et al (2013) Potential for combined use of parasitoids and generalist predators for biological control of the key invasive tomato pest Tuta absoluta. J Pest Sci 86:533–541. https://doi.org/10.1007/s10340-013-0498-6
Constant B, Grenier S, Febvay G, Bonnot G (1996) Host plant hardness in oviposition of Macrolophus caliginosus (Hemiptera: Miridae). J Econ Entomol 89:1446–1452
Courchamp F, Berec L, Gascoigne J (2008) Allee effects in ecology and conservation. Oxford University Press, New York
De Backer L, Megido RC, Haubruge E, Verheggen FJ (2014) Macrolophus pygmaeus (Rambur) as an efficient predator of the tomato leafminer Tuta absoluta (Meyrick) in Europe. A review. Biotechnol Agron Soc Environ 18:536–543
Dennis B (2002) Allee effects in stochastic populations. Oikos 96:389–401
Drake JM (2004) Allee effects and the risk of biological invasion. Risk Anal 24:789–802
Duncan RP, Blackburn TM, Rossinelli S, Bacher S (2014) Quantifying invasion risk: the relationship between establishment probability and founding population size. Methods Ecol Evol 5:1255–1263. https://doi.org/10.1111/2041-210X.12288
Ferracini C, Bueno VHP, Dindo ML, Ingegno BL, Luna MG, Salas-Gervassio NG, Sánchez SE, Siscaro G, van Lenteren JC, Zappalà L, Tavella L (2019) Natural enemies of Tuta absoluta in the Mediterranean basin, Europe and South America. Biocontrol Sci Technol 29:578–609. https://doi.org/10.1080/09583157.2019.1572711
Fischer S, Terrettaz C (2003) Release strategies of the Mirid Macrolophus caliginosus in protected tomato crops. Rev suisse Vitic Arboric Hortic 35:191–196
Frank SD (2010) Biological control of arthropod pests using banker plant systems: past progress and future directions. Biol Control 52:8–16. https://doi.org/10.1016/j.biocontrol.2009.09.011
Gemeno C, Alomar O, Riudavets J, Castañé C (2007) Mating periodicity and post-mating refractory period in the zoophytophagous plant bug Macrolophus caliginosus (Heteroptera: Miridae). Eur J Entomol 104:715–720
Gillespie RG, Roderick GK (2002) Arthropods on islands: colonization, speciation, and conservation. Annu Rev Entomol 47:595–632. https://doi.org/10.1146/annurev.ento.47.091201.145244
Grevstad FS (1999a) Factors influencing the chance of population establishment: implications for release strategies in biocontrol. Ecol Appl 9:1439. https://doi.org/10.2307/2641408
Grevstad FS (1999b) Experimental invasions using biological control introductions: the influence of release size on the chance of population establishment. Biol Invasions 1:313–323. https://doi.org/10.1023/A:1010037912369
Hothorn T, Bret F, Westfall P (2008) Simultaneous inference in general parametric models. Biom J 50:346–363. https://doi.org/10.1002/bimj.200810425
Huang N, Enkegaard A, Osborne LS et al (2011) The banker plant method in biological control. CRC Crit Rev Plant Sci 30:259–278. https://doi.org/10.1080/07352689.2011.572055
Ingegno BL, Pansa MG, Tavella L (2011) Plant preference in the zoophytophagous generalist predator Macrolophus pygmaeus (Heteroptera: Miridae). Biol Control 58:174–181. https://doi.org/10.1016/j.biocontrol.2011.06.003
Ingegno BL, La-Spina M, Jordan MJ, Tavella L, Sanchez JA (2016) Host plant perception and selection in the sibling species Macrolophus melanotoma and Macrolophus pygmaeus (Hemiptera: Miridae). J Insect Behav 29:117–142. https://doi.org/10.1007/s10905-016-9549-1
Jaworski CC, Bompard A, Genies L et al (2013) Preference and prey switching in a generalist predator attacking local and invasive alien pests. PLoS ONE 8:1–10. https://doi.org/10.1371/journal.pone.0082231
Kim KC, McPheron BA (1993) Evolution of insect pests: patterns of variation. Wiley, New York
Lenfant C, Ridray G, Schoen L (2000) Biopropagation of Macrolophus caliginosus Wagner for a quicker establishment in southern tomato greenhouses. IOBC wprs Bull 23:247–251
Lockwood JL, Cassey P, Blackburn T (2005) The role of propagule pressure in explaining species invasions. Trends Ecol Evol 20:223–228. https://doi.org/10.1016/j.tree.2005.02.004
Lykouressis D, Giatropoulos A, Perdikis D, Favas C (2008) Assessing the suitability of noncultivated plants and associated insect prey as food sources for the omnivorous predator Macrolophus pygmaeus (Hemiptera: Miridae). Biol Control 44:142–148. https://doi.org/10.1016/j.biocontrol.2007.11.003
Lykouressis DP, Perdikis DC, Konstantinou AD (2009) Predation rates of Macrolophus pygmaeus (Hemiptera: Miridae) on different densities of eggs and nymphal instars of the greenhouse whitefly Trialeurodes vaporariorum (Homoptera: Aleyrodidae). Entomol Gen 32:105–112
Margaritopoulos JT, Tsitsipis JA, and Perdikis DC (2003) Biological characteristics of the mirids Macrolophus costalis and Macrolophus pygmaeus preying on the tobacco form of Myzus persicae (Hemiptera: Aphididade). Bull Entomol Res 93:39–45. https://doi.org/10.1079/BER2002207
Martínez-Cascales JI, Cenis JL, Cassis G, Sanchez JA (2006) Species identity of Macrolophus melanotoma (Costa 1853) and Macrolophus pygmaeus (Rambur 1839) (Insecta: Heteroptera: Miridae) based on morphological and molecular data and bionomic implications. Insect Syst Evol 37:385–404
Messelink GJ, Bennison J, Alomar O et al (2014) Approaches to conserving natural enemy populations in greenhouse crops: current methods and future prospects. Biocontrol 59:377–393. https://doi.org/10.1007/s10526-014-9579-6
Mollá O, Montón H, Vanaclocha P et al (2009) Predation by the mirids Nesidiocoris tenuis and Macrolophus pygmaeus on the tomato borer Tuta absoluta. IOBC wprs Bull 49:209–214
Mollá O, Biondi A, Alonso-Valiente M, Urbaneja A (2014) A comparative life history study of two mirid bugs preying on Tuta absoluta and Ephestia kuehniella eggs on tomato crops: implications for biological control. Biocontrol 59:175–183. https://doi.org/10.1007/s10526-013-9553-8
Nannini M, Atzori F, Coinu M et al (2014) Developing improved methods for the release of Macrolophus pygmaeus (Rambur) (Heteroptera: Miridae) in sardinian tomato greenhouses. Acta Hortic 1041:163–170. https://doi.org/10.17660/ActaHortic.2014.1041.18
Naselli M, Zappalà L, Gugliuzzo A, Tropea Garzia G, Biondi A, Rapisarda C, Cincotta F, Condurso C, Verzera A, Siscaro G (2016) Olfactory response of the zoophytophagous mirid Nesidiocoris tenuis to tomato and alternative host plants. Arthropod Plant Interact 11:121–131. https://doi.org/10.1007/s11829-016-9481-5
Perdikis D, Lykouressis D (2000) Effects of various items, host plants, and temperatures on the development and survival of Macrolophus pygmaeus Rambur (Hemiptera: Miridae). Biol Control 17:55–60. https://doi.org/10.1006/bcon.1999.0774
Perdikis DC, Lykouressis DP (2002) Life table and biological characteristics of Macrolophus pygmaeus when feeding on Myzus persicae and Trialeurodes vaporariorum. Entomol Exp Appl 102:261–272
Perdikis D, Favas C, Lykouressis D, Fantinou A (2007) Ecological relationships between non-cultivated plants and insect predators in agroecosystems: the case of Dittrichia viscosa (Asteraceae) and Macrolophus melanotoma (Hemiptera: Miridae). Acta Oecol 31:299–306. https://doi.org/10.1016/j.actao.2006.12.005
Perdikis DC, Arvaniti ΚA, Paraskevopoulos A, Grigoriou Α (2015) Pre-plant release enhanced the earlier establishment of Nesidiocoris tenuis in open field tomato. Entomol Hell 24:11–21
Put K, Bollens T, Wäckers FL, Pekas A (2012) Type and spatial distribution of food supplements impact population development and dispersal of the omnivore predator Macrolophus pygmaeus (Rambur) (Hemiptera: Miridae). Biol Control 63:172–180. https://doi.org/10.1016/j.biocontrol.2012.06.011
R-Development-Core-Team (2017) A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna
Riudavets J, Castane C (1998) Identification and evaluation of native predators of Frankliniella occidentalis (Thysanoptera: Thripidae) in the Mediterranean. Environ Entomol 27:86–93
Rossinelli S, Bacher S (2015) Higher establishment success in specialized parasitoids: support for the existence of trade-offs in the evolution of specialization. Funct Ecol 29:277–284. https://doi.org/10.1111/1365-2435.12323
Sanchez JA, Gillespie DR, McGregor RR (2003a) The effects of mullein plants (Verbascum thapsus) on the population dynamics of Dicyphus hesperus (Heteroptera: Miridae) in tomato greenhouses. Biol Control 28:313–319. https://doi.org/10.1016/S1049-9644(03)00116-6
Sanchez JA, Martinez-Cascales JI, Lacasa A (2003b) Abundance and wild host plants of predator mirids (Heteroptera: Miridae) in horticultural crops in the Southeast of Spain. IOBC wprs Bull 26:147–151
Sanchez JA, Gillespie DR, McGregor RR (2004) Plant preference in relation to life history traits in the zoophytophagous predator Dicyphus hesperus. Entomol Exp Appl 112:7–19. https://doi.org/10.1111/j.0013-8703.2004.00174.x
Sanchez JA, del Amor FM, Flores P, López-Gallego E (2016) Nutritional variations at Nesidiocoris tenuis feeding sites and reciprocal interactions between the mirid and tomato plants. J Appl Entomol 140:161–173. https://doi.org/10.1111/jen.12246
Sanchez JA, López-Gallego E, Pérez-Marcos M et al (2018) How safe is it to rely on Macrolophus pygmaeus (Hemiptera: Miridae) as a biocontrol agent in tomato crops? Front Ecol Evol 6:1–10. https://doi.org/10.3389/fevo.2018.00132
Simberloff D (2009) The role of propagule pressure in biological invasions. Annu Rev Ecol Evol Syst 40:81–102. https://doi.org/10.1146/annurev.ecolsys.110308.120304
Smith RL (1980) Ecology and field biology. Harper & Row, Publishers, New York
Stiling P (1990) Calculating the establishment rates of parasitoids in classical biological control. Am Entomol 36:225–230. https://doi.org/10.1093/ae/36.3.225
Sylla S, Brévault T, Diarra K et al (2016) Life-history traits of Macrolophus pygmaeus with different prey foods. PLoS ONE 11:1–8. https://doi.org/10.1371/journal.pone.0166610
Trottin-Caudal Y, Baffert V, Leyre JM, Hulas N (2012) Experimental studies on Tuta absoluta (Meyrick) in protected tomato crops in France: biological control and integrated crop protection. EPPO Bull 42:234–240. https://doi.org/10.1111/epp.2560
Urbaneja A, Montón H, Molla O (2009) Suitability of the tomato borer Tuta absoluta as prey for Macrolophus pygmaeus and Nesidiocoris tenuis. J Appl Entomol 133:292–296. https://doi.org/10.1111/j.1439-0418.2008.01319.x
Urbaneja A, González-Cabrera J, Arnó J, Gabarra R (2012) Prospects for the biological control of Tuta absoluta in tomatoes of the Mediterranean basin. Pest Manag Sci 68:1215–1222. https://doi.org/10.1002/ps.3344
Urbaneja-Bernat P, Alonso M, Tena A, Bolckmans K, Urbaneja A (2013). Sugar as nutritional supplement for the zoophytophagous predator Nesidiocoris tenuis. BioControl 58:57–64. https://doi.org/10.1007/s10526-012-9466-y
Van Dam NM, Hare JD (1998) Differences in distribution and performance of two sap-sucking herbivores on glandular and non-glandular Datura wrightii. Ecol Entomol 23:22–32. https://doi.org/10.1046/j.1365-2311.1998.00110.x
van Lenteren JC (2012) The state of commercial augmentative biological control: plenty of natural enemies, but a frustrating lack of uptake. Biocontrol 57:1–20. https://doi.org/10.1007/s10526-011-9395-1
Venables WN, Ripley BD (2002) Statistic and computing. Modern applied statistic with S. Springer, New York
Wheeler AG, Krimmel BA (2015) Mirid (Hemiptera: Heteroptera) specialists of sticky plants: adaptations, interactions, and ecological implications. Annu Rev Entomol 60:393–414. https://doi.org/10.1146/annurev-ento-010814-020932
Zappala L, Biondi A, Alma A et al (2013) Natural enemies of the South American moth, Tuta absoluta, in Europe, North Africa and Middle East, and their potential use in pest control strategies. J Pest Sci 86:635–647. https://doi.org/10.1007/s10340-013-0531-9
Acknowledgements
We thank the workers at the IMIDA Torreblanca experimental station for technical assistance, María José Ramirez-Soria to help with sampling, and Javier Calvo from Koppert Spain for providing B. tabaci.
Funding
This work has been funded by European Regional Development Fund (FEDER 14-20-03).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Ethical approval
This article does not contain any studies with human participants or animals performed by any of the authors.
Additional information
Communicated by N. Desneux.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Supplementary material Table 1
Parameters of the logistic function (1) and statistics of the non-linear regression for the modeling of the population dynamics of M. pygmaeus in each greenhouse: No, the initial population—estimated number of M. pygmaeus in the greenhouse at t = 0—; r, population growth rate; df, degrees of freedom; R2, coefficient of determination; PRP, pre-plant release period in days; Rep, number of the replicate (DOCX 56 kb)
Rights and permissions
About this article

Cite this article
Sanchez, J.A., López-Gallego, E., Pérez-Marcos, M. et al. The effect of banker plants and pre-plant release on the establishment and pest control of Macrolophus pygmaeus in tomato greenhouses. J Pest Sci 94, 297–307 (2021). https://doi.org/10.1007/s10340-020-01257-z
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10340-020-01257-z