
Abstract
Key message
No soil nutrient differences between two plantations. In contrast to NSC, N and P concentrations were greater in black locust than in Chinese pine. NSC negatively, N and P positively related to precipitation for both plantations.
Abstract
Precipitation is a key environmental factor affecting carbon (C), nitrogen (N), and phosphorus (P) status of plants and soils, especially in water-limited regions. However, there are potential differences among species in their sensitivity to C, N, and P in relation to variation in precipitation. We presented paired measurements of non-structural carbon (NSC), N, and P concentrations in plantations of N-fixing black locust (Robinia pseudoacacia L.) and coniferous Chinese pine (Pinus tabulaeformis Carrière) along a mean annual precipitation gradient on the Loess Plateau, China. The results showed that soil nutrients positively related to precipitation, but their differences between two plantations were not clearly visible. NSC concentrations of tree tissues were significantly greater in Chinese pine than in black locust. In contrast, the N and P concentrations and the N:P ratios were significantly greater in black locust than in Chinese pine. Leaves contained the highest N and P concentrations, whereas coarse roots contained the highest NSC concentrations. The lowest concentrations of NSC were in the stem wood. NSC concentrations were negatively related to precipitation, while N and P concentrations were positively related to precipitation for both tree plantations. The constant leaf N:P ratios indicated that the growth of Chinese pine was limited by the soil N supply, whereas black locust was limited by P. These results indicate that inherent physiological and biological processes differ with tree species, and when coupled with environmental conditions, influence the variations of C, N, and P in plant tissues to adaptation and resilience under drought stress.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Forest ecosystems account for the storage of 45% of the total carbon (C) stored in terrestrial ecosystems and play a critical role in reducing greenhouse gas effects (Bonan 2008). Forest ecosystems take up atmospheric CO2 through photosynthesis to produce carbohydrates. Carbohydrates in trees consist of two primary components: structural (an immovable pool with a long lifetime, such as cellulose and lignin) and non-structural C (NSC, a metabolically active pool with a short lifetime, mainly soluble sugars and starch) (Dietze et al. 2014; Kozlowski 1992). In addition to having a role in non-growth mechanisms such as plant metabolism and maintaining cell turgor, NSC is also considered a mobile C supply for tree growth, and reflects the capital for flushing, reproduction, and buffering of trees to various stresses and disturbances, including drought, flooding, freezing, and pollution (Dietze et al. 2014; O’Brien et al. 2014; Villar-Salvador et al. 2015; Würth et al. 2005).
Precipitation is a key environmental factor in determining ecosystem structure and function, especially in water-limited regions (Meier and Leuschner 2014; Zhou et al. 2009). The changing patterns of precipitation due to climate change account for drought-induced tree die-back, declines in net primary production and C sequestration, alteration of biodiversity, and ecosystem functions and services (Anderegg et al. 2012). Under pressure from drought, low photosynthetic activity triggers stored NSC pools to fuel tree metabolism and provide energy for adaptive responses (Dietze et al. 2014; Gruber et al. 2012). Thus, NSC is posited as an important trait for resistance and resilience of forests to climate change-induced drought (Locosselli and Buckeridge 2017; O’Brien et al. 2014). An increasing number of studies have explored the variations of NSC concentrations to understand the mechanisms involved in tree response to water deficits, especially to test proposed C starvation hypotheses (Anderegg et al. 2012; Locosselli and Buckeridge 2017; O’Brien et al. 2014; Richardson et al. 2013; Ryan 2011). Trees are vulnerable to C starvation, which is induced by a reduction in photosynthetic activity from drought stress. Greater decreases in respiration activity relative to photosynthetic activity often result in the severe and eventually lethal decrease in NSC reserves (McDowell et al. 2008; Sala et al. 2012). However, due to the complexity of NSC responses to drought stress, factors such as drought intensity and progression, as well as tree size, age, tissue type, and species, a direct link between NSC and drought resistance and its relative importance to other variables remains unclear (Gruber et al. 2012; O’Brien et al. 2015; Sala et al. 2010; Zhang et al. 2015).
Within- and among-species variations in NSC concentrations have been widely studied in temperate and tropical forest trees, even in fruit trees (Boldingh et al. 2000; Hoch et al. 2003; Würth et al. 2005). It is widely accepted that evergreen conifers have greater NSC concentrations and pools than deciduous trees (Michelot et al. 2012; Richardson et al. 2015; Thomas and Martin 2012). However, a study observed that there were no significant differences between evergreen and deciduous groups or between ring- and diffuse-porous species (Zhang et al. 2014). Leaves usually have greater NSC concentrations than other plant tissues, but stems as the major pools of NSC because of their large biomass proportions (Barbaroux et al. 2003; Würth et al. 2005). Moreover, concentrations of NSC, sugar, and starch were much greater in bark than in stem wood (Richardson et al. 2015; Zhang et al. 2014). In addition, NSC-enriched seedlings showed greater stem water potentials and improved the survival advantage within ten tropical tree seedlings under drought (O’Brien et al. 2014). Moreover, NSC has been shown to increase with drought in Scots pine (Pinus sylvestris L.), Ponderosa pine (Pinus ponderosa (Dougl.) Laws), and Douglas-fir (Pseudotsuga menziesii), and declines in NSC have been observed in Longleaf pine (Pinus palustris Mill.) and Trembling aspen (Populus tremuloides Michx.), while remaining relatively unchanged in Pinon pine (Pinus edulis) and Utah juniper (Juniperus osteosperma) (Anderegg 2012; Anderegg and Anderegg 2013; Sala and Hoch 2009; Sayer and Haywood 2006; Woodruff and Meinzer 2011). However, NSC were three times lower in pinon pine than one-seed juniper (Juniperus monosperma) suggesting that pinon pine prefers to exhaust stored C during the growing season (Woodruff et al. 2015). Therefore, understanding patterns of NSC between forest types is highly relevant and critical in the context of tree resilience to global change (Richardson et al. 2015).
In addition to NSC, N and P also play vital roles in plant function, and are among the most important limiting nutrients in terrestrial ecosystems (Han et al. 2005; He and Dijkstra 2014; Sardans et al. 2013). Forest sites with greater precipitation and moister soils provide better conditions for the supply of nutrients to the roots through mass flow and diffusion (Cramer et al. 2009; Kreuzwieser and Gessler 2010). Drought can depress plant growth by reducing N and P uptake, transport, and redistribution (Sardans and Penuelas 2012). A meta-analysis showed that drought stress had negative effects on plant N (− 3.73%) and P (− 9.18%), but a positive effect on plant N:P (+ 6.98%) (He and Dijkstra 2014). In addition, numerous studies have focused on the N:P ratios to advanced our understanding ecosystem function and processes under drought stress (Cao and Chen 2017; He and Dijkstra 2014; Yuan and Chen 2009; Zhang et al. 2017). However, the inconsistent relationships between drought stress and plant N, P, and N:P in many field and plot studies were affected by various factors, including frequency and occurrence of rainfall, soil type affecting water potential, and evapotranspiration demand (Farooq et al. 2009; He and Dijkstra 2014).
In the present study, we investigated NSC, N, and P concentrations in leaves, branches, stems, bark, and fine and coarse roots between N-fixing black locust (Robinia pseudoacacia L.) and coniferous Chinese pine (Pinus tabulaeformis) growing in paired adjacent plantations along a mean annual precipitation gradient on the Loess Plateau, China. Black locust is a fast-growing exotic species and widely planted on the Loess Plateau, with more than 70,000 ha planted over the last few decades (Qiu et al. 2010). This exotic deciduous species is native to North America and is introduced to China from Europe at the beginning of the twentieth century (Cierjacks et al. 2013; Shan et al. 2002; Tsunekawa et al. 2014). Chinese pine is an indigenous conifer tree species and is widespread across central and northern China. Therefore, the ultimate objective of this study was to compare the varying response patterns of NSC, N, and P concentrations, and N:P ratios to a gradient of annual precipitation between a pair of neighboring plantations that rely on differing traits (deciduous vs. coniferous). We hypothesized that: (1) N-fixing black locust would have lower NSC, but greater N and P concentrations than coniferous Chinese pine and (2) the different relationships between the NSC, N, and P concentrations and the precipitation gradient for both plantations. The results of this investigation regarding the effect of drought stress on NSC, N, and P concentrations in N-fixing plantations compared to coniferous plantations may be useful for obtaining a comprehensive understanding of the ecological restoration process in this region.
Materials and methods
Study sites
Five counties, Huanglong (HL), Yichuan (YC), Ansai (AS), Suide (SD), and Shenmu (SM), were selected along an annual precipitation gradient from south to north in the Loess Plateau, China (Table 1). According to the previous studies on the spatial patterns of vegetation and climate on the Loess Plateau (Chen et al. 2008; Wang et al. 2011; Zhang et al. 2006), HL and YC are located within a forest region, where the annual precipitation varies ranges from 550 to 719 mm, the mean temperature ranges from 8.9 to 11.0 °C, the maximum evaporation ranges from 1150 to 1580 mm, and the accumulated temperature above 10 °C ranges from 3361 to 4700 °C. AS and SD are in the forest-steppe region, where the annual precipitation varies ranges from 400 to 550 mm, the mean temperature ranges from 6.7 to 9.0 °C, the maximum evaporation ranges from 1560 to 2000 mm, and the accumulated temperature ranges from 2350 to 3450 °C. SM is in the steppe region that has an annual precipitation ranges from 250 to 400 mm, a mean temperature ranges from 4.5 to 6.7 °C, a maximum evaporation ranges from 2004 to 2633 mm, and an accumulated temperature ranges from 2300 to 2890 °C.
Sampling and processing
With the guidance of local forestry bureau staff, three independent replicate sites of black locust and Chinese pine plantations with similar site conditions (located at the top of slopes, with an altitude ranging 1000–1400 m) were randomly selected in each county location. The distance among three selected sites was about 2–8 km within each county location, and the adjacent plantations of mature (around 40 years) black locust and Chinese pine were located around 30–500 m. All selected plantations were left unmanaged after established. A 20 m × 20 m plot was constructed in the central area of each plantation site for sampling. All the plots were studied during the time period of July–August 2013.
In each plot, soil samples were taken with a soil core (5 cm in diameter) from the top 0–20 cm soil layer of five randomly chosen locations. After removing the plant roots, fauna, and debris by hand, the samples were air-dried and mixed together to form a composite sample that was representative of the soil in the plot. Soil total N was measured according to the semi-micro Kjeldahl method and available N was analysed by the Alkali diffusion method (Bremner and Mulvaney 1982). Total P was digested with perchloric acid and sulfuric acid and determined using colorimetry (Parkinson and Allen 1975). Available P was extracted with sodium bicarbonate and measured with colorimetry (Olsen et al. 1954).
Diameter at breast height (DBH) and tree height was measured for all of the trees in each plot (Table 1). Samples of leaves, branches, stem wood, stem bark, and fine (< 2 mm in diameter) and coarse (> 2 mm in diameter) roots were collected from five trees representing the respective stand-specific DBH range in each plot. Living branches with leaves were collected using a pole pruner from the upper, middle, and lower parts of the canopy in all four cardinal compass directions, and divided into pooled samples of branches and leaves. Stem bark was obtained by scraping a 2 × 5 cm strip of bark from the sampled trees at DBH point, and then, samples of stem wood were obtained using an increment borer (Haglöf, Sweden) at both the south-north and east–west orientations to the centre of the stems at the DBH point of the sampled trees. Root samples were collected from the upper 50-cm soil layer using soil cores (9.8 cm in diameter) at a distance of 50 cm from the sampled trees in all four cardinal directions, and were then divided into fine and coarse roots. All the samples were immediately placed in a cooler until they were processed within 5 h after collection. The samples were microwaved at 600 W for 90 s to eliminate enzymatic activity and then dried to a constant weight at 60 °C (Gruber et al. 2012; Hoch et al. 2003; Li et al. 2013b). All dried samples were ground into fine powder in a Wiley Mill and stored in plastics before NSC, N, and P concentrations were analysed.
The NSC concentration is defined here as the sum of the concentrations of the soluble sugars and starch. Sugar and starch concentrations were analysed using the anthrone method (Li et al. 2013b; Yemm and Willis 1954; Zhang et al. 2015). The sample (0.1 g) was placed into a 10-ml centrifuge tube with an added 2 ml of 80% ethanol. The mixture was incubated at 80 °C in a shaking water bath for 30 min, and then centrifuged at 5000 g for 5 min. Two further extractions from the pellets were carried out with 80% ethanol. The supernatant was retained, combined, and stored at − 20 °C for soluble sugar determination. Starch was extracted from the ethanol-insoluble pellet after ethanol was first removed by evaporation. The starch in the residue was then released by boiling in 2-ml distilled water for 15 min. After cooling to room temperature, 2-ml 9.2-M HClO4 was added and the mixture was shaken for 15 min. Then, 4 ml of distilled water was added, and the mixture was centrifuged at 5000g for 5 min. An additional extraction was carried out with 2-ml 4.6-M HClO4. The supernatant was also retained, combined and stored at − 20 °C for starch determination. The supernatant for sugar and starch determination were both determined based on the absorbance at 625 nm in the presence of the same anthrone reagent in a spectrophotometer. Sugar and starch concentrations were calculated from the regression equations based on glucose standard solutions. However, the calculation of starch concentration needed to multiply the glucose concentration by a conversion factor of 0.9 (Osaki et al. 1991). Total N and P in tree tissues were also measured with the semi-micro Kjeldahl method and the digested with perchloric acid and sulfuric acid and determined using colorimetry, respectively (Bremner and Mulvaney 1982; Parkinson and Allen 1975).
Data analysis
All statistical analyses were performed with the SPSS software (version 20.0, IBM SPSS Statistics, USA). All comparisons were performed using analysis of variance (ANOVA) followed by multiple-comparison tests (Least Significant Difference tests). In addition, sample data were transformed to meet the assumptions of normality and homogeneity when needed. We performed regression analysis to express the response of the measured variables along the along the precipitation gradient. The level of statistical significance was taken as P < 0.05.
Results
Soil nutrients along the precipitation gradient
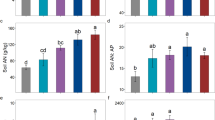
The concentrations of soil total N and the N:P ratios of black locust and Chinese pine plantations both decreased significantly with decreasing precipitation (Fig. 1a, c). Moreover, no significant difference on soil total N and the N:P ratios between two species was observed across all study sites (Fig. 1a, c). However, soil total P concentrations in both plantations remained relatively constant along the precipitation gradient, and a significant difference on soil total P concentrations between two tree plantations was observed at dryer sites (Fig. 1b).

Concentrations of soil total N (a), total P (b), N:P mass ratio (c), and soil available N (d) and available P (e) concentrations and their ratio (f) for black locust and Chinese pine plantations along a precipitation gradient on Loess Plateau, China. Error bars indicate the standard errors (n = 3). *Significant differences between black locust and Chinese pine stands at each site. The different uppercase and lowercase letters indicate significant differences among sites for black locust and Chinese pine stands, respectively. Only significant regression models are displayed
Soil available N and P concentrations accounted for approximately 6–7 and 0.3–0.6% of the total N and P concentrations in both tree species plantations, respectively. Both decreased significantly with decreasing precipitation (Fig. 1d, e). As a consequence, the soil available N:P ratio only in Chinese pine plantations decreased significantly with decreasing precipitation (Fig. 1f).
Trees NSC along the precipitation gradient
As we had hypothesized, NSC varied among different tree tissues of both plantations (Table 2; Fig. 2, see also Figs. S1 and S2). For all tree tissues except stem wood, the concentrations of NSC, as well as sugars and starch, were significantly lower in black locust than in Chinese pine (Table 2; Fig. 2). However, the concentration of NSC and sugars of stem wood in black locust were significantly greater than in Chinese pine, and there was no significant difference for starch between both plantations (Table 2). NSC and starch concentrations in the coarse roots and sugars in leaves were significant greater than those in other tree tissues, whereas NSC concentration, as well as sugar and starch, in stem wood was significantly lower than in other tree tissues for both tree plantations (Table 2).

Concentrations of non-structural carbon (NSC) in leaves (a), branches (b), stem wood (c), bark (d), fine roots (e), and coarse roots (f) for black locust and Chinese pine stands along a precipitation gradient on Loess Plateau. Error bars indicate the standard errors (n = 3). *Significant differences between black locust and Chinese pine stands. The different uppercase and lowercase letters indicate significant differences among sites for black locust and Chinese pine stands, respectively. Only significant regression models are displayed
The concentrations of NSC (including sugars and starch) in most tree tissues was linearly increased significantly with decreasing precipitation, but maintained relative stability in stem wood for both tree plantations (Fig. 2, see also Figs. S1 and S2). However, the concentrations of NSC and sugars in the coarse roots of Chinese pine were not sensitive to the precipitation gradient (Fig. 2, see also Figs. S1 and S2).
Tree N and p concentrations and N:P ratios along the precipitation gradient
The concentrations of N and P and their ratio also varied among different tree tissues (Table 2). Among all tree tissues, the highest and lowest concentrations of N and P were observed in leaves and stem wood, respectively, for both plantations (Table 2). Moreover, the N and P concentrations in all tree tissues and N:P ratio except in stem wood were significantly greater in black locust than in Chinese pine (Table 2; Figs. 3, 4, 5). As a consequence, the highest and lowest N:P ratios were in coarse roots and leaf for black locust, respectively, but were in stem wood and coarse roots for Chinese pine, respectively (Table 2).

N concentrations in leaves (a), branches (b), stem wood (c), bark (d), fine roots (e), and coarse roots (f) for black locust and Chinese pine stands for five sites along a precipitation gradient on Loess Plateau. Error bars indicate the standard errors (n = 3). *Significant differences between black locust and Chinese pine stands. The different uppercase and lowercase letters indicate significant differences along a precipitation gradient for black locust and Chinese pine stands, respectively. Only significant regression models are displayed

P concentrations in leaves (a), branches (b), stem wood (c), bark (d), fine roots (e), and coarse roots (f) for adjacent mature black locust and Chinese pine stands along a precipitation gradient on Loess Plateau. Error bars indicate the standard errors (n = 3). *Significant differences between black locust and Chinese pine stands. The different uppercase and lowercase letters indicate significant differences along a precipitation gradient for black locust and Chinese pine stands, respectively. Only significant regression models are displayed

N:P in leaves (a), branches (b), stem wood (c), bark (d), fine roots (e), and coarse roots (f) for black locust and Chinese pine stands along a precipitation gradient on Loess Plateau. Error bars indicate the standard errors (n = 3). *Significant differences between black locust and Chinese pine stands at each site. The different uppercase and lowercase letters indicate significant differences among sites for black locust and Chinese pine stands, respectively. Only significant regression models are displayed
Along the precipitation gradient, the concentrations of N and P in most of the tree tissues were linear and decreased significantly with decreasing precipitation, especially for black locust (Figs. 3, 4). For Chinese pine, the N concentrations in the stem wood and bark, as well as the P concentrations in leaves, stem wood, and fine roots, showed no significant trends over the precipitation gradient (Figs. 3, 4). As a result, the N:P ratios of most of the tree tissues for Chinese pine remained generally constant along the precipitation gradient (Fig. 5).
Discussion
Soil nutrients between plantations along the precipitation gradient
The concentrations of soil total N and P, and their ratios were demonstrated in this study between plantations of the deciduous N-fixing black locust and evergreen coniferous Chinese pine along a precipitation gradient. The annual precipitation of the five study sites we selected ranged from 440 to 602 mm. Precipitation influences soil chemistry and soil nutrient status via rock weathering, nutrient leaching, soil biological activity, the decomposition rate of plant litter, and the amount of nutrient return with litter fall (Austin and Vitousek 1998; Meier and Leuschner 2014; Sardans and Penuelas 2007). Many studies have observed that forest sites with greater precipitation and moister soils provide better nutrient conditions; thus, total soil nutrient and soil C:N and C:P ratios increase with increasing annual precipitation (Aranibar et al. 2004; Austin and Vitousek 1998; Kreuzwieser and Gessler 2010; Meier and Leuschner 2014). The results of this study agree with most previous studies that soil total N is highest at sites that receive the most rainfall and significantly decreases with decreasing precipitation (Fig. 1). In contrast to the positive relationship between soil total P and precipitation in the previous study sites (Meier and Leuschner 2014; Miller et al. 2001), soil total P (average 0.57 g kg−1 for both plantations) in the present study site remained relatively constant along a precipitation gradient, which is mainly attributed to the exceptionally low soil P concentration compared with global level (2.8 g kg−1), as well as to intensive leaching loss associated with serious soil erosion by wind and water on the Loess Plateau (Zhang et al. 2005, 2017; Zheng and Shangguan 2007).
The previous studies have found that soil available N and P concentrations were highest at drier sites, and decreased with increasing annual precipitation mainly because of the effect of increased rainfall on leaching exceed on weathering (Austin and Vitousek 1998; Vitousek et al. 1995). However, an inverse relationship between soil available N and P concentrations and precipitation in our study contradicts results from the previous studies. In addition, it was addressed that soil P availability is dependent on soil acidity and the amount of organic matter in the soil (Meier and Leuschner 2014). While we have no more data to further support our results, further studies may help to identify drivers that are directly or indirectly responsible for the trends in different soil N and P availability along the precipitation gradient.
It is widely understood that soil organic matter and N concentrations are greater in N-fixing forests than in non-N-fixing forests; this is because organic matter increases rates of N cycling and availability through plant fertilization (Deans et al. 1999; Macedo et al. 2008; Rice et al. 2004; Uselman et al. 2000; Wang et al. 2010). However, our study showed that concentrations of soil N and available N were not significantly different between black locust and Chinese pine plantations (Fig. 1). Tateno et al. (2007) also found no differences in soil N between black locust and oak forests on the Loess Plateau, and explained that black locust plantations might be more susceptible to soil erosion than oak forests. Therefore, the almost homogeneous soil nutrient substrate in our study between black locust and Chinese pine plantations at each site provided the precise environment to comprise the effects of tree species on the concentrations of NSC, N, and P along the precipitation gradient.
Trees NSC between plantations along the precipitation gradient
Our data showed that the concentrations of NSC, as well as sugar and starch, were significantly greater in Chinese pine than in black locust (Table 2; Fig. 2, see also Figs. S1 and S2). Our results were consistent with the previous reports that evergreen needles can sequester C concurrently and the needles from the previous year directly provided C for current growth; therefore, conifers used less NSC and also stored greater NSC than broadleaf evergreens (Michelot et al. 2012; Thomas and Martin 2012). However, a study found that broadleaf deciduous seedlings contain more NSC in roots than seedlings of broadleaf evergreens and conifers (Villar-Salvador et al. 2015). Richardson et al. (2015) found that the concentration and storage of NSC in woody tissues of white pine were significantly lower than in red oak. Whereas, other studies proved that there were no significant differences in concentrations of NSC for the 12 Chinese temperate tree species (Zhang et al. 2014) and 10 species in a European temperate forest (Hoch et al. 2003).
Concentrations of NSC also vary considerably with tree tissues. Leaves had the highest concentrations of sugar, whereas coarse roots had the highest values for starch and NSC for both plantations (Fig. 2). Our findings agree with the generally accepted observation that high concentration of NSC in tissues often have small biomass proportions (Zhang et al. 2014). The stem wood consistently had the lowest concentrations of NSC for both plantations (Table 2), which mainly due to C concentration in stems increases from the phloem to sapwood and heartwood, reaching its maximum in the bark (Bert and Danjon 2006; Chantuma et al. 2009; Li et al. 2013a; Zhang et al. 2014). Moreover, the functions of starch in stem wood acts as a long-term reserve of NSC, while soluble sugars as an intermediate, ready-to-use compartment of NSC (Bellasio et al. 2014; Richardson et al. 2013). However, the highest mean NSC concentrations in stems or branches and the lowest concentrations of NSC in fine roots were observed in 17 tree species in tropical forests in the Republic of Panama (Würth et al. 2005).
In addition, sampling height also had a significant influence on NSC concentrations. For example, Zhang et al. (2014) observed that the concentrations of NSC in the tree stumps were greater than other above heights (breast height, crown base, and mid-crown), and also sampling height significantly influenced sugar concentrations for all conifers, but not for broadleaved species. Moreover, we observed that the concentrations of NSC, as well as sugar and starch, were significantly greater in the coarse roots than in the fine roots for both tree plantations. However, Richardson et al. (2015) found similar results for red oak but the opposite results for white pine in Harvard Forest, USA. Therefore, species type, plant tissues, sampling data, and the protocol of select sampling trees may be attributed to the discrepancy in outcomes (Bert and Danjon 2006; Zhang et al. 2014, 2013). Recently, dendrobiochemistry as a fascination new concept proposed to study the connections between the dynamics of NSC and tree ring formation, which would provide new perspectives for understanding the responses of trees to the global climate change (Locosselli and Buckeridge 2017; Luttge 2017).
The NSC concentrations in most tree tissues increased with decreased precipitation (Fig. 2), which mainly due to the both increased sugar and starch concentrations along the precipitation gradient from wet to dry sites (Figs. S1 and S2). Körner (2003) and Würth et al. (2005) also observed that the NSC concentrations in all tree tissues increased as soil drought stress increased, and they addressed that this increase was largely due to starch. Moreover, it was notable that the concentrations of NSC (also starch and sugar) in the stem for both plantations remained constant along the precipitation gradient.
Tree N and p concentrations and N:P ratios between plantations along the precipitation gradient
Generally, leaves of N-fixing plants contain a greater concentration of N than P, and thus a greater N:P ratio than that of non-N-fixing plants (Killingbeck 1996; Li et al. 2014; Rice et al. 2004; Vergutz et al. 2012; Wright et al. 2004). Other tree tissues in our study also had significantly greater concentrations of N and P in black locust than in Chinese pine (Table 2). The highest N and P concentrations in leaves of black locust (Table 2) were consistent with the previous findings because of the photosynthetic machinery (N-rich compounds) and nuclear acid (P-rich compounds) production in metabolically active tissues (leaves vs. other tree tissues) or faster growing species (black locust vs. Chinese pine) (Li et al. 2013a; Matzek and Vitousek 2009; Reef et al. 2010). The stem wood consistently had the lowest concentrations of N and P for both plantations (Table 2) mainly due to the dilution effects of greater proportions of C-enriched structural compounds such as lignin and cellulose in stem (Li et al. 2013a). It is notable that there were no consistent rank trends among all tree tissues between the highest and lowest concentrations of NSC, N, and P for both plantations (Table 2).
Reduced rainfall significantly reduces the soil C, N, P, and other nutrients elements, and also accounts for a feedback on plant C and nutrient elements, and their growth (Kreuzwieser and Gessler 2010; Yuan and Chen 2015). In contrast to NSC, the concentrations of N and P and N:P ratio of most tree tissues decreased with decreasing precipitation (Figs. 2, 3, 4). The N and P concentrations in tree tissues exhibit different characteristic patterns for both tree plantations along the precipitation gradient. Moreover, a previous study also found more inconsistent results within the variation of N and P concentrations along the precipitation gradient. For example, at the global scale, leaf N concentrations significantly increased, but P concentrations decreased with increasing mean annual precipitation (Yuan and Chen 2009). Whereas leaf P concentrations of Ohi’a Lehua (Metrosideros polymorpha) in Hawai’i varied significantly with increasing precipitation, and especially increased from sites with a precipitation of 2000 mm to sites with a precipitation of 5500 mm, N concentrations did not (Austin and Vitousek 1998). Moreover, Santiago et al. (2004) found that both leaf N and P concentrations decreased with an increase in mean annual precipitation in tropical regions. Therefore, a better understanding of the factors, including soil type, region, temperature, availability of water, and interactions with other nutrients, that influence leaf N and P composition can contribute to a better understanding of subsequent consequences for plant communities (Ordonez et al. 2009; Sardans et al. 2011).
Leaf N:P ratios were used to indicate N limitation or P limitation in ecosystems, i.e., N:P ratios < 14 suggest N limitation and N:P ratios > 16 suggest P limitation (Reich 2005). In this study, the N:P ratios in leaves remained relatively constant along the precipitation gradient for both plantations, and they indicated P limitation for black locust plantations and N limitation for Chinese pine plantations (Fig. 5a). However, the leaf N:P and C:P ratios on a global scale increased, but C:N ratios decreased with increasing mean annual precipitation (Yuan and Chen 2009). However, other studies did not detect the correlations between leaf N:P ratios and precipitation in tropical areas (Townsend et al. 2007) or at the national level in the Loess Plateau area of China (Han et al. 2005; Zheng and Shangguan 2007).
Conclusion
The concentrations of soil total N and available N and P significantly decreased with decreasing mean annual precipitation along a south-to-north gradient on the Loess Plateau, China. Differing feedbacks on tree NSC, N, and P concentrations were observed between N-fixing black locust and coniferous Chinese pine. The concentrations of NSC were significantly greater in Chinese pines than in black locusts, and the NSC concentrations increased with decreased precipitation for both plantations. In addition, the concentrations of N and P were significantly greater in black locust than in Chinese pine and the concentrations of N and P in most tree tissues, especially for black locust, decreased with the decrease in precipitation for both plantations. Moreover, the constant N:P ratios in leaves were indicative of P limitation for black locust plantations and N limitation for Chinese pine plantations. Therefore, N-fixing black locust and coniferous Chinese pine exhibited differing eco-physiological characteristics on the patterns of NSC, N, and P concentrations when accounting for a shift from relatively high nutrient availability to relatively high carbon gain during periods of increased drought stress.
References
Anderegg WRL (2012) Complex aspen forest carbon and root dynamics during drought A letter. Clim Change 111:983–991
Anderegg WRL, Anderegg LDL (2013) Hydraulic and carbohydrate changes in experimental drought-induced mortality of saplings in two conifer species. Tree Physiol 33:252–260
Anderegg WRL, Berry JA, Smith DD, Sperry JS, Anderegg LDL, Field CB (2012) The roles of hydraulic and carbon stress in a widespread climate-induced forest die-off. Proc Natl Acad Sci 109:233–237
Aranibar JN, Otter L, Macko SA, Feral CJW, Epstein HE, Dowty PR, Eckardt F, Shugart HH, Swap RJ (2004) Nitrogen cycling in the soil-plant system along a precipitation gradient in the Kalahari sands. Global Change Biol 10:359–373
Austin AT, Vitousek PM (1998) Nutrient dynamics on a precipitation gradient in Hawai’i. Oecologia 113:519–529
Barbaroux C, Breda N, Dufrene E (2003) Distribution of above-ground and below-ground carbohydrate reserves in adult trees of two contrasting broad-leaved species (Quercus petraea and Fagus sylvatica). New Phytol 157:605–615
Bellasio C, Fini A, Ferrini F (2014) Evaluation of a high throughput starch analysis optimised for wood. PLoS One 9:e86645
Bert D, Danjon F (2006) Carbon concentration variations in the roots, stem and crown of mature Pinus pinaster (Ait.). For Ecol Manage 222:279–295
Boldingh H, Smith GS, Klages K (2000) Seasonal concentrations of non-structural carbohydrates of five Actinidia species in fruit, leaf and fine root tissue. Ann Bot 85:469–476
Bonan GB (2008) Forests and climate change: Forcings, feedbacks, and the climate benefits of forests. Science 320:1444–1449
Bremner J, Mulvaney C (1982) Nitrogen-total. Methods of soil analysis. Part 2. Chemical and microbiological properties, pp 595–624
Cao Y, Chen YM (2017) Coupling of plant and soil C:N:P stoichiometry in black locust (Robinia pseudoacacia) plantations on the Loess Plateau, China. Trees-Struct Funct 31:1559–1570
Chantuma P, Lacointe A, Kasemsap P, Thanisawanyangkura S, Gohet E, Clement A, Guilliot A, Ameglio T, Thaler P (2009) Carbohydrate storage in wood and bark of rubber trees submitted to different level of C demand induced by latex tapping. Tree Physiol 29:1021–1031
Chen HS, Shao MG, Li YY (2008) Soil desiccation in the Loess Plateau of China. Geoderma 143:91–100
Cierjacks A, Kowarik I, Joshi J, Hempel S, Ristow M, von der Lippe M, Weber E (2013) Biological flora of the british isles: robinia pseudoacacia. J Ecol 101:1623–1640
Cramer MD, Hawkins HJ, Verboom GA (2009) The importance of nutritional regulation of plant water flux. Oecologia 161:15–24
Deans JD, Diagne O, Lindley DK, Dione M, Parkinson JA (1999) Nutrient and organic-matter accumulation in Acacia senegal fallows over 18 years. For Ecol Manage 124:153–167
Dietze MC, Sala A, Carbone MS, Czimczik CI, Mantooth JA, Richardson AD, Vargas R (2014) Nonstructural carbon in woody plants. Annu Rev Plant Biol 65 65:667–687
Farooq M, Wahid A, Kobayashi N, Fujita D, Basra SMA (2009) Plant drought stress: effects, mechanisms and management. Agron Sustain Dev 29:185–212
Gruber A, Pirkebner D, Florian C, Oberhuber W (2012) No evidence for depletion of carbohydrate pools in Scots pine (Pinus sylvestris L.) under drought stress. Plant Biol 14:142–148
Han WX, Fang JY, Guo DL, Zhang Y (2005) Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China. New Phytol 168:377–385
He M, Dijkstra FA (2014) Drought effect on plant nitrogen and phosphorus: a meta-analysis. New Phytol 204:924–931
Hoch G, Richter A, Korner C (2003) Non-structural carbon compounds in temperate forest trees. Plant Cell Environ 26:1067–1081
Killingbeck KT (1996) Nutrients in senesced leaves: Keys to the search for potential resorption and resorption proficiency. Ecology 77:1716–1727
Körner C (2003) Carbon limitation in trees. J Ecol 91:4–17
Kozlowski TT (1992) Carbohydrate sources and sinks in woody-plants. Bot Rev 58:107–222
Kreuzwieser J, Gessler A (2010) Global climate change and tree nutrition: influence of water availability. Tree Physiol 30:1221–1234
Li H, Li J, He YL, Li SJ, Liang ZS, Peng CH, Polle A, Luo ZB (2013a) Changes in carbon, nutrients and stoichiometric relations under different soil depths, plant tissues and ages in black locust plantations. Acta Physiol Plant 35:2951–2964
Li MH, Cherubini P, Dobbertin M, Arend M, Xiao WF, Rigling A (2013b) Responses of leaf nitrogen and mobile carbohydrates in different Quercus species/provenances to moderate climate changes. Plant Biol 15:177–184
Li XW, Sun K, Li FY (2014) Variation in leaf nitrogen and phosphorus stoichiometry in the nitrogen-fixing Chinese sea-buckthorn (Hippophae rhamnoides L. subsp sinensis Rousi) across northern China. Ecol Res 29:723–731
Locosselli GM, Buckeridge MS (2017) Dendrobiochemistry, a missing link to further understand carbon allocation during growth and decline of trees. Trees-Struct Funct 31:1745–1758
Luttge U (2017) From dendrochronology and dendroclimatology to dendrobiochemistry. Trees-Struct Funct 31:1743–1744
Macedo MO, Resende AS, Garcia PC, Boddey RM, Jantalia CP, Urquiaga S, Campello EFC, Franco AA (2008) Changes in soil C and N stocks and nutrient dynamics 13 years after recovery of degraded land using leguminous nitrogen-fixing trees. For Ecol Manage 255:1516–1524
Matzek V, Vitousek PM (2009) N: P stoichiometry and protein : RNA ratios in vascular plants: an evaluation of the growth-rate hypothesis. Ecol Lett 12:765–771
McDowell N, Pockman WT, Allen CD, Breshears DD, Cobb N, Kolb T, Plaut J, Sperry J, West A, Williams DG, Yepez EA (2008) Mechanisms of plant survival and mortality during drought: why do some plants survive while others succumb to drought? New Phytol. 178:pp 719–739
Meier IC, Leuschner C (2014) Nutrient dynamics along a precipitation gradient in European beech forests. Biogeochemistry 120:51–69
Michelot A, Simard S, Rathgeber C, Dufrêne E, Damesin C (2012) Comparing the intra-annual wood formation of three European species (Fagus sylvatica, Quercus petraea and Pinus sylvestris) as related to leaf phenology and non-structural carbohydrate dynamics. Tree Physiol 32:1033–1045
Miller AJ, Schuur EAG, Chadwick OA (2001) Redox control of phosphorus pools in Hawaiian montane forest soils. Geoderma 102:219–237
O’Brien MJ, Leuzinger S, Philipson CD, Tay J, Hector A (2014) Drought survival of tropical tree seedlings enhanced by non-structural carbohydrate levels. Nat Clim Change 4:710–714
O’Brien MJ, Burslem DFRP., Caduff A, Tay J, Hector A (2015) Contrasting nonstructural carbohydrate dynamics of tropical tree seedlings under water deficit and variability. New Phytol 205:1083–1094
Olsen S, Cole C, Watanabe F, Dean L (1954) Estimation of available phosphorus in soils by extraction with sodium bicarbonate. Circular 939. Washington, DC, USDA
Ordonez JC, van Bodegom PM, Witte JPM, Wright IJ, Reich PB, Aerts R (2009) A global study of relationships between leaf traits, climate and soil measures of nutrient fertility. Global Ecol Biogeogr 18:137–149
Osaki M, Shinano T, Tadano T (1991) Redistribution of carbon and nitrogen compounds from the shoot to the harvesting organs during maturation in field crops. Soil Sci Plant Nutr 37:117–128
Parkinson J, Allen S (1975) A wet oxidation procedure suitable for the determination of nitrogen and mineral nutrients in biological material. Commun Soil Sci Plant Anal 6:1–11
Qiu LP, Zhang XC, Cheng JM, Yin XQ (2010) Effects of black locust (Robinia pseudoacacia) on soil properties in the loessial gully region of the Loess Plateau, China. Plant Soil 332:207–217
Reef R, Ball MC, Feller IC, Lovelock CE (2010) Relationships among RNA: DNA ratio, growth and elemental stoichiometry in mangrove trees. Funct Ecol 24:1064–1072
Rice SK, Westerman B, Federici R (2004) Impacts of the exotic, nitrogen-fixing black locust (Robinia pseudoacacia) on nitrogen-cycling in a pine-oak ecosystem. Plant Ecol 174:97–107
Richardson AD, Carbone MS, Keenan TF, Czimczik CI, Hollinger DY, Murakami P, Schaberg PG, Xu XM (2013) Seasonal dynamics and age of stemwood nonstructural carbohydrates in temperate forest trees. New Phytol 197:850–861
Richardson AD, Carbone MS, Huggett BA, Furze ME, Czimczik CI, Walker JC, Xu XM, Schaberg PG, Murakami P (2015) Distribution and mixing of old and new nonstructural carbon in two temperate trees. New Phytol 206:590–597
Ryan MG (2011) Tree responses to drought. Tree Physiol 31:237–239
Sala A, Hoch G (2009) Height-related growth declines in ponderosa pine are not due to carbon limitation. Plant Cell Environ 32:22–30
Sala A, Piper F, Hoch G (2010) Physiological mechanisms of drought-induced tree mortality are far from being resolved. New Phytol 186:274–281
Sala A, Woodruff DR, Meinzer FC (2012) Carbon dynamics in trees: feast or famine? Tree Physiol 32:764–775
Santiago LS, Kitajima K, Wright SJ, Mulkey SS (2004) Coordinated changes in photosynthesis, water relations and leaf nutritional traits of canopy trees along a precipitation gradient in lowland tropical forest. Oecologia 139:495–502
Sardans J, Penuelas J (2007) Drought changes phosphorus and potassium accumulation patterns in an evergreen Mediterranean forest. Funct Ecol 21:191–201
Sardans J, Penuelas J (2012) The role of plants in the effects of global change on nutrient availability and stoichiometry in the plant-soil system. Plant Physiol 160:1741–1761
Sardans J, Rivas-Ubach A, Penuelas J (2011) Factors affecting nutrient concentration and stoichiometry of forest trees in Catalonia (NE Spain). For Ecol Manage 262:2024–2034
Sardans J, Rivas-Ubach A, Estiarte M, Ogaya R, Penuelas J (2013) Field-simulated droughts affect elemental leaf stoichiometry in Mediterranean forests and shrublands. Acta Oecolog-Int J Ecol 50:20–31
Sayer MAS, Haywood JD (2006) Fine root production and carbohydrate concentrations of mature longleaf pine (Pinus palustris P. Mill.) as affected by season of prescribed fire and drought. Trees-Structure Function 20:165–175
Shan C, Liang Z, Hao W (2002) Review on growth of locust and soil water in Loess Plateau. Acta Bot Boreali-Occident Sinica 23:1341–1346
Tateno R, Tokuchi N, Yamanaka N, Du S, Otsuki K, Shimamura T, Xue Z, Wang S, Hou Q (2007) Comparison of litterfall production and leaf litter decomposition between an exotic black locust plantation and an indigenous oak forest near Yan’an on the Loess Plateau, China. For Ecol Manage 241:84–90
Thomas SC, Martin AR (2012) Carbon content of tree tissues: a synthesis. Forests 3:332–352
Townsend AR, Cleveland CC, Asner GP, Bustamante MMC (2007) Controls over foliar N: P ratios in tropical rain forests. Ecology 88:107–118
Tsunekawa A, Liu G, Yamanaka N, Du S (2014) Restoration and Development of the Degraded Loess Plateau, China. Springer, New York
Uselman SM, Qualls RG, Thomas RB (2000) Effects of increased atmospheric CO2, temperature, and soil N availability on root exudation of dissolved organic carbon by a N-fixing tree (Robinia pseudoacacia L.). Plant Soil 222:191–202
Vergutz L, Manzoni S, Porporato A, Novais RF, Jackson RB (2012) Global resorption efficiencies and concentrations of carbon and nutrients in leaves of terrestrial plants. Ecol Monogr 82:205–220
Villar-Salvador P, Uscola M, Jacobs DF (2015) The role of stored carbohydrates and nitrogen in the growth and stress tolerance of planted forest trees. New Forest 46(5–6):813–839
Vitousek PM, Turner DR, Kitayama K (1995) Foliar Nutrients during long-term soil development in Hawaiian montane rain-forest. Ecology 76:712–720
Wang FM, Li ZA, Xia HP, Zou B, Li NY, Liu J, Zhu WX (2010) Effects of nitrogen-fixing and non-nitrogen-fixing tree species on soil properties and nitrogen transformation during forest restoration in southern China. Soil Sci Plant Nutr 56:297–306
Wang YQ, Shao MA, Zhu YJ, Liu ZP (2011) Impacts of land use and plant characteristics on dried soil layers in different climatic regions on the Loess Plateau of China. Agric For Meteorol 151:437–448
Woodruff DR, Meinzer FC (2011) Water stress, shoot growth and storage of non-structural carbohydrates along a tree height gradient in a tall conifer. Plant Cell Environ 34:1920–1930
Woodruff DR, Meinzer FC, Marias DE, Sevanto S, Jenkins MW, McDowell NG (2015) Linking nonstructural carbohydrate dynamics to gas exchange and leaf hydraulic behavior in Pinus edulis and Juniperus monosperma. New Phytol 206:411–421
Wright IJ, Reich PB, Westoby M, Ackerly DD, Baruch Z, Bongers F, Cavender-Bares J, Chapin T, Cornelissen JHC, Diemer M, Flexas J, Garnier E, Groom PK, Gulias J, Hikosaka K, Lamont BB, Lee T, Lee W, Lusk C, Midgley JJ, Navas ML, Niinemets U, Oleksyn J, Osada N, Poorter H, Poot P, Prior L, Pyankov VI, Roumet C, Thomas SC, Tjoelker MG, Veneklaas EJ, Villar R (2004) The worldwide leaf economics spectrum. Nature 428:821–827
Würth MKR, Peláez-Riedl S, Wright SJ, Körner C (2005) Non-structural carbohydrate pools in a tropical forest. Oecologia 143:11–24
Yemm E, Willis A (1954) The estimation of carbohydrates in plant extracts by anthrone. Biochem J 57:508–514
Yuan ZYY, Chen HYH (2009) Global trends in senesced-leaf nitrogen and phosphorus. Global Ecol Biogeogr 18:532–542
Yuan ZY, Chen HY (2015) Decoupling of nitrogen and phosphorus in terrestrial plants associated with global changes. Nat Clim Change 5(5):465
Zhang C, Tian H, Liu J, Wang S, Liu M, Pan S, Shi X (2005) Pools and distributions of soil phosphorus in China. Global Biogeochem Cycles 19:GB1020. https://doi.org/10.1029/2004GB002296
Zhang JT, Ru WM, Li B (2006) Relationships between vegetation and climate on the Loess Plateau in China. Folia Geobotanica 41:151–163
Zhang H, Wu H, Yu Q, Wang Z, Wei C, Long M, Kattge J, Smith M, Han X (2013) Sampling date, leaf age and root size: implications for the study of plant C: N: P stoichiometry. PLoS One 8(4):e60360
Zhang HY, Wang CK, Wang XC (2014) Spatial variations in non-structural carbohydrates in stems of twelve temperate tree species. Trees-Struct Funct 28:77–89
Zhang T, Cao Y, Chen Y, Liu G (2015) Non-structural carbohydrate dynamics in Robinia pseudoacacia saplings under three levels of continuous drought stress. Trees 29(6):1837–1849
Zhang G, Zhang P, Peng S, Chen Y, Cao Y (2017) The coupling of leaf, litter, and soil nutrients in warm temperate forests in northwestern China. Sci Rep 7(1):11754
Zheng S, Shangguan Z (2007) Spatial patterns of leaf nutrient traits of the plants in the Loess Plateau of China. Trees-Struct Funct 21:357–370
Zhou XH, Talley M, Luo YQ (2009) Biomass, litter, and soil respiration along a precipitation gradient in Southern Great Plains, USA. Ecosystems 12:1369–1380
Acknowledgements
This research was supported by the National Nature Science Foundation of China (Nos. 41201088 and 41771556), and National Key R&D Program of China (2016YFC0501703 and 2017YFC0504605), and CAS “Light of West China” Program (XAB201702). The authors would like to acknowledge the contributions made by Christian J. Rivera (Princeton University, USA) regarding the English language revision of the manuscript.
Author information
Authors and Affiliations
Contributions
YC and YC designed the experiments. YC and YL carried out the experiments. YC wrote the manuscript with contributions from YL and YC.
Corresponding author
Ethics declarations
Conflict of interest
There are no conflicts of interest to declare.
Additional information
Communicated by M. Buckeridge.
Electronic supplementary material
Below is the link to the electronic supplementary material.
468_2018_1676_MOESM1_ESM.tif
Fig. S1. Sugars concentrations in leaves (a), branches (b), stem wood (c), bark (d), fine roots, (e) and coarse roots (f) for black locust and Chinese pine stands along a precipitation gradient on Loess Plateau. Error bars indicate the standard errors (n=3). * indicate significant differences between black locust and Chinese pine stands at each site. The different uppercase and lowercase letters indicate significant differences among sites for black locust and Chinese pine stands, respectively. Only significant regression models are displayed. (TIF 56527 KB)
468_2018_1676_MOESM2_ESM.tif
Fig. S2. Starch concentrations in leaves (a), branches (b), stem wood (c), bark (d), fine roots, (e) and coarse roots (f) for black locust and Chinese pine stands along a precipitation gradient on Loess Plateau. Error bars indicate the standard errors (n=3). * indicate significant differences between black locust and Chinese pine stands. The different uppercase and lowercase letters indicate significant differences along a precipitation gradient for black locust and Chinese pine stands, respectively. Only significant regression models are displayed. (TIF 56267 KB)
Rights and permissions
About this article

Cite this article
Cao, Y., Li, Y. & Chen, Y. Non-structural carbon, nitrogen, and phosphorus between black locust and chinese pine plantations along a precipitation gradient on the Loess Plateau, China. Trees 32, 835–846 (2018). https://doi.org/10.1007/s00468-018-1676-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00468-018-1676-1