
Abstract
Pressure–volume (P–V) curves are frequently used to analyze water relation properties of woody plants in response to transpiration-induced tissue water loss. In this study, P–V analyses were conducted on eight woody species growing in the semiarid Loess Plateau region of China during a relatively dry summer season using both the recently recommended instantaneous measurement and the traditional method with rehydration pretreatment. Generally, P–V-derived parameters in this study reflected conditions in a dry growth environment. Species-specific differences were also found among P–V parameters, suggesting each species uses different mechanisms to respond to drought. Based on the results from instantaneous measurements, a descending sequence for drought tolerance ranked by water potentials at the turgor loss point (Ψtlp) was Rosa hugonis > Syringa oblata = Armeniaca sibirica > Caragana microphylla > Pyrus betulaefolia > Acer stenolobum > Quercus liaotungensis > Robinia pseudoacacia. The first five species also showed lower levels of osmotic potential at full turgor (Ψ satπ ) and higher symplastic osmotic solute content per dry weight, suggesting they possess advantages in osmotic adjustment. Also, this study supports previous reports noting rehydration pretreatment resulted in shifts in P–V parameters. The magnitude of the shifts varied with species and water conditions. The effect of rehydration was stronger for species with higher drought tolerance or subjected to the influence of drought. Differences in the parameters among species were mitigated as a result of rehydration. Those with a lower Ψtlp or midday water potential were more deeply affected by rehydration. Application of instantaneous measurements was strongly recommended for proper analysis of P–V curves particularly in arid and semiarid areas.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Ci and Wu (1997) estimated that more than a third of China was composed of arid and semiarid lands. Vegetation in these areas is subjected to frequent stress from soil water shortages and the dry climate. In the semiarid Loess Plateau region of north-central China, easily eroded soils, aridity, and human activities have caused severe soil erosion and vegetation degradation. Rehabilitating and stabilizing these ecosystems is a major task related to forest and grassland management in the region.
Reforestation of semiarid land requires using selected species that can provide sustainable long-term ecological services. Eco-physiological properties of trees are commonly considered when their capacity for growth and stress tolerance are evaluated (Kozlowski and Pallardy 1997; Larcher 2003). Several characteristics have been addressed as being potentially responsible for the adaptability of tree species to dryland environments. These characteristics include, for example, the xerophytic structure of leaves, well-developed root systems, stems with high hydraulic conductivity and low cavitation risk, the ability to endure extremely low water potentials, stomatal control of transpiration, osmotic adjustment, activation of anti-oxidative enzymes, and possession of stiff cell walls (Kozlowski 1976; Kozlowski and Pallardy 1997; Larcher 2003). Several investigators have studied forest species growing in the central Loess Plateau region regarding their photosynthetic and water consumption responses to drought stress (Hara et al. 2008; Yan et al. 2010; Du et al. 2011). However, the investigation of water relation characteristics for in situ individuals is still very limited.
Pressure–volume (P–V) curves, which are plotted using measurements of leaf water potential (Ψ) and relative water content (RWC), have been widely applied as an important approach in the evaluation of drought tolerance and drought response patterns for trees and shrubs. Several parameters derived from a P–V curve may indicate a species ability to maintain cell turgor pressure under drying stress, or the drought tolerance of a species. Generally, lower water potential at the turgor loss point (Ψtlp) is highly correlated with lower osmotic potential at water saturation with full turgor (Ψ satπ ). This means the species has a strong ability for osmotic adjustment by having a large amount of osmotic material in the cell sap. As drought may stimulate synthesis and accumulation of osmotically effective constituents in some species, a reduction in Ψtlp may occur. Bulk elastic modulus (ε) reflects the rigidity of cell walls. Low ε helps tissues maintain their turgor during the process of water loss, while high ε means Ψ drops rapidly during water loss and the plant reaches Ψtlp quickly, which may help the plant absorb water during the early stages of water loss.
Two methods have been applied to the construction of P–V curves. The traditional method includes a rehydration pretreatment of plant materials (branches with leaves) lasting from several to more than 24 h before the measurements of leaf or xylem water potentials (Scholander et al. 1965; Tyree and Hammel 1972). Thus, the measurements are considered to start when the plant has full turgor and cease after turgor loss. Since rehydration pretreatment may lead to excessive apoplastic water uptake, high estimates of osmotic potential (Ψπ) and low estimates of relative water content at turgor loss point (RWCtlp) and ε have been detected (Evans et al. 1990; Meinzer et al. 1986). P–V parameters for rehydrated leaves shifted significantly for plants under soil moisture stress (Kubiske and Abrams 1991a, b). Seasonal variations and stress response patterns may not be reflected by the results in such cases, probably depending on the species involved, site conditions, and rehydration time (Kubiske and Abrams 1991b). Measurements with non-rehydrated samples have been applied to the P–V analyses in recent years. To obtain the changes in Ψ components over a wide range of RWC, researchers tend to start sampling and taking measurements in the predawn hours when the tissues may have been naturally rehydrated. However, shifts in P–V parameters have also been detected even in naturally rehydrated leaves and shoots (Parker and Pallardy 1987).
In the present study, we aimed (1) to comparatively evaluate the water relation characteristics based on P–V parameters for the common woody species in the region, and (2) to assess the shifts (if any) in P–V parameters induced during rehydration pretreatment. The results would contribute to both a broader understanding of drought response strategies for the species involved and proper application of P–V techniques for plants in this semiarid environment.
Materials and methods
Study site and plant materials
The study was carried out at Mount Gonglushan (36°25.40′ N, 109°31.53′ E; 1353 m a.s.l.) near the city of Yan’an, Shaanxi Province, in the semiarid Loess Plateau of central China. This area is located on the forest-grassland transition zone, where the forest ecosystems are subject to drought. The mean annual precipitation and air temperature for 1982–2003 were 514 mm and 10.2 °C, respectively, at a meteorological station in nearby Yan’an. Spring and early summer (from March to June) are usually dry in the area. July and August are hot during the summer season but with considerable precipitation. Otsuki et al. (2005) and Du et al. (2007) provide detailed descriptions of the study site. Measurements for this study were taken on relatively clear days from 17 to 23 in August 2003. Rainfall occurred scarcely during the 2 weeks before the measurements (Fig. 1).

Daily mean (solid line with filled diamond), maximum (broken line with open diamond), and minimum (dotted line with multi symbol) air temperature and daily precipitation (solid bars) in July and August in 2003 at Mt. Gonglushan. Meteorological instruments were set on a nearby open space. Field measurements for water relations were carried out during the 7 days (17–23 August) indicated by the shaded area
Plant materials include six deciduous tree species, i.e., Robinia pseudoacacia L. (black locust), Quercus liaotungensis Koidz. (Liaodong oak), Syringa oblata Lindl. (early lilac), Acer stenolobum Rehd. (narrow crack maple), Armeniaca sibirica Lam. (wild apricot), Pyrus betulaefolia Bunge (birch-leaf pear), and two deciduous shrubs, i.e., Caragana microphylla Lam. (littleleaf peashrub) and Rosa hugonis Hemsl. (Father Hugo rose). R. pseudoacacia trees were planted for reforestation on abandoned farmland 20 years ago on an east-facing slope. Q. liaotungensis trees were from a naturally developed stand about 50 years old and facing northeast. Others were from the nearby natural tree-shrub mixed vegetation around the ridge. Five sample trees of relatively similar size were selected for each species growing within 100 m from each other. Table 1 provides mean heights and diameters at breast height of the sample plants.
P–V curve analysis
Measurements for P–V analyses were performed using a pressure chamber (Model 1000, PMS instruments Corvallis, OR) during 17–20 August 2003, by periodically measuring sample weight and Ψ following the method described in previous studies (Tyree and Hammel 1972; Kubiske and Abrams 1990). Both rehydrated and non-rehydrated leaves were measured for each sample tree for comparison.
Sampling was conducted during predawn hours (around 4:00 h). Two terminal shoots with young, fully expanded leaves were collected from each of five individuals for each species. They were cut at 2–4 m height in the tall trees and at 1.0–1.5 m height in the two shrubs. The shoots of approximately 30–50 cm length were placed into a sealed lightproof plastic bag with a moist paper towel and carried to a nearby temporary laboratory. Each shoot for rehydration pretreatment was recut under tap water and the base hydrated in the dark for about 24 h before measurements. Another shoot from the same individual was used for instantaneous measurement in P–V analysis. An apical twig was cut from each sample shoot for periodical measurements.
Ψtlp, Ψ satπ , RWCtlp, relative symplastic water content at turgor loss point (SWCtlp), relative symplastic water content (V0), and relative apoplastic water content (Va) were derived from P–V curves based on the work of Tyree and Hammel (1972) and Kubiske and Abrams (1990, 1991b). Symplastic water at full turgor per dry weight (V0DW−1) and symplastic osmotic solute content per dry weight (NsDW−1) were also estimated (Maury et al. 2000; Tognetti et al. 2000). Maximum bulk elastic modulus (εmax) was calculated following Saito and Terashima (2004).
The saturated weight that is needed to calculate RWC was determined using linear regression on data above the turgor loss point in a plot of fresh weight versus water potential. Because several P–V curves of the rehydrated samples in some species contained a plateau region, the linear regression excluded this region to avoid the “plateau effect,” rehydration-induced shift in P–V curve parameters, following Kubiske and Abrams (1990, 1991a) and Dichio et al. (2003).
In situ measurements of water potentials
Water potentials at predawn (Ψpd) and midday (Ψmd) were measured in situ at about 0400 and 1300 hours, respectively, for each of the sampling individuals on 23 August 2003. Sampling was at the same height on the individuals as that for P–V curve analysis. The measurement was performed immediately after cutting.
Statistical analysis
Statistical analyses were performed using analysis of variance. Water potentials and P–V parameters were compared among species using Tukey’s honestly significant difference test. For P–V parameters, paired t tests were used to test the differences between rehydration treatment and non-rehydration. Differences between Ψmd and Ψtlp were also checked by paired t tests. Correlations among P–V parameters and between P–V parameters and Ψmd and Ψtlp were examined using Pearson (r) coefficient, considering the individual values of all sample trees for either the rehydrated or non-rehydrated method (n = 40). Analyses were performed using R software for Windows (R Foundation for Statistical Computing, Vienna, Austria).
Results and discussion
Predawn and midday water potentials
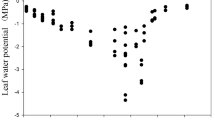
Species differences in Ψpd and Ψmd were observed (Fig. 2). R. pseudoacacia and Q. liaotungensis showed higher Ψpd and Ψmd than other species, though the differences with some species were not statistically significant. All the measured individuals for these two species recovered their Ψ to above −1.0 MPa by the predawn measurement time. Also, A. stenolobum, P. betulaefolia and C. microphylla also recovered relatively well over the night. On the other hand, S. oblata, A. sibirica and R. hugonis showed low values of both Ψpd and Ψmd, suggesting a poor recovery of leaf water status throughout, while C. microphylla showed a relatively high Ψpd but the Ψmd dropped to a much lower level.

Water potential at predawn and midday of the eight woody species growing at Mt. Gonglushan. Mean values and standard errors (n = 5) are presented for each species. Bars with the same letters are not significantly different among species (Tukey’s honestly significant difference test, P < 0.05)
These two parameters might reflect several water-related factors. High values of Ψpd and Ψmd are expected with sufficient soil moisture, efficient water uptake and high hydrological conductivities of the plants. Soil moisture conditions may be reflected by Ψpd very well. Water uptake by root systems of S. oblata, A. sibirica and R. hugonis was not able to compensate for their transpiration loss. Also, Ψmd may be more species-specific and also reflect the condition of soil moisture stress. In general, Ψmd values in the present study were much lower than those found in studies on plants growing at humid areas, suggesting these ecosystems were water-limited. S. oblata, A. sibirica, R. hugonis and C. microphylla in particular have developed drought resistance.
P–V parameters and rehydration effects
Figure 3 shows typical Höfler diagrams obtained from P–V curves for the eight species, including instant measurements and rehydration pretreatment. These diagrams represent dynamic changes of Ψ, Ψπ, and Ψp in relation to relative symplastic water content (SWC). Species differences in these P–V parameters can be found in the diagrams. Höfler diagrams particularly revealed differences between instant measurements and rehydration pretreatment. The plotted lines in the diagrams varied widely from one to another in S. oblata, A. sibirica, C. microphylla, and R. hugonis.

Typical Höfler diagrams, relating water potential (Ψ; filled diamond, open diamond), osmotic potential (Ψπ; filled triangle, open triangle), and pressure potential (Ψp; filled circle, open circle), to relative symplastic water contents from pressure–volume curves of the eight species using non-rehydrated (NR; solid lines with closed symbols) and rehydrated (R; broken lines with opened symbols) materials. Each diagram includes a pair of curves as a representative from one of the five replicated sampling individuals
R. pseudoacacia, Q. liaotungensis and A. stenolobum showed higher Ψtlp and Ψ satπ and lower NsDW−1 than the other species (Table 2). Other P–V parameters did not rank identically among species. Rehydration-induced shifts in P–V parameters occurred in most of the species regardless of correction for the “plateau effect.” Compared with instantaneous measurement, rehydration masked differences in several parameters (e.g., Ψtlp, Ψ satπ , and NsDW−1) among species. Rehydration resulted in overestimates of Ψtlp in every species. The shifts were significant in S. oblata, A. sibirica, P. betulaefolia, C. microphylla, and R. hugonis (P < 0.05, with asterisks). Ψ satπ was also significantly overestimated by rehydration in several species (S. oblata, A. sibirica, and R. hugonis). NsDW−1 was underestimated by rehydration for all species excluding R. pseudoacacia and Q. liaotungensis, and the underestimation was significant in S. oblata and R. hugonis. Other parameters showed relatively small differences between rehydration and non-rehydration and among species.
The results of this study support previous reports where rehydration resulted in shifts in P–V parameters, even after the correction of the “plateau” effects. Also, rehydration-induced shifts masked seasonal variations in P–V parameters that could be detected using instantaneous measurements (Kubiske and Abrams 1991b). In addition, the effect of rehydration was stronger for species with higher drought tolerance or subjected to the influence of a droughty environment (Evans et al. 1990; Kubiske and Abrams 1991a). The magnitude of the shifts in the present study also varied with species. Differences in the parameters among species were mitigated consequently. Those with lower Ψtlp and Ψmd, i.e., S. oblata, A. sibirica, C. microphylla, and R. hugonis, showed a stronger influence by the rehydration pretreatment.
In practice, full turgor does not occur under common weather conditions (non-rainy) in arid and semiarid areas. Artificial saturation in the laboratory may result in infiltration of water into air spaces, causing unreasonable level of RWC. Even re-watering the pot soil of water-stress conditioned poplars had caused substantial reduction in concentrations of some major solutes (Gebre et al. 1997). As the rehydration-induced shifts might result from the “plateau” effects of excessive Va, correction of the “plateau” has been applied to P–V analyses (Dichio et al. 2003; Patakas et al. 2002; Warren et al. 2007). However, such a correction turned out to be insufficient to eliminate the influence. They may also be caused by the loss of symplastic solutes (Kubiske and Abrams 1991b). Lower estimation of NsDW−1 and higher estimation of Va were also observed in the rehydration treatments of the present study, though results were statistically insignificant in many species. For the materials in arid and semiarid areas, or those subjected to water stress, osmotic substances would accumulate and play important roles in the water relation properties (Fernandez et al. 1999). In this case, rehydration-caused solute loss would particularly mask the real nature of the materials.
Relationships among P–V parameters
Correlation analyses for the parameters support the general understanding that low Ψtlp is strongly associated with low Ψ satπ which is affected by high NsDW−1, and high NsDW−1 is related to high V0DW−1 and low Va (Table 3). SWCtlp and εmax showed positive correlations. However, several correlations that are significant for parameters of non-rehydration became less significant or non-significant in data with rehydration pretreatment. In contrast, a positive relationship between Ψtlp and RWCtlp was only seen to be significant using the data with rehydration pretreatment.
There have been investigations on the synthesis of various osmotic substances in response to drought and their roles in Ψ satπ (Clifford et al. 1998; Guignard et al. 2005; Patakas et al. 2002; Warren et al. 2007). Increased NsDW−1 was found to be accompanied by an increase in V0 and decreases in Va and Ψ satπ in oak leaves, suggesting a role of osmotic solutes in the water distribution between inside and outside the cells (Aranda et al. 2001). Also, a seasonally observed increase in osmotic solutes was also found to be accompanied by a redistribution of apoplastic to symplastic water (Miki et al. 2003).
The rigidity of cell walls may be related to cell size and wall composition and structure, and so affects a cell’s tolerance to desiccation. Negative relationships were observed in previous reports and this study between ε and both Ψtlp and Ψ satπ (Corcuera et al. 2002; Kubiske and Abrams 1994; Ngugi et al. 2003; White et al. 2000). Rehydration treatment may not only have affected Va, NsDW−1, and RWCtlp, but also affected ε.
Relationships between P–V parameters and water potentials
Significant correlations were also observed for several P–V parameters of non-rehydration with respect to Ψpd and Ψmd, but not for parameters derived from rehydration (Table 4). Both Ψpd and Ψmd were positively correlated with Ψtlp and Ψ satπ , and negatively correlated with NsDW−1 and V0DW−1. This suggests P–V parameters substantially coincide with the in situ water status of the species and ecotypes. As Ψmd is likely to be a reasonable representative of minimum value throughout a day, the relationship between Ψmd and Ψtlp was also analyzed (Fig. 4). For most of the species, contrary to expectation, in situ measured Ψmd was even lower than Ψtlp from P–V analysis. Ψmd was significantly lower than the value of Ψtlp with rehydration pretreatment for most of the species (Fig. 4b). These results also agree with the shift of estimated parameters from the measurements after rehydration, especially the up-estimation for Ψtlp. However, for non-rehydration, Ψmd and Ψtlp values were not significantly different within individual species except for R. pseudoacacia and S. oblata (Fig. 4a).

Relationships between midday water potential and water potential at turgor loss point (Ψtlp) obtained from the pressure–volume curves using non-rehydrated (a) and rehydrated (b) materials. Data sets are presented as means and standard errors (n = 5) for each species. The dashed line indicates Y = X. Asterisks indicate significant difference between Midday Ψ and Ψtlp (paired t test, P < 0.05)
The present study was conducted during a period of relative drought. The soil moisture had barely recharged by effective rainfall over several weeks, while the evaporative demands were much higher than the rainfall’s recharge (Fig. 1). The measured lower values of Ψmd suggest these plants were subjected to drought. Species that have adapted such environments may have strong ability of osmotic adjustment in response to leaf water loss. R. pseudoacacia and S. oblata, which showed large differences between Ψmd and estimated Ψtlp, might have more actively transpired during the daytime and thus suffer water deficiency in their leaves. Some leaves in S. oblata were observed to be withering during the midday measurement period.
Conclusion
This study was conducted on a site of semiarid area during a drought period. Water relation characteristics of the six tree species and two shrub species reflected their dry growth environment. Ψtlp and Ψ satπ for all the species were lower than those for plants growing at humid areas (e.g., Hara et al. 2008; Kubiske and Abrams 1994; Saito et al. 2003). Species-specific differences were also found among P–V parameters, suggesting different species used different drought response mechanisms. Based on the results from non-rehydrated measurements, a descending sequence for drought tolerance ranked by Ψtlp was R. hugonis > S. oblata = A. sibirica > C. microphylla > P. betulaefolia > A. stenolobum > Q. liaotungensis > R. pseudoacacia. The former five species showed Ψtlp below −3 MPa, indicating their tolerance to severe water stress. These species also showed lower levels of Ψ satπ and higher contents of NsDW−1, suggesting their advantages in osmotic adjustment. From the viewpoint of cell wall rigidity, S. oblata, R. hugonis, A. sibirica and Q. liaotungensis shared the characteristics of high εmax, implying that their Ψ would rapidly decrease and the stoma would close at the early stage of water deficiency. With relatively higher Ψtlp and Ψ satπ , and lower εmax, R. pseudoacacia would continuously transpire during droughts and the cells tend to lose their turgor earlier.
This study supports previous reports that rehydration pretreatment resulted in shifts in P–V parameters. The magnitude of the shifts varied with species and water conditions. The effect of rehydration was stronger for species with higher drought tolerance or subjected to the influence of drought environment. Differences in the parameters among species were mitigated as a result of rehydration. Those with lower Ψtlp and Ψmd, i.e., S. oblata, A. sibirica, C. microphylla, and R. hugonis, showed higher influence by the rehydration treatment. Application of instantaneous measurements was strongly recommended for proper analysis of P–V curves particularly in arid and semiarid areas.
Author contribution
Mei-Jie Yan and Makiko Yamamoto prepared the manuscript. Fukuju Yamamoto, Norikazu Yamanaka and Sheng Du performed the field measurements. Guo-Bin Liu and Sheng Du edited the manuscript.
Abbreviations
- RWC:
-
Relative water content
- RWCtlp :
-
Relative water content at turgor loss point
- SWC:
-
Relative symplastic water content
- SWCtlp :
-
Relative symplastic water content at turgor loss point
- NsDW−1 :
-
Symplast osmotic solute content per dry weight
- V0DW−1 :
-
Symplastic water at full turgor per dry weight
- V0 :
-
Relative symplastic water content
- Va :
-
Relative apoplastic water content
- Ψ:
-
Water potential
- Ψpd :
-
Water potential at predawn
- Ψmd :
-
Water potential at midday
- Ψtlp :
-
Water potential at turgor loss point
- Ψp :
-
Pressure potential
- Ψπ :
-
Osmotic potential
- Ψ satπ :
-
Osmotic potential at water saturation with full turgor
- ε:
-
Bulk elastic modulus
- εmax :
-
Maximum bulk elastic modulus
References
Aranda I, Gil L, Pardos JA (2001) Effects of thinning in a Pinus sylvestris L. stand on foliar water relations of Fagus sylvatica L. seedlings, planted within the pinewood. Trees-Struct Funct 15(6):358–364
Ci LJ, Wu B (1997) Climatic type division and the potential extent determination of desertification in China. J Desert Res 17(2):107–111 (in Chinese)
Clifford SC, Arndt SK, Corlett JE, Joshi S, Sankhla N, Popp M, Jones HG (1998) The role of solute accumulation, osmotic adjustment and changes in cell wall elasticity in drought tolerance in Ziziphus mauritiana (Lamk). J Exp Bot 49(323):967–977. doi:10.1093/jexbot/49.323.967
Corcuera L, Camarero JJ, Gil-Pelegrin E (2002) Functional groups in Quercus species derived from the analysis of pressure–volume curves. Trees-Struct Funct 16(7):465–472. doi:10.1007/s00468-002-0187-1
Dichio B, Xiloyannis C, Angelopoulos K, Nuzzo V, Bufo SA, Celano G (2003) Drought-induced variations of water relations parameters in Olea europaea. Plant Soil 257(2):381–389
Du S, Yamanaka N, Yamamoto F, Otsuki K, Wang SQ, Hou QC (2007) The effect of climate on radial growth of Quercus liaotungensis forest trees in Loess Plateau, China. Dendrochronologia 25(1):29–36. doi:10.1016/j.dendro.2007.01.005
Du S, Wang Y-L, Kume T, Zhang J-G, Otsuki K, Yamanaka N, Liu G-B (2011) Sapflow characteristics and climatic responses in three forest species in the semiarid Loess Plateau region of China. Agric For Meteorol 151(1):1–10. doi:10.1016/j.agrformet.2010.08.011
Evans RD, Black RA, Link SO (1990) Rehydration-induced changes in pressure–volume relationships of Artemisia tridentata Nutt. ssp. tridentata. Plant Cell Environ 13(5):455–461
Fernandez M, Gil L, Pardos JA (1999) Response of Pinus pinaster Ait. provenances at early age to water supply. I. Water relation parameters. Annals For Sci 56(2):179–187. doi:10.1051/forest:19990209
Gebre GM, Brandle JR, Kuhns MR (1997) Influence of rewatering and time of sampling on solute accumulation of two Populus deltoides clones. Tree Physiol 17(5):341–346
Guignard C, Jouve L, Bogeat-Triboulot MB, Dreyer E, Hausman JF, Hoffmann L (2005) Analysis of carbohydrates in plants chromatography coupled with by high-performance anion-exchange electrospray mass spectrometry. J Chromatogr A 1085(1):137–142. doi:10.1016/j.chroma.2005.05.068
Hara Y, Zhang W, Du S, Tamai S, Yamanaka N (2008) Water relations of 4 afforestation species in the Loess Plateau, China. J Japanese For Soc 90(4):247–252. doi:10.4005/jjfs.90.247 (in Japanese)
Kozlowski TT (ed) (1976) Soil water measurement, plant responses, and breeding for drought resistance. In: Water deficits and plant growth, vol IV. Academic Press, New York
Kozlowski TT, Pallardy SG (1997) Physiology of woody plants, 2nd edn. Academic Press, San Diego
Kubiske ME, Abrams MD (1990) Pressure–volume relationships in non-rehydrated tissue at various water deficits. Plant Cell Environ 13(9):995–1000
Kubiske ME, Abrams MD (1991a) Rehydration effects on pressure–volume relationships in four temperate woody species: variability with site, time of season and drought conditions. Oecologia 85(4):537–542
Kubiske ME, Abrams MD (1991b) Seasonal, diurnal and rehydration-induced variation of pressure–volume relationships in Pseudotsuga menziesii. Physiol Plant 83(1):107–116
Kubiske ME, Abrams MD (1994) Ecophysiological analysis of woody species in contrasting temperate communities during wet and dry years. Oecologia 98(3–4):303–312. doi:10.1007/bf00324218
Larcher W (2003) Physiological Plant Ecology (trans: Huber-Sannwald E), 4th edn. Springer, Berlin
Maury P, Berger M, Mojayad F, Planchon C (2000) Leaf water characteristics and drought acclimation in sunflower genotypes. Plant Soil 223(1–2):153–160
Meinzer FC, Rundel PW, Sharifi MR, Nilsen ET (1986) Turgor and osmotic relations of the desert shrub Larrea tridentata. Plant Cell Environ 9(6):467–475
Miki N, Osuki K, Nishimoto T, Ken Y (2003) Leaf water relations in Pinus densiflora Sieb. et Zucc. on different soil moisture conditions. J For Res 8(3):153–161
Ngugi MR, Doley D, Hunt MA, Dart P, Ryan P (2003) Leaf water relations of Eucalyptus cloeziana and Eucalyptus argophloia in response to water deficit. Tree Physiol 23(5):335–343
Otsuki K, Yamanaka N, Du S, Yamamoto F, Xue ZD, Wang SQ, Hou QC (2005) Seasonal changes of forest ecosystem in an artificial forest of Robinia pseudoacacia in the Loess Plateau in China. J Agric Meteorol 60(5):613–616
Parker WC, Pallardy SG (1987) The influence of resaturation method and tissue type on pressure–volume analysis of Quercus alba L seedlings. J Exp Bot 38(188):535–549
Patakas A, Nikolaou N, Zioziou E, Radoglou K, Noitsakis B (2002) The role of organic solute and ion accumulation in osmotic adjustment in drought-stressed grapevines. Plant Sci 163(2):361–367. doi:10.1016/s0168-9452(02)00140-1
Saito T, Terashima I (2004) Reversible decreases in the bulk elastic modulus of mature leaves of deciduous Quercus species subjected to two drought treatments. Plant Cell Environ 27(7):863–875
Saito T, Tanaka T, Tanabe H, Matsumoto Y, Morikawa Y (2003) Variations in transpiration rate and leaf cell turgor maintenance in saplings of deciduous broad-leaved tree species common in cool temperate forests in Japan. Tree Physiol 23(1):59–66
Scholander PF, Hammel HT, Bradstre ED, Hemmings EA (1965) Sap pressure in vascular plants—negative hydrostatic pressure can be measured in plants. Science 148(3668):339–346
Tognetti R, Raschi A, Jones MB (2000) Seasonal patterns of tissue water relations in three Mediterranean shrubs co-occurring at a natural CO2 spring. Plant Cell Environ 23(12):1341–1351
Tyree MT, Hammel HT (1972) The measurement of turgor pressure and water relations of plants by pressure-bomb technique. J Exp Bot 23(74):267–282
Warren CR, Bleby T, Adams MA (2007) Changes in gas exchange versus leaf solutes as a means to cope with summer drought in Eucalyptus marginata. Oecologia 154(1):1–10. doi:10.1007/s00442-007-0803-2
White DA, Turner NC, Galbraith JH (2000) Leaf water relations and stomatal behavior of four allopatric Eucalyptus species planted in Mediterranean southwestern Australia. Tree Physiol 20(17):1157–1165
Yan MJ, Yamanaka N, Yamamoto F, Du S (2010) Responses of leaf gas exchange, water relations, and water consumption in seedlings of four semiarid tree species to soil drying. Acta Physiologiae Plantarum 32(1):183–189. doi:10.1007/s11738-009-0397-x
Acknowledgments
This research has been supported by the National Natural Science Foundation of China (No. 41171419), the Knowledge Innovation Project of Chinese Academy of Sciences (kzcx2-XB2-05, kzcx2-yw-BR-02), and the Core University Exchange Program of Japan Society for the Promotion of Science.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by J. Franklin.
M.-J. Yan and M. Yamamoto contributed equally to this work
Rights and permissions
About this article
Cite this article
Yan, MJ., Yamamoto, M., Yamanaka, N. et al. A comparison of pressure–volume curves with and without rehydration pretreatment in eight woody species of the semiarid Loess Plateau. Acta Physiol Plant 35, 1051–1060 (2013). https://doi.org/10.1007/s11738-012-1143-3
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11738-012-1143-3