
Abstract
The distribution of water across landscapes affects the diversity and composition of ecological communities, as demonstrated by studies on variation in vascular plant communities along river networks and in relation to groundwater. However, non-vascular plants have been neglected in this regard. Bryophytes are dominant components of boreal flora, performing many ecosystem functions and affecting ecosystem processes, but how their diversity and species composition vary across catchments is poorly known. We asked how terrestrial assemblages of mosses and liverworts respond to variation in (i) catchment size, going from upland-forest to riparian settings along increasingly large streams and (ii) groundwater discharge conditions. We compared the patterns found for liverworts and mosses to vascular plants in the same set of study plots. Species richness of vascular plants and mosses increased with catchment size, whereas liverworts peaked along streams of intermediate size. All three taxonomic groups responded to groundwater discharge in riparian zones by maintaining high species richness further from the stream channel. Groundwater discharge thus provided riparian-like habitat further away from the streams and also in upland-forest sites compared to the non-discharge counterparts. In addition, soil chemistry (C:N ratio, pH) and light availability were important predictors of vascular plant species richness. Mosses and liverworts responded to the availability of specific substrates (stones and topographic hollows), but were also affected by soil C:N. Overall, assemblages of mosses and vascular plants exhibited many similarities in how they responded to hydrological gradients, whereas the patterns of liverworts differed from the other two groups.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
One of the major goals of ecosystem ecology is to assess what factors and mechanisms underlie spatial variation in species composition (Wiens 1989). Significant advances have been made to understand how surface and sub-surface water flows link to landscape patterns in terrestrial biodiversity, with emphasis on riparian zones along longitudinal river gradients (Richardson and others 2005; Dunn and others 2006; Bendix and Stella 2013) and on groundwater (GW) fluxes (Harner and Stanford 2003; Zinko and others 2005; Jansson and others 2007; Kuglerová and others 2014a). Such studies have mainly focused on vascular plants (Planty-Tabacchi and others 1996; Naiman and Décamps 1997; Pollock and others 1998; Sabo and others 2005; Nilsson and others 2012), whereas non-vascular plant taxa, such as bryophytes, are poorly studied. In boreal forests, bryophytes are as abundant and species-rich as vascular plants, being dominant components of boreal ground-layer flora (Dynesius and Zinko 2006; Bagella 2014). Bryophytes are essential for many ecosystem processes (Cornelissen and others 2007), including nitrogen fixation in association with cyanobacteria (DeLuca and others 2002), control of below- and above-ground moisture and temperature, and they modify (Cornelissen and others 2007) or provide (Jonsson and others 2015) habitat for other taxa. Thus, understanding how bryophyte communities vary across landscapes and how they respond to hydrological gradients is important, yet such knowledge is missing.
Hydrological and environmental gradients along branching river systems (Grant and others 2007) affect riparian vascular plant assemblages, with an increase in species richness downstream (Honnay and others 2001; Dunn and others 2011; Kuglerová and others 2015). In many boreal and temperate systems, catchment size and fluvial disturbance progressively increase, soil chemistry systematically changes (for example, soil pH and N content increase), and the range of riparian habitat conditions widen, as the tributaries combine to create larger rivers (Kuglerová and others 2015). These factors, together with enhanced connectivity of waterways (Erős and others 2011) and increases in the efficiency of water dispersal downstream in the network (Honnay and others 2001; Bång and others 2007), help explain the successive increase in vascular plant species richness with increasing catchment size. Other types of water flow across the boreal landscape, unrelated to in-stream hydrology, also affect vascular plant distribution. Sites where GW flowing from surrounding hillslope discharges have elevated numbers of species in riparian as well as upland-forest settings (Kuglerová and others 2014a). Factors associated with GW discharge proposed to promote species richness are higher soil pH, more available nitrogen to plants, and higher soil moisture at GW discharge sites in comparison to their non-discharge counterparts (Zinko and others 2005; Jansson and others 2007; Kuglerová and others 2014a).
The relationships of non-vascular plant taxa such as mosses and liverworts to river network position and GW discharge remain poorly known. Research on moss and liverwort ecology in boreal riparian forests has focused on bryophyte-substrate affiliations (Hylander and others 2005; Hylander and Dynesius 2006), logging disturbances, and/or edge effects (Stewart and Mallik 2006; Dynesius and others 2009; Hylander and Weibull 2012; Selonen and Kotiaho 2013) and covers only short gradients in stream size and fluvial disturbance. Along boreal headwaters, bryophytes have been shown to respond strongly to variation in substrate availability and soil pH (Hylander and Dynesius 2006), but how these factors interact with fluvial regimes and disturbance gradients to drive moss and liverwort diversity across stream networks is not known. The amount and quality of substrates do not, necessarily, relate to network position and it is thus likely that the same set of riparian habitat conditions represent different niches for mosses, liverworts, and vascular plants (Lee and Laroi 1979; Bagella 2014). Further, bryophyte species composition may be favored by higher soil moisture and humidity in GW discharge areas to a larger extent than vascular plants, because mosses and liverworts do not depend only on below-ground sources to acquire water and nutrients (Glime 2007). Taken together, species composition of mosses and liverworts may not respond to surface and sub-surface water flows across the stream network in a similar manner as vascular plants. However, ecological similarities and differences among riparian mosses, liverworts, and vascular plants have rarely been addressed (Bagella 2014) and whether the three taxonomic groups coincide in diversity patterns along hydrological gradients is not known.
We asked how liverwort and moss assemblages are affected by hydrological gradients in terms of (i) catchment size increment from upland (non-riparian, 0 order) to riparian sites situated along increasingly large streams (1st to 7th stream order) and (ii) GW recharge/discharge conditions. Further, we addressed whether these two groups respond to the availability of water in a similar manner as vascular plants in the same set of study plots, and how and why their diversity patterns coincide across the landscape. Because many bryophyte substrates (for example, rocks and wood) are not utilized by vascular plants and because bryophytes and vascular plants acquire water and other resources in different ways, we hypothesized that species richness and community composition of bryophytes will be less linked to the catchment size gradient in comparison to vascular plants and be more linked to specific substrates. We also hypothesized that species assemblages of mosses and liverworts will respond strongly to GW discharge in both riparian and upland-forest settings because it can modify substrate composition and chemistry (for example, increase soil pH), and increase soil moisture (Kuglerová and others 2014a), factors to which bryophytes respond positively (for example, Hylander and Dynesius 2006). Thus, more diverse bryophyte communities can be expected at GW discharge compared to non-discharge sites.
Methods
Study Area and Site Selection
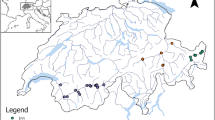
The study area is situated in boreal Sweden, approximately 60 km north-west of the city of Umeå (Figure 1A). In total 44 forested upland and riparian sites were established to represent a gradient in catchment size. The catchment size (that is, the land area drained into an upland-forest site or to the stream adjacent to a riparian site) determines the hydrological conditions of a site, and closely corresponds to the magnitude of fluvial disturbance (for example, stream power, sedimentation rates, plant injuries, Figure A1 in Appendix A in Electronic Supplementary Material). In addition, water level regimes (inundation magnitude, Appendix A in Electronic Supplementary Material), and soil moisture also vary along the catchment size gradient and thus catchment size is a good proxy for the fluvial disturbance gradient (Kuglerová and others 2015). The majority of the sites (36) were located in the Krycklan catchment (Laudon and others 2013) where 30 sites were placed in riparian zones along streams ranging in size from stream orders 1–4, whereas six sites were placed in upland forest with no permanent surface flow (hereafter called order 0) (Figure 1B). The upland-forest sites were included to extend the lower end of the catchment size gradient and represented sites without GW discharge (2 sites) and a gradient in GW discharge (4 sites). We also extended the upper end of the catchment size gradient by selecting four riparian sites along each of two large rivers in the region, namely the Sävar and Vindel Rivers (5th and 7th stream order, respectively, Figure 1A). The catchment size of the selected sites ranged from 25 m2 (upland non-discharge) to 12,000 km2 (Vindel River). Riparian sites (38) were placed in pairs along the same stream reach, representing contrasting GW conditions. One site had high and the other none or little GW discharge (Kuglerová and others 2014a). The determination of GW conditions was based on a flow accumulation algorithm (D8 in ArcHydro Tools, ArcMap version 10, Esri), predicting GW flow paths within riparian zones based on a digital elevation model, and on a topographic wetness index which predicts soil moisture based on contributing area and slope (details in Kuglerová and others 2014a, b). Visual field inspections were made to confirm the model predictions. Each site was further assigned to one of two main geomorphic areas, reflecting the glaciation history. The northern part of the Krycklan catchment is above the former highest postglacial coastline (FHC, Wastenson and Fredén 2002), formed by a pre-stage of the Gulf of Bothnia, and is characterized by glacial till soils, whereas the soils in the southern part, below the FHC, are mainly sorted sedimentary deposits (Figure 1B). The differences in soil types (grain size) may be important for vegetation. Upland-forest and small stream sites (up to 3rd order) were located in both geomorphic areas, but larger stream/river sites (≥4th order) were located below the FHC only. The bedrock in the study area is generally acidic.

The study area in northern Sweden (A) with black markers representing approximate location of study sites placed along the Krycklan (36 sites), Vindel (4), and Sävar (4) Rivers. Thirty of the Krycklan study sites (B) were placed in pairs (one site with groundwater discharge and one without) in riparian zones along the stream network (red markers). Four of the Krycklan sites were placed in upland forest with GW discharge (black circles) and two sites were placed in upland forest with no discharge (black stars). The former highest coastline (FHC), marking the boundary between till and sedimentary deposits, is outlined by a black line (Color figure online).
The Krycklan catchment is dominated by secondary forests with a dominance of Scots pine (Pinus sylvestris) and Norway spruce (Picea abies) with birch (Betula pubescens), alder (Alnus incana), aspen (Populus tremula), and willows (Salix spp.) found in mesic-wet and riparian habitats. Land use is dominated by forestry with clear cuts representing about 7% of the catchment. Arable land and built areas represent only about 2% of the catchment. The mean annual temperature is 1.8°C and average precipitation is 614 mm of which about 40% falls as snow. The hydrology is snow melt driven with peak flows occurring during spring flood, usually in May (Laudon and others 2013). Krycklan is a tributary to the Vindel River, whereas the Sävar River is unconnected hydrologically but closely situated (~20 km, Figure 1A), controlling for similarity in species pool, geology, climate, and hydrological regimes. All selected sites were placed in mature forest stands, with no nearby recent clear cuts.
Species Survey
At each site we established one 5 × 10 m plot. At the 38 riparian sites, the 10-m side of the plot was placed perpendicular to the stream and the 5-m side bordered the stream channel at summer low water level. The streams (that is, the permanently wetted area during summer low flows) were not included in the plots. The 10-m side of the four upland-forest plots with GW discharge were placed parallel to the GW flow paths and the two non-discharge upland-forest plots were randomly placed. Along all streams, except for the Vindel River (the largest river), the 10-m-long plot laterally encompassed the entire riparian zone and its upper part extended into areas that are not regularly flooded (that is, beyond the riparian zone). To apply the same plot size at the Vindel River sites and encompass the entire riparian zone, we excluded the lowest 5 m of the riparian zone so that the plots were established at the distance 5–15 m from the water edge. Vascular plant species richness was low within the excluded 5 m closest to the channel and the species were also found at higher elevations, and this zone was almost devoid of bryophytes.
Additionally, we divided each 5 × 10 m plot into two sub-plots as the lower and an upper halves (5 × 5 m). We recorded all species of liverworts, mosses, and vascular plants in the entire 5 × 10 m plots, whereas mosses and liverworts were also recorded in the two 5 × 5 m sub-plots separately. Vascular plants and bryophytes were inventoried in summers of 2011 and 2013, respectively. The taxonomy and nomenclature followed Krok and Almquist (1994) for vascular plants and Hallingbäck and others (2006) for liverworts and mosses. Some combinations of species were treated as one taxon (Appendix B in Electronic Supplementary Material). We classified each moss and liverwort species according to the substrate (wood and bark, ground, rocks and boulders) where it most commonly occurs in the region. The vascular plant species found are all ground living (Appendix B in Electronic Supplementary Material).
Plot Measurements
In each 5 × 10 m plot, we measured several variables describing habitat and substrates, thought to be important for bryophytes, henceforth called plot measurements. We visually estimated the area of (i) dead wood (m2), (ii) open soil not covered by vegetation (m2), (iii) stones not covered by ground-living bryophytes (m2), and (iv) topographic hollows (m2), that is, small scale depressions between rocks, roots, stumps, and so on. We also recorded diameter at breast height (DBH) of all trees within the 5 × 10 m plot and calculated the total basal area (m2 ha−1), which corresponds to the amount of available bark surfaces (tree bases) as well as to light penetrating to the understory. We also counted the number of tree saplings (that is, trees <2 m high and <2 cm in DBH) in each plot, with high numbers indicating high below canopy availability of light. Canopy openness was also directly measured by image analysis of canopy photos following the methodology in Canham (1988). We calculated potential annual direct incident radiation (PADIR, MJ cm−2 y−1) from the digital elevation model using Spatial Analyst Tools in ArcGIS (version 10.2) to account for variation in solar energy inputs related to slope and aspect. As indicators of soil chemistry, we measured soil pH, and total C and N (expressed as C:N ratio) in each plot (see Kuglerová and others [2014a] for methodology). Soil chemistry and canopy measurements were conducted in 2011, and substrates and tree measurements in 2014.
Statistical Analysis
All statistical analyses were performed in R (R Development Core Team 2013). Unless specified otherwise, data represent results from the entire 5 × 10 m plots. First, we tested whether species richness correlated among the three groups (vascular plants, mosses, and liverworts) using Pearson product-moment correlations because species richness data were normally distributed (Shapiro–Wilk tests, P > 0.05). Second, we related species assemblages to hydrological variables and all plot measurements.
Hydrological Gradients
We investigated if species richness of mosses and liverworts respond to hydrological factors in similar ways as vascular plants across the same set of study sites (Kuglerová and others 2014a, 2015). We used linear mixed effect models (LMMs, Baayen and others 2008) with species richness as response variable and catchment size, groundwater conditions (GW discharge vs. non-discharge) and geomorphic area (below vs. above FHC) and their interactions as fixed factors, and stream identity (because pairs of sites along the same stream may be spatially dependent) as a random factor. In these analyses, the catchment size gradient is used as a surrogate for increasing fluvial disturbance, because it correlates tightly with stream power, inundation magnitude, sedimentation rate, and an index of plant injury (Kuglerová and others 2015, Appendix A in Electronic Supplementary Material). In the LMMs, species richness was assumed to have Poisson error structures. Selection of the best model to predict species richness of each plant group was based on likelihood ratio tests and on the Akaike information criterion (AIC). The LMMs were performed in the package lme4 (Bates and Maechler 2009). Additionally, we tested how GW discharge (that is, its presence or absence based on the hydrological model) affects the widths of riparian vegetation by looking for differences in bryophyte species richness between GW discharge and non-discharge sites separately for the lower and upper 5 × 5 m riparian sub-plots (LMMs). We removed the six upland-forest sites from the last analysis.
To investigate how community composition responds to catchment size and GW discharge, we performed permutational multivariate analysis of variance (PERMANOVA) for all three plant groups in the vegan package (Oksanen and others 2013). Vascular plant data were also analyzed, because in contrast to species richness, species composition analyses had not been done before. PERMANOVA was developed for categorical factors (Anderson 2001); hence, we used stream order to describe the catchment size gradient (these two variables were closely correlated; r = 0.9). For mosses and liverworts, we also performed PERMANOVAs for the lower and upper sub-plots separately. The six upland-forest sites were included in all composition analyses. In these sites, the two sub-plots were assigned randomly because they had no relation to stream distance.
Plot Measurements
Before analyzing the influence of plot measurement variables (basal area of trees, canopy openness, amount of dead wood, number of tree saplings, area of open soil, PADIR, soil C:N and pH, cover of stones, and area of topographic hollows) on species richness, we addressed the problem of multicollinearity (Graham 2003), although only a few plot measurements were strongly correlated (r > 0.5, Table A1 in Appendix A in Electronic Supplementary Material). For this purpose, we reduced the number of predictor variables by using principal component regressions. First, we performed principal component analysis (PCA) with all plot measurement variables (standardized data). The scores of the first four principal components (PC) were then used as explanatory factors in multiple regressions, with species richness of vascular plants, mosses, and liverworts, respectively, as response variables. To evaluate the individual independent contribution of each plot measurement variable to the explained variance in species richness, we performed hierarchical partitioning (package hier.part, Walsh and MacNally 2013), a method robust to multicollinearity between explanatory factors (Chevan and Sutherland 1991). Variables were transformed to fulfill the assumption of normality when needed.
To relate species composition of the three groups to variation in plot measurement variables, we used non-metric multidimensional scaling (NMDS) (Kruskal 1964). The presence/absence data of species and Bray–Curtis distances were used to project the assemblage data onto ordination space. All plot measurements, plus catchment size, were fitted onto community ordination diagrams, and the function envfit in package vegan (Oksanen and others 2013) provided squared correlation coefficients and P values for each of the variables with assemblage data. Joint plots were used to depict relationships of assemblages to fitted plot variables as well as to visualize how stream orders and GW discharge conditions segregate species assemblages.
Results
Species Richness
In total, we found 137 species of vascular plants, 113 species of mosses, and 51 species of liverworts in all 44 study plots combined. Seventy percent of all moss species (79 species) and 57% of all liverwort species (29) were ground living, 19% (21 mosses) and 20% (10 liverworts) were stone-associated species, and 11% (13 mosses) and 23% (12 liverworts) were species associated with wood or bark (Appendix B in Electronic Supplementary Material). Only one nationally red-listed species was found in the study, the liverwort Anastrophyllum hellerianum (near threatened).
Species richness of the three groups was significantly positively correlated (Person’s correlation coefficients, P < 0.01) with the strongest correlation between vascular plants and mosses (r = 0.67) and the weakest between vascular plants and liverworts (r = 0.40, Appendix C in Electronic Supplementary Material). The two upland-forest sites with no GW discharge had high leverage on correlations (Appendix C in Electronic Supplementary Material). When removing these two sites (n = 42), the correlation between vascular plants and liverworts became insignificant (r = 0.26, P = 0.09), whereas mosses still correlated significantly (P < 0.01) with vascular plants (r = 0.60) and liverworts (r = 0.45). Restricting the correlation analysis to ground-living species across all 44 sites, thus excluding 34 moss and 22 liverwort species, improved the correlations between species richness of vascular plants and mosses (r = 0.72, P < 0.01) and vascular plants and liverworts (r = 0.48, P = 0.01). The correlations between mosses and liverworts were similar when ground-living (r = 0.56, P < 0.01) or non-ground-living (stone-, wood- and bark-living) species (r = 0.54, P < 0.01) were analyzed.
Hydrological Gradients
Across all 44 study sites, significant trends were found between catchment size and species richness of mosses (z = 5.63, P < 0.001, df = 20) and liverworts (z = 3.25, P = 0.001, df = 16) in the linear mixed effect models (LMMs). Similarly, as for vascular plants (Kuglerová and others 2015, Appendix D in Electronic Supplementary Material), species richness of mosses increased with catchment size (Figure 2A), whereas liverworts exhibited a quadratic pattern (Figure 2B). In contrast to vascular plants (Kuglerová and others 2014a, Appendix D in Electronic Supplementary Material), we did not detect any overall significant effect of GW discharge on species richness either for mosses (z = 1.24, P = 0.22, df = 20, Figure 2C) or liverworts (z = 0.45, P = 0.65, df = 16, Figure 2D). However, in the best models the interaction between GW discharge and catchment size was retained and close to significant for mosses (z = −1.76, P = 0.08, df = 20) and significant for liverworts (z = −2.55, P = 0.01, df = 16). This indicates that effects of GW discharge varied with catchment size with essentially no consistent effects of GW in riparian but a strong effect in the upland-forest sites (0 order in Figure 2C, D). The three-way interaction of GW discharge, catchment size, and geomorphic area (below vs. above FHC) was also retained in the best model for liverworts (z = 2.22, P = 0.03, df = 16). For mosses, geomorphic area was not retained in the final model. The results remained similar (Appendix D in Electronic Supplementary Material) after removing the two non-discharge upland-forest sites (n = 42), that is, positive relationships for moss and liverwort (quadratic) species richness with catchment size and no overall effect of GW discharge. When restricting the analyses for non-ground-living species of mosses and liverworts across all sites (n = 44), the results for moss species richness resembled those for total species richness (including ground living), that is, catchment size and its interaction with GW were significant in the final model but not GW on its own (Appendix D in Electronic Supplementary Material). For non-ground-living liverworts, geomorphic area was the only individual variable that was significant in the final model, but also in its two-way interaction with catchment size and the three-way interaction including GW discharge (Appendix D in Electronic Supplementary Material). In contrast, for ground-living mosses and liverworts catchment size was the only retained variable explaining species richness in the best models (n = 44, Appendix D in Electronic Supplementary Material).

Species richness of mosses (A) and liverworts (B) in relation to catchment size across all study sites (n = 44). Mean species richness (±SE) of mosses (C) and liverworts (D) is also displayed for GW discharge (gray bars) and non-discharge (white bars) sites separately for each stream order. Note the differences in the scale of the y-axes between mosses and liverworts.
Species richness of mosses and liverworts was generally higher in the lower than in the upper sub-plots across all riparian sites (Figure 3, n = 38). This difference was smaller in sites with GW discharge, where the upper plot had 25% fewer moss species in comparison to 41% fewer at non-discharge sites and 16% fewer liverworts at GW discharge sites in comparison to 60% fewer liverworts at non-discharge sites (Figure 3). In the lower riparian sub-plots, adjacent to the stream, GW discharge did not have any significant effect on species richness of mosses (z = −0.86, P = 0.39, df = 20, Figure 3A) and liverworts (z = −1.48, P = 0.14, df = 20, Figure 3C), whereas in the upper riparian sub-plots, further from the stream, we found significantly more species of both (mosses: z = 2.53, P = 0.01, df = 20; liverworts: z = 3.06, P = 0.002, df = 20) at the GW discharge sites (Figure 3B, D). This trend was similar across the entire catchment size gradient, except the smallest streams (Figure 3).

Mean species richness (±SE) per stream order of mosses (A, B) and liverworts (C, D) in relation to GW conditions (discharge vs. non-discharge) displayed separately for the lower and upper 5 × 5 m sub-plots. Only riparian sites were used (n = 38). Significantly higher species richness was found at the GW discharge sites at the upper sub-plots for both mosses and liverworts (LLMs). Note the differences between mosses and liverworts in the scale of the y-axes.
Plot Measurements
Some of the plot variables were correlated (Spearman rank correlation) with each other (r < 0.6, P < 0.05, Table A1 in Appendix A in Electronic Supplementary Material) as well as with catchment size and GW discharge (LMMs). In general, tree basal area and soil C:N ratio were lower along larger streams, and lower basal area, lower soil C:N, less acidic soils, less stones, and more topographic hollows were found at GW discharge sites (Table A2 in Appendix A in Electronic Supplementary Material).
In the PCA of the ten plot measurements, the first four components explained 67% of the variation in habitat conditions. The first axis, explaining about 25% of the total variance, was strongly (r > 0.7) positively correlated with soil pH and strongly negatively correlated with soil C:N ratio. The first axis was also positively correlated (r > 0.4) with the amount of dead wood, open soil, and number of saplings, and negatively correlated with tree basal area (Table 1). The second axis, explaining 15% of the total variance, was mostly correlated with canopy openness (negatively), cover of stones, and topographic hollows (both positive). The third and fourth components were correlated with the amount of open soil (positive) and PADIR (negative), individually explaining 14 and 13% of the variance, respectively (Table 1). Multiple regressions with the scores of the four principal components (PC) as independent and species richness as response variables revealed differences in how the species richness of three groups responded to the plot measurements. PC1 was significant for vascular plants and mosses (β standard = 0.49, P < 0.01 and β standard = 0.31, P = 0.03, respectively), whereas PC2 was significant only for liverworts (β standard = 0.36, P = 0.01). PC4 was significant for all three groups (vascular plants: β standard = −0.35, P < 0.01; mosses: β standard = −0.36, P = 0.01, liverworts: β standard = −0.39, P < 0.01) and PC3 was not significant for any group.
In the hierarchical partitioning models, five, two and three of the ten plot measurement variables independently explained significant variance in species richness of vascular plants, mosses and liverworts, respectively (Table 1). Twenty-six, 26, 17, 13, and 11% of the variance of vascular plant species richness was explained by basal area, soil C:N, number of tree saplings, soil pH, and canopy openness, respectively. Variation in moss richness was significantly explained by soil C:N (34%) and amount of stones (26%). Soil C:N (18%) and stones (23%) also explained the variation in liverwort species richness, but the strongest predictor for this group was the area of topographic hollows (28%) (Table 1).
Assemblage Composition
The PEMANOVAs performed at the entire 5 × 10 m plot level showed that stream order was a strong predictor of species composition, significantly segregating assemblages of all three groups (Table 2; Figure 4A–C). GW discharge significantly affected species composition of vascular plants and mosses, but not liverworts (Table 2; Figure 4D–F). When we performed the PERMANOVAs for the upper and lower sub-plots separately, the results for the lower sub-plots mirrored the results from the entire 5 × 10 m plots for both mosses and liverworts (vascular plants were not analyzed; Appendix E in Electronic Supplementary Material). In contrast, the moss and liverwort species assemblages in the upper sub-plots were less segregated by stream order (lower r 2, although still significant) and more segregated by GW discharge (higher r 2 and significant) than the assemblages in the lower sub-plot adjacent to the stream channel (Appendix E in Electronic Supplementary Material).

Results of NMDS for assemblages of vascular plants, mosses, and liverworts across all study sites (n = 44). The top and middle rows of panels display how assemblages of vascular plants (A, D), mosses (B, E), and liverworts (C, F) segregate by stream order and GW discharge conditions, respectively. The bottom three figures display plot measurement variables (vectors displayed in blue) which were identified as significantly correlated with community patterns (DW dead wood; BA basal area; CN soil C:N ratio; Canopy canopy openness; Catchment catchment size; pH soil pH; Open.soil area of open soil). Note that the positions of the points in all three panels for a given plant group are identical because they are from the same ordination (Color figure online).
Non-metric multidimensional scaling of assemblage composition in the entire 5 × 10 m plots suggested two-dimensional ordination solutions for all three plant groups. Assemblage patterns of all three groups in NMDS ordination space were most correlated with catchment size and soil C:N ratio (Table 2; Figure 4G–I), which were also negatively correlated with each other (Appendix B in Electronic Supplementary Material). For vascular plants and mosses, several other plot measurements (total basal area, dead wood, open soil, canopy openness, soil pH) showed significant relationships with assemblage data (Table 2; Figure 4G, H), while for liverworts, the only other significant variable was the area of topographic hollows (Table 2; Figure 4I).
Discussion
Catchment Size
Species richness of vascular plants has previously been shown to monotonically increase with catchment size (Kuglerová and others 2015) and here we documented a similar, although not as strong, trend for mosses in the same set of river network plots (Figure 2), and consequently, vascular plant and moss species richness correlated well (Appendix C in Electronic Supplementary Material). This shows that across boreal ecosystems, sites favorable for vascular plants are also likely favorable for mosses and the two groups can thus function as surrogates when predicting biodiversity (Bagella 2014). The correlations of species richness between vascular plants and bryophytes (also liverworts, Appendix C in Electronic Supplementary Material) can mostly be explained by their similar response to variation in environmental drivers. At the same time, it has been shown that bryophytes can facilitate habitat for vascular plants (although negative interactions were also reported in, for example, Nilsson and Wardle 2005), primarily by controlling soil moisture, temperature, and nutrient availability (Cornelissen and others 2007), factors that are important for plant establishment and growth, especially in cold regions. It is possible that in our case, high bryophyte abundance and diversity at some of the most species-rich sites may be associated with high facilitation function and thus contributes to the diversity of vascular plants.
The effect of increasing catchment size on moss species richness could be interpreted as a positive response to progressively increasing fluvial disturbance, inundation range, and heterogeneity of riparian habitat conditions downstream in the network (Appendix A in Electronic Supplementary Material). This is also supported by the fact that the additional habitat variables in this study (plot measurements) which affected bryophyte assemblages had none or weak correlations with catchment size. On the other hand, affiliation of mosses to specific substrates was likely the reason for mosses having a weaker relationship with catchment size than vascular plants. Liverwort species richness exhibited a quadratic relationship with catchment size (Figure 2). Bryophyte species richness peaking along medium-sized streams was shown in another stream network study, which, however, did not distinguish between mosses and liverworts (Jonsson 1996). Jonsson (1996) suggested that mid-sized streams had intermediate levels of flooding and erosion, allowing species of both disturbed and stable habitats to coexist. Intermediate disturbances along mid-river reaches have also been linked to peaks in species richness for riparian vascular plants (Nilsson and others 1989; Tabacchi and others 1996; Dunn and others 2011). These trends were observed mostly along longitudinal gradients of single channels not covering the range of catchment sizes and connectivity (Erős and others 2011) of the river network in our study. The likely explanation for the quadratic pattern of liverwort species richness along the catchment size gradient in our study is the peak in stones and topographic hollows, substrates important for liverworts, along some of the mid-sized streams. Together with this, competition from the other two plant groups (Bruun and others 2006) along the largest streams might have reduced liverwort species richness despite availability of suitable substrates. The differences between mosses and liverworts can be explained by a smaller proportion of liverwort than moss species being ground living and thus fluvial effects might have less influence on liverwort species composition (Appendix D in Electronic Supplementary Material). Finally, the stronger link between catchment size and vascular plants than bryophytes can also be explained by their dispersal strategy. While many riparian vascular plant species rely on hydrochory which is more efficient downstream in the network (Kuglerová and others 2015), bryophytes disperse by small, wind-dispersed spores and unlikely benefit from dispersal by water.
We also documented directional changes in assemblage composition with respect to catchment size, supporting previous findings of changes in bryophyte species assemblages along stream size gradients (Jonsson 1997; Gould and Walker 1999). Assemblages of vascular plants and mosses changed substantially with catchment size only at the upper end of the size gradient, with small changes between stream order one and four (vascular plants) and one and three (mosses). In contrast, liverwort species assemblages were more continuously segregated by stream size category (Figure 4).
Groundwater Discharge
GW discharge did not affect species richness of mosses and liverworts in the same way as vascular plants, for which higher species richness was found at GW discharge sites across the entire catchment (Kuglerová and others 2014a). Although the GW discharge sites provided more bryophyte-favorable habitat (higher variability of substrates, soil pH, and water availability), the high above- and below-ground moisture favored specific species, such as Sphagnum spp., which had high abundance and may have restricted the frequency of other bryophyte species. Despite the lack of differences across the entire catchment size gradient, GW discharge increased species numbers of mosses and liverworts in the upland-forest sites and in the upper riparian sub-plots in comparison to the non-discharge sites (Figures 2, 3). GW discharge thus compensated for the lower fluvial influence further away from the stream channels, which is, at the non-discharge sites, associated with a sharp decrease in bryophyte richness (Dynesius and others 2009; Selonen and Kotiaho 2013). This is likely caused by high soil moisture, higher soil pH, lower C:N ratio, and more topographic variation (Appendix B in Electronic Supplementary Material) being provided further from the streams at GW discharge sites. This finding confirms that discharge of GW laterally extend riparian-like habitat upwards into hillslopes and maintain higher diversity of plants further from the streams in comparison to non-discharge riparian areas nearby (Kuglerová and others 2014a). At the same time, the results suggest that GW discharge increases bryophyte richness only when the direct effect of fluvial disturbance is small or absent.
The largest differences in bryophyte assemblage composition between GW discharge and non-discharge sites were observed at the upland-forest sites and in the riparian sites along the largest rivers (5th and 7th order), whereas there was little effect along small to medium-sized streams (Figure 4). Hylander and Dynesius (2006) also did not find a strong effect of area of wet/moist ground, a typical feature of GW discharge, on bryophyte richness and species composition along small and medium boreal streams. In accordance with the species richness results, moss and liverwort assemblages in the upper sub-plots were more strongly affected by GW discharge than catchment size in comparison to the lower sub-plots (Appendix E in Electronic Supplementary Material) highlighting the importance of GW discharge when fluvial influence is small.
Plot Measurements
Of the ten plot variables, only one—soil C:N ratio—was significantly related to species richness and composition of all three plant groups (Table 1; Figure 4). Lower soil C:N translates to higher relative availability of nitrogen, which is generally the most limiting plant nutrient in the region (Tamm 1991). The result that low C:N promotes riparian vascular plant species richness has been previously documented (Kuglerová and others 2014a, 2015) but it has not been linked to bryophyte assemblages. Bryophytes lack root systems and are thus poor competitors for soil nitrogen (DeLuca and others 2002). In that sense, the close link between soil C:N and bryophytes is surprising. On the other hand, soil chemistry may control the occurrence of bryophyte species growing at the soil surface even if they do not acquire N by roots. At the same time, the higher relative N content in riparian soils may be a feedback from the bryophyte community, which is associated with N fixing bacteria (Cornelissen and others 2007), providing nitrogen into the system during bryophyte decay. The fact that higher relative N content was found in places with either high bryophyte diversity (large streams) or high bryophyte cover (Sphagnum mats at GW discharge areas, Appendix A in Electronic Supplementary Material) supports this hypothesis. This could be an ecologically important aspect of bryophyte ecosystem function; however, to confirm this speculation, experimental manipulation needs to be conducted. Nevertheless, in both the hierarchical partitioning models and the PCA regressions for species richness, and the NMDS of assemblage composition, soil C:N was less important for liverworts than for mosses and vascular plants, presumably because a smaller proportion of the liverwort species pool is ground living compared to mosses and vascular plants (Appendix B in Electronic Supplementary Material). Similarly, soil pH was positively associated with species richness in the PCA regressions and assemblage composition (NMDS) of mosses and vascular plants but not liverworts. Hierarchical partitioning positively associated soil pH only with vascular plants. Soil pH has been shown to be a strong predictor of bryophyte richness and composition along small streams in northern Sweden (Hylander and Dynesius 2006). Across a gradient of six stream orders in an Arctic river system, soil pH did not correlate with richness of bryophytes, but did have an effect on species composition (Gould and Walker 1999). Here we conclude that although variation in soil pH may be important for riparian bryophytes, stronger gradients related to fluvial disturbance and substrate availability likely override this effect on moss and especially liverwort assemblages in the study network.
Moss and liverwort species richness was positively influenced by the increasing cover of stones in the plots, a factor that did not influence vascular plants. This was expected because stones and rocks represent important bryophyte substrates (Jonsson 1997; Hylander and others 2005; Hylander and Dynesius 2006) utilized by very few vascular plants. Another plot measurement variable important for both bryophyte groups but not for vascular plants was the area of topographic hollows. For both bryophyte taxa, the area of topographic hollows affected composition, but only for liverworts a positive correlation with species richness was detected (Table 1; Figure 4). The area of depressions has been found to be important for bryophytes previously (Hylander and others 2005; Hylander and Dynesius 2006), the hypothesized mechanism being that topographic hollows provide more moisture, more shelter, and larger variation in microhabitat conditions. This is important for bryophytes and especially for small liverworts also because depressions may represent microhabitats with low competition for space due to low light levels (Dynesius and others 2009), an environment suitable for the many bryophyte species that are shade adapted (Glime 2007). This conclusion is confirmed by the PCA results showing that only liverwort species richness was positively associated with decreasing light availability.
It was unexpected that we did not find a stronger relationship between wood/bark substrates (apart from its contribution in the PCA regression) and richness of mosses and liverworts, because decaying wood and tree bases accommodate specific bryophyte species (Appendix B in Electronic Supplementary Material). These substrates, however, occurred across the study sites in similar volumes, and thus, many common species associated with them were present everywhere, not contributing much to species richness variability. Also, very few species demanding high-quality wood and bark were present in the study plots (M. Dynesius, personal observation). Similarly, the lack of relationships between moss community composition and variables related to light availability (basal area, canopy openness, PADIR) was unexpected. It is possible that the measurements we took for estimating light availability were too coarse to show correlations with mosses. Liverwort species richness was positively associated with decreasing canopy openness in the PCA, indicating that larger proportions of liverwort than moss species in our study plots are physiologically adapted to low light intensities (Glime 2007). Vascular plant species richness and composition were affected by variation in light penetrating to the understory layer more than bryophytes: vascular plants were positively correlated with canopy openness and amount of tree saplings (which positively respond to increasing light levels) and negatively correlated with total basal area of trees (Table 1; Figure 4). Higher light availability influences species richness of riparian vascular plants, because it can promote the abundance and number of individuals, and favor species-rich groups such as graminoids and herbs (Pabst and Spies 1998; Lite and others 2005).
Conclusions
Here we tested how plant taxa related to hydrological gradients across the boreal landscape. Importantly, we found that community composition of mosses and liverworts are driven by fluvial regimes and groundwater conditions, aspects of ecosystem science that have previously been inadequately addressed. Against our expectations, we documented an overall similarity in how boreal assemblages of vascular plants and mosses responded to increasing catchment size, although some of the similarity was offset by the stronger response of mosses to substrate variation. In contrast, liverwort assemblages were affected by catchment size in a different way and their richness patterns along the river network diverged from those of vascular plants and mosses, mainly because many liverwort species were affiliated to stone substrates and topographic hollows. Further, we confirmed the significance of GW discharge in boreal forests because along streams it laterally widened the areas with high species numbers of all three groups and maintained more riparian-like assemblages further away from the stream channels and in upland-forest plots. It has been suggested that riparian buffers, spared from disturbance by timber harvest, should be wider in areas with groundwater discharge to mitigate negative effects of forestry on aquatic ecosystems (for example, increased sedimentation, pollutants and metal export, soil rutting, Kuglerová and others [2014b]). Here we showed that plant diversity follows GW flow paths, and thus, application of hydrologically determined riparian buffers would benefit multiple ecosystem services, including biodiversity. With knowledge about similarities and differences in species composition and diversity across catchments, we may address questions about their relative importance for ecosystem processes. For example, what is the role of mosses and liverworts for variation in soil nutrient dynamics and stream water chemistry across catchments and how their abundance and diversity influence other organisms?
References
Anderson MJ. 2001. A new method for non-parametric multivariate analysis of variance. Austral Ecol 26:32–46.
Baayen RH, Davidson DJ, Bates DM. 2008. Mixed-effects modeling with crossed random effects for subjects and items. J Mem Lang 59:390–412.
Bagella S. 2014. Does cross-taxon analysis show similarity in diversity patterns between vascular plants and bryophytes? Some answers from a literature review. CR Biol 337:276–82.
Bates DM, Maechler M. 2009. lmer4: linear mixed-effects models using S4 classes. R package version 0.999375-32.
Bendix J, Stella JC. 2013. Riparian vegetation and the fluvial environment: a biogeographic perspective. In: Shroder J, Ed. Treatise on geomorphology. San Diego: Academic Press. p 53–74.
Bruun HH, Moen J, Virtanen R, Grytnes JA, Oksanen L, Angerbjorn A. 2006. Effects of altitude and topography on species richness of vascular plants, bryophytes and lichens in alpine communities. J Veg Sci 17:37–46.
Bång A, Nilsson C, Holm S. 2007. The potential role of tributaries as seed sources to an impoundment in Northern Sweden: a field experiment with seed mimics. River Res Appl 23:1049–57.
Canham CD. 1988. An index for understory light levels in and around canopy gaps. Ecology 69:1634–8.
Chevan A, Sutherland M. 1991. Hierarchical partitioning. Am Stat 45:90–6.
Cornelissen JHC, Lang SI, Soundzilovskaia NA, During HJ. 2007. Comparative cryptogam ecology: a review of bryophyte and lichen traits that drive biogeochemistry. Ann Bot 99:987–1001.
DeLuca TH, Zackrisson O, Nilsson MC, Sellstedt A. 2002. Quantifying nitrogen-fixation in feather moss carpets of boreal forests. Nature 419:917–20.
Dunn RR, Colwell RK, Nilsson C. 2006. The river domain: why are there more species halfway up the river? Ecography 29:251–9.
Dunn WC, Milne BT, Mantilla R, Gupta VK. 2011. Scaling relations between riparian vegetation and stream order in the Whitewater River network, Kansas, USA. Landsc Ecol 26:983–97.
Dynesius M, Hylander K, Nilsson C. 2009. High resilience of bryophyte assemblages in streamside compared to upland forests. Ecology 90:1042–54.
Dynesius M, Zinko U. 2006. Species richness correlations among primary producers in boreal forests. Divers Distrib 12:703–13.
Erős T, Schmera D, Schick RS. 2011. Network thinking in riverscape conservation—a graph-based approach. Biol Conserv 144:184–92.
Glime, JM. 2007. Bryophyte ecology. Volume 1. Physiological ecology. Ebook sponsored by Michigan Technological University and the International Association of Bryologists. http://www.bryoecol.mtu.edu/. Accessed 14 Jan 2015.
Gould WA, Walker MD. 1999. Plant communities and landscape diversity along a Canadian arctic river. J Veg Sci 10:537–48.
Graham MH. 2003. Confronting multicollinearity in ecological multiple regression. Ecology 84:2809–15.
Grant EHC, Lowe WH, Fagan WF. 2007. Living in the branches: population dynamics and ecological processes in dendritic networks. Ecol Lett 10:165–75.
Hallingbäck T, Hedenas L, Weibull H. 2006. New checklist of Swedish bryophytes. Svensk Botanisk Tidskrift 100:96–148.
Harner MJ, Stanford JA. 2003. Differences in cottonwood growth between a losing and a gaining reach of an alluvial floodplain. Ecology 84:1453–8.
Honnay O, Verhaeghe W, Hermy M. 2001. Plant community assembly along dendritic networks of small forest streams. Ecology 82:1691–702.
Hylander K, Dynesius M. 2006. Causes of the large variation in bryophyte species richness and composition among boreal streamside forests. J Veg Sci 17:333–46.
Hylander K, Dynesius M, Jonsson BG, Nilsson C. 2005. Substrate form determines the fate of bryophytes in riparian buffer strips. Ecol Appl 15:674–88.
Hylander K, Weibull H. 2012. Do time-lagged extinctions and colonizations change the interpretation of buffer strip effectiveness?—a study of riparian bryophytes in the first decade after logging. J Appl Ecol 49:1316–24.
Jansson R, Laudon H, Johansson E, Augspurger C. 2007. The importance of groundwater discharge for plant species number in riparian zones. Ecology 88:131–9.
Jonsson BG. 1996. Riparian bryophytes of the HJ Andrews Experimental Forest in the western Cascades, Oregon. Bryologist 99:226–35.
Jonsson BG. 1997. Riparian bryophyte vegetation in the Cascade mountain range, Northwest U.S.A.: patterns at different spatial scales. Can J Bot 75:744–61.
Jonsson M, Kardol P, Gundale MJ, Bansal S, Nilsson M-C, Metcalfe DB, Wardle DA. 2015. Direct and indirect drivers of moss community structure, function, and associated microfauna across a successional gradient. Ecosystems 18:154–69.
Krok TOBN, Almquist S. 1994. Svensk flora. Fanerogamer och ormbunksväxter. Stockholm: Liber Utbildning.
Kruskal JB. 1964. Nonmetric multidimensional scaling—a numerical method. Psychometrika 29:115–29.
Kuglerová L, Jansson R, Ågren A, Laudon H, Malm-Renöfält B. 2014a. Groundwater discharge creates hotspots of riparian plant species richness in a boreal forest stream network. Ecology 95:715–25.
Kuglerová L, Ågren A, Jansson R, Laudon H. 2014b. Towards optimizing riparian buffer zones: ecological and biogeochemical implications for forest management. For Ecol Manag 334:74–84.
Kuglerová L, Jansson R, Sponseller RA, Laudon H, Malm-Renöfält B. 2015. Local and regional processes determine plant species richness in a river-network metacommunity. Ecology 96:381–91.
Laudon H, Taberman I, Ågren A, Futter M, Ottosson-Löfvenius M, Bishop K. 2013. The Krycklan Catchment Study—A flagship infrastructure for hydrology, biogeochemistry, and climate research in the boreal landscape. Water Resour Res 49:7154–8.
Lee TD, Laroi GH. 1979. Bryophyte and understory vascular plant beta diversity in relation to moisture and elevation gradients. Vegetatio 40:29–38.
Lite SJ, Bagstad KJ, Stromberg JC. 2005. Riparian plant species richness along lateral and longitudinal gradients of water stress and flood disturbance, San Pedro River, Arizona, USA. J Arid Environ 63:785–813.
Naiman RJ, Décamps H. 1997. The ecology of interfaces: Riparian zones. Annu Rev Ecol Syst 28:621–58.
Nilsson C, Grelsson G, Johansson M, Sperens U. 1989. Patterns of plant species richness along riverbanks. Ecology 70:77–84.
Nilsson C, Jansson R, Kuglerová L, Lind L, Ström L. 2012. Boreal riparian vegetation under climate change. Ecosystems 16:401–10.
Nilsson MC, Wardle DH. 2005. Understory vegetation as a forest ecosystem driver: evidence from the northern Swedish boreal forest. Front Ecol Environ 3:421–8.
Oksanen J, Blanchet FG, Kindt R, Legendre P, Minchin PR, O’Hara RB, Simpson GL, Solymos P, Stevens MHH, Wagner H. 2013. Vegan: R Community ecology packge.
Pabst RJ, Spies TA. 1998. Distribution of herbs and shrubs in relation to landform and canopy cover in riparian forests of coastal Oregon. Can J Bot 76:298–315.
Planty-Tabacchi AM, Tabacchi E, Naiman RJ, Deferrari C, Décamps H. 1996. Invasibility of species rich communities in riparian zones. Conserv Biol 10:598–607.
Pollock MM, Naiman RJ, Hanley TA. 1998. Plant species richness in riparian wetlands—a test of biodiversity theory. Ecology 79:94–105.
R Developmental Core Team. 2013. R: a language and environment to statistical computing. Vienna: R Foundation for Statistical Computing.
Richardson JS, Naiman RJ, Swanson FJ, Hibbs DE. 2005. Riparian communities associated with Pacific Northwest headwater streams: assemblages, processes, and uniqueness. J Am Water Resour Assoc 41:935–47.
Sabo JL, Sponseller R, Dixon M, Gade K, Harms T, Heffernan J, Jani A, Katz G, Soykan C, Watts J, Welter A. 2005. Riparian zones increase regional species richness by harboring different, not more, species. Ecology 86:56–62.
Selonen VAO, Kotiaho JS. 2013. Buffer strips can pre-empt extinction debt in boreal streamside habitats. BMC Ecol 13:24.
Stewart KJ, Mallik AU. 2006. Bryophyte responses to microclimatic edge effects across riparian buffers. Ecol Appl 16:1474–86.
Tabacchi E, PlantyTabacchi AM, Salinas MJ, Decamps H. 1996. Landscape structure and diversity in riparian plant communities: a longitudinal comparative study. Regul Rivers Res Manag 12:367–90.
Tamm CO. 1991. Nitrogen in terrestrial ecosystems. Ecol Stud 81:1–115.
Walsh C, Mac Nally R. 2013. hier.part: hierarchical partitioning. R package version 1.0-4. https://cran.r-project.org/web/packages/hier.part/.
Wastenson L, Fredén C, Eds. 2002. Sveriges Nationalatlas. Berg och Jord. Vällingby: Sveriges Nationalatlas.
Wiens JA. 1989. Spatial scaling in ecology. Funct Ecol 3:385–97.
Zinko U, Seibert J, Dynesius M, Nilsson C. 2005. Plant species numbers predicted by a topography-based groundwater flow index. Ecosystems 8:430–41.
Acknowledgments
We thank Henrik Weibull for inventorying and identifying all bryophytes in the study plots, Johan Lingegård, Julia Jansson, and Isak Lindmark for helping with the fieldwork, and three anonymous reviewers for valuable comments on the manuscript. Funding was provided by the Swedish Research Council Formas (to R. Jansson), SITES, Mistra Future Forests, and Formas Forwater (to H. Laudon), and SJCKMS Kempe foundation and Gunnar and Ruth Björkmans Fund for Botanical Research in Norrland (to L. Kuglerová).
Author information
Authors and Affiliations
Corresponding author
Additional information
Author contribution
LK performed research and data analyses with contribution from RJ and MD. LK wrote the manuscript with active contribution from all co-authors.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Kuglerová, L., Dynesius, M., Laudon, H. et al. Relationships Between Plant Assemblages and Water Flow Across a Boreal Forest Landscape: A Comparison of Liverworts, Mosses, and Vascular Plants. Ecosystems 19, 170–184 (2016). https://doi.org/10.1007/s10021-015-9927-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10021-015-9927-0