
Abstract
An important ecosystem function of vegetation on riverbanks is the control of soil erosion. Riverbanks also offer potential semi-natural habitats for grassland species. Previous classification of riverbank vegetation in Japan ignored both seasonal changes of dominant species and species composition, offering no information on ecosystem functions. Here we aimed at clarifying the association between dominant species, vegetation composition and species attributes in spring and autumn. We surveyed vegetation in 125 plots on a riverbank in Japan. We identified seven vegetation types on the basis of dominant species in autumn. Imperata cylindrica var. koenigii (Type I) and Pleioblastus chino (Type P) dominated their vegetation year-round. Dominant species changed seasonally in the other five vegetation types, but Lolium multiflorum was dominant in spring. Since the dominance of annuals is linked to fast turnover of root systems and thus poor control of soil erosion, Types P and I are superior in controlling soil erosion. Both the total number of species and the number of grassland species were small, showing species-poor grasslands in all vegetation types. There were no clear differences in numbers of species between species attributes. Nevertheless, this classification was linked to differences in species composition, reflecting several representative species in each vegetation type. Type P has more indigenous representative species and typical grassland species. Since some representative species may be suitable for natural processes (e.g., for pollination), this classification reflects biodiversity functions.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Biodiversity is important for ecosystem functions (Millennium Ecosystem Assessment 2005), which sustain both human needs (through supporting, provisioning, regulating and cultural functions) and ecological needs (through natural processes). The assessment of ecosystem functions is a valuable planning and management tool (Cardinale et al. 2012). Research should focus on both human and ecological needs at the same time for both existing and planned sustainable ecosystems (Cardinale et al. 2012; Higgs 2017).
Flooding caused by embankment breaches can have devastating consequences for surrounding areas. An important regulating ecosystem function of vegetation is the control of soil erosion (e.g., De Groot et al. 2002; Wallace 2007). Both above- and below-ground plant parts are important (Gyssels et al. 2005; Vannoppen et al. 2016). Vegetation cover reduces splash detachment and inter-rill erosion (Zuazo and Pleguezuelo 2009). Plant roots are effective in controlling concentrated flow erosion and shallow mass movements by contributing to both mechanical and hydrological soil properties (e.g., Simon and Collison 2002; Eviner and Chapin 2003), and they are more pronounced on splash detachment and inter-rill erosion (e.g., Zuazo and Pleguezuelo 2009; Shinohara et al. 2016). Although less reliable than below-ground traits, above-ground traits are useful for predicting root length density when information on below-ground traits is lacking (Helsen et al. 2016).
Riverbanks are traditionally revegetated with perennial grass species in Asia (Ezaki and Sakurai 1992; Lee and Nakagoshi 2010) and in Europe (Coppin and Stiles 1995; Scheres and Schüttrumpf 2019), which provide year-round soil cover, limiting erosion sometimes even with continued biomass harvest (Zuazo and Pleguezuelo 2009). They support typical species-rich plant communities (Liebrand and Sykora 1996; Bátori et al. 2016; Koyanagi et al. 2019). This also enhances other trophic groups, leading to important roles as semi-natural grassland in natural processes (e.g., habitat creation, ecological connectivity enhancement, integrated pest control; Reijers et al. 2014; Korvroëdan et al. 2021). Berendse et al. (2015) and Gould et al. (2016) demonstrated the positive influences of greater plant species richness and functional diversity on preventing soil erosion in grasslands on a simulated embankment. If this is applicable for real riverbanks, it is important to evaluate the effectiveness of components of species diversity in mitigating runoff and soil erosion (Hazebroek and Sprangers 2002). However, positive influences are still debatable. Scheres and Schüttrumpf (2020) suggested that species mixtures with a greater percentage of herbs and higher diversity lead in early development stages and lessen erosion resistance relative to a standard grass-dominated mixture. Thus, the engineering of embankments should consider both ecological needs and human needs at the same time for more sustainable design (Sasaki et al. 2000).
In Japan, means of improving the cost efficiency of managing embankments include reducing the frequency of mowing, which leads to unfavourable vegetation changes that replace short-grass monocultures with taller species (Asami et al. 1994). The mown plant material is removed and incinerated to improve surface visibility on riverbanks. At a given root density, a fibrous root system is hypothesized to have a larger erosion-reducing potential than a taproot system, owing to the larger root–soil contact (Dissmeyer and Foster 1985). Taller grass species are less suitable for land managers owing to their sparse root systems (Sasaki et al. 2000), and their large above-ground biomass is unsuitable because of the high incineration costs (Yamamoto and Yatsuki 2019).
The distribution of vegetation can be patchy along riverbanks (Koyanagi et al. 2019; Tisserant et al. 2021). Vegetation classification is important in evaluating the ecosystem functions in riverbank sections. Riverbanks are currently classified on the basis of dominant species. This is reasonable because local river managers can identify the vegetation type easily, even if they are unfamiliar with the species; and the mass ratio hypothesis (Grime 1998) stipulates that ecosystem properties are driven by the traits of the dominant species in the community. Sasaki et al. (2000) indicated seven vegetation types on riverbanks in Japan, among which they paid attention to three major types, dominated in turn by Zoysia japonica, Imperata cylindrica var. koenigii and Lolium multiflorum. They measured root densities as an indicator of the potential to control soil erosion (Hattori et al. 1997) and found that vegetation dominated by Z. japonica had the highest root density, and therefore high soil erosion control, followed by I. cylindrica var. koenigii and L. multiflorum. However, this classification does not take into account floristic composition: it remains unclear whether it is linked to differences in floristic diversity (i.e., species richness in each species attribute) among vegetation types. Moreover, it does not focus on seasonal vegetation change. Although L. multiflorum is replaced by other species in summer and later, the identity of those species is unclear. Understanding the relationship of vegetation between spring and autumn is important to evaluating ecosystem functions year-round.
This study was designed to compare the vegetation of major vegetation types on a riverbank in spring and autumn. We aimed at clarifying the association of both dominant species, floristic composition and species attributes between spring and autumn, and we discuss the evaluation of ecosystem functions desirable for riverbanks.
Materials and methods
Study site
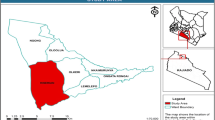
The study site is located in Ageo, Saitama Prefecture, Japan (35°56′20″N, 139°33′03″E; Fig. 1). The mean annual precipitation (30 years period, between 1991 and 2020), recorded at the nearby Saitama meteorological station, is 1371 mm; the mean annual temperature is 15.2 °C, the mean minimum is 3.9 °C (January), and the mean maximum is 27.0 °C (August) (JMA 2022). The study site is on the east bank (left bank), and both the land- and the river side of the Arakawa River, with a slope of 20°. The vegetation was mown twice a year, which is conventional for maintaining the riverbank. This bank was constructed before the 1950s. Since then, no broadscale reconstructions have taken place. Nevertheless, patchy maintenance (introduction of surface soil and subsequent planting of Z. japonica or Ophiopogon japonicus) has been done, though it is difficult to clarify where and when (personal communication with H. Sugama, former Head of the Organization for the Conservation of Arakawa River, Japan).

Location of the study site
Vegetation survey
The data were obtained by one of us (Saito H.) to evaluate the effects of past grazing activities on the vegetation composition on the riverbank. The riverbank had been grazed by cattle owned by a farmer until sixty years ago. According to the interview with the farmer (Sugama H.) the nearer the location of the riverbank, the higher the grazing intensity. Vegetation plots were established on the riverbank which had been grazed until sixty years ago along 1700 m of the riverbank. The effects of grazing on the vegetation were unclear (unpublished data). The survey monitored a total of 43 transects from the bottom to the top of the riverbank. In each transect, up to six consecutive quadrats were established, for a total of 125 plots. To monitor species diversity, the percentage cover of each species was estimated by eye from 15 to 28 May, on 18 June and on 6 and 11 September 2016. The plot size was 3 m × 3 m (9 m2) in 120 plots but was either 6.0 m2 or 7.5 m2 in 5 plots owing to limited slope length. The difference in the number of species between those 5 plots and the other 120 plots was not significant (P = 0.140). Hence, we used all 125 plots for the analyses.
Major dominant species
We based our classification of vegetation type on dominant species in autumn (Sasaki et al. 2000). Dominant species in each plot were defined as those with highest cover and those with ≥ 20% cover.
Species attributes
All plants collected were classified as exotic or native and as annuals (plus biennials), perennial herbs or woody species, as per Numata and Yoshizawa (1979). Characteristic species representing grassland vegetation in Miscanthetea sinensis communities were defined as “typical grassland species” (Miyawaki 1994). Following Chiba Historical Materials Research Foundation (2003), all species were distinguished among short plants, medium-statured plants and larger perennials. Plant height classification was applied only to perennials (though not climbers), too, as per Numata and Yoshizawa (1979). The reference classified P. chino as tall, but it reached only 50 cm in our study area, as seen also on other riverbanks (Yamada et al. 2017). Continuously mown P. chino remains low for several years even if mowing is resumed (Hori et al. 1998). Therefore, we classified P. chino as medium-statured. Nomenclature is based on BG Plants (Yonekura and Kajita 2003).
Analysis
To confirm any differences in species composition among vegetation types, we analysed the data by detrended correspondence analysis (DCA; Hill and Gauch 1980) of presence–absence data. To evaluate the occurrence of each species in different vegetation types and in different seasons, we performed indicator species analysis (INSPAN, Dufrêne and Legendre 1997) of cover data. INSPAN identified “representative species” considered representative of each habitat type (P < 0.05, Monte Carlo test; McCune and Mefford 1999) in PC-ORD for Windows v. 6 software (McCune and Mefford 1999).
We calculated one-way ANOVA and Tukey’s post hoc test to test differences in number of species, coverage and DCA axis scores between vegetation types. We modelled numbers of species with a Poisson error distribution, coverage with a negative binomial error distribution and DCA axis scores with a Gaussian error distribution. P < 0.05 was considered significant. Statistical analyses were performed in R v. 2.13.1 software (R Development Core Team 2015). As the monitoring period in spring spanned 2 months and thus may affect species cover, we included the month (May or June) as an explanatory variable in models of cover in spring. Because total vegetation cover was ~ 5% lower in June than in May and it is unlikely that any species would be competitively excluded under the less competitive conditions, we believe that the difference in the number of species and vegetation composition in the survey period had little effect.
Evaluation of ecosystem function in each vegetation type
We used vegetation cover as an indicator of the control of splash detachment and inter-rill erosion (Zuazo and Pleguezuelo 2009; Shinohara et al. 2016). We used the cover of annuals as an indicator of root instability (Scheres and Schüttrumpf 2019), cover of tall species as an indicator of incineration cost, and species richness, number of indigenous species and occurrence of typical grassland species as indicators of positive natural processes (Reijers et al. 2014; Korvroëdan et al. 2021). As the precise assessment of the ecological function and conservation value of different habitats cannot be based solely on species-based diversity indices, which are not sensitive to functional redundancy and other functional consequences of species identity (Petchey and Gaston 2006), we also evaluated each representative species in each vegetation type to evaluate natural processes.
Results
General description
We recorded 161 species, including 1 unknown: 56 annuals, 95 perennials and 9 woody species (Appendix 1). Of these, 104 species were indigenous and 56 were exotic. Seven species were typical grassland species; the most frequent was Arundinella hirta, followed by Miscanthus sinensis and Cirsium oligophyllum. In spring, half of the plots were dominated by Lolium multiflorum (Table 1). In autumn, I. cylindrica var. koenigii, S. altissima, Causonis japonica, P. chino and Sorghum halepense were dominant, with frequencies of > 5% (7/125 plots).
Most plots (108) were represented by seven vegetation types. Pleioblastus chino (Type P) and Imperata cylindrica var. koenigii (Type I) are indigenous perennials. Solidago altissima (Type Sa) and Sorghum halepense (Type Sh) are tall exotic perennials. Climber-dominated plots, all featuring and most dominated by Causonis japonica, were defined as Type C. Plots dominated by Ophiopogon japonicus or Zoysia japonica, both planted during construction of the riverbank, were combined as short species (Type OZ). Plots dominated by annuals such as Bidens pilosa var. pilosa were defined as Type A. Ten plots were dominated by other species, and seven plots had no dominant species. Plots in Type P and plots in Type I in autumn were linked to dominance by the same species in spring (Fig. 2). Most plots in Types Sa, C, A, Sh and OZ in autumn were dominated by Lolium multiflorum in spring.

Change of dominant species between spring and autumn. Values show the number of plots. Bold arrows show that more than half of plots in each vegetation type (right) originated from a dominant species
Floristic composition
The proportion of variance in the distance matrix of the DCA ordination in autumn totalled 0.539 on the first three axes (1, 0.391; 2, 0.097; 3, 0.051; Fig. 3). Plots in Types P and I were located to the left. Their scores differed significantly from those of the other five types on axis 1 (Appendix 2). Differences in scores between Types P and I on axes 1, 2 and 3 were not significant. Scores of Type OZ differed significantly from the other six types on axis 2. The proportion of variance in the distance matrix of the DCA ordination in spring totalled 0.497 on the first three axes (1, 0.250; 2, 0.142; 3, 0.105; Fig. 4). Again, scores of Types P and I differed significantly from those of the other five types on axis 1. Scores differed significantly between Types P and I on axis 3. The other five types were not significantly distinguished on any DCA axis.

Mean and SD of ordination scores in each quadrat in autumn by detrended correspondence analysis (DCA) in the first three dimensions

Mean and SD of ordination scores in each quadrat in spring by detrended correspondence analysis (DCA) in the first three dimensions
Number of species and cover values in each species attribute
Vegetation cover tended to be low in Type A in autumn but was generally > 80% in all vegetation types (Table 2). Cover of tall species was significantly larger in Types Sa and Sh than in the other five vegetation types, which nevertheless had moderate cover. All vegetation types had high total cover in spring, too. Species richness was significantly lower in Type OZ than in Type Sa in spring and autumn but was not significantly different between most vegetation types. There were more perennials than annuals and more indigenous than exotic species in all vegetation types. Type P averaged significantly more typical grassland species than several other vegetation types in autumn, but still < 1.0.
Representative species
Solidato altissima and Paederia foetida in Type Sa and Zoysia japonica, Ophiopogon japonicus and Calystegia pubescens in Type OZ were representative in both spring and autumn (Table 3). All representative species in Type P were native perennials.
Discussion
Association of vegetation types in spring and autumn
Species dominant in both spring and autumn were linked to an extent (Fig. 2). Dominant species in Types P and I were consistent throughout the year in most plots. Conversely, dominant species changed seasonally in the majority of plots in the other five vegetation types. Most of these plots were dominated by L. multiflorum in spring. Most plots in Types Sa, Sh, A, C and OZ were dominated by L. multiflorum in spring. Types Sa and Sh produced tall vegetation in summer and autumn, whereas Types C and OZ produced shorter vegetation. Yet despite this difference, the vegetation compositions were not distinct in either season (Figs. 3, 4). A high dominance of L. multiflorum until June would strongly determine species available in these plots in spring and thereafter. Indeed, several species dominant in autumn can grow fast: warm temperatures promoted rhizome bud sprouting and rapid shoot growth in Sorghum halepense (Monaghan 1980); and climbers are generally known to quickly spread over the ground (Dickinson et al. 2021).
Although vegetation composition in Type I was not distinct from that in Type P (Fig. 3), the cover of exotic species and L. multiflorum was significantly larger in Type I in spring (Table 2). Asami et al. (1994) reported vegetation characteristics of I. cylindrica var. koenigii on riverbanks and suggested that vegetation dominated by it was composed mostly of perennials, along with 10% to 40% exotic species, as observed here. The root system of I. cylindrica var. koenigii stretches from shallow to deep soil layers (Tominaga 2003). This range would probably enable coexistence between I. cylindrica var. koenigii and L. multiflorum, with their roots in different layers (McKane et al. 2002).
Vegetation characteristics of this study site
We classified vegetation on a riverbank into seven major types on the basis of dominant species in autumn. Among them, Z. japonica, I. cylindrica var. koenigii, P. chino and annual weeds were reported as major vegetation types by Sasaki et al. (2000). The other three types were reported as dominant species on riverbanks elsewhere (Suzaki and Tanaka 2004; Aizawa et al. 2015; Yamane et al. 2016; Koyanagi et al. 2019; Yamada and Nemoto 2020). Therefore, we assumed that all seven vegetation types were more or less commonly distributed on riverbanks in Japan.
The literatures suggest that species-rich semi-natural grasslands on riverbanks in Japan are dominated by either I. cylindrica var. koenigii or P. chino (Yamada et al. 2017; Koyanagi et al. 2019). However, neither Type I nor Type P resembled semi-natural grasslands. Yamada et al. (2017) reported 34.1 species per 5 m2 in the semi-natural grasslands on a riverbank; our numbers were distinctly lower: 10.4 species in Type I and 12.8 species in Type P per 9 m2. These lower values are in line with the report that species-poor meadow has 10–20 species per 25 m2 and species-rich meadow has 30–40 species in 25 m2 in the Netherlands (Hazebroek and Sprangers 2002). We assume that all vegetation types belong to species-poor grasslands. One reason for the marked difference in species richness would be the difference in the cover of dominant species: total cover in Type P in autumn (175.9%) was significantly larger than the total cover in all other vegetation types except Sa (Table 2). Abe et al. (2015) reported that species-rich semi-natural grassland on a riverbank was sparse, with a biomass of ~ 350 g/m2. Vigorous overgrowth would inhibit the persistence of diverse species in the vegetation, as generally known in grassland (Vermeer and Berendse 1983).
Implications for the evaluation of vegetation on riverbanks
A major ecosystem function required of the vegetation on riverbanks is the control of erosion (De Groot et al. 2002; Wallace 2007). Since vegetation dominated by Z. japonica is desirable to control erosion (Hattori et al. 1997), Type OZ is superior among the seven vegetation types. However, it is generally difficult to maintain such low vegetation on lowland riverbanks by mowing twice a year (Asami et al. 1994). Indeed, Type OZ was overgrown by medium species to some extent in spring (Table 2). It is likely that Z. japonica and O. japonicus here were planted recently. Persistence of the vegetation type in this area is uncertain.
This study clarified the relationship of vegetation between spring and autumn, enabling us to evaluate the ecosystem function of vegetation on riverbanks all year round. The dominance of annuals is linked to fast turnover of root systems and thus poor control of erosion on riverbanks (Scheres and Schüttrumpf 2019). Types P and I were superior to other vegetation types (except Type OZ), being dominated by the same species throughout the year (i.e., P. chino and I. cylindrica var. koenigii, respectively), and were not dominated by annuals (e.g., L. multiflorum). Imperata cylindrica var. koenigii has relatively large root biomass (Hattori et al. 1997; Sasaki et al. 2000) and is assumed to be persistent. I. cylindrica has smaller above-ground biomass than taller species (Yamada et al. 2021). Thus, I. cylindrica is more suitable because of the low incineration costs than taller species. This is applicable to P. chino, too, because continuously mown P. chino remains short for several years even if mowing is resumed (Hori et al. 1998). Information on the root system of P. chino is lacking; further research is needed to evaluate its root system for the control of soil erosion. Information on the root systems of Types Sa, Sh, C and A is needed, too.
Dense vegetation cover is important to prevent erosion by rain and water flow (Zuazo and Pleguezuelo 2009; Shinohara et al. 2016). However, vegetation cover was consistently high (> 80%) in each vegetation type in both spring and autumn, and it was similar or much larger in the six other vegetation types than in Type OZ. The relationship between vegetation cover and soil erosion has usually been reported as a negative exponential curve in a wide range of environmental conditions (Gyssels et al. 2005), meaning that a wide range of vegetation offers similarly high soil erosion resistance. Therefore, vegetation cover in all vegetation types would be large enough to prevent erosion.
In terms of biodiversity, no clear difference was observed in total species richness between vegetation types, except for a lower number in Type OZ. Typical grassland species were low overall (Table 2). In this regard, no special attention should be paid to a particular vegetation type in relation to natural processes. However, the occurrence of individual species differed among vegetation types. Type P was characterized by indigenous species. Thus, to sustain the abundance of indigenous species, Type P is valuable. Two of the representative species in Type P are grasses (i.e., Luzula capitata and Bromus remotiflorus), which are generally important food sources for several Orthoptera species (Wünsch et al. 2012). Pollination is important in grassland species (Sydenham et al. 2022). Representative species included several with entomophilous flowers (e.g., Aster iinumae in Type C and Agrimonia pilosa var. japonica in Type Sa). Indeed, among frequent typical grassland species, Cirsium oligophyllum has entomophilous flowers and is more frequent in Type P. Land managers should prioritize such natural processes, considering landscape perspectives (e.g., proportion of grasslands in surrounding areas in relation to dispersal availability and habitat uniqueness; Bátori et al. 2020).
Conclusion
Since the positive influences of greater plant species richness and functional diversity on preventing soil erosion in grasslands are still unclear (Scheres and Schüttrumpf 2020), developments in the engineering of embankments, at least, should investigate both at the same time for their more sustainable and adaptive design (Sasaki et al. 2000). We can identify a clear seasonal pattern in dominant species on a riverbank. The ecosystem properties are driven mainly by the traits of the dominant species in a community (the mass ratio hypothesis; Grime 1998). Dominant species in spring can be estimated from those in autumn. Thus, dominant species in autumn offer an effective indicator of the potential to control soil erosion on riverbanks. Vegetation dominated by Imperata cylindrica var. koenigii is superior for erosion control in terms of the persistence of its dominance throughout the year and the scarcity of Lolium multiflorum in spring. Vegetation dominated by Pleioblastus chino is also superior in terms of the scarcity of L. multiflorum. Since the root architecture of P. chino is unclear, further research is needed to evaluate the potential of the vegetation type to control soil erosion.
Our plots featured generally species-poor grassland, without a clear difference in species richness between vegetation types. However, this vegetation classification is linked to species composition, which depends on the presence of representative species in each vegetation type. By this measure, the P. chino vegetation type is superior in terms of larger numbers of indigenous representative species and typical grassland species. Nevertheless, natural processes should be evaluated by multiple traits (e.g., support for pollinators and herbivores) (Petchey and Gaston 2006). The long flowering period and the presence of insect-pollinated plants in these secondary grasslands are favourable for pollinator assemblages and for palynivores in Europe (Bátori et al. 2020). Since such functional traits are not been generally assessed in Japan, further research is needed on this aspect.
Data availability
The datasets generated and/or analyzed during the current study are available in the Mendeley Data, at https://doi.org/10.17632/ybdkcnz9sj.1.
References
Abe S, Yamada S, Nemoto M (2015) Influences of gap size on growth of native plants transplanted into an Imperata cylindrica grassland: does growth form matter? J Jpn Soc Revegetn Technol 41:91–96 (in Japanese with English summary)
Aizawa A, Tanaka A, Kobayashi H, Kobayashi T (2015) Evaluation of the effect of alien plants on the indigenous grassland community on the river bank of the Tone canal. J Jpn Soc Reveget Tech 40:527–533 (in Japanese with English summary)
Asami K, Hattori T, Akamatsu H, Takeda Y (1994) Characteristics of the Imperata cylindrica var. koenigii-Erigeron annuus Community on the embankment slopes. J Phytogeogr Taxon 42:75–81 (in Japanese with English summary)
Bátori Z, Körmöczi L, Zalatnai M, Erdős L, Ódor P, Tölgyesi C, Margóczi K, Torma A, Gallé R, Cseh V, Török P (2016) River dikes in agricultural landscapes: the importance of secondary habitats in maintaining landscape-scale diversity. Wetlands 36:251–264
Bátori Z, Kiss PJ, Tölgyesi C, Deák B, Valkó O, Török P, Erdős L, Tóthmérész B, Kelemen A (2020) River embankments mitigate the loss of grassland biodiversity in agricultural landscapes. River Res Appl 36:1160–1170
Berendse F, van Ruijven J, Jongejans E, Keesstra S (2015) Loss of plant species diversity reduces soil erosion resistance. Ecosystems 18:881–888
Cardinale BJ, Duffy JE, Gonzalez A, Hooper DU, Perrings C, Venail P, Narwani A, Mace GM, Tilman D, Wardle DA, Kinzig AP, Daily GC, Loreau M, Grace JB, Larigauderie A, Srivastava DS, Naeem S (2012) Biodiversity loss and its impact on humanity. Nature 486(7401):59–67
Chiba Historical Materials Research Foundation (2003) Chibaken-no-shizenshi (4) Chibaken-shokubutsushi [Natural source book of Chiba Prefecture (4) Flora of Chiba Prefecture]. Chiba-Nippousha, Chiba (In Japanese)
Coppin N, Stiles R (1995) Slope stabilisation and erosion control a bioengineering approach. Morgan RPC, Rickson RJ (eds) Ecological principles for vegetation establishment and maintenance. E & F N Spon, Chapman & Hall, Cambridge, pp. 59–94
De Groot RS, Wilson MA, Boumans RM (2002) A typology for the classification, description and valuation of ecosystem functions, goods and services. Ecol Economics 41:393–408
Dickinson CC, Jelesko JG, Barney JN (2021) Habitat suitability and establishment limitations of a problematic liana. Plants 10:263
Dissmeyer GE, Foster GR (1985) Modifying the universal soil loss equation for forest land. Ecol Monographs 67:345–366
Dufrêne M, Legendre P (1997) Species assemblages and indicator species: the need for a flexible asymmetrical approach. Ecol Monographs 67:345–366
Eviner VT, Chapin FS (2003) Functional matrix: a conceptual framework for predicting multiple plant effects on ecosystem processes. Annu Rev Ecol Evol Syst 34:455–485
Ezaki T, Sakurai Y (1992) Standing crop and strength of root system of trees and grasses for landscaping. Jpn J Landsc Archit 55:181–186 (in Japanese with English summary)
Gould IJ, Quinton JN, Weigelt A, De Deyn GB, Bardgett RD (2016) Plant diversity and root traits benefit physical properties key to soil functions in grasslands. Eco Let 19:1140–1149
Grime JP (1998) Benefits of plant diversity to ecosystems: immediate, filter and founder effects. J Ecol 86:902–910
Gyssels G, Poesen J, Bochet E, Li Y (2005) Impact of plant root characteristics on the resistance of soils to erosion by water: a review. Prog Phys Geogr 29:189–217
Hattori A, Mochizuki T, Fujita K (1997) Estimation of erosion resistance of the levee slopes covered by vegetation which is managed by two cuttings per year. Proc Hydraul Eng 41:367–372 (in Japanese with English summary)
Hazebroek E, Sprangers JTCM (2002) Richtlijnen voor dijkgraslandbeheer. Wageningen, Alterra, Research Instituut voor de Groene Ruimte. Alterra-rapport 469 (in Dutch)
Helsen K, Vannoppen W, Honnay O, Poesen J (2016) Both below-ground and aboveground functional traits can help predict levee grassland root length density as a proxy for flow erosion resistance. J Veg Sci 27:1254–1263
Higgs E (2017) Novel and designed ecosystems. Restor Ecol 25:8–13
Hill MO, Gauch HG Jr (1980) Detrended correspondence analysis: an improved ordination technique. Vegetatio 42:47–58
Hori Y, Kawarasaki S, Kobayashi T (1998) Plasticity of C/F ratio of aerial parts and its ecological significance of a bamboo grass, Pleioblastus chino. J Jpn for Soc 80:165–169 (in Japanese with English summary)
JMA (Japan Meteorological Agency) (2022) Past Meteorological Data in Japan. http://www.data.jma.go.jp/obd/stats/etrn/index.php. Accessed 23 Sep 2022.
Korvroëdan L, Armand R, Rey F, Faucon M-P (2021) Trait-based sediment retention and runoff control by herbaceous vegetation in agricultural catchment: a review. Land Degrad Develop 32:1077–1089
Koyanagi FT, Yamada S, Matsuzaki H, Kato Y (2019) Impacts of previous maintenance of river embankments on the grassland communities by changing soil properties. Ecol Eng 131:73–80
Lee HJ, Nakagoshi N (2010) Grassland vegetation change after riverbank restoration in Jungnang River, Seoul, Korea. Hikobia 15:377–384
Liebrand CIJM, Sykora KV (1996) Restoration of semi-natural, species-rich grasslands on river dikes after reconstruction. Ecol Eng 7:315–326
McCune B, Mefford MJ (1999) PC-ORD multivariate analysis of Ecological data. Version 5.0. MjM software. Gleneden Beach, Oregon
McKane RB, Johnson LC, Shaver GR, Nadelhoffer KJ, Rastetter EB, Fry B, Giblin AE, Kielland K, Kwiatkowski BL, Laundre JA, Murray G (2002) Resource-based niches provide a basis for plant species diversity and dominance in arctic tundra. Nature 415(6867):68–71
Millennium Ecosystem Assessment (2005) Ecosystems and Human Well-being: Synthesis. Island Press, Washington
Miyawaki A (ed) (1994) Vegetation of Japan. Volume 7 Kanto. Shibundo Co., LTD. Publishers, Tokyo, p 641 (in Japanese)
Monaghan N (1980) The biology of Johnson grass (Sorghum halepense). Weed Res 19:261–267
Numata M, Yoshizawa N (eds) (1979) Weed flora of Japan. Nipponshokubutsuchouseizai-kenkyukyokai [Japan Association for Advancement of Phyto-Regulators], Tokyo (in Japanese)
Petchey OL, Gaston KJ (2006) Functional diversity: back to basics and looking forward. Ecol Letters 9:741–758
R Development Core Team (2015) R: a language and environment for statistical computing. r foundation for statistical computing, Vienna, Austria. http://www.R-project.org/. Accessed 23 Sept 2022
Reijers VC, Visser EJW, Paulissen MPCP, de Kroon H (2014) De invloed van vegetatie op de erosiebestendigheid van dijken: de start van een monitoringsexperiment naar de effecten van de vegetatiesamenstelling op de erosiebestendigheid van de Purmerringdijk. (Alterra-rapport; No. 2622). Alterra, Wageningen-UR. https://edepot.wur.nl/329789. Accessed 10 Jan 2023. (in Dutch)
Sasaki Y, Toya H, Ishibashi A, Isaka M, Hirata M (2000) Teiboushokusei-no-tokusei-to-teiboushokuseikanrikeikaku [Characteristics of riverbank vegetation and management planning of the vegetation]. Kasenkankyosougoukenkyujohoukoku 6:69–105 (in Japanese)
Scheres B, Schüttrumpf H (2019) Enhancing the ecological value of sea dikes. Water 11:1617
Scheres B, Schüttrumpf H (2020) Investigating the erosion resistance of different vegetated surfaces for ecological enhancement of sea dikes. J Marine Sci Eng 8:519
Shinohara Y, Otani S, Kubota T, Otsuki K, Nanko K (2016) Effects of plant roots on the soil erosion rate under simulated rainfall with high kinetic energy. Hydrol Sci J 61:2435–2442
Simon A, Collison AJC (2002) Quantifying the mechanical and hydrological effects of riparian vegetation on streambank stability. Earth Surf Processes Landforms 27:527–546
Suzaki T, Tanaka B (2004) Examination of mowing method which supports the inhabits of plants, insects and spiders on the embankment. Yahagigawa Kenkyu 8:149–169 (in Japanese with English summary)
Sydenham MAK, Venter ZS, Eldegard K, Moe SR, Steinert M, Staverløkk A, Dahlec S, Skoog DIJ, Hanevik KA, Skrindo A, Rusch GM (2022) High resolution prediction maps of solitary bee diversity can guide conservation measures. Lands Urban Plan 217:104267
Tisserant M, Bourgeois B, González E, Evette A, Poulin M (2021) Controlling erosion while fostering plant biodiversity: a comparison of riverbank stabilization techniques. Ecol Eng 172:106387
Tominaga T (2003) Growth of Seedlings and Plants from Rhizome Pieces of Cogongrass (Imperata cylindrica (L.) Beauv.). Weed Biol Manag 3:193–195
Vannoppen W, Honnay O, Poesen J (2016) Both below-ground and above-ground functional traits can help predict levee grassland root length density as a proxy for flow erosion resistance. J Veg Sci 27:1254–1263
Vermeer JG, Berendse F (1983) The relationship between nutrient availability, shoot biomass and species richness in grassland and wetland communities. Vegetatio 53:121–126
Wallace KJ (2007) Classification of ecosystem services: problems and solutions. Biol Conserv 139:235–246
Wünsch Y, Schirmel J, Fartmann T (2012) Conservation management of coastal dunes for Orthoptera has to consider oviposition and nymphal preferences. J Insect Conserv 16:501–510
Yamada S, Nemoto M (2020) Effects of weed abundance and frequency of hand weeding on the establishment of transplanted Imperata cylindrica. Landsc Ecol Eng 16:1–9
Yamada S, Nemoto M, Koyanagi FT, Yamamoto Y, Yatsuki H (2017) Vegetation and environmental attributes in species-rich semi-natural grasslands in rivedikes. J Jpn Soc Reveget Technol 42:428–432 (in Japanese)
Yamada S, Nemoto M, Okuro T (2021) Season and plant life history stage in revegetation influence competition of foundation species, subordinate species, and weeds in a reclaimed grassland. Ecol Eng 162:106178
Yamamoto Y, Yatsuki H (2019) Attempt to utilize pellets of levee mowed grass in cooperation with local residents. Kasensougoukenkyujohoukoku 24:27–35 (in Japanese with English summary)
Yamane A, Harada S, Uchida T (2016) Control of invasive alien species, Sorghum halepense (L.) Pers. flourishing on the river bank by cutting. J Jpn Soc Revegetn Technol 41:272–278 (in Japanese)
Yonekura K, Kajita T (2003) BG Plants Y List, an online service of Japanese plant names, including a nomenclature index. http://ylist.info/. Accessed 23 Sept 2022
Zuazo VHD, Pleguezuelo CRR (2009) Soil-erosion and runoff prevention by plant covers: a review. In: Lichtfouse E, Navarrete M, Debaeke P, Véronique S, Alberola C (eds) Sustainable agriculture. Springer, Dordrecht
Acknowledgements
We thank M. Hiroko Sugama (former head) and other members of the Organization for the Conservation of the Arakawa River for their assistance with field surveys. This research was partly supported by JSPS KAKENHI Grant Numbers JP#17H03958 and #20H03015 and by the River Fund of the River Foundation Japan.
Funding
Japan Society for the Promotion of Science, #20H03015, Susumu Yamada, #17H03958, Susumu Yamada
Author information
Authors and Affiliations
Corresponding author
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Yamada, S., Saito, H., Nemoto, M. et al. Relationship between dominant species, vegetation composition and species attributes in spring and autumn on a riverbank: implications for river management to enhance ecosystem functions. Landscape Ecol Eng 19, 435–446 (2023). https://doi.org/10.1007/s11355-023-00551-z
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11355-023-00551-z