
Abstract
Key message
Year-round observation suggests that sap flow in growing season represents species-specific transpiration characteristics and sap flow of deciduous broadleaved species in non-growing season is related to daily changes in stem diameter.
Abstract
Year-round observation is necessary to investigate species-specific characteristics of sap flow and its possible contribution in non-growing season. The middle Loess Plateau in China is characterized by temperate climate and vegetation cover of generally drought-tolerant deciduous broadleaved and some evergreen coniferous trees. In this study, Granier-type thermal dissipation probes were applied to measure year-round xylem sap flow in three species from April 2017 to April 2018. Solar radiation, air temperature, relative air humidity, precipitation, and soil moisture were monitored continuously. The results showed that the peak time of sap flux density (Fd) in the diurnal courses of Robinia pseudoacacia was later than that of Quercus liaotungensis and Platycladus orientalis. The peak time of Fd for R. pseudoacacia was significantly postponed during the period with higher soil moisture when compared with that at lower soil moisture, whereas the peak time was reached slightly earlier in the other two species. Sap flow during the non-growing season was relatively higher in P. orientalis than in the two broadleaved species. The diurnal course of Fd in non-growing season showed high values at nighttime and low values in daytime, opposite to that in growing season. Daily variation in stem diameter was inverse of the change in sap flow in the two broadleaved species, whereas the daily change in stem diameter of P. orientalis was very small and showed no clear relation with sap flow. The results suggest that changes in transpiration characteristics with soil moisture conditions vary with tree species; sap flow in deciduous broadleaved trees in non-growing season is small and closely related to daily changes in stem diameter.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Transpiration is the main pathway for plant water loss and constitutes a significant part of water budget in forest ecosystems (Siegert and Levia 2011). Sap flow, which represents the water transport from roots to leaves through the stem xylem, is commonly used to characterize transpiration process in relation to environmental and internal factors. Year-round observations of sap flow not only provide useful information about water use throughout the year, but also reveal species-specific characteristics in transpiration responses to meteorological conditions and soil moisture environments.
Transpiration in the growing season is interactively determined by environmental factors and species features. On a daily timescale, transpiration rate closely follows its meteorological driving factors, i.e., solar radiation (RS) and vapor pressure deficit (VPD) (e.g., Granier et al. 1996; Zhang et al. 2015). As transpiration is ultimately supported by soil water supply, it is highly dependent on the plant’s ability of absorbing water from soil. Transpiration often varies greatly among species at any point of a single wet-dry cycle due to differences in sensitivity to soil moisture changes (Oren and Pataki 2001). The influence of soil moisture on transpiration has been detected by many studies. Llorens et al. (2010) found that the transpiration rate in Scots pine (Pinus sylvestris) is strongly limited by soil water availability. Chang et al. (2006) showed that sap flow in Gansu poplar (Populus gansuensis) growing in a shelter belt experienced a remarkable improvement after soil moisture was significantly increased by irrigation. Previous studies have shown species differences in their responses to fluctuations in soil moisture. A study conducted in North Carolina suggested that transpiration of Acer rubrum had a greater response to soil moisture depletion than did Quercus alba, and ring-porous species had weaker stomatal sensitivity to soil moisture compared with diffuse-porous species (Oren and Pataki 2001). In a temperate broad-leaved forest, the relationship between sap flow and VPD changed with increasing soil drought in all species except in Fraxinus excelsior, suggesting that this species was the best adapted to prolonged drought (Kocher et al. 2009). In contrast, an investigation of four co-occurring tree species in a Mediterranean riparian forest indicated that these species were more limited by evaporative demand than by water availability, while ash (F. excelsior) experienced transpiration constraints at soil water content less than 8% (Nadal-Sala et al. 2017). These findings were mainly inferred from sap flow measurements based on either instantaneous data or daily sums or averages. The analyses of responses to environmental factors based on diurnal patterns have been less reported. Investigation of the transpiration characteristics among species should be helpful to quantify the impact of short-term soil moisture deficit and evaluate potential adaptation of the tree species to future climate regimes.
During a non-growing season, physiological activities are extremely weak in temperate trees, whereas evergreen species may exhibit stomatal transpiration through leaves. However, quantitative measurements are relatively few, particularly those on possible sap flow in deciduous species. Sap flow of low intensity was detected in Norway spruce (Picea abies) in winter months and it reacted to temperature changes, suggesting that spruce was physiologically active during this dormant period (Matisons et al. 2017). A measurement using eddy covariance in a subtropical plantation of several evergreen species detected winter transpiration and suggested that the transpiration was closely correlated with environmental factors (Song et al. 2006). By monitoring oxygen potential difference in tree trunk of Tilia cordata, Totzke et al. (2017) concluded that physiological activity was still present in the leafless state in winter. Nadezhdina et al. (2010) observed a weak sap flow during winter in several tree species in Europe, but they contributed it to water redistribution driven by water potential difference, rather than transpiration, caused by the cold weather conditions. These results highlight the need to study the existence of sap flow during non-growing season to better understand the inadequately studied water consumption in cold conditions.
The semiarid Loess Plateau in central China has a temperate climate and the vegetation cover consists of deciduous broadleaved and some evergreen coniferous trees that are generally drought tolerant. These species show seasonal changes in foliage conditions and physiological characteristics. Water availability during the growing season can be a limiting factor affecting the plant growth and ecosystem stability (Tsunekawa et al. 2014). In this study, we investigated sap flow characteristics throughout the year in three tree species from two adjacent stands. The main objectives were to (1) comparatively analyze the responses of sap flux density to environmental factors among the three tree species based on diurnal patterns and variations under different soil moisture conditions to obtain species-specific water use strategies and adaptations to arid environments, and (2) explore the possible sap flow in non-growing season and its determining factors to provide comprehensive insight into year-round transpiration.
Material and methods
Study site
The study was conducted at Mount Gonglushan (36° 25.40′ N, 109° 31.53′ E, 1353 m a.s.l.) near Yan’an city, Shaanxi Province in northern China. The area is located in the ecological transition zone between forest and forest-steppe ecosystems with a temperate semiarid climate. The mean annual precipitation and air temperature during 1971–2010 were 504.7 mm and 10.1 °C, based on data obtained from the local meteorological station (Shi et al. 2014). Precipitation mainly occurs from July to September. Details of the study site are presented by Du et al. (2007).
We investigated three tree species, two deciduous broadleaved species, Quercus liaotungensis and Robinia pseudoacacia, and an evergreen coniferous species, Platycladus orientalis. Q. liaotungensis and P. orientalis are present in a natural secondary forest dominated by Q. liaotungensis, the major natural forest type in the region. R. pseudoacacia, a fast-growing tree species that was the main reforestation species in the region, is located in an approximately 30-year-old plantation. There was a permanent research plot in each of the two forest stands for multipurpose investigations of, for example, forest productivity, evapotranspiration, soil moisture dynamics, and soil microbial biology (Du et al. 2011; Shi et al. 2014; Tian et al. 2017; Yan et al. 2016; Zhang et al. 2015).
Sap flow measurement
Xylem sap flow was measured in five individuals for each species from April 2017 to April 2018 (Table 1) using Granier-type sensors (Granier 1987). Each sensor consisted of a pair of cylindrical probes (10 mm long, 2 mm in diameter): a continuously heated upper probe, supplied with constant power at 0.15 W, and an unheated lower probe as a temperature reference (James et al. 2002). Each probe pair was inserted into the sapwood of the sample trees vertically 15 cm apart on the northern side of the stem at breast height. The temperature difference between the probes was converted into a voltage value and the average at every 30 min was recorded in a data logger (CR1000, Campbell Scientific Inc., Logan, UT, USA) with a multiplexer (AM16/32, Campbell Scientific, USA).
The measured temperature difference was converted to sap flux density using the empirical relationship determined by Granier (1987):
where Fd is the sap flux density (mL m−2 s−1), and K is dimensionless and related to the temperature difference between the two probes ΔT:
where (ΔTmax) is the maximum value between heated and un-heated probe when sap is not moving (Fd = 0).
The sapwood thickness of some sample trees was less than the length of the probe (10 mm). To avoid underestimation of Fd in those trees, corrected values (ΔTSW) were used instead of measured values (ΔT) for the temperature difference in Eq. 2, calculated following the correction equation proposed by Clearwater et al. (1999):
where a is the proportion of the probe in the sapwood, ΔT is the measured temperature difference between the two probes, and ΔTSW is the actual temperature difference between the two probes.
Sapwood area and thicknesses of sample trees were calculated by core sample analysis. Core samples were collected using an increment borer. For Q. liaotungensis, regression equation derived from a previous study was used to calculate sapwood area (Yan et al. 2016). Sapwood area of R. pseudoacacia trees was determined by regression equation of AS with DBH that was derived from core samples taken from 15 randomly selected trees as AS = 0.1197 × DBH1.5546. For P. orientalis, core samples were taken from five sample trees due to the limited number of trees from this species, and they were used to calculate sapwood area of this species. As Fd may vary among individuals within each species, the sapwood area-weighted average was calculated by multiplying each Fd with different weight according to the tree’s relative amount of sapwood area.
Meteorological and soil moisture measurements
Meteorological factors, namely solar radiation (RS), air temperature (T), relative humidity (RH), and precipitation (P), were measured in an open area outside the stands. Solar radiation was measured using a pyranometer (Li-200; Li-Cor Inc., Lincoln, NE, USA), and air temperature and relative humidity were measured using a thermohygrograph (HMP50; Vaisala, Helsinki, Finland). The pyranometer and thermohygrograph were mounted 2 m above the ground. Precipitation was measured using a tipping bucket rain gauge (Davis 7852; Davis Instruments, Hayward, CA, USA) at approximately 0.3 m above ground. These data were sampled every 30 s and average values were recorded every 30 min in a CR1000 data logger (Campbell Scientific, USA). Vapor pressure deficit (VPD) was calculated from air temperature and relative humidity according to Campbell and Norman (1998), as follows:
Soil water content of the 0–100 cm soil profile in each plot was monitored with S-SMC-M005 soil moisture sensors connected to a HOBO data logger (H21-001; Onset Computer Corporation, Bourne, MA, USA) and recorded as one-hour averages. The sensors were installed at depths of 6, 12, 30, 50, 70, and 90 cm. Weighted average calculation was used to obtain the soil moisture value of the 0–100 cm layer.
Dendrometer measurements
To further elucidate the daily sap flow characteristics in non-growing season, stem diameter variation experiment was conducted during the non-growing season (November–March) of 2018 using a dendrometer (Type DC3; Ecomatik, Dachau, Germany). Three trees of each species were selected from among the sample trees in which Fd was also measured. The dendrometers were mounted on the north face above the sap flow sensors and covered with aluminum foil to protect them from direct solar radiation and contact with water. Dendrometer data were recorded every 1 h using a data logger (UX120-006M; Onset, USA).
Data analysis
We assumed that stems exhibited zero growth during non-growing season. The minimum value obtained from dendrometer measurements for each day was used as reference value. Stem diameter variation was the difference between current stem radial reading and the minimum value.
Significant difference in the peak time of Fd among the three species and between different soil moisture conditions were assessed with t-test. SPSS 16.0 (IBM Corp., Armonk, NY, USA) and SigmaPlot 14.0 (Systat Software Inc., San Jose, CA, USA) were used for statistical analysis and graph plotting, respectively.
Results
Climate factors and soil water conditions during the study period
Climate factors presented obvious seasonal variations in that the values of each factor rose gradually from April, decreased in October, and started increasing in February of next year (Fig. 1). We thus defined the growing season as a period from April to October, and the non-growing season from November to March. The daily mean temperature was 10.46 °C during the entire study period, 17.66 °C in the growing season, and 0.89 °C in the non-growing season. Fluctuations in temperature and relative humidity resulted in a large range of VPD (0.04–3.17 kPa). The maximum daily RS was 33.44 MJ m−2.

Daily sum of solar radiation (RS), mean air temperature (T), mean relative humidity (RH), and mean vapor pressure deficit (VPD) during the study period
Precipitation was unevenly distributed during the study period (Fig. 2). Most rainfall in the growing season occurred from July to October (83%). The trend in soil moisture in the two plots was identical during the study period. Soil moisture declined since the start of the growing season due to tree germination and growth. When entering summer, changes in soil moisture closely followed the precipitation pattern. Pronounced rise in soil moisture was observed between 13 and 29 July after two rainfall events. Soil moisture increased again after another rainfall event of 136.4 mm. Total rainfall during 2–11 October was 92 mm, which clearly recharged the soil water.

Changes in soil moisture in Robinia pseudoacacia plot (soil moisture-R) and Quercus liaotungensis plot (soil moisture-Q) and precipitation during the study period
Variations in diurnal course of sap flow during the growing season
The diurnal courses of RS, VPD, and sapwood area-weighted average Fd for each tree species on a representative bright day are shown in Fig. 3. The diurnal courses of Fd generally coincided with those of environmental factors, but they were not necessarily synchronized. RS reached its maximum at 13:00, 1.5 h ahead of VPD. Fd in all species was close to zero from 00:00 to 06:00, gradually increased after sunrise, and reached its daily peak earlier than or simultaneously with RS and VPD; it decreased to a low level towards sunset and reached the minimum at midnight. The value of Fd peaked relatively late in R. pseudoacacia and early (at around 11:00) in Q. liaotungensis, whereas in P. orientalis, it showed an intermediate pattern between the other two species.

Diurnal courses of solar radiation (RS), vapor pressure deficit (VPD), and sapwood area-weighted average sap flux density (Fd) for Platycladus orientalis, Robinia pseudoacacia, and Quercus liaotungensis on 20 June, 2017
To further clarify these differences among the species, peak times of Fd, encompassing the data sets for rain-free days and daytime maximum VPD ≥ 2 kPa, were calculated for the period June–September. The data sets were divided into two periods based on soil moisture conditions, those before and after an intense rainfall event, when 111.6 mm of shower precipitation fell between 13 and 18 July (Table 2). The peak time of Fd in R. pseudoacacia occurred significantly later compared with that in P. orientalis and Q. liaotungensis under both soil moisture conditions, and it was distinctly delayed under higher soil moisture mainly owing to relatively longer hours of active sap flow. However, in P. orientalis and Q. liaotungensis, Fd in higher soil moisture period peaked slightly earlier than it did at lower soil moisture, probably due to rapid increase in sap flow in morning hours under higher soil moisture conditions; nevertheless, these differences were not statistically significant in both species.
Contribution in sap flow in growing season and non-growing season
The average Fd of the three species over the whole growing season and non-growing season is presented in Fig. 4. The mean Fd of the three species in the non-growing season ranged from 0.65–3.03 mL m−2 s−1, and it was significantly lower than those measured in the growing season (4.68–17.39 mL m−2 s−1). Different from that in the growing season, the Fd values in the non-growing season decreased from P. orientalis to R. pseudoacacia and to Q. liaotungensis. The ratio between Fd in the two periods was the highest in the evergreen conifer P. orientalis. The two deciduous species showed smaller ratios, but revealed the existence of sap flow in the non-growing season.

Average sap flux density (Fd) in Platycladus orientalis, Robinia pseudoacacia, and Quercus liaotungensis in the growing season (201 days) and non-growing season (151 days), and the ratio between the two periods. Error bars represent the unit weight medium error (n = 5)
Sap flow diurnal course and its affecting factors in non-growing season
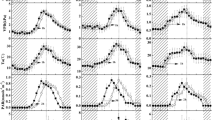
The diurnal variations in sap flow, stem diameter variation, and major meteorological factors on two typical days of non-growing season are presented in Fig. 5. As the sap flow was small in the non-growing season, we presented K instead of Fd for better interpretation of diurnal variations. The results showed that the diurnal courses of sap flow in non-growing season were almost the opposite of those in the growing season; they reached the lowest levels at daytime and the highest at nighttime. For P. orientalis and Q. liaotungensis, K decreased to the lowest levels at around 13:00 and in R. pseudoacacia at around 16:00, later than in the former two species. In addition, the diurnal courses in stem diameter variation were similar for all species. The dynamics of diameter variation reflected temperature values, but they were not necessarily synchronized. The diameter variation continuously decreased from 00:00, and reached the smallest value around 09:00, when it started expanding, until reaching the maximum at about 18:00, and then decreased again and entered the next daily cycle. Furthermore, the dynamics of sap flow showed an opposite trend in daily changes to that of diameter variation in all species. Diurnal lag effects were observed in all species, showing that diameter variation lagged behind sap flow.

Diurnal courses of solar radiation (RS), temperature (T), vapor pressure deficit (VPD), K (a dimensionless parameter related to temperature difference between probes), and stem diameter variation on 9–10 December, 2018
To illustrate the factors affecting sap flow in non-growing season, responses of the hourly sap flow to T, VPD, and diameter variation during a 10-day period under normal weather conditions were analyzed (Fig. 6). The results suggested that K had a significantly negative linear correlation with the three factors, except for P. orientalis in which no significant relationship was detected between K and diameter variation. In addition, K of P. orientalis showed smaller correlation coefficients with T and VPD compared with those in other two species, and K of Q. liaotungensis and R. pseudoacacia showed higher level of correlation with T.

Relationships between K (a dimensionless parameter related to temperature difference between probes) and stem diameter variation, temperature (T), and vapor pressure deficit (VPD). The data sets covered hourly data for 10 days. Linear function y = ax + y0 was fitted to several data sets, where y is K and x is the factor affecting the sap flow
Discussion
Sap flow characteristics and responses to soil moisture during growing season
Typical daytime patterns of the sap flux density and responses of the sap flow to soil moisture in tree species may indicate species-specific water use strategies (Du et al. 2011; Kocher et al. 2009; Yan et al. 2018). Reaching a peak in sap flow earlier suggested that the plants quickly initiated stomatal regulation to preserve water, whereas later peak time indicated they actively transpired water for longer time. In P. orientalis and Q. liaotungensis, Fd peaked earlier and there was no significant difference in Fd peak time between two soil moisture conditions, implying an early stomatal regulation of transpiration in response to increasing VPD and lower sensitivity of the transpiration process to changing soil water conditions. The major natural forest type on the Loess Plateau is the secondary forest dominated by Q. liaotungensis. A 3-year study on stand transpiration of the secondary oak forest at our study site showed that this forest maintained a normal magnitude of annual water consumption (Yan et al. 2016). Another study reported a relatively small amplitude of sap flow variation in Q. liaotungensis compared with that in R. pseudoacacia in response to soil water changes (Du et al. 2011). The present study further demonstrated that Q. liaotungensis tends to regulate its stomata earlier compared with the other two species to maintain stable transpiration during a relatively long period with a wider range of soil moisture conditions.
The native evergreen coniferous species, P. orientalis, is highly adaptable to drought in the Loess Plateau (Wang et al. 2017). Similarly, a study conducted in northern China suggested that P. orientalis can seize every opportunity to store water and thus rapidly adjust its water use pattern to the soil moisture conditions (Liu et al. 2019; Niinemets et al. 2011). Lu et al. (2019) indicated that P. orientalis is effective and responsive to water supply and better adapted to semiarid areas. These findings were corroborated by our results which showed that P. orientalis tends to reduce its transpiration earlier in the morning of a day compared with R. pseudoacacia to use water more conservatively.
Fd in R. pseudoacacia peaked significantly late compared with other two species and under higher soil moisture conditions, indicating an extended transpiration without stomatal closure under sufficient soil moisture supply. This was consistent with the findings by Mantovani et al. (2014), who reported that transpiration of R. pseudoacacia at both the individual-leaf level and at the whole-plant level was not limited by the stomata at optimal soil moisture and, therefore, concluded that the species is not a water-saving tree species. A 3-year study conducted on the semiarid Loess Plateau similarly showed that transpiration in R. pseudoacacia was not regulated due to lower sensitivity of stomata to VPD (Chen et al. 2014). Lower stomata sensitivity indicated that the trees vigorously transpired under both soil moisture conditions. R. pseudoacacia may undergo physiological and structural adjustment to minimize water loss and avoid hydraulic system failure under drought conditions, whereas the transpiration would largely increase after soil moisture was recharged by rainfall following short-term drought (Breda et al. 2006; Mantovani et al. 2015; Zhang et al. 2015). However, prolonged drought would suppress the capacity of transpiration resilience by causing recovery insufficient and changing leaf traits (He et al. 2020). In addition, similar observations in R. pseudoacacia and other indigenous species suggested a larger water demand and higher influence of soil water conditions on transpiration of R. pseudoacacia (Du et al. 2011; Wang et al. 2017). Yan et al. (2013; 2010) found that, compared with Q. liaotungensis, R. pseudoacacia exhibited a higher water use based on leaf area and larger difference between water potential at midday (Ψmd) and estimated water potential at turgor loss point (Ψtlp), implying a more active transpiration during daytime and thereby increased water deficiency in their leaves. R. pseudoacacia stands have been declining in recent years, and the forest composition is changing in favor of P. orientalis (Wang et al. 2017). Our results indicate that, due to the risky water use strategy of R. pseudoacacia, P. orientalis may negatively impact R. pseudoacacia when the two species compete for water resources, which suggests that a likely forest succession in the mixed forest would be favorable to P. orientalis (Chen et al. 2018; Wang et al. 2017). Species-specific water use differences may ensue from different wood hydraulic conductivity, rooting patterns, and soil water uptake depths (Kunert et al. 2010; Nadal-Sala et al. 2017). Variations in water use and physiological traits between wet and dry years need further investigation to clarify water use strategies and long-term hydrological regime of these species.
Characteristics and factors influencing sap flow in non-growing season
Water loss during the non-growing season is an important component of the annual plant water budget, although previous studies have neglected this process. In our study, the ratio of Fd between non-growing and growing season was 0.14–0.35. The evergreen species P. orientalis had the highest sap flow during this season among the tree species. Our estimates were lower than the value reported for Myrica faya (0.52), but higher than that of Laurus azorica (0.30), two broad-leaved evergreen species present in a laurel forest with humid Mediterranean climate (Jimenez et al. 1996, 1999). Evergreen species need to maintain an adequate water balance in their foliage to survive the winter season (Boyce and Lucero 1999). Sap flow in two Abies forests located at different elevations and with a continental temperate climate showed that high-elevation species did not transpire during the winter period contrary to low-elevation species (Chan and Bowling 2017). This could be attributed to the elevation-caused differences in temperature and liquid water available for sap flow.
In our study, sap flow in deciduous species was detected even after losing all their foliage, although at a relatively low ratio. In Panamanian forest plantations, normalized daily sap flux density of the broad-leaved deciduous species Cedrela odorata was less than 6% when trees shed all their leaves (Kunert et al. 2010). Yoshifuji et al. (2011) monitored the sap flow of the tropical deciduous species Tectona grandis in northern Thailand for 8 years and found that sap flow existed when the leaf area index was zero. Fisher et al. (2007) indicated that daytime and nighttime sap flow of the broad-leaved deciduous species Quercus douglasii and that of the coniferous evergreen species Pinus ponderosa in regions of California with Mediterranean climate were low in winter, and the values for evergreen species were higher than those for the deciduous species; these findings were similar to the results of our research.
Our results showed that the variation in stem diameter followed a similar diurnal course with that for temperature. Zweifel and Hasler (2000) reported similar results for subalpine Norway spruce. However, temperature alone cannot explain the great contraction in stem diameter due to small expansion of the wood. Shrinkage of this magnitude is possible when living cells are dehydrated and lose turgor (Zweifel and Hasler 2000). Therefore, the sap flow and stem diameter variation followed opposite diurnal patterns in all species. Similar results were reported for Betula pendula, a deciduous species in boreal regions in Finland (Holtta et al. 2018). These authors suggested that this opposite trend may be caused by conversion between substances. Namely, during the day, the osmotic potential in the parenchyma increased due to sugar-to-starch turnover in the parenchyma cells. The increasing osmotic potential resulted in increased sap pressure in the xylem and decreased water uptake from soil. Therefore, sap flow rate slowed down and diameter increased with reduced water transportation into various tissues and organs. During the night, starch was degraded into sugars to protect cells from low temperature, the osmotic potential decreased leading to lower xylem sap pressure, and water uptake from soil increased and water was transported into tissues and organs, resulting in the diameter decreasing. Alternatively, as the temperature was below 0 °C when sap flow occurred in the current study, exposed twigs of the crown periphery might freeze and stem diameter shrank. Xylem water potential would decrease and water transported along an osmotic potential gradient. Temperature increased with sunrise and sap flow declined, implying xylem sap pressure would be relaxing with thawing of frozen soil (Lindfors et al. 2019; Zweifel and Hasler 2000). A water potential gradient during the freeze–thaw cycle may result in daily courses of sap flow and stem diameter changes in cold seasons.
During the non-growing season, although the sap flow was low, it was negatively correlated with T, VPD, and variation in stem diameter on an hourly timescale; the exception is P. orientalis in which sap flow was not significantly correlated with the variation in stem diameter. These relationships may, nevertheless, indicate that some physiological activity is still present. In a broad-leaved evergreen woody vegetation in a seasonally dry region in southwestern Australia, transpiration decreased dramatically with increasing VPD as plants closed their stomata under severe moisture deficit in the non-growing season (Gwenzi et al. 2012). Transpiration in a coniferous evergreen species, Picea engelmannii, in Colorado was generally low during early- to mid-winter, but it rose in mid- to late-winter with increasing solar angle and air temperature (Boyce and Lucero 1999). Boyce et al. (2002) concluded that winter water relations in three evergreen conifers in Vermont were affected by winter temperature and relative humidity. A measurement using eddy-covariance in a subtropical plantation of several evergreen species showed that winter transpiration increased significantly along with ascending VPD, but was not obviously related to canopy conductance, suggesting that transpiration was controlled mainly by environmental factors and weakly influenced by stomatal regulation (Song et al. 2006). Species, climate, time scale, and soil property may thus be responsible for different factors affecting sap flow during the non-growing season.
Uncertainty and expectations about sap flow in non-growing season
Thermal dissipation probes (TDP) developed by Granier (1987), owing to their simplicity, high degree of accuracy, and relatively low cost, are commonly used to monitor tree transpiration (Lu et al. 2004). Most researchers used the TDP method in the growing season (Kunert et al. 2010; Ma et al. 2017; Yan et al. 2018; Zhang et al. 2016). Lower temperature and the risk of freezing damage in non-growing season complicate the use of TDP. In addition, uncertainty exists whether TDP can detect small sap flow in non-growing season (Chan and Bowling 2017; Holtta et al. 2015). Chan and Bowling (2017) conducted laboratory and field experiments to examine the feasibility of using TDP during the cold season, and they discovered that this method can be reliably applied to monitor water transport during cold periods with low sap flow. Therefore, TDP method could be applied to measure sap flow in non-growing season.
In our study, sap flow in non-growing season was detected in both the evergreen species and deciduous species. We focused on the diurnal courses and the daily factors affecting sap flow in non-growing season. Similar results were reported for Betula pendula, a deciduous broadleaved species, and the authors highlighted the reasons why sap flow generates in non-growing season (Holtta et al. 2018). Their results help us to speculate about our results. Where and why the sap flow generates in the semi-arid Loess Plateau region are questions that should be addressed in future research. The observation will help us to better understand winter water relations and cold-resistance of species.
Conclusions
The results revealed differences in peak times in diurnal courses of sap flow, suggesting that the three tested species vary in stomatal sensitivity to atmospheric conditions. P. orientalis and Q. liaotungensis tended to reduce their instantaneous transpiration earlier and did not show significant difference between different soil moisture conditions. In contrast, R. pseudoacacia transpired water actively for longer time during the midday hours, particularly when soil water had been restored by rainfall. Sap flow during the non-growing season was detected in all the species. The ratio of Fd in non-growing season compared with that in the growing season was higher in the evergreen P. orientalis than in the two deciduous species. The sap flow in non-growing season was correlated with T, VPD, and stem diameter variation. Considering the foliage and temperature conditions, species variations and mechanisms in non-growing season sap flow need further investigation.
References
Boyce RL, Lucero SA (1999) Role of roots in winter water relations of Engelmann spruce saplings. Tree Physiol 19:893–898. https://doi.org/10.1093/treephys/19.13.893
Boyce RL, Vostral CB, Friedland AJ (2002) Winter water relations of New England conifers and factors influencing their upper elevational limits II modelling. Tree Physiol 22:801–806. https://doi.org/10.1093/treephys/22.11.801
Breda N, Huc R, Granier A, Dreyer E (2006) Temperate forest trees and stands under severe drought: a review of ecophysiological responses, adaptation processes and long-term consequences. Ann Forest Sci 63:625–644. https://doi.org/10.1051/forest:2006042
Campbell GS, Norman JM (1998) An introduction to environmental biophysics. Springer, New York
Chan AM, Bowling DR (2017) Assessing the thermal dissipation sap flux density method for monitoring cold season water transport in seasonally snow-covered forests. Tree Physiol 37:984–995. https://doi.org/10.1093/treephys/tpx049
Chang XX, Zhao WZ, Zhang ZH, Su YZ (2006) Sap flow and tree conductance of shelter-belt in arid region of China. Agric For Meteorol 138:132–141. https://doi.org/10.1016/j.agrformet.2006.04.003
Chen LX et al (2014) Response of transpiration to rain pulses for two tree species in a semiarid plantation. Int J Biometeorol 58:1569–1581. https://doi.org/10.1007/s00484-013-0761-9
Chen XD, Tang M, Zhang XL, Hamel C, Li W, Sheng M (2018) Why does oriental arborvitae grow better when mixed with black locust: Insight on nutrient cycling? Ecol Evol 8:744–754. https://doi.org/10.1002/ece3.3578
Clearwater MJ, Meinzer FC, Andrade JL, Goldstein G, Holbrook NM (1999) Potential errors in measurement of nonuniform sap flow using heat dissipation probes. Tree Physiol 19:681–687. https://doi.org/10.1093/treephys/19.10.681
Du S, Yamanaka N, Yamamoto F, Otsuki K, Wang SQ, Hou QC (2007) The effect of climate on radial growth of Quercus liaotungensis forest trees in Loess Plateau, China. Dendrochronologia 25:29–36. https://doi.org/10.1016/j.dendro.2007.01.005
Du S, Wang YL, Kume T, Zhang JG, Otsuki K, Yamanaka N, Liu GB (2011) Sapflow characteristics and climatic responses in three forest species in the semiarid Loess Plateau region of China. Agric For Meteorol 151:1–10. https://doi.org/10.1016/j.agrformet.2010.08.011
Fisher JB, Baldocchi DD, Misson L, Dawson TE, Goldstein AH (2007) What the towers don't see at night: nocturnal sap flow in trees and shrubs at two AmeriFlux sites in California. Tree Physiol 27:597–610. https://doi.org/10.1093/treephys/27.4.597
Granier A (1987) Evaluation of transpiration in a Douglas-fir stand by means of sap flow measurements. Tree Physiol 3:309–320. https://doi.org/10.1093/treephys/3.4.309
Granier A, Huc R, Barigah ST (1996) Transpiration of natural rain forest and its dependence on climatic factors. Agric For Meteorol 78:19–29. https://doi.org/10.1016/0168-1923(95)02252-x
Gwenzi W, Veneklaas EJ, Bleby TM, Yunusa IAM, Hinz C (2012) Transpiration and plant water relations of evergreen woody vegetation on a recently constructed artificial ecosystem under seasonally dry conditions in Western Australia. Hydrol Process 26:3281–3292. https://doi.org/10.1002/hyp.8330
He Q-Y et al (2020) Sap flow changes and climatic responses over multiple-year treatment of rainfall exclusion in a sub-humid black locust plantation. For Ecol Manag 457:117730. https://doi.org/10.1016/j.foreco.2019.117730
Holtta T, Linkosalo T, Riikonen A, Sevanto S, Nikinmaa E (2015) An analysis of Granier sap flow method, its sensitivity to heat storage and a new approach to improve its time dynamics. Agric For Meteorol 211:2–12. https://doi.org/10.1016/j.agrformet.2015.05.005
Holtta T, Carrasco M, Salmon Y, Aalto J, Vanhatalo A, Back J, Lintunen A (2018) Water relations in silver birch during springtime: How is sap pressurised? Plant Biol 20:834–847. https://doi.org/10.1111/plb.12838
James SA, Clearwater MJ, Meinzer FC, Goldstein G (2002) Heat dissipation sensors of variable length for the measurement of sap flow in trees with deep sapwood. Tree Physiol 22:277–283. https://doi.org/10.1093/treephys/22.4.277
Jimenez MS, Cermak J, Kucera J, Morales D (1996) Laurel forests in Tenerife, Canary Islands: the annual course of sap flow in Laurus trees and stand. J Hydrol 183:307–321. https://doi.org/10.1016/0022-1694(95)02952-4
Jimenez MS, Morales D, Kucera J, Cermak J (1999) The annual course of transpiration in a laurel forest of Tenerife: estimation with Myrica faya. Phyton-Ann Rei Bot A 39:85–90
Kocher P, Gebauer T, Horna V, Leuschner C (2009) Leaf water status and stem xylem flux in relation to soil drought in five temperate broad-leaved tree species with contrasting water use strategies. Ann Forest Sci 66:101. https://doi.org/10.1051/forest/2008076
Kunert N, Schwendenmann L, Holscher D (2010) Seasonal dynamics of tree sap flux and water use in nine species in Panamanian forest plantations. Agric For Meteorol 150:411–419. https://doi.org/10.1016/j.agrformet.2010.01.006
Lindfors L, Atherton J, Riikonen A, Holtta T (2019) A mechanistic model of winter stem diameter dynamics reveals the time constant of diameter changes and the elastic modulus across tissues and species. Agric For Meteorol 272:20–29. https://doi.org/10.1016/j.agrformet.2019.03.016
Liu ZQ, Yu XX, Jia GD (2019) Water uptake by coniferous and broad-leaved forest in a rocky mountainous area of northern China. Agric For Meteorol 265:381–389. https://doi.org/10.1016/j.agrformet.2018.11.036
Llorens P, Poyatos R, Latron J, Delgado J, Oliveras I, Gallart F (2010) A multi-year study of rainfall and soil water controls on Scots pine transpiration under Mediterranean mountain conditions. Hydrol Process 24:3053–3064. https://doi.org/10.1002/hyp.7720
Lu P, Urban L, Zhao P (2004) Granier's thermal dissipation probe (TDP) method for measuring sap flow in trees: Theory and practice. Acta Bot Sin 46:631–646. https://doi.org/10.3321/j.issn:1672-9072.2004.06.001
Lu WW, Yu XX, Jia GD (2019) Instantaneous and long-term CO2 assimilation of Platycladus orientalis estimated from 13C discrimination. Ecol Indic 104:237–247. https://doi.org/10.1016/j.ecolind.2019.05.007
Ma C, Luo Y, Shao M, Li X, Sun L, Jia X (2017) Environmental controls on sap flow in black locust forest in Loess Plateau. China Sci Rep 7:13160. https://doi.org/10.1038/s41598-017-13532-8
Mantovani D, Veste M, Freese D (2014) Black locust (Robinia pseudoacacia L.) ecophysiological and morphological adaptations to drought and their consequence on biomass production and water-use efficiency. NZ J For Sci 44:29. https://doi.org/10.1186/s40490-014-0029-0
Mantovani D, Veste M, Boehm C, Vignudelli M, Freese D (2015) Spatial and temporal variation of drought impact on black locust (Robinia pseudoacacia L.) water status and growth. iForest 8:743–747. https://doi.org/10.3832/ifor1299-008
Matisons R, Bardulis A, Kanberga-Silina K, Krisans O, Jansons A (2017) Sap flow in xylem of mature Norway spruce: a case study in Northwestern Latvia during the season of 2014–2015. Balt For 23:477–481
Nadal-Sala D, Sabate S, Sanchez-Costa E, Poblador S, Sabater F, Gracia C (2017) Growth and water use performance of four co-occurring riparian tree species in a Mediterranean riparian forest. For Ecol Manage 396:132–142. https://doi.org/10.1016/j.foreco.2017.04.021
Nadezhdina N et al (2010) Trees never rest: the multiple facets of hydraulic redistribution. Ecohydrology 3:431–444. https://doi.org/10.1002/eco.148
Niinemets U, Flexas J, Penuelas J (2011) Evergreens favored by higher responsiveness to increased CO2. Trends Ecol Evol 26:136–142. https://doi.org/10.1016/j.tree.2010.12.012
Oren R, Pataki DE (2001) Transpiration in response to variation in microclimate and soil moisture in Southeastern deciduous forests. Oecologia 127:549–559. https://doi.org/10.1007/s004420000622
Shi WY, Yan MJ, Zhang JG, Guan JH, Du S (2014) Soil CO2 emissions from five different types of land use on the semiarid Loess Plateau of China, with emphasis on the contribution of winter soil respiration. Atmos Environ 88:74–82. https://doi.org/10.1016/j.atmosenv.2014.01.066
Siegert CM, Levia DF (2011) Stomatal conductance and transpiration of co-occurring seedlings with varying shade tolerance. Trees-Struct Funct 25:1091–1102. https://doi.org/10.1007/s00468-011-0584-4
Song X, Yu GR, Liu YF, Sun XM, Lin YM, Wen XF (2006) Seasonal variations and environmental control of water use efficiency in subtropical plantation. Sci China Ser D 49:119–126. https://doi.org/10.1007/s11430-006-8319-X
Tian Q, Taniguchi T, Shi W-Y, Li G, Yamanaka N, Du S (2017) Land-use types and soil chemical properties influence soil microbial communities in the semiarid Loess Plateau region in China. Sci Rep 7:45289. https://doi.org/10.1038/srep45289
Totzke C, Cermak J, Nadezhdina N, Tributsch H (2017) Electrochemical in-situ studies of solar mediated oxygen transport and turnover dynamics in a tree trunk of Tilia cordata. iForest 10:355–361. https://doi.org/10.3832/ifor1681-010
Tsunekawa A, Liu GB, Yamanaka N, Du S (2014) Restoration and development of the degraded loess plateau. Springer, Tokyo, China. https://doi.org/10.1007/978-4-431-54481-4
Wang L, Dai YX, Sun JZ, Wan XC (2017) Differential hydric deficit responses of Robinia pseudoacacia and Platycladus orientalis in pure and mixed stands in northern China and the species interactions under drought. Trees-Struct Funct 31:2011–2021. https://doi.org/10.1007/s00468-017-1605-8
Yan CH, Wang B, Zhang Y, Zhang XN, Takeuchi S, Qiu GY (2018) Responses of sap flow of deciduous and conifer trees to soil drying in a subalpine forest. Forests 9:32. https://doi.org/10.3390/f9010032
Yan MJ, Yamamoto M, Yamanaka N, Yamamoto F, Liu GB, Du S (2013) A comparison of pressure-volume curves with and without rehydration pre-treatment in eight woody species of the semiarid Loess Plateau. Acta Physiol Plant 35:1051–1060. https://doi.org/10.1007/s11738-012-1143-3
Yan MJ, Yamanaka N, Yamamoto F, Du S (2010) Responses of leaf gas exchange, water relations, and water consumption in seedlings of four semiarid tree species to soil drying. Acta Physiol Plant 32:183–189. https://doi.org/10.1007/s11738-009-0397-x
Yan MJ, Zhang JG, He QY, Shi WY, Otsuki K, Yamanaka N, Du S (2016) Sapflow-based stand transpiration in a semiarid natural oak forest on China's Loess Plateau. Forests 7:227. https://doi.org/10.3390/f7100227
Yoshifuji N, Komatsu H, Kumagai T, Tanaka N, Tantasirin C, Suzuki M (2011) Interannual variation in transpiration onset and its predictive indicator for a tropical deciduous forest in northern Thailand based on 8-year sap-flow records. Ecohydrology 4:225–235. https://doi.org/10.1002/eco.219
Zhang JG, Guan JH, Shi WY, Yamanaka N, Du S (2015) Interannual variation in stand transpiration estimated by sap flow measurement in a semi-arid black locust plantation, Loess Plateau, China. Ecohydrology 8:137–147. https://doi.org/10.1002/eco.1495
Zhang ZZ et al (2016) Influence of the decoupling degree on the estimation of canopy stomatal conductance for two broadleaf tree species. Agric For Meteorol 221:230–241. https://doi.org/10.1016/j.agrformet.2016.02.018
Zweifel R, Hasler R (2000) Frost-induced reversible shrinkage of bark of mature subalpine conifers. Agric For Meteorol 102:213–222. https://doi.org/10.1016/s0168-1923(00)00135-0
Acknowledgments
This research was supported by the National Key R&D Program of China (Grant Number 2017YFC0504601) and the National Natural Science Foundation of China (Grant Numbers 41171419, 41471440, 41411140035).
Author information
Authors and Affiliations
Contributions
JL and SD conceived the research. JL, QYH, JY, QWC and RRC conducted the field investigations. JL analyzed the data and wrote the manuscript. All members commented on the data analyses and reviewed the manuscript.
Corresponding author
Additional information
Communicated by Ishii.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Lyu, J., He, QY., Yang, J. et al. Sap flow characteristics in growing and non-growing seasons in three tree species in the semiarid Loess Plateau region of China. Trees 34, 943–955 (2020). https://doi.org/10.1007/s00468-020-01972-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00468-020-01972-1