
Abstract
To investigate influences of forest plantations on soil nutrient properties, biomass accumulation, major nutrient elements (NPK) and their stoichiometric couplings in different tissues and aged plants, and correlations between major nutrient contents in soils and in foliage of plants, 5-, 10-, 15- and 20-year-old plantations of black locust (Robinia pseudoacacia L.) and farmland were selected. Black locust plantations increased soil organic carbon (SOC) and N stocks by 23–327 and 23–119 %, respectively, in the 0–10 cm top soil layer compared to those in farmland. Soil C:N, C:P, C:K, N:P, N:K and P:K ratios were 10.1, 22.9, 0.7, 2.2, 0.7 and 0.03, respectively. These ratios were higher in the 0–10 cm soil layer than those in the 10–20 cm soil layer and increased under older plantations. Higher C contents in stem, N contents in leaf, the largest C pools in stem and N pools in root in 20-year-old plantation were observed. Correspondingly, the highest C:N, C:P and C:K and the lowest N:P and N:K ratios in stem, decreased C:N and C:P ratios in older trees were found. No strong correlations were observed between element contents in soils and in leaves of black locust trees. These results suggest that black locust plantations can increase soil nutrient concentrations, SOC and N stocks resulting in changes in element stoichiometric relations. CNPK contents and their stoichiometries vary with tissues and tree ages of black locust. No strong coupling relations exist between major nutrient element contents in the top soil and in foliage of black locust.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
The Loess Plateau is one of the most fragile ecosystems in China. Serious soil erosion occurred in this area mainly due to traditional farming activities. To prevent soil erosion, programs for afforestation and reforestation have been launched since 1978 (Cao et al. 2009). Since that time forest plantations have been chronologically established on previous farmlands on the Loess Plateau. To assess the sustainability of these measures, it is important to understand how reforestation processes influence soil carbon and nutrient concentrations.
Soil organic carbon (SOC) and other major nutrients such as nitrogen (N), phosphorus (P) and potassium (K) are often observed to increase after conversion of farmlands or grasslands to forests due to the accumulation of plant litter in soils under forest canopy (Menyailo et al. 2002; Cao et al. 2007; Hazlett et al. 2007; Dong et al. 2009; Qiu et al. 2010; Roy et al. 2010). Decomposition of litter, in turn, results in increases in SOC, N, P and K in the developing forest soils (Chen and Xu 2005; An and Huang 2006; Dong et al. 2009; Roy et al. 2010).
For ecosystem sustainability, both soil nutrient contents and soil nutrient stoichiometries are important (Sardans et al. 2011). According to Liebig’s “law of the minimum”, the nutrient, supply of which to plants is the most limiting, constrains the forest growth (Xia and Wan 2008). Accumulating evidence suggests that not only soil nutrient contents, but also nutrient element stoichiometric relations are important factors controlling ecosystem net primary productivity and sustainability (Agren 2008; Ladanai et al. 2010). Soil nutrient stoichiometric relations have been examined in soil microbial biomass and in the 0–10 cm top soil on the global scale (Cleveland and Liptzin 2007). In Chinese soils the mean C:N:P atomic ratio is 134:9:1, however, with high spatial heterogeneity and large variations in different soil depths (Tian et al. 2010). Previous studies on soil nutrient properties focus on forestland conversion to grassland or arable land (Lal et al. 2007). Although SOC, N, P and K concentrations in soil are often elevated after conversion of farmland to forestland (Menyailo et al. 2002; Cao et al. 2007; Hazlett et al. 2007; Laik et al. 2009; Qiu et al. 2010), little information is available on alteration in soil nutrient stoichiometric relations after land use changes (Wang et al. 2011; Yang et al. 2011).
Plants play a pivotal role in ecosystem nutrient cycling and stoichiometric relations (Matzek and Vitousek 2009; Yang et al. 2011; Reed et al. 2012). N, P and K are macronutrients for plants, which often limit growth in natural ecosystems (Agren 2008; Reef et al. 2010). Currently, our understanding of the specific functions of nutrients in plants has made advances, but the relative demand for these elements in different plant organs and ontogenetic stages is not well understood (Niklas and Cobb 2006; Agren 2008; Yang and Luo 2011). Previous studies have examined the N:P ratio at the whole plant level and reported N:P atomic ratios in the range from 5.5 to 66.4; an optimal N:P atomic ratio is suggested to be 22–44 depending on plant species and tissue ages (Gusewell 2004; Agren 2008; Elser et al. 2010). Most of these studies concentrate on natural communities/ecosystems in which nutrient status may be different from forest plantations (Agren 2008; Elser et al. 2010). Currently, scarce data are available on plant nutrient concentrations and nutrient element stoichiometric relations in forest plantations.
Plant tissue and age are important factors affecting nutrient concentrations and stoichiometry (Niklas and Cobb 2006; Yang and Luo 2011). In perennial plants, N, P and K are enriched in active growth tissues and the concentrations of these macronutrients may decrease with ontogenetic development (Niklas and Cobb 2006; Agren 2008). In trees N:C atomic ratio may vary from more than 8.6 in fast growing tissues such as leaves to less than 0.086 in woody stems (Agren 2008). In addition, increasing plant age also alters metabolite partitioning between tissues leading to changes in nutrient stoichiometric relations (Niklas and Cobb 2006; Agren 2008; Yang and Luo 2011). Decline in N:C, P:C and K:C mass ratios in foliage is observed during a growing season (Agren 2008). This decline in nutrient element:C mass ratio with ontogenetic development of plants is attributable to a more rapid formation of supportive, element-poor tissues (stems) in older plants than that of metabolically more active, element-rich tissues (leaves) (Agren 2008; Yang and Luo 2011). Furthermore, in forest plantations dynamics, forest age is regarded as a primary driver for changes in forest structure and function because age of woody plants alters photosynthetic capacity (Zheng et al. 2011), carbon storage in live biomass (Bradford et al. 2008), soil organic layer (Pregitzer and Euskirchen 2004), carbon balance (Bradford et al. 2008) and nutrient contents and stoichiometry (Agren 2008). Due to the chronological afforestation and reforestation on the Loess Plateau, forest age is expected to affect the nutrient concentrations and stoichiometry in these forest ecosystems, but no experimental evidence supports such assumption.
Black locust (Robinia pseudoacacia L.) is a fast growing tree species with ability to fix atmospheric N. This species has been widely planted on the Loess Plateau with more than 70,000 ha since the last few decades (Qiu et al. 2010). Previous studies mainly focused on effects of black locust plants on soil properties (Khan et al. 2010; Qiu et al. 2010) and no integrated investigations have been carried out on nutrient concentrations and stoichiometry in both plants and soils on the Loess Plateau. Thus, the aim of this study was to investigate changes in carbon, nutrients and stoichiometric relations under different soil depths, plant tissues and ages in black locust plantations. We used 5-, 10-, 15- and 20-year-old black locust plantations and the farmland to test following hypotheses: (i) black locust plantations increase soil nutrient concentrations and storage leading to changes in soil nutrient stoichiometric relations, (ii) the nutrient concentrations and stoichiometry in black locust trees display tissue- and age-specific patterns, and (iii) correlations exist between major nutrient concentrations in the top soils and those in leaves of black locust plantations. The experimental evidence to these hypotheses may provide novel insights for us to better understand ecological functions of forest plantations on degraded lands for sustainability in the context of global change.
Materials and methods
Experimental site and sampling
The study area is located in the Wuliwan watershed of Ansai County on the Loess Plateau of China (36°51′N, 109°20′E; altitude 1,245–1,285 m). The study area is characterized by a warm temperate sub-humid continental climate. The annual mean air temperature is 8.8 °C. The annual mean precipitation is 505 mm and 70 % of annual precipitation falls from July to September. The soil is typical loess as described by Zheng et al. (2011). In previous time the study area was farmland. At the beginning in the 1990s, forest plantations were gradually established on the farmlands aiming to alleviate soil erosion in this region.
In the study area, the age of black locust (R. pseudoacacia) plantation was determined by an increment borer and three plantations for each age (5, 10, 15 and 20 years) were randomly chosen on former farmlands. Further characteristics of these plantations are given in Table S1. In each plantation, three sample plots (plot size: 20 m × 20 m) were randomly selected. In each sample plot, six soil columns of 20 cm depth were randomly sampled by a soil auger (5 cm in diameter). Each soil column at the 0–10 cm and the 10–20 cm in depth was collected separately and brought back to the laboratory for further analysis. To compare the land use changes leading to alteration of nutrients between plantations and farmlands, three farmlands ca. 2 km away from the plantations of black locust were selected as the control and considered as the plantation of black locust (R. pseudoacacia L.) grown for 0 year. However, no plant tissues were harvested for analysis on the farmlands. The owners of farmlands confirmed that the management regime of farmlands (e.g., species cultivated, tillage frequency, organic/mineral fertilizations) remained unchanged since the establishment of the first plantations at the beginning in the 1990s. Soil samples in each farmland were collected according to the same scheme as the plantations.
In addition, the diameter at breast height of each tree in each sample plot was measured using calipers. In each sample, plot six sample trees were randomly selected for harvesting to determine biomass and biochemical properties. To harvest the selected tree, the whole root system was manually excavated. Subsequently, the root system, stem (including branches) and leaves were separated and fresh weight of each fraction (i.e., roots, stem with branches and leaves) was determined. Sub-samples of main root, stem (at breast height) and leaves were collected and kept on ice before being carried to the laboratory and stored at −80 °C for further analysis. The sampling was done between July and August 2009.
Analysis of soil major nutrient concentrations, soil bulk density and soil nutrient storage
To determine SOC, N, P and K, the samples from the 0 to 10 cm and the 10 to 20 cm soil layers were air-dried, then manually ground to fine powder (<0.25 mm) as proposed by Wang et al. (2011). SOC was determined by oil bath-K2CrO7 titration method (Nelson and Sommers 1975). Total soil N was determined by the Kjeldahl method (Qiu et al. 2010). Total soil P was determined colorimetrically after the digestion with H2SO4 and HClO4 (Parkinson and Allen 1975). K in soil was extracted with neutral ammonium acetate and subsequently measured by atomic absorption spectrometry (SpectrAA-220 Zeeman, Varian Inc., Palo Alto, CA) according to the method of Qiu et al. (2010).
To calculate soil C, N, P and K atomic ratios, the concentrations (g kg−1) of SOC, soil N, P and K were transformed to mol kg−1, and ratios of the elements (C:N, C:P, C:K, N:P, N:K and P:K) in the 0–10 cm and the 10–20 cm soil layers were calculated as molar ratios (atomic ratios), rather than mass ratios.
The soil bulk density was determined according to the method proposed by Wang et al. (2011). Subsequently, the SOC, soil N, P and K storage were calculated using the following formula suggested by Wang et al. (2011):
SOC, soil N, P and K (gm−2) = α × ρ × 0.1
where α is the concentrations of SOC, soil N, P or K (g kg−1), ρ is the soil bulk density (g cm−3 = ton m−3) and 0.1 represents the depth of each soil layer.
Biomass estimation of black locust
To determine tree biomass, sub-samples (ca. 15 % of fresh weight of each fraction) of main root, stem and leaves were ground to fine powder by a miller. About 500 g fine powder of fresh samples was dried to constant weight at 60° C and the dry weight was recorded. Subsequently, the ratio of dry to fresh weight of each tissue of each harvested tree was determined. The dry weight of each tissue of each harvested tree was estimated by the fresh weight multiplying the ratio of dry to fresh weight. Afterwards, an allometric relationship was established between dry biomass and diameters of harvested trees after the method proposed by Luo et al. (2006) (Table S2). Biomass per square meter of black locust plantations was estimated using the density of trees in the sample plots and the allometric relationship for mass and stem diameter.
Determination of main nutrients in black locust
Total C and N in roots, stem and leaves of black locust were determined by a CN analyzer (Luo and Polle 2009). Aliquots of 0.6–1.0 mg milled materials were weighed into 5 mm × 9 mm tin capsules and determined by the Elemental Analyzer EA1108 (Carlo Erba Strumentazione, Rodano, Milan, Italy). Acetanilide (71.09 % C, 10.36 % N; Carlo Erba Strumentazione, Rodano) was used as a standard. After acid digestion of fine powder according to Luo and Polle (2009), total P and K in black locust were determined as above. Using biomass of different organs and the nutrient concentrations in each organ, the pools of C, N, P and K were calculated. Nutrient ratios in plants were expressed as molar ratios.
Correlation analysis between leaf and soil nutrients
To investigate whether relationships existed between leaf and soil nutrients, correlation analysis was performed. The data of nutrient concentrations in leaf and soil were expressed as molar nutrient per kilogram dry material. After log10 transformation, linear correlation analysis was carried out between leaf nutrients and those in the two layers of soil (the 0–10 cm and the 10–20 cm), respectively.
Analysis of sugars, phenolics and lignin in black locust
Tree tissue composition, particularly, lignin is important for litter decomposition and nutrient release. Thus, sugars, phenolics and lignin were determined in black locust trees. Concentrations of soluble sugar and starch in plants were analyzed by the anthrone reagent (Zhang et al. 2009). Soluble phenolics in roots, stem and leaves of black locust were extracted and determined spectrophotometrically at 765 nm (UV-3802, Unico Instruments Co. Ltd., Shanghai, China) using the Folin-Ciocalteus reagent (Luo et al. 2008). Acid-insoluble (Klason) lignin in roots, stem and leaves of black locust was determined as described by Luo et al. (2008).
Statistical analysis
All statistical tests were performed with STATGRAPHICS (STN, St Louis, MO). The biomass of different aged plantations was analyzed by one-way ANOVA. For soil samples, the soil depth and age of black locust plantation were treated as two independent variables. For plant samples, the tissue and age of black locust plantation were considered as two independent variables. The data for each variable within a stand were used for statistical analysis. Two-way ANOVA was conducted with these independent variables. When the interaction was significant, a post hoc comparison of means was made. To reduce the chance of type I errors, all P values of these multi-comparisons were corrected by the Tukey-HSD method. Data were tested for normality prior to the statistical analysis. Differences between means were considered significant when the P value of the ANOVA F test was less than 0.05.
Results
SOC and major nutrients in soils
Concentrations of SOC were significantly higher in the 0–10 top soil layer than that in the 10–20 cm soil layer in different aged black locust plantations (Fig. 1a). In the top soil layer, the concentration of SOC was 2.2 g kg−1 in farmland, but enhanced to 2.7–10.9 g kg−1 in soils of black locust plantations and the most pronounced change was that SOC content in 20-year-old plantation was ca. five times higher than that in farmland. Among the soils of plantations, SOC contents in 15- and 20-year-old plantations were two and four times higher than that in 5-year-old plantation, respectively. In the 10–20 cm soil layer, SOC contents in farmland and soils of black locust plantations grown for 5 and 10 years were ca. 2.3, 2.4 and 2.8 g kg−1, respectively, which were similar to those in the top soil of these plantations. However, SOC contents in the 10–20 cm soil layer of 15- and 20-year-old plantations were 3.4 and 3.7 g kg−1, respectively, which only counted for 54 and 34 % of SOC contents in the top soil layer of these plantations. In the 10–20 cm soil layer, SOC contents displayed a trend of gradual increases from farmland to older forest plantations.

SOC, total N, total P and total K in soils under plantations of Robinia pseudoacacia grown for 0, 5, 10, 15 or 20 year(s). Bars indicate mean ± SE (n = 3). Different letters on the bars for the same soil depth indicate significant difference between soils under different aged plantations. P values of the ANOVAs of soil depth (P Depth) and plantation ages (P Age) were also shown. No interaction was significant. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001, ns not significant
N concentrations were also analyzed in soils (Fig. 1b). In the 0–10 cm soil layer, total N concentration was 0.34 g kg−1 in soils of farmland and 0.43–0.87 g kg−1 in soils of plantations. In the top soil layer, total N concentrations in plantations were higher by 27–156 % than that in farmland. Moreover, total N concentrations in the top soil layer were significantly higher under older plantations than that under younger plantations. In the lower (the 10–20 cm) soil layer, total N concentrations were similar to those in the top soil layer of farmland and plantations grown for 5 and 10 years, but were significantly decreased in plantations grown for 15 and 20 years in comparison with those in top soils of these plantations.
In the 0–10 cm soil layer, total P concentration was 0.57 g kg−1 in farmland and 0.55–0.59 g kg−1 in plantations, respectively (Fig. 1c). In the top soil layer, the highest P content was detected in 5-year-old plantation and P contents decreased in older plantations. In lower (the 10–20 cm) soil layer, total P concentrations were lower than those in the top soil layers of farmland and plantations and total P contents were higher in older plantations than those of younger plantations.
In the 0–10 cm soil layer, total K concentration was 18.3 g kg−1 in farmland and 17.2–19.9 g kg−1 in different aged black locust plantations (Fig. 1d). No soil depth effects were observed on soil total K contents. Total K concentrations in the 0–10 cm soil layer were pronouncedly higher in 5- and 10-year-old plantations than those of farmland, 5- and 20-year-old plantations. In the 10–20 cm soil layer, total K displayed similar pattern as the top soil layer.
To investigate the nutrient element balance in soils of farmland and black locust plantations, the molar ratios of SOC:elements were calculated (Fig. 2). In 0–20 cm soil, molar ratios of C:N, C:P, C:K, N:P, N:K and P:K were 6.9–14.9, 12.1–59.7, 0.395–2.099, 1.5–4.2, 0.051–0.145 and 0.029–0.034, respectively. All ratios were significantly higher in the 0–10 cm soil layer than those in the 10–20 cm soil layer except the C:N ratio. The nutrient element ratios were significantly higher in soils under older plantations compared to those in soils of farmland and younger plantations, and this effect was more obvious in the top (the 0–10 cm) soil layer.

Molar ratios of SOC to total N (C:N), SOC to total P (C:P), SOC to total K (C:K), total N to total P (N:P), total N to total K (N:K), total P to total K (P:K) in the 0–10 cm and the 10–20 cm depth soil under plantations of black locust grown for 0, 5, 10, 15 and 20 year(s). Data indicate mean ± SE (n = 3). Different letters on the bars indicate significant difference. P values of the ANOVAs of soil depth (P Depth) and plantation ages (P Age) were also shown. No interaction was significant. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001, ns not significant
Soil bulk density was also analyzed in farmlands and forestlands of black locust (Fig. 3). In farmland and forestlands of black locust with 15- and 20-year-old plantations, soil bulk density was lower by 7–11 % in the top (0–10 cm) soil layer than that in the 10–20 cm soil layer. Soil bulk densities were higher by ca. 17 % in the top soil layer of farmland and forestlands of black locust with 5- and 10-year-old plantations than that in the top soil layer of forestlands of black locust with 15- and 20-year-old plantations. Soil bulk density was higher by 20 % in the 10–20 cm soil layer of farmland than that in the 10–20 cm soil layer of forestlands with different ages which had similar soil bulk density.

Soil bulk density in the 0–10 cm and the 10–20 cm depth soil under plantations of black locust grown for 0, 5, 10, 15 and 20 year(s). Data indicate mean ± SE (n = 3). Different letters on the bars indicate significant difference. P-values of the ANOVAs of soil depth (P Depth) and plantation ages (P Age) were also shown. No interaction was significant. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001, ns not significant
Based on the SOC, soil N, P and K concentrations and soil bulk densities, storage of SOC, soil N, P and K in each soil layer was estimated in farmlands and forestlands (Fig. 4). SOC storage was higher by 15–169 % in the top soil layer than that in the 10–20 cm soil layer of forest plantations, but the opposite was true for farmland (Fig. 4a). SOC storage was gradually elevated in both soil layers of older forest plantations (Fig. 4a). SOC storage was significantly higher by 23–327 % in the top soil layers of black locust plantations than that in farmland (Fig. 4a). Similar to SOC, soil N storage was higher by 17–109 % in the top soil layer than that in the 10–20 cm soil layer of forest plantations (Fig. 4b), but soil N storage was higher by 7 % in the 10–20 cm layer than that in the top soil layer of farmland (Fig. 4b). Soil N storage was the lowest in the top soil layer of farmland and it was gradually enhanced in older plantations of black locust (Fig. 4b). In the oldest plantation of black locust, soil N storage was higher by 119 % in the top soil layer than that in farmland (Fig. 4b). Soil P storage was higher by ca. 20 % in the top soil layers of farmland and of 5- and 10-year-old plantations than that of 15- and 20-year-old plantations (Fig. 4c). Soil P storage was the highest in the 10–20 cm soil layer of farmland and the lowest in the 15-year-old plantation (Fig. 4c). Soil K storage was the highest in soil of farmland and the lowest of the 15-year-old plantation (Fig. 4d). Soil K storage was lower by 21 % in soil of the 15-year-old plantation than that of farmland (Fig. 4d).

Storage of SOC a, soil N b, soil P c and soil K d under plantations of Robinia pseudoacacia grown for 0, 5, 10, 15 or 20 year(s). Bars indicate mean ± SE (n = 3). Different letters on the bars indicate significant difference between soils under different aged plantations. P values of the ANOVAs of soil depth (P Depth) and plantation ages (P Age) were also shown. No interaction was significant. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001, ns not significant
Biomass of black locust plantations
Biomass of black locust plantations grown for 5, 10, 15 and 20 years was estimated based on the allometric relationship and the diameter of each tree in sample plots (Fig. 5). Biomass of black locust plantations grown for 5, 10, 15 and 20 years was 100, 320, 963 and 1,067 g m−2 for roots (Fig. 5a), 238, 970, 6,010 and 6,323 g m−2 for stem (Fig. 5b), and 59, 102, 370 and 383 g m−2 for leaves (Fig. 5c), respectively. Biomass of black locust plantations grown for 5 and 10 years was similar, but biomass was significantly larger for 15- and 20-year-old plantations.

Dry biomass of roots a, stem b and leaves c of Robinia pseudoacacia grown for 0, 5, 10, 15 or 20 year(s). Bars indicate mean ± SE (n = 3). Different letters on the bars for the same organs indicate significant difference between different aged plantations. P-values of the ANOVAs of plantation ages (P Age) were also shown. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001, ns not significant
Main nutrients in black locust plantations
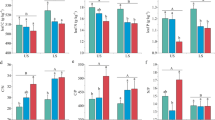
Concentrations of C, N, P and K in black locust trees were shown (Fig. 6). Among root, stem and leaf tissues, the C concentration was the highest in stem and the lowest in leaves (Fig. 6a). The mean C concentrations of roots, stem and leaves of black locust did not show any age-dependent changes (Fig. 6a). N concentrations in roots/leaves of 5-year-old black locust were significantly lower than those in roots/leaves of plantations grown for 10, 15 and 20 years, but N concentrations were quite similar in stem of black locust with different ages (Fig. 6b). P concentration was the highest in leaves of 20-year-old plants and the lowest in leaves of 5-year-old black locust (Fig. 6c). No age effects were found on total P concentrations in root/stem of black locust. Total K concentration was the lowest in roots of 10-year-old black locust and the highest in 5-year-old plants (Fig. 6d). Total K concentrations were similar in stem of different aged plants, but total K concentration was the lowest in leaves of 5-year-old plants in comparison with those in leaves of black locust grown for 10, 15 and 20 years (Fig. 6d).

Total C a, total N b, total P c and total K d in roots, stem and leaves of Robinia pseudoacacia grown for 0, 5, 10, 15 or 20 year(s). Bars indicate mean ± SE (n = 3). Different letters on the bars for the same organs indicate significant difference between different aged plantations. P-values of the ANOVAs of plant tissues (P Tissue) and plantation ages (P Age) were also shown. No interaction was significant. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001, ns not significant
To evaluate the total nutrient pools in black locust plantations, the amounts of C, N, P and K in root, stem and leaf biomass were calculated (Fig. 7). Among roots, stems and leaves, the C pools in stems were always the highest, irrespective of plantation age (Fig. 7a). The C pools were significantly elevated with increases in plantation age, irrespective of plant tissue (Fig. 7a). Among the three tissue types, the N pools were the highest in roots, irrespective of plantation age (Fig. 7b). Among different aged plantations, the N pools were the highest in 20-year-old plantation and the lowest in 5-year-old plantation, irrespective of plant tissue (Fig. 7b). Among different aged plantations, the P pools were the largest in 20-year-old plantation, irrespective of plant tissues (Fig. 7c). Roots and stems held large amounts of K in the plantations (Fig. 7d). The K pools were the largest in 20-year-old plantation among different aged plantations, irrespective of plant tissues (Fig. 7d).

The C a, N b, P c and K d pools in roots, stem and leaves of Robinia pseudoacacia grown for 0, 5, 10, 15 or 20 year(s). Bars indicate mean ± SE (n = 3). Different letters on the bars for the same organs indicate significant difference between different aged plantations. P-values of the ANOVAs of plant tissues (P Tissue) and plantation ages (P Age) were also shown. No interaction was significant. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001, ns not significant
The C:N, C:P and C:K ratios were significantly higher in stem than those in root and leaf, whereas the N:P and N:K ratios in stem were the lowest among the three tissues (Table 1). The P:K ratios in stem were higher than those in root, but lower than those in leaf. The C:N and C:P ratios decreased in older trees in comparison with those in younger trees.
Correlations between leaf and soil nutrients
Black locust is a deciduous legume species which increases N concentrations and storage in leaves and soil (Figs. 1b, 4b, 6b, 7b). Since the leaves are shed annually, recycling of nutrient elements from the leaf litter of this species may play a crucial role in soil nutrients. As a result, a positive impact of increasing N availability on leaf and soil nutrients may be expected, which in turn may influence the correlation relations between soil and plant nutrients. A significantly negative linear correlation was observed between foliar and soil P concentrations in the 10–20 cm layer (P = 0.0168, Table 2). For other elements (C, N and K), no significant correlations were found between foliar and soil element concentrations (Table 2).
Carbohydrates, phenolics and lignin in black locust
Tree tissue composition such as sugars, phenolics and lignin is particularly important for litter decomposition and nutrient release to the soil. Therefore, concentrations of soluble sugar, starch, soluble phenolics and lignin were determined in black locust trees (Table S3). Concentrations of soluble sugar were the highest in leaves and the lowest in stem, irrespective of plant ages. Concentrations of soluble sugar decreased in plants with increases in ages of plantations. In contrast to soluble sugar, concentrations of starch were the highest in roots, but the lowest in leaves of plants. Concentration of starch in roots of 15-year-old plant was the lowest in comparison with plants grown for 5, 10 and 20 years. Concentrations of soluble phenolics were the highest in leaves of plants. Concentrations of soluble phenolics were significantly higher in leaves and roots of plants grown for 5 and 10 years compared with plants grown for 15 and 20 years. Lignin contents were the highest in stems and the lowest in leaves of plants. Elevated lignin contents of plants with increases in plantation ages were found.
Discussion
Black locust plantations alters soil major nutrient storage resulting in changes in nutrient stoichiometric relations
SOC, total N, P and K in soil are crucial for ecosystem sustainability and productivity (Agren 2008; Dong et al. 2009; Laik et al. 2009; Ladanai et al. 2010). Current data indicate that higher SOC, soil N, P and K contents and storage of black locust plantations have led to changes in soil nutrient element ratios in comparison with those of farmland. After conversion of farmlands to Caragana korshinskii and larch (Larix gmelinii) plantations, SOC and soil N contents and storage are also significantly enhanced (An and Huang 2006; Wang et al. 2011). Reforestation in the degraded forest fringe areas also increases SOC and soil nutrients (Roy et al. 2010). These results suggest that restorative land uses such as afforestation/reforestation can play a crucial role in soil C storage and N accumulation (Jimenez et al. 2007). SOC is mainly derived from soil organic matter (litter, humus and living microbial biomass) which can stabilize soil structure against erosion and enhance soil nutrient availability (Wang et al. 2008; Roy et al. 2010). In this study, elevated SOC contents and storage in forestlands of black locust is likely due to accumulation of organic matter resulting from leaf and root litter, whereas on farmlands crop biomass is removed by harvesting. Moreover, black locust is characteristic for its atmospheric N fixation since it is a legume species. This species is estimated to have fixation as much as 75 kg N hm−2 y−1 (Olesniewicz and Thomas 1999). Consistently, SOC and soil N stocks have been enhanced by ca. 3.3- and 1.2-fold, respectively, in the 0–10 top soil layer of the 20-year-old black locust plantation compared to that of farmland. These results are consistent with previous studies which indicate that using woody legume species for afforestation/reforestation on degraded lands can enhance SOC and soil nutrition (Duponnois et al. 2005; Ferrari and Wall 2007; Lee et al. 2011). Furthermore, current data also suggest that increases in forest age lead to significant accumulation of SOC and soil N, although soil P and K stocks tend to be decreased in older plantations. This is probably because (i) litter is gradually accumulated and soil organic matter increases with the decomposition of litter which releases a significant amount of organic C and N to the soil (Lee et al. 2011; Forrester et al. 2013) and (ii) some fixed N by this woody legume species may be released to the soil via exudation of roots (Duponnois et al. 2005; Ferrari and Wall 2007). Overall, these data suggest that woody legume species such as R. pseudoacacia are good candidates for afforestation/reforestation on degraded land to prevent soil erosion and augment SOC and soil nutrients.
Changes in SOC, total N, P and K concentrations in soil inevitably lead to alterations in nutrient stoichiometric relations (Agren 2008; Tian et al. 2010). In this study, the altered nutrient element ratios in soil indicate that higher proportions of SOC, N and P are held in the 0–10 cm top soil layer compared to those in the 10–20 cm deep soil layer and these nutrient elements are enriched in soils of older plantations. The C:N (7.5–14.9), C:P (12.1–59.7) and N:P (1.6–4.2) molar ratios in the 0–10 cm soil layer of black locust plantations are consistent with a previous finding where the C:N, C:P and N:P mean ratios of 10.7, 38 and 3.6, respectively, have been found in the 0–10 cm soil layer of China (Tian et al. 2010). Consistent with our results, higher ratios of C:N, C:P and N:P in the 0–10 cm soil layer compared to those in deep soil have been observed by Tian et al. (2010). The higher C:N, C:P and C:K ratios in the top soil are probably attributable to marked accumulation of SOC in the top soil layer under 15- and 20-year-old plantations. The higher N:P and N:K ratios in the top soil layer are mainly due to (i) the significant increases in total N contents and stocks in the top soil layer, (ii) the relatively stable P and K contents in both layers and (iii) higher inputs of litter and fixed-N in the top soil layer of the older black locust plantations than those of young plantations. Similarly, higher element ratios in soils under black locust plantations compared to those in farmland are mainly due to the higher accumulation of SOC and N in soils of the older black locust plantations than those of farmlands, which is probably associated with higher inputs of litter and fixed-N of the black locust plantations. Our results are in line with previous findings which indicate that alterations in soil nutrient stoichiometric relations may occur in soils after land use changes (Wang et al. 2011; Chen et al. 2012). These results indicate that increases in SOC and soil N concentrations and stocks by black locust plantations can lead to alterations of soil nutrient stoichiometric relations.
Biomass, nutrients and stoichiometric relations in black locust display tissue- and age-specific patterns
Plants take up atmospheric CO2, soil N, P, K and other nutrient elements to synthesize various metabolites including structural compounds such as lignin and cellulose to build up plant body, i.e., biomass. In this study, biomass increments in roots, stems and leaves of black locust trees indicate that this species has slowed growth after ca. 15–20 years because no biomass increments are observed in 15- and 20-year-old plantations. Moreover, current data suggest that C, N, P and K contents, their pools and stoichiometric relations in black locust trees displayed tissue- and age-specific patterns. Higher C contents in stem compared to roots and leaves are largely attributable to the higher proportion of C-enriched structural compounds such as lignin and cellulose in stem. Among roots, stem and leaves, higher C contents in stem have also been observed in other tree species (Luo et al. 2006). Furthermore, the lowest N, P and K contents in stem among the three tissues are mainly due to the dilution effects of higher proportion of C-enriched structural compounds in stem. The highest N and P contents in leaves of black locust are probably attributable to the photosynthetic machinery (N-rich compounds) and nuclear acids (P-rich compounds) production in this most metabolically active tissue. These data are consistent with previous findings which indicate that the more metabolically active tissues or faster growing species contain higher proportions of N and P relative to C because proteins (N-rich compounds) and nuclear acids are essential for C gain and growth (Matzek and Vitousek 2009; Reef et al. 2010). Increases in N, P and K contents in leaves of 10-, 15-, 20-year-old plantations compared to that of 5-year-old plantation imply that these nutrient elements reach stable levels in foliage of older plants. By contrast, the N, P and K contents in woody plants are often observed to reduce with the ontogenetic development of individual tree (Agren 2008; Yang and Luo 2011). These results suggest that N-fixing plants may delay the ontogenetic decline of nutritional partitioning to leaves. The C and nutrient pools in black locust plantations reflect these element contents in tissues and their accumulation in biomass. The highest C pools in stems of different aged plantations result from the highest C contents and biomass accumulation of stems. The highest N pools in roots of different aged plantations are mainly due to the relative higher N contents and biomass in roots. For P and K pools, no consistent tissue patterns appear. Although significant amounts of C, N, P and K are accumulated in biomass including relatively short-lived leaves and fine roots (a few weeks to several months), some of these elements, especially organic C, may be released to the soil via decomposition and exudation, affecting soil nutrient pools (Khan et al. 2010; Qiu et al. 2010; Wang et al. 2011; Lambers et al. 2012).
Large variation in C, N, P and K stoichiometric relations is found between tissue types in woody plants (Niklas and Cobb 2006; Agren 2008). In this study, the results of foliar C:N molar ratios (20.8–25.5) are consistent with a previous study in which mean C:N molar ratio of 24.7 is observed in leaves of a few species on the Loess Plateau (Zheng and Shangguan 2007). However, foliar C:P (1,013–1,380) and N:P (48.8–54.5) ratios in the current study are much higher than the C:P (806) and N:P (34.1) ratios, respectively, in leaves of several species on the Loess Plateau (Zheng and Shangguan 2007). This is largely due to the significantly lower P contents (0.08–0.12 %) in leaves of black locust plantations compared to those (0.06–0.35 %) in foliage used by Zheng and Shangguan (2007). These results indicate that growth and productivity of black locust plantations in this study area are mainly constrained by P. As an immobile element in soil, P is often found to constrain plant growth and ecosystem productivity (Marleau et al. 2011; Marklein and Houlton 2012; Reed et al. 2012). The presence of N-fixing species in plant communities may alleviate P limitation of plant growth and ecosystem productivity because N inputs can be allocated to phosphatase enhancing P uptake of plants (Novotny et al. 2007; Cech et al. 2008; Venterink 2011). As a result, legume species can become abundant on P- and N-poor soils (Venterink 2011). The higher C:N, C:P and C:K ratios, lower N:P and N:K ratios in stems of black locust plantations are mainly due to the lower N contents caused by the dilution effects of higher proportion of C-enriched compounds such as lignin in stems. Similar results are also found in other plant species (Niklas and Cobb 2006; Agren 2008). Decreases in C:N and C:P ratios with increases in ages of black locust plantations suggest that the carbon gain per unit N and/or P declines in older plantations, which is associated with the ontogenetic decline in aging plants (Agren 2008). As the plant age increases, the N- and P-poor tissues increase more rapidly than the N- and P-rich tissues (Agren 2008). However, it is reported that C:N ratios of the plants increased with the ages of secondary forests (Yang and Luo 2011). Overall, these data indicate that changes in C, N, P and K concentrations in different tissues of black locust plantations can lead to alterations in nutrient stoichiometric relations and that the tissue- and age-specific patterns of nutrient stoichiometry are associated with chemical composition of the tissues and age-related ontogenetic development of black locust plantations.
No strong correlations exist between major nutrient concentrations in top soils and in leaves of black locust
In natural ecosystems, nutrient elements are cycled between soils and plants (Agren 2008). Exchange of nutrient elements such as N, P and K between the soil and plants underlies reciprocal control of element composition in both sides (Ladanai et al. 2010). Elements such as C, N, P and K are stoichiometrically coupled in both soils (Cleveland and Liptzin 2007; Tian et al. 2010) and plants (Agren 2008). However, whether correlations of these elements exist between the soils and plants currently remains unknown. As an attempt to address this question, we have performed the correlation analysis in this study. Although there is a correlation between P contents in the 10–20 cm soil layer and in leaves of black locust plants, no correlations exist between other elements (C, N and K) in soils and in leaves of black locust. Coupling analysis between elements in soils and in foliage has been performed in Scots pine (Pinus sylvestris) and Norway spruce (Picea abies) forests in Sweden, and no correlations have been found (Ladanai et al. 2010). These results suggest that coupling relations of major elements such as C, N, P and K between soils and forest plants did not always exist, although forest plantations can significantly alter soil physical, chemical and biological properties (Khan et al. 2010; Wang et al. 2011).
An important factor affecting the coupling relations of C, N, P and K between soils and forest plants is the chemical properties of forest litter because the chemical properties of tree tissues, especially leaves and fine roots, can substantially impact soil element stoichiometric relations (Khan et al. 2010; Qiu et al. 2010). The difference in chemical properties of root, stem and leaf tissues of different-aged black locust plantations may partially explain the contribution of these tissues to changes in nutrient stoichiometric relations in soils. For instance, higher concentrations of soluble sugar and lower lignin concentrations of leaves than those of roots indicate that the decomposition of leaf litter may be faster than that of roots since soil microorganisms favor for plant litter with higher sugar concentrations and lower lignin contents (Cotrufo et al. 2005; Luo et al. 2008). In the same line, lower concentrations of soluble sugar and elevated lignin concentrations in roots and leaves of older black locust plantations than those of young plantations may delay the release of nutrient elements in these tissues to soils during the decomposition processes. These results indicate that the chemical properties of plant tissues may affect the decomposition processes of litter, resulting in decoupling of nutrients between the soil and the plant.
In summary, black locust plantations, particularly 15- and 20-year-old plantations, have increased SOC and N contents and stocks in the top soil layer, leading to changes in soil C, N, P and K stoichiometric relations. The C:P, C:K, N:P, N:K and P:K molar ratios are higher in the 0–10 cm soil layer than those in the 10–20 cm soil layer. All analyzed element ratios in soils are elevated under older plantations. Biomass of plantations is accumulated in older plantations. The C, N, P and K contents in black locust have displayed tissue- and age-specific patterns. Higher C contents in stem and N contents in leaves have been found in black locust. The largest C pools in stem and N pools in roots were detected in 20-year-old black locust. The highest C:N, C:P and C:K and the lowest N:P and N:K in stem have been observed among the three tissues. The C:N and C:P ratios decrease in older black locust. No strong correlations exist between elements (C, N and K) in soils and leaves of plantations. These results suggest that black locust plantations can increase soil nutrient contents resulting in changes in element stoichiometric relations, nutrient element contents and their stoichiometric couplings, which vary in different tissues and aged plants, and no strong coupling relations exist between major nutrient element contents in soils and in leaves of plants. These data also indicate that N-fixing woody plants such as R. pseudoacacia are good candidates for afforestation/reforestation on degraded land to prevent soil erosion and increase SOC and N storage contributing to mitigation of global change.
Author contribution
H. Li wrote the manuscript, H. Li, J. Li, Y. He and S. Li carried out the field work and laboratory analysis, Z. Liang, C. Peng and A. Polle helped in drafting the manuscript, Z.B. Luo conceived the experiment and helped in writing the manuscript and interpretation of the results.
References
Agren GI (2008) Stoichiometry and nutrition of plant growth in natural communities. Annu Rev Ecol Evol Syst 39:153–170
An S, Huang Y (2006) Rapid changes of soil properties following Caragana korshinskii plantations in the hillygully Loess Plateau. Frontiers For China 1:394–399
Bradford JB, Birdsey RA, Joyce LA, Ryan MG (2008) Tree age, disturbance history, and carbon stocks and fluxes in subalpine Rocky mountain forests. Glob Change Biol 14:2882–2897
Cao S, Chen L, Xu C, Liu Z (2007) Impact of three soil types on afforestation in China’s Loess Plateau: growth and survival of six tree species and their effects on soil properties. Landsc Urban Plan 83:208–217
Cao SX, Chen L, Yu XX (2009) Impact of China’s grain for Green Project on the landscape of vulnerable arid and semi-arid agricultural regions: a case study in northern Shaanxi Province. J Appl Ecol 46:536–543
Cech PG, Kuster T, Edwards PJ, Venterink HO (2008) Effects of herbivory, fire and N2-fixation on nutrient limitation in a humid African Savanna. Ecosystems 11:991–1004
Chen C, Xu Z (2005) Soil carbon and nitrogen pools and microbial properties in a 6-year-old slash pine plantation of subtropical Australia: impacts of harvest residue management. For Ecol Manage 206:237–247
Chen FS, Feng X, Liang C (2012) Endogenous versus exogenous nutrient affects C, N, and P dynamics in decomposing litters in mid-subtropical forests of China. Ecol Res 27:923–932
Cleveland CC, Liptzin D (2007) C:N:P stoichiometry in soil: is there a ‘‘Redfield ratio’’ for the microbial biomass? Biogeochemistry 85:235–252
Cotrufo MF, De Angelis P, Polle A (2005) Leaf litter production and decomposition in a poplar short-rotation coppice exposed to free air CO2 enrichment (POPFACE). Glob Change Biol 11:971–982
Dong XW, Zhang XK, Bao XL, Wang JK (2009) Spatial distribution of soil nutrients after the establishment of sand-fixing shrubs on sand dune. Plant Soil Environ 55:288–294
Duponnois R, Founoune H, Masse D, Pontanier R (2005) Inoculation of Acacia holosericea with ectomycorrhizal fungi in a semiarid site in Senegal: growth response and influences on the mycorrhizal soil infectivity after 2 years plantation. For Ecol Manag 207:351–362
Elser JJ, Fagan WF, Kerkhoff AJ, Swenson NG, Enquist BJ (2010) Biological stoichiometry of plant production: metabolism, scaling and ecological response to global change. New Phytol 186:593–608
Ferrari AE, Wall LG (2007) Nodulation and growth of black locust (Robinia pseudoacacia) on a desurfaced soil inoculated with a local Rhizobium isolate. Biol Fertil Soils 43:471–477
Forrester DI, Pares A, O’Hara C, Khanna PK, Bauhus J (2013) Soil organic carbon is increased in mixed-species plantations of Eucalyptus and nitrogen-fixing Acacia. Ecosystems 16:123–132
Gusewell S (2004) N:P ratios in terrestrial plants: variation and functional significance. New Phytol 164:243–266
Hazlett P, Gordon A, Voroney R, Sibley P (2007) Impact of harvesting and logging slash on nitrogen and carbon dynamics in soils from upland spruce forests in northeastern Ontario. Soil Biol Biochem 39:43–57
Jimenez JJ, Lal R, Leblanc HA, Russo RO (2007) Soil organic carbon pool under native tree plantations in the Caribbean lowlands of Costa Rica. For Ecol Manage 241:134–144
Khan B, Ablimit A, Mahmood R, Qasim M (2010) Robinia pseudoacacia leaves improve soil physical and chemical properties. J Arid Land 2:266–271
Ladanai S, Agren GI, Olsson BA (2010) Relationships between tree and soil properties in Picea abies and Pinus sylvestris forests in Sweden. Ecosystems 13:302–316
Laik R, Kumar K, Das DK, Chaturvedi OP (2009) Labile soil organic matter pools in a calciorthent after 18 years of afforestation by different plantations. Appl Soil Ecol 42:71–78
Lal R, Follett F, Stewart BA, Kimble JM (2007) Soil carbon sequestration to mitigate climate change and advance food security. Soil Sci 172:943–956
Lambers H, Bishop JG, Hopper SD, Laliberte E, Zuniga-Feest A (2012) Phosphorus-mobilization ecosystem engineering: the roles of cluster roots and carboxylate exudation in young P-limited ecosystems. Ann Bot 110:329–348
Lee YC, Nam JM, Kim JG (2011) The influence of black locust (Robinia pseudoacacia) flower and leaf fall on soil phosphate. Plant Soil 341:269–277
Luo ZB, Polle A (2009) Wood composition and energy content in a poplar short rotation plantation on fertilized agricultural land in a future CO2 atmosphere. Glob Change Biol 15:38–47
Luo ZB, Calfapietra C, Liberloo M, Scarascia-Mugnozza G, Polle A (2006) Carbon partitioning to mobile and structural fractions in poplar wood under elevated CO2 (EUROFACE) and N fertilization. Glob Change Biol 12:272–283
Luo ZB, Calfapietra C, Scarascia-Mugnozza G, Liberloo M, Polle A (2008) Carbon-based secondary metabolites and internal nitrogen pools in Populus nigra under free air CO2 enrichment (FACE) and nitrogen fertilisation. Plant Soil 304:45–57
Marklein AR, Houlton BZ (2012) Nitrogen inputs accelerate phosphorus cycling rates across a wide variety of terrestrial ecosystems. New Phytol 193:696–704
Marleau JN, Jin Y, Bishop JG, Fagan WF, Lewis MA (2011) A stoichiometric model of early plant primary succession. Am Nat 177:233–245
Matzek V, Vitousek PM (2009) N:P stoichiometry and protein: RNA ratios in vascular plants: an evaluation of the growth-rate hypothesis. Ecol Lett 12:765–771
Menyailo O, Hungate B, Zech W (2002) The effect of single tree species on soil microbial activities related to C and N cycling in the Siberian artificial afforestation experiment: tree species and soil microbial activities. Plant Soil 242:183–196
Nelson DW, Sommers LE (1975) A rapid and accurate method for estimating organic carbon in soil. Proc Ind Acad Sci 84:456–462
Niklas KJ, Cobb ED (2006) Biomass partitioning and leaf N, P-stoichiometry: comparisons between tree and herbaceous current-year shoots. Plant Cell Environ 29:2030–2042
Novotny AM, Schade JD, Hobbie SE, Kay AD, Kyle M, Reich PB, Elser JJ (2007) Stoichiometric response of nitrogen-fixing and non-fixing dicots to manipulations of CO2, nitrogen, and diversity. Oecologia 151:687–696
Olesniewicz KS, Thomas RB (1999) Effects of mycorrhizal colonization on biomass production and nitrogen fixation of black locust (Robinia pseudoacacia) seedlings grown under elevated atmospheric carbon dioxide. New Phytol 142:133–140
Parkinson JA, Allen SE (1975) A wet oxidation procedure suitable for determination of nitrogen and mineral nutrients in biological material. Commun Soil Sci Plant Anal 6:1–11
Pregitzer KS, Euskirchen ES (2004) Carbon cycling and storage in world forests: biome patterns related to forest age. Glob Change Biol 10:2052–2077
Qiu L, Zhang X, Cheng J, Yin X (2010) Effects of black locust (Robinia pseudoacacia) on soil properties in the loessial gully region of the Loess Plateau, China. Plant Soil 332:207–217
Reed SC, Townsend AR, Davidson EA, Cleveland CC (2012) Stoichiometric patterns in foliar nutrient resorption across multiple scales. New Phytol 196:173–180
Reef R, Ball MC, Feller IC, Lovelock CE (2010) Relationships among RNA: DNA ratio, growth and elemental stoichiometry in mangrove trees. Funct Ecol 24:1064–1072
Roy PK, Samal NR, Roy MB, Mazumdar A (2010) Soil carbon and nutrient accumulation under forest plantations in Jharkhand State of India. Clean-Soil Air Water 38:706–712
Sardans J, Rivas-Ubach A, Penuelas J (2011) Factors affecting nutrient concentration and stoichiometry of forest trees in Catalonia (NE Spain). Forest Ecol Manag 262:2024–2034
Tian HQ, Chen GS, Zhang C, Melillo JM, Hall CAS (2010) Pattern and variation of C:N:P ratios in China’s soils: a synthesis of observational data. Biogeochemistry 98:139–151
Venterink HO (2011) Legumes have a higher root phosphatase activity than other forbs, particularly under low inorganic P and N supply. Plant Soil 347:137–146
Wang QL, Bai YH, Gao HW, He J, Chen H, Chesney RC, Kuhn NJ, Li HW (2008) Soil chemical properties and microbial biomass after 16 years of no-tillage fanning on the Loess Plateau, China. Geoderma 144:502–508
Wang WJ, Qiu L, Zu YG, Su DX, An J, Wang HY, Zheng GY, Sun W, Chen XQ (2011) Changes in soil organic carbon, nitrogen, pH and bulk density with the development of larch (Larix gmelinii) plantations in China. Glob Change Biol 17:2657–2676
Xia J, Wan S (2008) Global response patterns of terrestrial plant species to nitrogen addition. New Phytol 179:428–439
Yang Y, Luo Y (2011) Carbon: Nitrogen stoichiometry in forest ecosystems during stand development. Global Ecol Biogeogr 20:354–361
Yang YH, Luo YQ, Lu M, Schadel C, Han WX (2011) Terrestrial C:N stoichiometry in response to elevated CO2 and N addition: a synthesis of two meta-analyses. Plant Soil 343:393–400
Zhang LX, Li SX, Liang ZS (2009) Differential plant growth and osmotic effects of two maize (Zea mays L.) cultivars to exogenous glycinebetaine application under drought stress. Plant Growth Regul 58:297–305
Zheng SX, Shangguan Z (2007) Spatial patterns of leaf nutrient traits of the plants in the Loess Plateau of China. Trees-Struct Funct 21:357–370
Zheng Y, Zhao Z, Zhou JJ, Zhou H, Liang ZS, Luo ZB (2011) The importance of slope aspect and stand age on the photosynthetic carbon fixation capacity of forest: a case study with black locust (Robinia pseudoacacia) plantations on the Loess Plateau. Acta Physiol Plant 33:419–429
Acknowledgments
This work is supported by the State Key Basic Research Development Program (Grant No. 2012CB416902), the National Natural Science Foundation of China (Grant No. 31070539, 31100481, 31270647), the Program for New Century Excellent Talents in University from the Ministry of Education of China (Grant No. NCET-08-0468), the Fok Ying Tung Education Foundation (Grant No. 121026), the Specialized Research Fund for the Doctoral Program of Higher Education of China (Grant No. 20090204110027), the Fundamental Research Funds for the Central Universities of China (Grant No. YQ2013005) and the Knowledge Innovation Program of the Chinese Academy of Sciences (Grant No. KZCX2-YW-443).
Conflict of interest
The authors declare that they have no conflict of interest.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by L. Bavaresco.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article
Cite this article
Li, H., Li, J., He, Y. et al. Changes in carbon, nutrients and stoichiometric relations under different soil depths, plant tissues and ages in black locust plantations. Acta Physiol Plant 35, 2951–2964 (2013). https://doi.org/10.1007/s11738-013-1326-6
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11738-013-1326-6